

Зоологический журнал, 2021, T. 100, № 8, стр. 914-926
Биология размножения хрустана (Eudromias morinellus) на плато Путорана, Средняя Сибирь
А. А. Романов a, *, В. В. Тарасов b, **
a Географический факультет Московского государственного университета имени М.В. Ломоносова
119991 Москва, Россия
b Институт экологии растений и животных УрО РАН
620144 Екатеринбург, Россия
* E-mail: putorana05@mail.ru
** E-mail: grouse@bk.ru
Поступила в редакцию 23.04.2020
После доработки 03.09.2020
Принята к публикации 08.09.2020
Аннотация
Приведены данные по биологии и экологии хрустана (Eudromias morinellus), полученные в 1988–2018 гг. в крупнейшем горном массиве Российской Арктики. Установлено, что в период размножения вид осваивает широкий диапазон высот, почти повсеместно населяет плоские вершины в пределах гольцового пояса, локально – некоторые участки подгольцового пояса и крайне редко встречается в горно-таежном поясе. Его ареал имеет явно выраженный трехмерный характер. Оптимальные местообитания в гольцовом поясе приурочены к сухим щебнистым участкам горной тундры, которые имеют бугристый микрорельеф и покрыты скудной травянистой растительностью. Обилие вида в гольцовом поясе в гнездовой период варьирует от 0.4 до 10.0 особей/км2, в среднем 1.7 особей/км2. Обилие хрустана максимально в средней части гольцового пояса (11.5 особей/км2) и сокращается в сторону вершин (1.3 особей/км2) и подножий склонов (3.5 особей/км2). В средней части гольцового пояса, составляющей 42% жизненного пространства в вертикальной плоскости, в гнездовой период концентрируется 70% всех особей вида. Самки до середины 3-й декады июня активно токуют в полете, а также на земле – поодиночке и группами до 15 особей. Откладка яиц начинается 7–22 июня, часто – при еще почти сплошном снежном покрове. Гнезда располагаются на высотах 850–1036 м над ур. м., в среднем 935 ± 28 (n = 8). Локальная гнездовая плотность, определяемая по среднему минимальному расстоянию между соседними гнездами, может составлять 1.67 гнезд/км2. Наиболее ранняя расчетная дата появления птенцов 3 июля, наиболее поздняя – 18 июля, средняя – 9 июля ± 1 день (n = 13). В одном гнезде птенцы вылупляются в течение одних суток.
Хрустан (Eudromias morinellus Linnaeus 1758) имеет арктоальпийское распространение по всему северу Палеарктики (Степанян, 2003; Лаппо и др., 2012; Рябицев, 2014). Особенности распространения и экологии этого кулика на значительной части ареала позволяют отнести его к типично горным видам. До 1980-х гг. его справедливо признавали слабоизученным видом (Рогачева, 1988). Многие аспекты его биологии на значительной части ареала до сих пор изучены неудовлетворительно. Недостаточны даже сведения о распространении хрустана в Российской Федерации, в т.ч. на плато Путорана, где находится один из предполагаемых оптимумов гнездового ареала (Лаппо и др., 2012). При этом констатируются снижение численности и сокращение ареала вида (Томкович, 2007) – по-видимому, во многом в результате гибели птиц на зимовках в Северной Африке, где используются пестициды против саранчи (Hable, Präsent, 1990; Saari, 1995).
Основная цель – обобщение и анализ данных по экологии и биологии хрустана на плато Путорана для использования при мониторинге и разработке мер сохранения вида. В соответствии с этим решали три основные задачи: 1 – выявление закономерностей пространственного размещения и показателей обилия вида в гнездовой период; 2 – выявление основных элементов поведения, параметров устройства гнезд, размеров кладок и яиц; 3 – установление календарных дат основных фенологических явлений в период гнездования.
МАТЕРИАЛЫ И МЕТОДЫ
Материалы по биологии и экологии хрустана собраны в 1988–2018 гг. на плато Путорана – крайней северо-западной оконечности Средне-Сибирского плоскогорья и одновременно срединной части гнездового ареала вида. Район исследований расположен между 65°00′–71°00′ с.ш. и 90°00′–100°00′ в.д. Площадь арены исследований составила около 250 тыс. км2. Сведения, приводимые в настоящей работе, получены во время стационарных наблюдений и пеших маршрутов, которые были проведены с мая по август в вышеуказанные сезоны и которые охватили северные, южные, центральные, западные и восточные районы плато Путорана (см. табл. 1). Один и тот же географический объект плато Путорана на разных картах может быть назван по-разному. Например, оз. Нералах в ряде картографических источников именуется как Нералак, оз. Негу-Икэн – как Негу-Икон. В нашей работе названия обследованных рек и озер соответствуют названиям на среднемасштабной топографической карте масштаба 1 : 200 000.
Таблица 1.
Протяженность маршрутов (км), обилие хрустана (особей/км2) и доля его участия (%) в общем населении птиц в гнездовой период в различных пунктах плато Путорана
| № | Пункты регистрации вида | Гольцовый пояс | Подгольцовый пояс | ||
|---|---|---|---|---|---|
| Обилие/ протяженность маршрутов |
Доля | Обилие/ протяженность маршрутов |
Доля | ||
| Север плато | |||||
| 1 | оз. Боковое (70°10′ с.ш., 94°15′ в.д.) | 3.7/25 | 3.2 | – | – |
| 2 | р. Аян (69°00′–69°20′ с.ш., 93°30′– 94°30′ в.д.) | 10.0/86 | 22.4 | – | – |
| 3 | озера Богатырь, Нералах (69°35′–69°43′ с.ш., 92°10′–92°35′ в.д.) | 0.5/227 | 0.7 | – | – |
| 4 | оз. Негу-Икэн (69°35′–69°47′ с.ш., 93°05′–93°15′ в.д.) | 0.5/141 | 0.5 | – | – |
| 5 | оз. Богатырь-Хуолу (69°42′–69°49′ с.ш., 92°30′–92°45′ в.д.) | 6.0/289 | 2.5 | – | – |
| Центр плато | |||||
| 6 | оз. Аян (69°00′–69°20′ с.ш., 93°30′–94°30′ в.д.) | 1.0/104 | 4.3 | 0.6/68 | 1.3 |
| Запад плато | |||||
| 7 | оз. Кутарамакан (68°35′–68°50′ с.ш., 91°30′–92°30′ в.д.) | 0.6/60 | 2.0 | 8.0/11 | 9.9 |
| 8 | Гусиные озера в бассейне р. Курейки (68°22′ с.ш., 93°30′ в.д.) | 0.4/113 | 1.1 | 0.2/41 | 0.05 |
| 9 | оз. Качук (69°00′ с.ш., 94°30′ в.д.)* | – | – | – | – |
| 10 | Норильские озера (68°41′–68°49′ с.ш., 89°36′–90°26′ в.д.)** | – | – | – | – |
| Восток плато | |||||
| 11 | оз. Харпича (68°46′ с.ш., 96°57′ в.д.) | 0.7/21 | 1.9 | 1.5/17 | 2.1 |
| 12 | оз. Дюпкун Котуйский (68°00′ с.ш., 98°30′ в.д.) | 1.2/21 | 3.4 | – | – |
| 13 | оз. Нерангда (69°00′ с.ш., 98°00′ в.д.)*** | – | – | – | – |
| Юг плато | |||||
| 14 | оз. Някшингда (67°00′ с.ш., 93°30′ в.д.) | 1.0/30 | 2.2 | – | – |
Для анализа обилия вида использованы результаты маршрутных учетов, проведенных по методике Равкина (1967) на высотах до 1412 м над ур. м., протяженность маршрутов указана в табл. 1. Высоту местности и длину пройденного пути определяли по навигаторам GPS. Суммарная протяженность маршрутов составила 6921 км: 4063 км – в лесных ландшафтах горно-таежного пояса, 238 км – в горных редколесьях подгольцового пояса, 1204 км – в горных тундрах гольцового пояса. Птиц на маршрутах регистрировали визуально и по голосу. Все показатели обилия, приводимые в настоящей работе, получены на учетах в период размножения, включая этапы брачных демонстраций, насиживания кладок и вождения птенцов, возраст которых не превышал 7–10 дней. Различия в уровне встречаемости по указанным этапам цикла размножения не выявлены, поэтому все данные суммированы и усреднены.
Гнезда хрустана обнаруживали, наблюдая за беспокоящимися особями, а также случайным образом при вспугивании птиц на маршрутах. Настойчиво беспокоившихся особей со второй половины июля относили к птицам, сопровождавшим затаившихся птенцов.
Степень насиженности яиц и дату появления первого яйца в кладках, сроки формирования которых неизвестны, определяли методом флотации (для куликов – по: Liebezeit et al., 2007), который является общепринятым в исследовании биологии размножения птиц (Westerkov, 1950; Rahn, 1974).
ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА РАЙОНА ИССЛЕДОВАНИЙ
Плато Путорана – самый крупный горный массив Российской Арктики. В соответствии с вертикальной дифференциацией растительности различают высотно-ландшафтные пояса: горно-северотаежный (лесной), подгольцовый (горно-лесотундровый) и гольцовый (горно-тундровый) (Норин, 1986). Хрустан в своем распространении на плато Путорана экологически связан почти исключительно с гольцовым поясом. Физико-географическое описание этого пояса содержится в ряде монографий (Пармузин, 1964, 1976; Гвоздецкий, Голубчиков, 1987; Голубчиков, 1996; Куваев, 2006), поэтому приведем лишь краткую характеристику основных местообитаний вида.
Гольцовый пояс расположен на высотах более 500–800 м над ур. м., где при полном отсутствии деревьев и кустарников перемежаются участки, покрытые горной тундрой и практически лишенные растительности (Куваев, 2006). Рельеф формируют возвышающиеся над окружающим пространством крутосклонные массивы плато, сильно разрушенные горные массивы со сглаженными куполообразными вершинами и пологими террасированными склонами, а также столовые горные плато с плоскими вершинами, приподнятыми в среднем на 900–1200 м над ур. м. Современный облик окружающих вершины низкогорных ландшафтов сформировался под воздействием поздне-плейстоценового оледенения.
Горные вершины плато отличаются особой экстремальностью климатических условий (Голубчиков, 1996). Снег здесь регулярно выпадает и летом, и даже в июле может на несколько дней образовать устойчивый покров глубиной до 15–20 см. В годы с холодным летом, когда температура воздуха не превышает +5°С, мощный лед на озерах сохраняется в почти неизменном виде все лето (Пармузин, 1976), образуются лишь узкие (шириной 2–15 м) полыньи, вытянутые вдоль берегов. Повсеместно распространены мохово-лишайниково-осоковые, мохово-лишайниково-кустарничковые, мохово-осоково-разнотравные тундры. Проективное покрытие растительностью на разных участках гольцового пояса составляет 25–100%.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Распространение вида в пределах региона
Наблюдения, проведенные нами и другими исследователями, указывают на то, что хрустан распространен на плато Путорана почти повсеместно и зарегистрирован в большинстве обследованных пунктов: в 11 из 15 – по нашим данным (Романов, 1996, 2006а, 2013, 2015; Романов и др., 2007, 2018, 2019) и в 14 из 18 – по обобщенным данным, полученным нами и другими исследователями (Кречмар, 1966; Морозов, 1984; Зырянов, 1988). Не удалось встретить хрустана лишь в долине р. Микчангда и в котловинах озер Собачье, Кета, Агата Верхняя (Романов, 2006, 2013; Рупасов, Журавлев, 2006). Вид достоверно гнездится в северной, западной и восточной частях плато. Гнездование в центральной и южной его частях также не вызывает сомнений, но фактами не подтверждено.
Высотно-ландшафтное размещение
Хрустан – типичный обитатель горно-тундровых ландшафтов вершин плато, неотъемлемый элемент авифауны гольцового пояса, входит в число видов, формирующих ее ядро. В гнездовой период подавляющее большинство встреч зарегистрировано в гольцовом поясе на высотах 750–1245 м над ур. м., преимущественно 890–1070 м над ур. м. Вид предпочитает участки с небольшим уклоном, которые имеют бугристый микрорельеф и которые покрыты скудной травянистой растительностью. Его оптимальные местообитания – сухие каменистые мохово-лишайниковые тундры с различной долей участия дриады (Dryas sp.), кассиопеи (Cassiope sp.) и осоки (Carex sp.), с обилием каменистых и щебнистых россыпей, а также мерзлотных медальонов. Охотно держится на вершинах крупных щебнистых бугров, поверхность которых, если не считать лишайниковых куртин, лишена всякой растительности. Избегает крутых горных склонов, скал и осыпей.
Значительно в меньшей степени хрустан населяет подгольцовый пояс, куда наряду с некоторыми другими видами – тундровой куропаткой (Lagopus muta), золотистой ржанкой (Pluvialis apricaria), краснозобым коньком (Anthus cervinus), гольцовым коньком (A. rubescens), обыкновенной каменкой (Oenanthe oenanthe), – экологически тесно связанными с гольцовыми ландшафтами, может проникать при наличии достаточно обширных участков низкорослой горно-тундровой растительности. В подобных местообитаниях подгольцового пояса хрустан встречается в районе озер Аян, Кутарамакан, Дюпкун Курейский, Харпича. По разделяющим заросли ерника (Betula fruticosa), ольховника (Duschekia fruticosa), ивняка (Salix glauca, S. lanata) участкам каменистых горных тундр с пятнами мерзлотных форм рельефа хрустан может опускаться ниже верхней границы подгольцового пояса, которая на севере плато проходит в среднем по высоте 500, на юге – 800, на западе и в центре – 750 м над ур. м. Хрустан и тундровая куропатка в пределах подгольцового пояса строго придерживаются горно-тундровых участков. Остальные из перечисленных видов не проявляют явной биотопической избирательности и встречаются также среди редин из лиственниц (Larix gmelinii), куртин ольховника, на опушках горных лиственничных редколесий.
В пределах горно-таежного пояса хрустан распространен в буквальном смысле точечно (Кречмар, 1966). Основным лимитирующим фактором, препятствующим более широкому распространению вида в этом поясе, следует считать не абсолютную высоту местности, а ничтожно малую площадь пригодных для гнездования местообитаний и отсутствие экологических “русел проникновения”. Но, несмотря на это, там, где на высотах 100–120 м над ур. м. формируются подходящие биотопы, часть птиц охотно селятся локальными изолированными очагами и успешно гнездятся (Кречмар, 1966). Известны случаи гнездования в тундроподобных ландшафтах по берегам Норильских озер в пределах горно-таежного пояса (Кречмар, 1966). Формирование подходящих гнездовых местообитаний в пределах экологически чуждых виду северо-таежных ландшафтов обусловлено температурной и, соответственно, ландшафтной инверсией. Благодаря охлаждающему влиянию крупных озер, имеющих “полярный” гидрологический режим, вода весь вегетационный период имеет значительно более низкую температуру, чем воздух, и в совокупности с постоянными ветрами оказывает сильнейшее охлаждающее воздействие на растительность прибрежных пространств (приозерий) на расстоянии до 1–1.5 км (Водопьянова, 1976). В приозерьях получают широкое распространение открытые тундроподобные каменистые ландшафты. Таких мест на плато Путорана немного, но единичные пары хрустанов, вероятно, гнездятся там регулярно.
В целом, на плато Путорана хрустан в гнездовой период осваивает широкий диапазон высот, при этом почти повсеместно населяет плоские вершины гольцового пояса, локально – некоторые участки подгольцового пояса, и крайне редко встречается у подножий склонов в пределах горно-таежного пояса. Ареал этого вида, обитающего во всех трех высотных поясах, имеет явно выраженный трехмерный характер. Полученные нами данные свидетельствуют о том, что высотное распределение вида в пределах высотно-поясного спектра обусловлено не столько абсолютной высотой местности, сколько экологическими параметрами предпочитаемых местообитаний и границами распространения наиболее оптимальных из них.
Дифференциация обилия в пределах гольцового пояса
Выявлены существенные различия показателей обилия в разных пунктах, обследованных в разные годы (см. табл. 1). Эти различия могут быть обусловлены как межгодовой динамикой численности (Лаппо и др., 2012), так и экологическими особенностями местообитаний.
Из 11 пунктов плато Путорана, где в 1988–2018 гг. мы встретили хрустана, многочислен он был лишь в одном (р. Аян, где в населении птиц он оказался в числе доминантов с долей участия 22.4%), обычен в 5 и редок в 5 пунктах. В 8 пунктах он входил в число субдоминантов с долей участия 1.1–4.3%. По данным учетов, среднее обилие вида по всему гольцовому поясу составило 1.7 особей/км2, а по подгольцовому – 0.8 особей/км2. В гольцовом поясе обилие хрустана в гнездовой период варьировало от 0.4 до 10.0 особей/км2, достигая максимальных значений в северных частях региона. Локальные участки повышенной плотности населения вида обнаружены в 1989 г. на вершинах плато в среднем течении р. Аян (10.0 особей/км2) и в 2018 г. в котловине оз. Богатырь-Хуолу (6.0 особей/км2). В подгольцовом поясе показатели обилия изменялись в пределах 0.2–8.0 особей/км2, наибольшая плотность (8.0 особей/км2) обнаружена в 1990 г. на вершинах плато у юго-западной оконечности оз. Кутарамакан.
В котловине оз. Богатырь-Хуолу на северо-западе плато выявлена вертикальная дифференциация показателей обилия хрустана в пределах нижней (730–950 м над ур. м.), средней (950–1100 м над ур. м.) и верхней (1100–1412 м над ур. м.) частей гольцового пояса, предопределенная различиями флористического состава, структуры и высоты растительного покрова, показателей проективного покрытия, широты распространения открытых каменистых поверхностей и увлажнения (Романов и др., 2019).
Нижняя часть пояса, где обилие хрустана составило 3.5 особей/км2, представлена влажной мелкокочкарной мохово-ивняково-осоковой горной тундрой с дриадой. В поймах рек и на пологих склонах озерных котловин широко распространены дерновинные разнотравно-луговые тундры с обилием дриады, копеечника арктического (Hedysarum arcticum), остролодочника (Oxytropis sp.), горца (Polygonus sp.), новосиверсии ледяной (Novosieversia glacialis) с проективным покрытием до 100%. Обилие трав приближает дерновинную тундру к луговым ценозам (Павлов и др., 1988).
В средней части обилие хрустана 11.5 особей/км2. Проективное покрытие здесь не превышает 60%, повсеместно господствуют мохово-лишайниковые, мохово-осоковые, разнотравно-осоковые тундры с ивой арктической (Salix arctica), карликовой березкой (Betula nana), голубикой (Vaccinium uliginosum). Сухие пологие склоны котловин озер изобилуют мерзлотными медальонами, покрыты лишайниками, кассиопеей четырехгранной (Cassiope tetragona) и дриадой. Широко распространены песчано-щебнистые холмы, мозаично поросшие куртинами лишайников, дриады, злаков и незабудочника (Eritrichinum sp.). Холмы чередуются с обширными плоскими переувлажненными долинами речек и ручьев, задернованная мелкобугристая поверхность которых занята густым растительным покровом изо мхов, лишайников, дриады, кассиопеи, злаков, осок, карликовых ив. Повсюду многочисленны снежники, каменистые россыпи (курумы) и скопления валунов.
Верхняя часть гольцового пояса, где обилие хрустана составило 1.3 особей/км2, отличается крайне низкими показателями проективного покрытия (лишь местами до 40%). Террасированные и крутые склоны заняты почти безжизненными холодными гольцовыми пустынями (Куваев, 2006), где мозаично чередуются многочисленные горные обрывы, скалы-останцы, обширные снежники, пятна мерзлопученного грунта, каменистые россыпи с фрагментарным вкраплением маленьких куртинок изо мхов, лишайников, осок, дриады, новосиверсии.
Количественные учеты птиц в котловине оз. Богатырь-Хуолу, целенаправленно проведенные на разных высотах, показали, что на фоне снижения общей плотности населения птиц от подножий к вершинам обилие хрустана максимально в средней части гольцового пояса и постепенно сокращается в сторону как вершин, так и подножий склонов (Романов и др., 2019). В средней части гольцового пояса, составляющей 42% жизненного пространства в вертикальной плоскости, в гнездовой период концентрируется 70% всех особей вида. С увеличением высоты, в условиях снижения теплообеспеченности, сокращения продолжительности бесснежного периода и фрагментации растительного покрова обилие снижается значительно более интенсивно.
Аналогичные закономерности вертикальной дифференциации обилия хрустана установлены при анализе соответствующих показателей в котловинах всех обследованных в 2010, 2013 и 2018 гг. гольцовых озер: Богатырь (970 м над ур. м.), Нералах (920 м над ур. м.), Богатырь-Хуолу (854 м над ур. м.) и Негу-Икэн (760 м над ур. м.). Обилие хрустана здесь оказалось максимальным (6.0 особей/км2) в котловине оз. Богатырь-Хуолу (на средних относительно других озер высотах), и его сокращение прослеживается как в направлении вышерасположенного оз. Богатырь (0.5 особей/км2), так и в направлении нижерасположенного оз. Негу-Икэн (0.5 особей/км2).
Картирование в 2018 г. в котловине оз. Богатырь-Хуолу найденных гнезд (n = 5) и выводков с недавно вылупившимися птенцами (n = 3) показало более низкое значение обилия, чем по результатам маршрутных учетов. Точки этих находок оказались ограничены участком горной тундры площадью 4 км2 (см. рис. 1). При условии тотального обнаружения гнезд (что на практике недостижимо) и равного соотношения полов это соответствует обилию в 4.0 особей/км2 (по данным маршрутных учетов – 6.0 особей/км2). Различия в полученных разными методами результатах можно объяснить или нарушением баланса соотношения полов (по визуальным наблюдениям, действительно, было много холостых самок), или тем, что мы нашли слишком мало гнезд, не более трети от их реального числа. Второе предположение вряд ли возможно, так как мы обследовали контрольный участок достаточно продолжительное время, а найденные гнезда размещались вполне равномерно. Это позволяет надеяться, что мы пропустили не более 1–2 гнезд. В таком случае плотность гнездования в этом “центре” составила 2–2.5 условных пар (гнезд)/км2 (Романов и др., 2018). Расчет гнездовой плотности по среднему минимальному расстоянию между соседними гнездами дает еще более низкое значение. Расстояние между ближайшими гнездами и местами обнаружения выводков (n = 14) составило в среднем 774 ± 78 (SE) м (от 369 до 1167). Если бы все гнезда располагались равномерно по территории с таким расстоянием одно от другого, плотность составила бы 1.67 гнезд/км2.
Рис. 1.
Схема размещения гнезд (1), выводков (2) и стай холостых самок (3) в 2018 г. в котловине оз. Богатырь-Хуолу; 4 – контрольный участок (4 км2), 5 – участок наибольшей токовой активности самок.
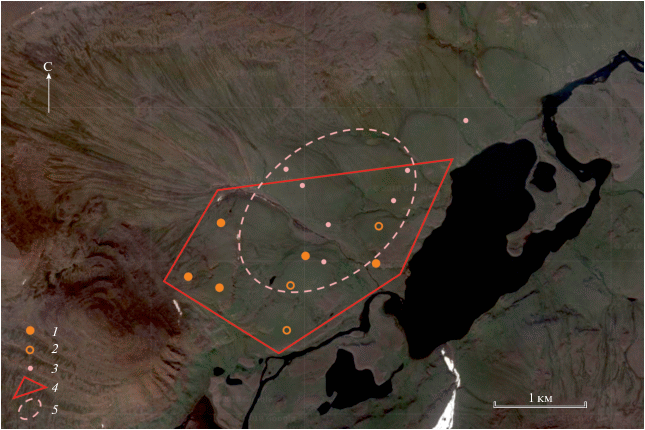
При этом здесь же, в котловине оз. Богатырь-Хуолу, выявлена выраженная неравномерность распределения птиц по территории. Так, у подножия высоких столовых гор на границе нижней и средней частей гольцового пояса, на пологих склонах с высотами от 870 до 970 м над ур. м., мы обнаружили участок повышенной концентрации птиц размером примерно 1.5 × 2 км. На этом участке заметно чаще, чем где-либо в других местах, мы встречали токующих самок – как в воздухе, так и на земле. Они собирались в группы до 15 особей и перемещались по этому участку, устраивая своеобразные “блуждающие тока” подобно самцам турухтана (Philomachus pugnax) (см.: Рябицев и др., 2003; Рябицев, 2014). Вблизи этого “центра активности самок” располагались все найденные в тот сезон гнезда и выводки. Мы предполагаем, что на северо-западе плато Путорана такие “центры” могут быть удалены один от другого на несколько десятков километров.
Хрустан – один из 17 видов птиц, которые, несмотря на экстремальные условия среды верхней части гольцового пояса, держатся в гнездовой период на практически безжизненных вершинах плато, напоминающих по своим условиям зональные арктические тундры или полярные пустыни (Романов, 2013; Романов и др., 2019). Экстремально сжатые (из-за значительно более поздних в сравнении с меньшими высотами сроков таяния снега) сроки гнездования на вершинах плато, скорее всего, не относятся к непреодолимым препятствиям для обитания хрустана.
Брачное поведение
В 1989 г. в долине р. Аян наиболее активные брачные демонстрации пришлись на 20 июня и весьма четко совпали с окончанием массового прохода диких северных оленей (Rangifer tarandus) по вершинам плато. К 25 июня брачные демонстрации заметно ослабели, а к 3 июля прекратились.
Более подробные сведения о брачном поведении хрустанов удалось получить в 2018 г., к которому относятся все описанные ниже в данном разделе наблюдения. Со дня нашего прибытия в котловину оз. Богатырь-Хуолу (18 июня) мы наблюдали токовые полеты самок – до 2–3 особей находились одновременно в поле зрения. Они летали порознь на высоте 30–50 м в разных направлениях широкими (радиусом 1–2 км) кругами или почти по прямой, скрываясь за горизонтом. На земле мы встречали их поодиночке, иногда 2 птицы гонялись друг за другом с “жужжанием”. Так, 18 июня наблюдали одиночную самку, которая отдыхала и кормилась, а затем, увидев вторую птицу (пол не определен), стала преследовать ее в полете.
Известно, что хрустаны в сезон гнездования образуют брачные пары, причем зачастую еще на пролете (Cramp, Simmons, 1983; Kålås, Byrkjedal, 1984; Pulliainen, Saari, 1996), однако пар мы практически не видели. Расчетные сроки откладки яиц в тот год пришлись в среднем на 10–12 июня. По-видимому, спустя неделю после завершения кладки, ко времени начала наших работ, большинство самок уже оставили своих самцов, и пары прекратили существование.
Активное токование самок в полете продолжалось примерно до середины 3-й декады июня. Группы токующих птиц встречали и на земле. Так, 20 июня обнаружена группа из 15 самок. Они собрались на склоне приречной террасы, где шумно, с “жужжанием”, бегали по земле, демонстрировали позы угрозы и периодически атаковали друг друга. Голова при этом была опущена вниз, спина – горбом, хвост – веером, оперение распушено – как это демонстрируют дерущиеся самцы галстучника (Charadrius hiaticula) (см.: Cramp, Simmons, 1983). Мы были свидетелями, как одна из самок прижала соперницу к земле и долго (с полминуты) не отпускала, не переставая клевать ее в спину. Время от времени бóльшая или меньшая часть птиц взлетала и улетала прочь от стаи, но вскоре возвращалась обратно. Ни одного самца в стае надежно разглядеть мы не сумели, хотя не исключено, что самец в стае все же был (возможно, даже в паре с самкой), и именно к нему слетелись холостые (или покинувшие самцов, приступивших к насиживанию кладок) самки, как это делают, например, холостые селезни речных уток (род Anas), преследуя брачные пары (см., например: Коблик, 2001). На следующий день токующих самок здесь мы не нашли – по-видимому, они токовали где-то в другом месте. Такое поведение птиц было похоже на “блуждающее токование” самцов турухтана или тетерева (Lyrurus tetrix) (см.: Рябицев, 2014). По-видимому, совместное токование самок хрустана на земле встречается нечасто, описания этого явления в литературе мы не нашли.
Второй раз группа из 5 токующих птиц встречена 24 июня в 1 км от первого места. Поведение их было таким же – они шумно бегали по земле, демонстрировали позы угрозы и периодически атаковали друг друга. Вскоре 3 птицы улетели, а две другие (вероятно, самец и самка, хотя внешне надежно различить их с расстояния 30 м не удалось даже по фотографиям) долгое время продолжали ходить вместе, издавая негромкие высокие трели (явно адресованные друг другу), и кормиться. Одна из них (самка), помимо этого, непрерывно токовала, вторая (очевидно, самец) периодически присаживалась, демонстрируя гнездовые ямки. Это был единственный эпизод, когда две птицы (предположительно, брачная пара) наблюдались вместе. К моменту нашего прибытия в район исследований большинство самцов уже более недели насиживали кладки, и отсутствие пар на фоне довольно частых встреч токующих самок подтверждает данные о том, что пары у хрустанов обычно формируются на очень непродолжительное время, лишь на несколько дней, до окончания кладки (Nethersole-Thompson, 1973; Cramp, Simmons, 1983). А описанную выше встречу предполагаемой пары, судя по срокам, можно интерпретировать проявлением последовательной полиандрии. Известно, что самка может отложить кладку второму самцу уже через 5 дней после завершения первой кладки (Cramp, Simmons, 1983; Pulliainen, Saari, 1992).
В дальнейшем группы из 3–9 самок, уже не проявлявших взаимной агрессии, мы встречали еще 5 раз – до 8 июля. Они вместе кормились, токовали, улетали вместе при вспугивании, издавая при взлете “жужжание”. Вскоре они перестали и токовать. Последний раз звуки токования на земле (монотонно повторяющийся короткий свист) слышали 1 июля в группе из 3 самок. Токование в полете мы продолжали регистрировать регулярно (1–2 раза в день) до 10 июля, к 14 июля оно полностью прекратилось. В горах Норвегии активное токование самок длится около месяца (Kålås, Byrkjedal, 1984), на плато Путорана, по нашим наблюдениям, – заметно меньше.
Большинство встреч самок мы регистрировали на участке размером 1.5 × 2 км (см. рис. 1), расположенном у подножия гор на пологих спускающихся к руслу реки террасах – в 0.5–1.5 км от реки, на высотах 870–970 м над ур. м., в среднем 905 ± 15 (SE), n = 7. При удалении от этого “центра” на 4–12 км число встреч хрустанов сокращалось в 5–10 раз. Вблизи этого “центра”, на контрольном участке площадью 4 км2, мы нашли 5 гнезд и 3 выводка с недавно вылупившимися (3–5 дней назад) птенцами, т.е., наверное, половину потомства в данной локальной группировке (судя по количеству токующих самок, при условии равного соотношения полов). Возможно, как многие виды птиц, для которых характерно совместное токование особей, – глухари (род Tetrao), тетерева и др. – хрустаны стремились устраивать гнезда в местах расположения токовищ или неподалеку от них. Известно, что самки турухтана, например, устраивают гнезда на расстоянии не более 400 м от токовища (Hayman et al., 1986). Кроме того, очевидно, что локальная группировка в данном месте сформировалась во многом благодаря расположению первых проталин на обращенных к югу склонах котловины озера (см. рис. 1). Это согласуется с данными о том, что пары обычно образуются из стаи в 5–8 особей каждого пола, которая собирается на плоской поверхности, часто поблизости от водного потока (Cramp, Simmons, 1983). По-видимому, на данном участке, который раньше других освободился от снега, осела пролетная стая птиц. Хотя в другие годы хрустаны появлялись весной на плато Путорана в основном парами и лишь изредка группами из 6–9 особей (см. ниже).
Фенология размножения
В гольцовом поясе долины р. Аян 20 июня 1989 г., когда горная тундра еще на 50% была покрыта снегом, найдено гнездо с одним свежим яйцом. В том же году в долине р. Неракачи у оз. Бокового (в 35 км севернее р. Аян) на границе гольцового и подгольцового поясов 4 августа неоднократно видели птиц, отводивших от молодых, которые к этому времени достигли 1/3 величины взрослой особи. Длина клюва, крыла, цевки одного из них составила соответственно 13, 60, 32 мм, кроющие плеча и спины распустились на 3/4 их длины, все маховые и верхние кроющие крыла находились в стадии кисточек, а на месте нижних кроющих крыла и контурных головы были заметны лишь пеньки.
В горной тундре у юго-западной оконечности оз. Кутарамакан, еще почти полностью покрытой снегом, 21 июня 1990 г. найдено гнездо с 3 ненасиженными яйцами. У средней части этого озера 5 августа наблюдали взрослую птицу, сопровождавшую нелетную молодую. Последняя была почти полностью оперена, достигала размеров взрослой особи, имела не полностью сформированные маховые и рулевые (длиной 10–20 мм), клюв, крыло, цевку (длиной соответственно 9, 113, 40 мм).
В гольцовом поясе у Гусиных озер 28 июня и 1 июля 2006 г. найдены 2 гнезда, в каждом из которых была полная кладка из 3 слабо насиженных яиц. Расчетные даты начала этих кладок 21 и 22 июня.
В нижней части гольцового пояса у восточной оконечности оз. Харпича 18 июня 2007 г. найдено гнездо с 3 ненасиженными яйцами.
В котловинах озер Богатырь и Нералах с 6 июля по 5 августа 2010 г. и в котловине оз. Негу-Икэн с 25 июня по 26 июля 2013 г. периодически отмечали особей, проявлявших беспокойство у гнезд или выводков. Самцы, сопровождающие птенцов, встречены 9 и 14 июля 2010 г.
В котловине оз. Богатырь-Хуолу в 2018 г. найдены 5 гнезд с полными кладками из 3 яиц. Птенцы в них появились довольно дружно: в двух гнездах 7 июля, еще в двух гнездах 8 июля и в одном гнезде 11 июля. Сроки начала гнездования определены по реконструированным датам начала насиживания кладок. Согласно данным о динамике изменения удельного веса яиц (ее определяли методом флотации), инкубация одного яйца длилась в среднем примерно 24 дня, в разных гнездах ее длительность различалась на 1–2 дня. В Шотландии и финской Лапландии инкубационный период длился дольше – около 26 дней (Nethersole-Thompson, 1973; Pulliainen, Saari, 1992). В дни, когда в известных нам гнездах появились птенцы (7–8 июля), мы встретили еще 3 выводка с птенцами 3–5-дневного возраста (уже без яйцевого зуба), в каждом выводке было по 3 птенца. С учетом данных об этих выводках, расчетная средняя дата откладки первого яйца в гнездах (n = 8) пришлась в 2018 г. на 10 июня ± 1 день (SE), наиболее ранняя дата – на 7 июня, наиболее поздняя дата – на 15 июня.
Когда 18 июня в 2018 г. мы прибыли в котловину оз. Богатырь-Хуолу, снег в долине реки и котловине озера уже практически растаял (оставались лишь снежники в горах). Вместе с тем, судя по нашим наблюдениям в окрестностях г. Норильска с 14 по 17 июня, весна в тот год была холодная и затяжная, таяние снега в горах началось, вероятно, не ранее 10 июня. И в эти же дни, как показывают наши расчеты, в гнездах хрустанов в районе исследований уже были яйца. Это говорит о том, что они начали гнездиться на первых проталинах при еще практически сплошном снежном покрове. В горах северной Скандинавии сроки гнездования вида более растянуты и приходятся на фенологически более поздние даты. Так, в Норвегии хрустаны прилетали в середине мая, занимали гнездовые участки в конце мая–начале июня, когда снег покрывал 75% поверхности, и через несколько дней начинали откладывать яйца. При этом самки продолжали токовать до второй недели июля, а сезон откладки яиц длился до 1 месяца (Kålås, Byrkjedal, 1984).
По наблюдениям за 4 гнездами в 2018 г., процесс вылупления птенцов в каждом из них занял менее суток. На севере Европы этот процесс длился от 12 часов (Pulliainen, Saari, 1992) или менее (Nethersole-Thompson (1973) до 24 часов (Kålås, Byrkjedal, 1984). В котловине оз. Богатырь-Хуолу видеонаблюдение за одним из гнезд показало, что между появлением первого и третьего птенцов прошло 16 часов, затем вся семья продолжала оставаться в гнезде еще 1.5 суток. Самец временами оставлял птенцов и уходил кормиться, отсутствуя не дольше минуты. Пока он грел младшего птенца, двое старших тоже периодически покидали гнездо и гуляли в его окрестностях, что-то склевывая с поверхности грунта и растений, и затем возвращались обратно в гнездо под брюхо самца.
Итак, по обобщенным данным (n = 13) за 1988–2018 гг., хрустаны приступали к откладке яиц с 7 по 22 июня, в среднем 13 июня ± 2 дня (SE), медиана 12 июня. В дни, когда в гнездах хрустанов появлялись первые яйца, горная тундра часто еще полностью оставалась покрытой снегом. Наиболее ранняя расчетная дата появления птенцов 3 июля, наиболее поздняя – 18 июля, средняя – 9 июля ± 1 день (SE), медиана – 8 июля.
Устройство гнезд, размеры кладок и яиц
Все известные нам полные кладки хрустана (n = 9) содержали по 3 яйца.
Гнездо, найденное в долине р. Аян, представляло собой слабозаметное углубление диаметром 9 см и глубиной 5 см в кочке, образованной мхом, осокой и дриадой среди мерзлотных медальонов.
Гнездо, обнаруженное у оз. Кутарамакан, представляло собой бесформенное углубление в грунте диаметром 13 см и глубиной 1 см и было расположено на вытаявшем глинисто-щебнистом бугре размером 10 × 15 м, покрытом мхами, лишайниками, дриадой, стелющимся ерником, ивкой и багульником (Ledum palustre). Размеры яиц: 29.2 × × 41.3, 29.8 × 42.5, 29.0 × 42.5 мм; их вес соответственно: 18.0, 19.15, 18.1 г.
В горной тундре у Гусиных озер 2 осмотренных гнезда располагались на высотах 875 и 890 м над ур. м. Они были устроены на плоских поверхностях террас, покрытых дриадой, лишайником, мхом, кассиопеей, осокой. Не менее 30% площади этих местообитаний занимали каменистые россыпи и мерзлотные медальоны. Гнезда выглядели как ямки в примятой растительной куртинке, их внешний диаметр: 12 и 14 см, глубина лотка: 4 и 4.5 см. Размеры яиц (n = 6): 39.2–43.8 × × 28.9–30.1, в среднем 41.3 × 29.4 мм.
Найденное у оз. Харпича гнездо располагалось в горной тундре нижней части гольцового пояса (850 м над ур. м.) в 150 м от бурного ручья и в 15 м от одиночных угнетенных лиственниц высотой 0.5–0.8 м. Оно представляло собой углубление на поверхности сырой, мохово-злаково-дриадовой кочки, расположенной среди каменистых россыпей, и было выстлано сухими листьями ивы и арктоуса (Arctous alpina). Внешний диаметр гнезда 12 см, глубина – 6 см. Размеры яиц: 29.1 × 43.0, 30.2 × 42.8, 29.3 × 44.1 мм.
В котловине оз. Богатырь-Хуолу обнаруженные гнезда (n = 5) были устроены на ледниковых моренах, пологих террасах с углом наклона до 10° (по глазомерным оценкам), среди камней и редкой растительности с проективным покрытием 30–50%. Выстилка 5 гнездовых лунок состояла из измельченного растительного мусора толщиной около 1 см. Средний диаметр лотка (n = 3): 10.3 ± ± 0.2 (SE) см, глубина – 4.2 ± 0.9 см. Средние размеры яиц (n = 12): 28.13 ± 0.19 × 41.74 ± 0.34 мм (27.2–29.2 × 40.4–43.6). Высота местности, где были обнаружены гнезда и выводки, составила в среднем 941 ± 26 м над ур. м. (от 871 до 1036, n = 8).
Таким образом, все найденные в 1988–2018 гг. гнезда располагались на высоте от 850 до 1036 м над ур. м., в среднем 935 ± 28 (n = 8). Они представляли собой углубление в грунте диаметром от 9 до 14 см, в среднем 11, и глубиной от 1 до 6 см, в среднем 4 (n = 8). Размеры яиц (n = 25) составили в среднем 28.80 ± 0.17 × 41.88 ± 0.25 мм (27.2–30.2 × × 39.2–44.1).
Успешность гнездования
Скорость таяния снежного покрова и степень его мозаичности весной в значительной мере определяют распределение гнезд по территории, сроки откладки яиц и саму возможность гнездования. По-видимому, сильные снегопады в начале лета могут стать причиной гибели кладок (Cramp, Simmons, 1983), о чем косвенно свидетельствуют и наши наблюдения в конце июня 1988 и 1989 гг. за вертикальными перемещениями хрустанов (см. ниже). В годы с благоприятными погодными условиями случаи гибели кладок или яиц не отмечены. Так, в 2018 г. во всех 5 гнездах, которые находились под наблюдением, благополучно вывелись все птенцы. Следует отметить, что в тот год успешность гнездования у всех видов птиц была высокой. Из 39 гнезд, судьбу которых удалось проследить, неудачными оказались лишь 4. Из отложенных всеми видами 167 яиц благополучно вывелись и покинули гнезда 144 птенца, таким образом, общая успешность гнездования птиц составила 86% (Романов и др., 2018). Столь высокая успешность гнездования, помимо благоприятных погодных условий, была обусловлена также низким уровнем хищничества на фоне обилия мелких грызунов. Здесь, в котловине оз. Богатырь-Хуолу мы не встречали песцов (Vulpes lagopus), лисиц (V. vulpes), лишь однажды был отмечен горностай (Mustela erminea). Из птиц потенциальным разорителем гнезд был лишь длиннохвостый поморник (Stercorarius longicaudus), но в качестве кормовых объектов его в полной мере обеспечивали многочисленные полевки Миддендорфа (Microtus middendorffi) и копытные лемминги (Dicrostonyx torquatus). В другие годы судьбу найденных гнезд проследить не удалось.
Реакция насиживающих птиц на фактор беспокойства
В 2018 г. при контрольных посещениях гнезд мы заставали на кладках только самцов. Самки участия в насиживании не принимали, хотя известно, что в других частях ареала это иногда случается (Cramp, Simmons, 1983; Lücker et al., 2011; Bassi et al., 2014). Поведение самцов при приближении человека было разным: в 13 случаях из 31 они затаились и взлетели в самый последний момент из-под ног, в 18 случаях – незаметно сошли с гнезда заранее. При этом 4 самца 10 раз подпустили человека вплотную к гнезду и лишь 5 раз сошли с него заблаговременно. А пятый самец, на которого пришлась половина всех наблюдений (из-за непосредственной близости гнезда к нашему палаточному лагерю), почти каждый раз (в 13 визитах из 16) заранее незаметно сходил с гнезда. Этот же самец 4 июля трижды кратковременно сходил с насиживаемой кладки, чтобы отогнать оказавшегося вблизи гнезда галстучника.
Сроки миграций
Во время весеннего движения хрустанов их встречали в основном парами и лишь изредка группами из 6–9 особей. Наиболее ранние даты встреч – 5 июня 1988 г. в районе оз. Аян, 14 июня 1989 г. в среднем течении р. Аян (Романов, 1996). В этих районах птицы появились в горной тундре, когда более 30% ее поверхности было покрыто не стаявшим снегом. В некоторые годы хрустан прилетает фенологически еще раньше, когда снежный покров находится в зимнем состоянии, а его таяние только начиналось. Подобная ситуация была зафиксирована нами, например, 16 июня 1990 г. у оз. Кутарамакан, а Морозовым (1984) – 25 июня 1980 г. у оз. Капчук.
Сильные снегопады и похолодание 20–22 июня 1988 г. заставили птиц, державшихся на вершинах плато, спуститься в межгорные долины. В это время продолжался прилет хрустанов, которые из-за неблагоприятной погоды не имели возможности переместиться в горную тундру. В результате птицы скапливались на болотах среди лиственничников, в редколесьях, на береговых галечниках оз. Аян, заросших злаками. Видимо, даже у воды не все могли прокормиться, т.к. на берегу озера находили погибших от истощения птиц (n = 5). Скудное содержимое желудков двух вскрытых особей состояло из измельченной растительной массы, ягод шикши, личинок комаров и гастролитов. В 1989 г. в долине р. Аян 24 и 25 июня сильные снегопады стимулировали вертикальные перемещения и концентрации птиц у реки, подобные тем, что происходили годом ранее у оз. Аян (Романов, 1996).
Вероятно, сильные снегопады также стали причиной кочевок хрустанов 23–25 июля 2010 г. у берегов озер Богатырь и Нералах, где активно перемещавшиеся птицы держались одиночно или группами по 2–5 особей.
Осенью большинство птиц отлетает из региона, вероятно, к 20 августа. Из бассейна р. Аян в 1989 г. хрустаны исчезли к 16 августа. Отлет хрустанов из окрестностей оз. Капчук Морозов (1984) в 1980 г. наблюдал 16–20 августа. В 1990 г. у оз. Кутарамакан интенсивный осенний пролет шел 18–19 августа и совпал с резким похолоданием и мощными снегопадами, образовавшими сплошной снежный покров в гольцовом и подгольцовом поясах плато. В эти дни стаи из 10–30 хрустанов постоянно летели низко над водой на юго-запад, а на берегах озера повсеместно встречались одиночки, пары и группы из 3–4 особей, собиравших корм.
ЗАКЛЮЧЕНИЕ
Установлено, что хрустан распространен по территории плато Путорана почти повсеместно, достоверно гнездится в его северной, западной и восточной частях. В гнездовой период он осваивает широкий диапазон высот, населяя преимущественно плоские вершины гольцового пояса, локально – некоторые участки подгольцового пояса и крайне редко встречаясь в пределах горно-таежного пояса. Высотное распределение вида обусловлено не столько абсолютной высотой местности, сколько экологическими параметрами предпочитаемых местообитаний. Ареал вида в пределах Путорана имеет явно выраженный трехмерный характер.
Хрустан – типичный обитатель горно-тундровых ландшафтов вершин плато, характерный элемент авифауны гольцового пояса, входящий в число видов, формирующих ее ядро. В гнездовой период держится преимущественно в интервале 890–1070 м над ур. м. Предпочитает участки с небольшим уклоном, имеющие бугристый микрорельеф и покрытые скудной травянистой растительностью. Оптимальные местообитания – сухие каменистые мохово-лишайниковые тундры с различной долей участия дриады, кассиопеи и осоки, обилием каменистых и щебнистых россыпей, а также мерзлотных медальонов. Охотно держится на вершинах крупных щебнистых бугров, поверхность которых, если не считать лишайниковых куртин, почти лишена всякой растительности.
Распределение хрустана в пространстве неравномерно. Обилие в оптимальных горно-тундровых местообитаниях гольцового пояса в гнездовой период варьирует в пределах 0.4–10.0 особей/км2, составляя в среднем 1.7 особей/км2. Обилие вида максимально в средней части гольцового пояса и постепенно сокращается в сторону как вершин, так и подножий склонов. Кроме того, очаги повышенной плотности можно наблюдать на относительно небольших участках местности, где формируются локальные центры токовой активности самок. В таких случаях птицы придерживаются ограниченной территории, на которой не только совершают обычные для этого вида одиночные рекламные полеты, но и собираются на земле для совместного токования группами до полутора десятков особей. К таким “центрам” может быть приурочено и распределение гнезд на местности.
Птицы прилетают в места размножения в основном парами, которые распадаются вскоре после откладки яиц. Покинув приступивших к насиживанию самцов, самки еще около двух недель продолжают активно токовать, пытаясь привлечь новых самцов, затем активность токования заметно снижается и по истечении еще двух недель полностью прекращается. В годы наших исследований птицы начинали откладывать яйца с 7 по 22 июня, в среднем – 13 июня ± 2 дня (n = 13). Экстремальные погодно-климатические условия в гнездовой период не являются непреодолимыми для хрустана. Он может приступать к гнездованию, даже когда гольцовые вершины плато еще полностью покрыты снегом. В целом, сроки размножения хрустанов на плато Путорана более сжаты, чем в североевропейской части ареала, а отдельные фазы этого цикла имеют меньшую продолжительность.
Найденные гнезда располагались на высоте от 850 до 1036 м над ур. м., в среднем 935 ± 28 (n = 8). Они представляли собой углубление в грунте диаметром от 9 до 14 см, в среднем 11, и глубиной от 1 до 6 см, в среднем 4 (n = 8). Все полные кладки (n = 9) содержали по 3 яйца. Размеры яиц (n = 25) составили в среднем 28.80 ± 0.17 × 41.88 ± 0.25 мм (27.2–30.2 × 39.2–44.1). Наиболее ранняя расчетная дата появления птенцов – 3 июля, наиболее поздняя – 18 июля, средняя – 9 июля ± 1 день (n = 13). Птенцы из одного выводка появляются на свет в течение одних суток. Случаев разорения гнезд хищниками или гибели яиц из-за неблагоприятных погодных условий не отмечено. Примерно к 20 августа птицы покидают места гнездования.
Список литературы
Водопьянова Н.С., 1976. О влиянии озер Путорана на прибрежную растительность // Природно-ландшафтные основы озер Путорана // Труды Лимнологического ин-та СО АН СССР. Т. 22 (42). Новосибирск. С. 86–92.
Гвоздецкий Н.А., Голубчиков Ю.Н., 1987. Горы. М.: Мысль. 399 с.
Голубчиков Ю.Н., 1996. География горных и полярных стран. М.: Изд-во МГУ. 304 с.
Зырянов В.А., 1988. Орнитофауна окрестностей оз. Нерангда // Животный мир плато Путорана, его рациональное использование и охрана. Сб. науч. трудов ВАСХНИЛ, Сиб. отделение НИИ СХ Крайнего Севера. Новосибирск. С. 88–96.
Коблик Е.А., 2001. Разнообразие птиц (по материалам экспозиции Зоологического музея МГУ). Ч. 1. М.: Изд-во МГУ. 384 с.
Кречмар А.В., 1966. Птицы Западного Таймыра // Биология птиц. М.–Л.: Изд-во АН СССР. С. 185–312.
Куваев В.Б., 2006. Флора субарктических гор Евразии и высотное распределение ее видов. М.: Товарищество научных изданий КМК. 568 с.
Лаппо Е.Г., Томкович П.С., Сыроечковский Е.Е.-мл., 2012. Атлас ареалов гнездящихся куликов Российской Арктики. М.: УФ Офсетная печать. 448 с.
Морозов В.В., 1984. Орнитофауна окрестностей оз. Капчук, плато Путорана // Орнитология. Вып. 19. С. 30–40.
Норин Б.Н., 1986. Общая характеристика растительности // Горные фитоценологические системы Субарктики. Л.: Наука. С. 164–168.
Павлов Б.М., Александрова А.С., Шелковникова Т.А., 1988. Природные комплексы и их охрана // Животный мир плато Путорана, его рациональное использование и охрана. Сб. науч. трудов ВАСХНИЛ. Сиб. отделение. НИИСХ Крайнего Севера. Новосибирск. С. 102–117.
Пармузин Ю.П., 1964. Средняя Сибирь. М.: Наука. 310 с.
Пармузин Ю.П., 1976. Основные особенности массо- и энергообмена в озерных котловинах путоранской провинции Субарктики // Природно-ландшафтные основы озер Путорана. Новосибирск. С. 4–10.
Равкин Ю.С., 1967. К методике учета птиц лесных ландшафтов // Природа очагов клещевого энцефалита на Алтае. Новосибирск: Наука. С. 66–75.
Рогачева Э.В., 1988. Птицы Средней Сибири: распространение, численность, зоогеография. М.: Наука. 309 с.
Романов А.А., 1996. Птицы плато Путорана. М.: тип. Россельхозакадемии. 297 с.
Романов А.А., 2006. Видовой состав, численность и ландшафтно-биотопическое размещение птиц в бассейне р. Северной // Изучение и охрана животных сообществ плато Путорана. М. С. 9–70.
Романов А.А., 2006а. Фауна и население птиц центральной части котловины оз. Кета // Изучение и охрана животных сообществ плато Путорана. М. С. 71–102.
Романов А.А., 2013. Авифауна гор Азиатской Субарктики: закономерности формирования и динамики. М.: Русское общество сохранения и изучения птиц им. М.А. Мензбира. 475 с.
Романов А.А., 2015. Авифауна плато Путорана. М.: Русское общество сохранения и изучения птиц им. М.А. Мензбира. 196 с.
Романов А.А., Рупасов С.В., Журавлев Е.А., Голубев С.В., 2007. Птицы бассейна р. Курейки // Биоразнообразие экосистем плато Путорана и сопредельных территорий. М. С. 7–70.
Романов А.А., Тарасов В.В., Мелихова Е.В., Тимченко А.С., Зарубина М.А., Яковлев В.О., 2018. Авифауна котловины озера Богатырь-Хуолу (северо-запад плато Путорана, Красноярский край) // Фауна Урала и Сибири. № 2. С. 92–104.
Романов А.А., Мелихова Е.В., Зарубина М.А., 2019. Птицы гор Северной Азии: итоги исследований 2010–2018 гг. М.: Русское общество сохранения и изучения птиц им. М.А. Мензбира. 240 с.
Рупасов С.В., Журавлев Е.А., 2006. Орнитофауна долины р. Микчангда и прилегающих территорий // Изучение и охрана животных сообществ плато Путорана. М. С. 122–154.
Рябицев В.К., 2014. Птицы Сибири: справ.-определитель. Т. 2. М.; Екатеринбург: Кабинетный ученый. 452 с.
Рябицев В.К., Алексеева Н.С., Тюлькин Ю.А., 2003. К распространению, биологии, экологии и поведению турухтана Philomachus pugnax на Ямале // Русский орнитологический журнал. Вып. 243. С. 1277–1290.
Степанян Л.С., 2003. Конспект орнитологической фауны России и сопредельных территорий. М.: Наука. 727 с.
Томкович П.С., 2007. Динамика численности хрустана: тревожные мысли на “зыбкой почве” // Информационные материалы Рабочей группы по куликам. Вып. 20. С. 43–45.
Bassi E., Vigano E., Scieghi G., 2014. New breeding of the Eurasian Dotterel, Eudromias morinellus, in the Italian Alps // Rivista Italiana di Ornitologia – Research in Ornithology. V. 84 (1). P. 5–10.
Cramp S., Simmons K.E.L. (eds), 1983. The Birds of the Western Palearctic. Oxford: Oxford University Press. V. 3. 913 p.
Hable E., Präsent I., 1990. Die Forschungsstatte “Pater Blasius Hanf” am Furtnerteich (Gemeinde Mariahof, Bezirk Murau) // Schwarzenbergische Archive. P. 181–257.
Hayman P., Marchant J., Prater T., 1986. Shorebirds: an identification guide to the waders of the world. Boston: Houghton Mifflin. 412 p.
Kålås J.A., Byrkjedal I., 1984. Breeding chronology and mating system of the Eurasian Dotterel (Charadrius morinellus) // Auk. V. 101. P. 838–847.
Liebezeit J.R., Smith P.A., Lanctot R.B., Schekkerman H., Tulp I., Kendall S.J., Tracy D.M., Rodrigues R.J., Meltofte H., Robinson J.A., Gratto-Trevor C., Mccaffery B.J., Morse J., Zack S.W., 2007. Assessing the development of shorebird eggs using the flotation method: species-specific and generalized regression models // Condor. V. 109. P. 32–47.
Lücker L., Kraatz S., Kraatz B., 2011. Field notes on the breeding biology of the Dotterel Charadrius morinellus in arctic Norway // Ornis Svecica. V. 21. P. 109–118.
Nethersole-Thompson D., 1973. The Dotterel. London: Collins. 288 p.
Pulliainen E., Saari L., 1992. Breeding biology of the Dotterel Charadrius morinellus in eastern Finnish Lapland. Ornis Fennica. V. 69. P. 101–107.
Pulliainen E., Saari L., 1996. Pre- and non-breeding biology of Dotterel Charadrius morinellus on Varriotunturi fell area, NE Finland // Wader Study Group Bull. V. 81. P. 54–58.
Rahn H., 1974. The avian egg: incubation time and water loss // Condor. V. 76 (2). P. 147–152.
Saari L., 1995. Population trends of the dotterel Charadrius morinellus in Finland during the past 150 years // Ornis Fennica. V. 721. P. 29–36.
Westerkov K., 1950. Method for determining the age of game bird eggs // Journal of Wildlife Management. V. 14. P. 56–57.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал