

Зоологический журнал, 2021, T. 100, № 8, стр. 927-937
Биоакустическая, генетическая и морфологическая изменчивость некоторых подвидов варакушки (Cyanecula svecica, Muscicapidae, Aves) в центральной части ареала
О. А. Дмитриева a, *, В. В. Иваницкий a, **, Я. А. Редькин b, ***, М. М. Белоконь c, ****, Е. Л. Мацына d, *****, И. М. Марова a, ******
a Биологический факультет Московского государственного университета
имени М.В. Ломоносова
119991 Москва, Россия
b Научно-исследовательский Зоологический музей Московского государственного университета
имени М.В. Ломоносова
125009 Москва, Россия
c Институт общей генетики имени Н.И. Вавилова РАН
119991 Москва, Россия
d Экоцентр “Дронт”
603000 Новгород Нижний, Россия
* E-mail: neun.welle@gmail.com
** E-mail: vladivanit@yandex.ru
*** E-mail: yardo@mail.ru
**** E-mail: mbelokon@gmail.com
***** E-mail: elmatzuna@yandex.ru
****** E-mail: collybita@yandex.ru
Поступила в редакцию 10.07.2020
После доработки 16.08.2020
Принята к публикации 08.09.2020
Аннотация
В центральной части ареала изучали биоакустическую, генетическую и морфологическую изменчивость варакушки (Cyanecula svecica) – политипического вида c транспалеарктическим распространением и сложной подвидовой дифференциацией. Наибольшее внимание уделено полиморфному подвиду C. s. volgae, по всему ареалу которого совместно встречаются белозвездные, рыжезвездные и промежуточные морфотипы. Проведенный анализ свидетельствует о незначительном в целом уровне дивергенции C. s. volgae по сравнению с тремя сопредельными подвидами (C. s. cyanecula, C. s. svecica и C. s. pallidogularis) по всем изученным признакам. По песне отличие C. s. volgae наблюдается только при сравнении с C. s. cyanecula, по морфологическим признакам C. s. volgae широко перекрывается со всеми подвидами, по двум изученным нами генетическим маркерам отличий от C. s. svecica и C. s. pallidogularis также не выявлено. Таким образом, основной спецификой подвида C. s. volgae остается его ярко выраженный полиморфизм. При этом значимых различий между рыжезвездными и белозвездными морфотипами мы не выявили ни по вокальным, ни по размерным, ни по генетическим характеристикам, вследствие чего форму C. s. volgae можно рассматривать как единый самостоятельный подвид. Высказано предположение о том, что возникновение современной формы C. s. volgae могло происходить в результате трансформации местообитаний хозяйственной деятельностью человека, которая привела к возникновению зоны вторичного контакта европейской белозвездной расы C. s. cyanecula и северной рыжезвездной формы C. s. svecica.
Варакушка (Cyanecula svecica (Linnaeus 1758)) – политипический вид со сложной подвидовой дифференциацией. В границах ее огромного ареала, охватывающего почти всю Палеарктику и часть Неарктики (Аляска), выделяют до 15 подвидов (географических рас) (Tugarinov, 1929; Козлова, 1945; Степанян, 1990). Географическая изменчивость проявляется в общих размерах и пропорциях тела, окраске верхней стороны тела и головы. У самцов варьирует интенсивность голубого тона груди (пластрона). Но самым ярким проявлением морфологической изменчивости варакушки, неизменно привлекающим к себе внимание, являются наличие, цвет и размеры центрального пятна на грудном пластроне – так называемой звезды. Известны беззвездные, белозвездные и рыжезвездные морфотипы, в ряде районов распространены полиморфные популяции, где совместно встречаются самцы с разными типами окраски пластрона и не представляют редкости промежуточные варианты (Сушкин, 1925; Tugarinov, 1929; Портенко, 1954; Мацына, 2006; Pavel, Chutny, 2013; Petrilák, 2014).
По молекулярно-генетические данным, базальной формой считают беззвездный испанский подвид C. s. аzuricollis (Rafinesque 1814) и эволюционно тесно связанные с ним подвиды белозвездных варакушек C. s. cyanecula (Meisner 1804) и C. s. namneum (Mayaud 1934), тогда как все рыжезвездные варакушки – производные от него (Questiau et al., 1998; Hogner et al., 2013 цит. по: Petrilák, 2014; Zink et al., 2003; Johnsen et al., 2006). Межпопуляционная дифференциация мтДНК незначительна и не соответствует делению на подвиды, основанному на морфологических признаках. Достоверно различаются лишь все южные и все северные популяции, причем широко распространенный северный подвид C. s. svecica генетически более однороден (Zink et al., 2003). Анализ микросателлитов выявил значительно большее соответствие принятой подвидовой структуре (Johnsen et al., 2006). В частности, найдены различия между белозвездным C. s. cyanecula, с одной стороны, и рыжезвездными C. s. svecica и C. s. pallidogularis Zarudny 1897, с другой. Различия между C. s. svecica и C. s. pallidogularis гораздо менее значительные, нежели между каждым из них и C. s. cyanecula (Johnsen et al., 2006). В упомянутой работе не использовали экземпляры из ареала C. s. volgae (O. Kleinschmidt 1907), поскольку, как полагают авторы, это могло привести к размыванию границ между выборками и утрате четко выраженных генетических паттернов, соответствующих разным подвидам.
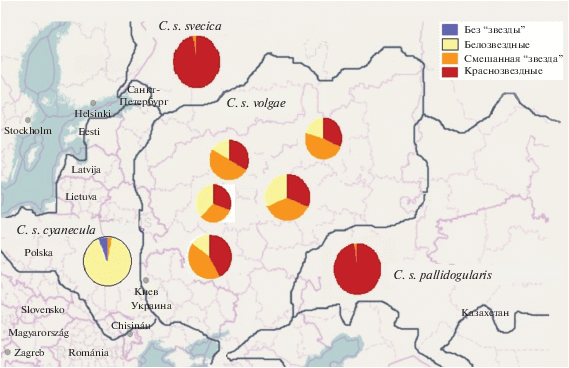
Между тем подвид C. s. volgae, распространенный в центральной России и в восточных областях Украины и Белоруссии, представляет особый интерес. Он обитает почти в центре видового ареала и граничит с другими подвидами: белозвездным C. s. cyanecula на западе, рыжезвездными svecica (на севере и северо-востоке) и C. s. pallidogularis (на востоке и юго-востоке). По всему ареалу C. s. volgae совместно, но в разной пропорции, встречаются белозвездные и рыжезвездные морфы, а также разнообразные промежуточные варианты окраски (рис. 1). Предполагают, что C. s. volgae представляет собой результат гибридизации рыжезвездных и белозвездных варакушек (Tugarinov, 1929; Матюхин, 2000; Мацына, 2006; Petrilák, 2010), однако природа полиморфизма C. s. volgae до конца не разгадана.
Рис. 1.
Распространение подвидов C. s. cyanecula, C. s. svecica, C. s. pallidogularis, C. s. volgae и изменчивость окраски нагрудного пятна (пластрона) внутри подвида C. s. volgae (по данным анализа коллекционных материалов).
Вопрос о подвидовых особенностях песни варакушки – с учетом ее впечатляющего таксономического разнообразия – изучен недостаточно. Нам известна единственная публикация на эту тему о сравнении песни двух подвидов – рыжезвездного C. s. svecica и белозвездного C. s. cyanecula. По данным Turčoková et al. (2010) C. s. svecica обладают более низкочастотной песней, нежели C. s. cyanecula, что авторы связывают с разной биотопической приуроченностью этих подвидов. Прочие подвиды варакушки с биоакустической точки зрения не изучены.
Цель нашей работы состоит в том, чтобы изучить биоакустическую и генетическую изменчивость в полиморфной популяции подвида C. s. volgae, оценить сходство и различия в пении самцов белозвездного, рыжезвездного и промежуточного морфотипов, проанализировать морфологические признаки, а также провести сравнение с данными по сопредельным подвидам: C. s. cyanecula, C. s. svecica и C. s. pallidogularis.
МАТЕРИАЛЫ И МЕТОДЫ
Нами записаны 27 самцов C. s. volgae близ Нижнего Новгорода (16 рыжезвездных, 7 белозвездных и 4 с промежуточной окраской). Подробное описание этой полиморфной популяции опубликовано (Мацына, 2006). Пение 13 самцов C. s. pallidogularis записала Л.А. Немченко в заказнике Дьяковский лес в Саратовской обл. Записи сделаны на цифровой диктофон OLYMPUS LS-5 с выносным конденсаторным микрофоном Philips SBC ME570 и параболическим рефлектором Sony PBR-330 и на профессиональный магнитофон Marantz PMD 660 c конденсаторным микрофоном Sennheiser МЕ 66 с предусилителем К6. Все записи выполнены в формате Windows PCM (wav). От одного самца делали по одной записи песни в течение 10 мин. Для каждого из записанных самцов в полиморфной популяции C. s. volgae определена окраска нагрудного пятна; 17 из этих самцов были пойманы для измерений, описания окраски и взятия крови для генетического анализа.
Использованы также 15 фонограмм, находящихся в свободном доступе на сайте www.xeno-canto.org (подвиды C. s. cyanecula и C. s. svecica). Общий объем данных, участвующих в сравнении вокализации варакушек из разных регионов, составил 358 песен.
Записи проанализированы в программе Syrinx v.2.6. Мы применяли стандартную методику иерархического описания песни птиц с выделением вокальных компонентов разного уровня: элементов (нот), слогов и фраз (Панов, Опаев, 2011; Иваницкий и др., 2013). Элементы – это компоненты, изображаемые на сонограмме в виде отдельных сплошных (не имеющих внутри себя перерывов) фигур. Слоги – компоненты, состоящие из 2–3 элементов, разделенных короткими паузами и всегда исполняемых вместе. Фразы – последовательности, состоящие из сходных комбинаций нот (слогов), или же комбинации разных слогов, которые всегда исполняются вместе.
Для каждой отдельной песни измеряли ряд параметров, следуя методике Turčoková et al. (2010): 1) максимальная частота, 2) минимальная частота, 3) частотный диапазон, 4) длительность песни, 5) число слогов, 6) число типов слогов, 7) число переключений между разными типами слогов, 8) число фраз, 9) пластичность – отношение числа переключений между фразами к максимально возможному числу переходов в песне данной длительности (пластичность песни рассчитывается так: число переходов/(число фраз в песне 1), 10) разнообразие (отношение количества слогов в песне к количеству типов слогов в песне). Поскольку из каждой записи было изучено несколько песен от одного самца, то для того чтобы избежать псевдорепликации, для дальнейшего анализа использовали средние значения по каждой записи (от каждого самца брали по возможности не менее 5 песен для получения усредненного значения). Каждое среднее значение для самца варакушки принимали за отдельное независимое наблюдение. Результаты измерений приведены в табл. 1.
Таблица 1.
Параметры песни четырех подвидов варакушки
| Подвиды и морфы | n | Fmin | Fmax | Fdif | Длина песни | Число элементов | Число типов элементов | Число переходов | Разнообразие | Размер репертуара | Пластичность | Скорость пения |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C. s. volgae (красная звезда) | 15 | 7.69 ± ± 0.36 | 2.05 ± ± 0.20 | 5.64 ± ± 0.52 | 7.72 ± ± 2.52 | 26.96 ± ± 6.72 | 9.03 ± ± 1.21 | 11.36 ± ± 2.5 | 3.38 ± ± 1.31 | 43.36 ± ± 13.25 | 0.33 ± ± 0.07 | 3.50 ± ± 1.52 |
| C. s. volgae (белая звезда) | 8 | 7.63 ± ± 0.69 | 2.18 ± ± 0.22 | 5.44 ± ± 0.82 | 9.01 ± ± 1.69 | 26.94 ± ± 6.74 | 8.56 ± ± 1.51 | 10.79 ± ± 2.22 | 3.51 ± ± 0.78 | 50.66 ± ± 16.86 | 0.34 ± ± 0.06 | 3.92 ± ± 0.93 |
| C. s. volgae (смешанная звезда) | 6 | 7.22 ± ± 0.76 | 2.09 ± ± 0.32 | 5.13 ± ± 0.92 | 10.14 ± ± 5.79 | 25.2 ± ± 10.8 | 9.44 ± ± 2.69 | 11.09 ± ± 5.30 | 3.03 ± ± 1.30 | 48.5 ± ± 20.3 | 0.38 ± ± 0.26 | 2.75 ± ± 0.45 |
| C. s. cyanecula | 7 | 7.44 ± ± 0.15 | 1.63 ± ± 0.39 | 5.81 ± ± 0.43 | 11.15 ± ± 1.76 | 31.97 ± ± 2.36 | 10.16 ± ± 0.76 | 13.09 ± ± 2.43 | 3.35 ± ± 0.68 | 50.33 ± ± 14.3 | 0.17 ± ± 0.03 | 4.42 ± ± 1.13 |
| C. s. pallidogularis | 10 | 7.39 ± ± 0.29 | 2.17 ± ± 0.34 | 5.22 ± ± 0.33 | 7.57 ± ± 2.48 | 24.24 ± ± 5.78 | 8.50 ± ± 1.33 | 11.24 ± ± 2.63 | 3.27 ± ± 0.94 | 46.75 ± ± 9.95 | 0.22 ± ± 0.05 | 4.05 ± ± 0.89 |
| C. s. svecica | 7 | 7.37 ± ± 0.65 | 1.77 ± ± 0.28 | 5.60 ± ± 0.78 | 9.58 ± ± 2.32 | 33.83 ± ± 9.87 | 9.37 ± ± 1.63 | 12.96 ± ± 2.31 | 4.09 ± ± 1.81 | – | 0.14 ± ± 0.03 | 3.85 ± ± 0.69 |
Для генетического анализа взяты образцы крови 17 самцов из полиморфной популяции C. s. volgae близ Нижнего Новгорода. Кровь брали из подключичной вены на бумажные фильтры. Для сравнения с “чистыми” белозвездными C. s. cyanecula (Белоруссия) использовали спиртовой мышечный материал (4 экз.) и фрагменты кожи с тушек (3 экз.) из коллекции Зоологического музея МГУ (сборы А. Шаповала, Н. Дацкевича и В. Гаврина). Анализировали два участка мтДНК – цитохром b (белок-кодирующий регион, 456 п.н.) и контрольный регион (850 п.н.). ДНК выделяли наборами Diatom™ DNA Prep 100 (ООО “Лаборатория Изоген”). Качество выделенной ДНК проверяли при помощи электрофореза 2–3 мкл образца в 0.7–1.0% агарозном геле (Белоконь и др., 2014). Для амплификации, секвенирования и очистки применяли наборы реагентов GenePak® PCR Core и Diatom™ DNA Clean-Up (ООО “Лаборатория Изоген”). Использовали праймеры, указанные в работе Zink et al. (2003): L14841 (5'-CCATCCAACATCTCAGCATGATGAAA-3'); H15299 (5'-GGAGGAAGTGCAGGGCGAAGAATCG-3') для цитохрома b и LCR4 (5′-CTCACGAGAACCGAGCTACT-3′); H1248 (5′‑CATCTTCAGTGTCATGCT-3′) для контрольного региона. После амплификации качество проверяли на наличие продуктов в 1.5% агарозном геле. Полученные продукты реакций хранили при температуре –20°С. Секвенирование ДНК проводили в фирме “Евроген” (Москва). Идентификацию осуществляли путем сравнения полученных последовательностей нуклеотидов с базой данных NCBI (National Center for Biotechnology Information, США). Сравнение полученных последовательностей проводили в программе Bioedit Sequence Alignment Editor 7.2.6.
Морфологические признаки варакушек изучали по коллекциям Научно-исследовательского Зоологического музея МГУ, Зоологического института РАН, кафедры зоологии позвоночных биологического факультета МГУ, Государственного Дарвиновского музея, Кировского городского зоологического музея, Института биологии и химии Московского педагогического государственного университета, Сибирского федерального университета (Красноярск), личной коллекции В.В. Гричика (Минск). Также использованы прижизненные измерения 17 птиц, пойманных нами близ Нижнего Новгорода. Измеряли длину крыла, хвоста, цевки, клюва от переднего края ноздри, клюва от верхнего края рамфотеки, а также длину вершины, а также определяли формулу крыла. Рассчитаны индексы отношения длины хвоста к длине крыла и индексы отношения длина вершины к общей длине крыла. Результаты измерений и число изученных экземпляров приведены в табл. 2. При изучении окраски оперения самцов мы использовали шкалу цветов (Smithe, 1975) и эталонные серии экземпляров, с помощью которых оценивали: 1) интенсивность синего цвета на груди по (3 градации), 2) ширину рыжей полосы на груди, 3) размер центрального нагрудного пятна (4 градации), 3) цвет центрального нагрудного пятна (7 градаций).
Таблица 2.
Размеры взрослых самцов четырех подвидов варакушки
| Признак | Подвиды и число измерений (n) | |||||||
|---|---|---|---|---|---|---|---|---|
| n | C. s. cyanecula | n | C. s. svecica | n | C. s. pallidogularis | n | C. s. volgae | |
| Длина крыла, мм | 37 | 73.0–79.2 (76.5 ± 1.6) |
126 | 70.5–80.0 (75.3 ± 1.9) | 63 | 68–78 (72.8 ± 1.8) |
110 | 70–77.7 (74.0 ± 1.5) |
| Длина вершины крыла, мм | 15 | 17.0–20.6 (18.8 ± 1) |
119 | 13.2–20.7 (17.6 ± 1.4) | 97 | 12.0–19.0 (15.6 ± 1.2) | 169 | 14–20.1 (16.8 ± 1.2) |
| Длина хвоста, мм | 36 | 52.8–58.8 (56.0 ± 1.6) |
103 | 50–61.6 (55.1 ± 2.3) |
61 | 51.0–60.5 (56.5 ± 2) | 107 | 50.2–61.7 (56.3 ± 2.1) |
| Отношение длины хвоста к длине крыла |
36 | 0.69–0.76 (0.73 ± 0.01) |
103 | 0.65–0.81 (0.73 ± 0.02) | 59 | 0.73–0.83 (0.77 ± 0.02) | 105 | 0.7–0.82 (0.76 ± 0.02) |
| Индекс заостренности крыла | 15 | 0.22–0.26 (0.24 ± 0.01) |
119 | 0.18–0.28 (0.23 ± 0.02) | 62 | 0.17–0.26 (0.2 ± 0.016) | 108 | 0.19–0.26 (0.23 ± 0.01) |
| Длина цевки, мм | 37 | 25.0–27.2 (26.0 ± 0.6) |
161 | 23.7–28.7 (26.6 ± 0.8) | 98 | 22.4–28.3 (25.7 ± 0.9) | 169 | 23.2–28.1 (25.9 ± 0.8) |
| Длина клюва от ноздри, мм | 37 | 7.8–9.4 (8.7 ± 0.4) |
149 | 7.5–9.8 (8.6 ± 0.4) |
60 | 8.0–9.7 (8.8 ± 0.3) |
161 | 7.5–9.8 (8.6 ± 0.4) |
| Длина клюва от рамфотеки, мм | 37 | 11.1–12.8 (12.1 ± 0.4) |
147 | 10.7–13 (11.8 ± 0.5) | 93 | 11.0–13.7 (12.0 ± 0.6) | 157 | 10.3–13.1 (11.8 ± 0.5) |
Статистическую обработку проводили в программе Statistica 8.0. Достоверным считалось отличие при Р < 0.05. Для сравнения каждого типа сигналов между регионами использовали тест Kruskal-Wallis ANOVA. Комплексное сравнение регионов по выбранным признакам провели с помощью дискриминантного анализа.
РЕЗУЛЬТАТЫ
Песню варакушки отличают значительная структурная сложность, большое разнообразие составляющих элементов и способов их компоновки. Отдельного упоминания заслуживают превосходные способности к имитации звуковых сигналов других видов птиц (Симкин, 1990; Мальчевский, 2012; Зиновьев, Зиновьев, 2016; Naguib, Kolb, 1992; Metzmacher, 2008).
Песня варакушки тяготеет к раздельной манере исполнения, т.е. состоит из отдельных (единичных) песен, разделенных четкими паузами, однако при этом характеризуется очень высокой изменчивостью всех параметров: частной модуляции, длительности, ритмической структуры. В то же время в песне, как правило, отчетливо заметны повторяющиеся (стереотипные) вокальные паттерны, многие из которых имеют достаточно сложную структуру и состоят из десятков разнообразных нот, слогов и фраз. В известной мере эти последние можно рассматривать как “типы песен”. Они представляют собой стереотипные вокальные конструкции, повторяющиеся многократно во время пения данного самца. Принимая во внимание огромное разнообразие слогов в индивидуальном репертуаре и практически бесконечное число их возможных комбинаций друг с другом, возможность случайного повторения их длительных последовательностей представляется пренебрежимо малой. Примеры таких повторяющихся стереотипных конструкций приведены на рис. 2.
Рис. 2.
Образцы песен четырех подвидов варакушки: А–Ж – S. c. volgae, З–К – S. c. pallidogularis, Л–М – S. c. cyanecula, Н–П – S. c. svecica. По оси ординат – частота, кГц; по оси абсцисс – время, с.
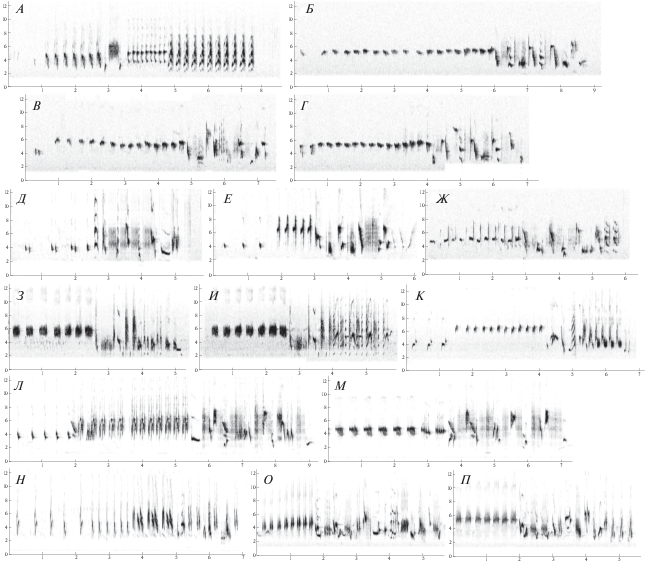
Наряду с высокой изменчивостью, песня варакушки несет и отчетливые видоспецифические признаки. Наиболее заметны среди них длинные серии одинаковых элементов (слогов), которые исполняются обычно в самом начале очередной песни и которые выполняют функцию своеобразных запевок. Запевки, включающие до 20 элементов, типичны для всех четырех изученных нами подвидов. Во многих случаях последовательно исполняются две или три гомотипические серии, состоящие из разных элементов. После запевки исполняется основная часть песни, обычно представляющая собой сложный конгломерат из тоновых, гармонических и шумовых элементов. Часто последовательно несколько раз подряд исполняются одинаковые запевки, однако при этом всякий раз исполняется новая основная часть песни. Эта особенность песни, впервые отмеченная у C. s. сyanecula (Naguib, Kolb, 1992), оказалась столь же характерной и для изученных нами C. s. volgae и C. s. pallidogularis.
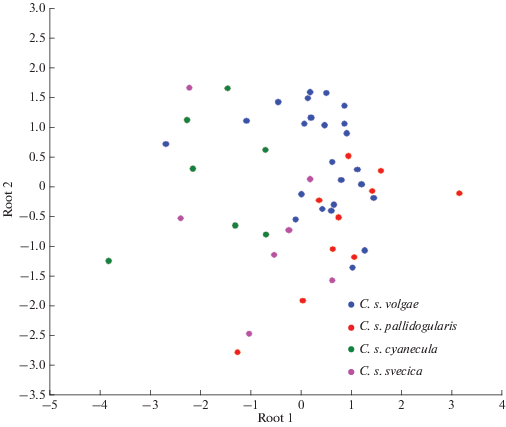
Основные частотно-временны́е параметры песен варакушки для 4-х изученных нами подвидов приведены в табл. 1. Статистический анализ (тест Kruskal-Wallis) выявил достоверные различия (Р < 0.05) между некоторыми подвидами только по четырем из 10 параметров, используемых нами при анализе единичных песен. Примечательно, что по всем четырем признакам C. s. volgae отличается от C. s. cyanecula, но не отличается от C. s. svecica, тогда как между C. s. volgae и C. s. pallidogularis имеется только одно различие (рис. 3). При использовании всех 10 параметров в процедуре дискриминантного анализа получен сходный результат, причем наибольший вклад в разделение дает минимальная частота (р < 0.01; лямбда Уилкса 0.352). Белозвездный C. s. cyanecula достаточно четко отделяется от рыжезвездных C. s. svecica и C. s. pallidogularis, а также от полиморфного C. s. volgae (рис. 4). Что касается изученной нами нижегородской полиморфной популяции C. s. volgae, то статистически достоверных различий между пением белозвездных и рыжезвездных самцов и самцов с промежуточной окраской ни по одному параметру не выявлено.
Рис. 3.
Изменчивость параметров песни четырех подвидов варакушки: А – максимальная частота, Б – минимальная частота, В – пластичность, Г – длительность, Д – число элементов.
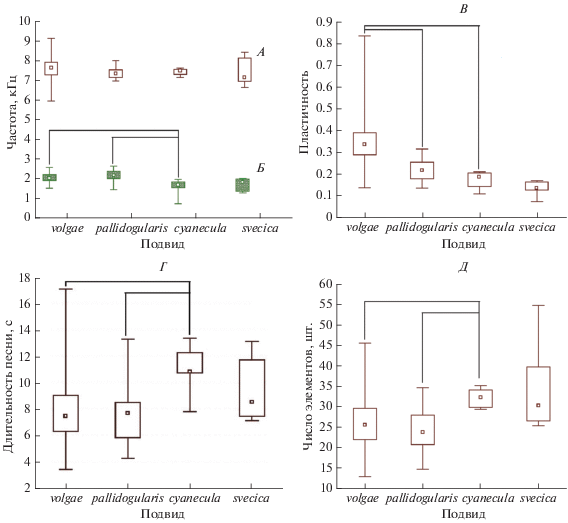
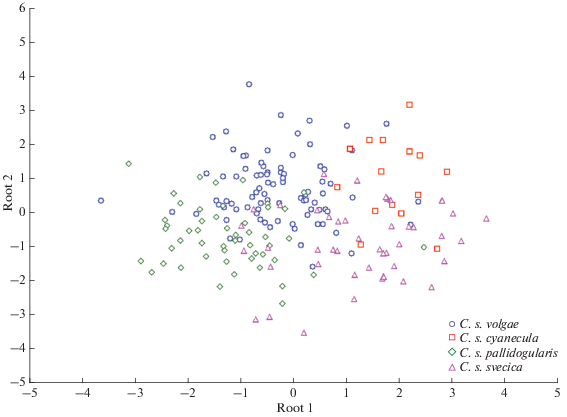
Рис. 4.
Визуализация результатов канонического дискриминантного анализа по параметрам песни (табл. 1) четырех подвидов варакушки.
Для выявления генетических различий между изучаемыми подвидами варакушки и особями с разной окраской нагрудного пятна из полиморфной популяции C. s. volgae мы использовали 24 пробы по цитохрому b и 20 – по контрольному региону. Ни по участку цитохрома b (456 п.н.), ни по контрольному региону (850 п.н.) между C. s. volgae и C. s. cyanecula, а также между ними и C. s. svecica (последовательности из Генбанка) значимых различий не выявлено. Не найдены также генетические различия в полиморфной популяции C. s. volgae в Нижнем Новгороде между особями, различавшимися окраской звезды (рыжезвездными, белозвездными и промежуточными).
Данные о размерах и некоторых размерных пропорциях самцов изученных подвидов приведены в табл. 2. Результаты дискриминатного анализа (рис. 5), выполненного по всем признакам, свидетельствует о достоверном в целом разделении выборок (Wilks’ Lambda: 0.27, F = 11,27, p < 0.001). Первая дискриминантная ось достаточно четко отделяет C. s. pallidogularis от C. s. cyanecula и C. s. svecica, тогда как две последние формы разделены второй осью, хотя и не так полно. Что касается C. s. volgae, то облако точек, соответствующее этой форме, широко перекрывается с прочими формами, что можно рассматривать, как свидетельство весьма значительной морфологической изменчивости и вполне согласуется с распространенными представлениями о гибридной природе этого полиморфного подвида. Дискриминантный анализ набора морфологических признаков самцов из ареала C. s. volgae не показал статистически достоверного разделения белозвездных и рыжезвездных морф (Wilks’ Lambda: 0.86, F = 1,802, p < 0.099).
Рис. 5.
Визуализация результатов канонического дискриминантного анализа по размерным (табл. 2) и окрасочным (интенсивность синего цвета пластрона и величина нагрудного пятна) признакам для взрослых самцов четырех подвидов варакушки.
ОБСУЖДЕНИЕ
Проведенный нами анализ позволяет сделать вывод о весьма незначительном уровне дивергенции полиморфного подвида C. s. volgae по всем изученным признакам. По песне его отличие наблюдается только при сравнении с C. s. cyanecula, по морфологическим признакам C. s. volgae широко перекрывается с тремя сопредельными подвидами, по двум генетическим маркерам, изученным нами, отличий от C. s. svecica и C. s. pallidogularis не выявлено. Таким образом, основной спецификой подвида C. s. volgae остается его ярко выраженный полиморфизм, когда практически во всех популяциях бок о бок гнездятся и белозвездные, и рыжезвездные особи. Подчеркнем, что между двумя этими вариантами мы не выявили значимых различий ни по вокальным, ни по размерным, ни по генетическим характеристикам, вследствие чего форму C. s. volgae можно рассматривать как единый самостоятельный подвид.
Формирование данной расы, вероятнее всего, началось вследствие возникновения зоны вторичного контакта европейской белозвездной расы C. s. cyanecula, прежде довольно широко распространенной к востоку в пойменных местообитаниях преимущественно по берегам водоемов, и северной рыжезвездной формы C. s. svecica, обитавшей на севере Европы строго в тундровой и лесотундровой зоне. Гнездовой ареал cyanecula в прошлом, по-видимому, простирался к востоку значительно шире, по крайней мере, до бассейна Дона, верхнего и, возможно, среднего течения Волги. Лесные пространства таежной зоны исторически служили препятствием, изолирующим эту расу от распространенного на севере номинативного подвида. Область распространения svecica была приурочена к тундроподобным ландшафтам севера, а к югу в пределы таежной зоны эта форма проникала, по-видимому, диффузно, по пространствам обширных верховых болот. Освоение и трансформация лесной зоны к востоку от Балтийского моря человеком, приведшие к возникновению на месте прежде непрерывных лесных массивов мозаичного ландшафта, а также запруживание рек с образованием большого количества новых искусственных водоемов, создали условия для проникновения белозвездных варакушек (C. s. cyanecula) к северу и возникновения зоны вторичного контакта с номинативным подвидом. Дальнейшим следствием гибридизации этих форм стало широкое распространение генов номинативного подвида в пределы прежде мономорфных белозвездных популяций, приведшее к возникновению современного полиморфизма окрасочных признаков. “Модифицированная” таким образом форма, вероятно, обладала уже значительно большей экологической пластичностью по сравнению с родительскими подвидами, что в сочетании с продолжающимся хозяйственным освоением исходных ландшафтов европейской части России создало условия для ее широкого распространения во всех подходящих местообитаниях. Вторично на юго-востоке европейской части обсуждаемая форма вступила в контакт и образовала зону интерградации со степной рыжезвездной расой C. s. pallidogularis, гнездовой ареал которой, вероятно, простирался к северо-западу до долин Волги и нижнего течения Камы. Геном C. s. pallidogularis также влился в общий пул полиморфных популяций, дополнив фенотипическую картину некоторыми окрасочными чертами этой формы, однако фенотип рыжезвездной степной формы по большей части сохранил свои исходные географические позиции. Пределы расселения варакушек со смешанными признаками на востоке достигли западных предгорий Урала, а на юге – побережья Азовского моря и Предкавказья, что соответствует современным пределам распространения С. s. volgae (рис. 1). Здесь же следует отметить, что область зимовок C. s. volgae охватывает и северо-восточную Африку (совпадая с областью, занимаемой C. s. cyanecula) и юго-западную Азию (где помимо нее зимует и C. s. pallidogularis) (Dickinson, 2003).
Приведенная гипотеза возникновения современной формы C. s. volgae в результате трансформации исходных местообитаний хозяйственной деятельностью человека хорошо согласуется с данными, обосновывающими возникновение на севере и востоке Европы гибридогенного полиморфизма популяций желтой трясогузки (Motacilla flava) и серого сорокопута (Lanius excubitor), вызванного теми же историческими причинами (Тайкова, Редькин, 2014; Редькин и др., 2015).
Список литературы
Белоконь М.М., Белоконь Ю.С., Банникова А.А., Политов Д.В., 2014. Молекулярно-генетические методы в зоологической практике на Звенигородской биостанции МГУ. Город: Изд-во ООО “Цифровичок”. 68 с.
Зиновьев В.И., Зиновьев А.В., 2016. Некоторые материалы по биологии варакушки в Калининской области // Русский орнитологический журнал. Т. 25. Экспресс-выпуск 1299. С. 2199–2201.
Иваницкий В.В., Марова И.М., Антипов В.А., 2013. Принципы построения и особенности дифференциации песни в московской популяции восточного соловья (Luscinia luscinia, Turdidae) // Зоологический журнал. Т. 92. № 2. С. 206–220.
Козлова Е. В., 1945. Индивидуальная изменчивость варакушки [Cyanecula svecica (L.)] и вероятные пути ее эволюции // Зоологический журнал. Т. 24. № 5. С. 299–307.
Мальчевский А.С., 2012. К вопросу о голосовой имитации птиц // Русский орнитологический журнал. Т. 21. Экспресс-выпуск 799. С. 2353–2360.
Матюхин А.В., 2000. Фенотипический состав полиморфных популяций варакушки Luscinia svecica Linnaeus, 1758 Восточной Европы и возможные пути его становления // Теоретические проблемы экологии и эволюции (Третьи Любищевские чтения). Тольятти: ИЭВБ РАН. 222 с.
Мацына Е.Л., 2006. Территориальный консерватизм и филопатрия в полиморфной популяции варакушки подвида Luscinia svecica volgae (Zarudny, 1897) // Бутурлинский сборник. Материалы II Всерос. научно-практической конференции, посвященной памяти С.А. Бутурлина. С. 220–229.
Панов Е.Н., Опаев А.С., 2011. Акустическое поведение птиц: структура, функция, эволюция (на примере избранных семейств отряда Воробьинообразных). Электронный журнал “Этология и зоопсихология”. № 1(3). http://www. etholpsy.ru
Портенко Л.А., 1954. Отряд воробьиные. Птицы СССР. Ч. 3. М.-Л.: Изд-во АН СССР. 256 с.
Редькин Я.А., Архипов В.Ю., Волков В.Ю., Мосалов А.А., Коблик Е.А., 2015. Вид или не вид? Спорные таксономические трактовки птиц Северной Евразии // XIV Международная орнитологическая конференция Северной Евразии (Алматы, 18–24 августа 2015 г.). Т. 2. Доклады. С. 104–138.
Симкин Г.Н., 1990. Певчие птицы. Справочное пособие. М.: Лесная промышленность. 399 с.
Степанян Л.С., 1990. Конспект орнитологической фауны СССР. М.: Наука. 726 с.
Сушкин П.П., 1925. Список и распределение птиц Русского Алтая и ближайших частей северо-западной Монголии с описанием новых и малоизвестных форм. Л.: Изд-во АН СССР. 79 с.
Тайкова С.Ю., Редькин Я.А., 2014. О формах группы серых сорокопутов в Европейской России и Украине в свете современных взглядов на систематику этого комплекса // Редкие виды птиц Нечерноземного центра России. Материалы V совещания “Распространение и экология редких видов птиц Нечернозёмного центра России”. Москва. С. 223–240.
Dickinson E.C. (Ed.), 2003. The Howard and Moore complete checklist of the birds of the world. Princeton University Press. 1056 p.
Johnsen A., Andersson S., Fernandez J.G., Kempenaers B., Pavel V., Questiau S., Raess V., Rindal E., Lifjeld J.T., 2006. Molecular and phenotypic divergence in the bluethroat (Luscinia svecica) subspecies complex // Journal of Molecular Ecology. V. 15. P. 4033–4047.
Metzmacher M., 2008. Les Grillons, muses de la Gorgebleue à miroir blanc Luscinia svecica cyanecula? // Parcs et Réserves. V. 63. № 2. P. 17–19.
Naguib M., Kolb H., 1992. Vergleich des Strophenaufbaus und der Strophenabfolge an Gesängen von Sprosser (Luscinia luscinia) und Blaukehlchen (Luscinia svecica) // Journal für Ornithologie. Bd. 133. S. 133–145.
Pavel V., Chutný B., 2013. Slavík modráček (Luscinia svecica) – komplex poddruhů v rané fázi speciace // Opera Corcontica. V. 50. P. 9–26.
Petrilák L., 2010. Poddruhová determinace samic slavíka modráčka (Luscinia svecica) // Ms. (bakalářská práce, Přírodovědecká fakulta, Univerzita Palackého, Olomouc). Univerzita Palackého v Olomouci Přírodovědecká fakulta Katedra zoologie a ornitologická laboratoř. 34 p.
Petrilák L., 2014. Subspecies differences in the Bluethroat (Luscinia svecica) // Ms. (Diploma, Přírodovědecká fakulta, Univerzita Palackého, Olomouc). Univerzita Palackého v Olomouci Přírodovědecká fakulta Katedra zoologie a ornitologická laboratoř. 40 p.
Questiau S., Eybert M.C., Gaginskaya A.R., Gielly L., Taberlet P., 1998. Recent divergence between two morphologically differentiated subspecies of bluethroat (Aves: Muscicapidae: Luscinia svecica) inferred from mitochondrial DNA sequence variation // Journal of Molecular Ecology. V. 7. P. 239–245.
Smithe B.F., 1975. Naturalist’s color guide. American Museum of Natural History. New York. 366 p.
Tugarinov A., 1929. The bluethroats of Eastern Palerctic // Annuaire du Muséé Zoologic de l`Academie des Sciene de l`URRS XXIX 1928 (1929). P. 1–14.
Turčoková L., Osiejuk T.S., Pavel V., Glapan J., Petrusková T., 2010. Song divergence of two bluethroat subspecies (Luscinia s. svecica and cyanecula) // Ornis Fennica. V. 87. P. 168–179.
Zink R.M., Drovetski S.V., Questau S., Fadeev I.V., Nesterov E.V., Westberg M.C., Rohwer S., 2003. Recent evolutionary history of the bluethroat (Luscinia svecica) across Eurasia // Journal of Molecular Ecology. V. 12. P. 3069–3075.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал