

Зоологический журнал, 2021, T. 100, № 8, стр. 885-896
Новые данные о распространении видов рода Bittacus (Mecoptera, Bittacidae) в европейской части России и Казахстане и замечания об их диагностике
В. Ю. Савицкий a, *, А. В. Тимохов b, **
a Зоологический музей Московского государственного университета
имени М.В. Ломоносова
125009 Москва, Россия
b Биологический факультет Московского государственного университета
имени М.В. Ломоносова
119234 Москва, Россия
* E-mail: alophus@gmail.com
** E-mail: atimokhov@mail.ru
Поступила в редакцию 29.06.2020
После доработки 23.11.2020
Принята к публикации 30.11.2020
Аннотация
Обобщены и уточнены известные сведения о распространении, биотопической приуроченности и фенологии видов рода Bittacus Latreille 1805 в европейской части России и прилегающих регионах. Bittacus italicus (Müller 1766) впервые указан для Волгоградской области и Казахстана. Проанализированы диагностические признаки B. italicus и B. hageni Brauer 1860. Предложены новые признаки жилкования крыльев для определения обоих полов этих видов.
Род Bittacus Latreille 1805 включает более 110 видов, распространенных в Евразии, Африке, Северной и Южной Америке (Penny, Byers, 1979; Плутенко, 1995). В Европе, в том числе и на юге европейской части России, встречаются 2 вида этого рода: Bittacus italicus (Müller 1766) и B. hageni Brauer 1860 (Esben-Petersen, 1921; Мартынова, 1959; Дорохова, Мартынова, 1987; Tillier, 2008; Tillier et al., 2009; Щуров, Макаркин, 2017). Еще 3 вида рода Bittacus известны на Дальнем Востоке России: B. sinensis Walker 1853, B. cirratus Tjeder 1956 и B. ussuriensis Plutenko 1985 (Плутенко, 1995). Самцы B. italicus и B. hageni хорошо различаются строением вершинных сегментов брюшка и гениталий (Esben-Petersen, 1921; Мартынова, 1959; Petschenka, 2006). Самки этих видов очень сходны и их точное определение затруднительно (Przybyłowicz, 2006; Щуров, Макаркин, 2017). Для их различения используют главным образом некоторые признаки жилкования крыльев, которые могут быть подвержены довольно сильной изменчивости (Devetak, 1991; Przybyłowicz, 2006).
В настоящей работе обобщены сведения о распространении видов рода Bittacus в европейской части России и прилегающих регионах, проанализированы диагностические признаки B. italicus и B. hageni, предложены новые признаки жилкования крыльев для определения обоих полов этих видов.
МАТЕРИАЛ И МЕТОДИКА
Материалом для настоящей работы послужили сборы авторов и коллекция Зоологического музея Московского государственного университета (ЗММУ). Также авторы имели возможность ознакомиться с географическими этикетками отдельных экземпляров Bittacus italicus и B. hageni из коллекции Зоологического института РАН (Санкт-Петербург, ЗИН), определенных О.М. Мартыновой.
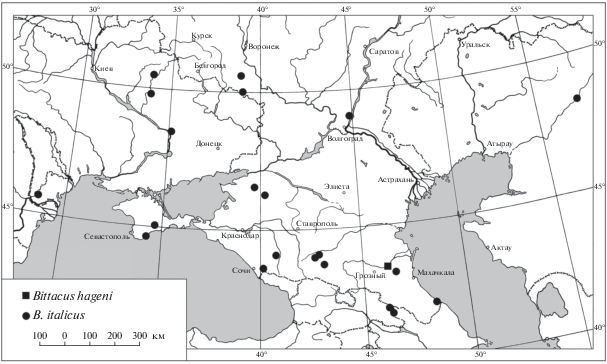
Карта распространения видов рода Bittacus в европейской части России и на сопредельных территориях (рис. 1) составлена на основании изученных материалов, литературных данных (Мартынова, 1959; Щуров, Макаркин, 2017) и информации, полученной от А.В. Присного.
Рис. 1.
Распространение видов рода Bittacus в европейской части России и на сопредельных территориях.
Фотографии деталей строения B. italicus (рис. 2; 3, 10–13; 4, 1–5, 8) сделаны с изученных нами экземпляров. Фотографии дистальной части брюшка самца B. italicus сделаны с экземпляра, хранящегося в спирту. Фотографии и рисунки деталей строения B. hageni (рис. 3, 1–9; 4, 6, 7) заимствованы нами с изменениями из работ других авторов (Brauer, 1860; Мартынова, 1959; Devetak, 1991; Petschenka, 2006; Przybyłowicz, 2006; Tillier, 2008; Tourneur, 2019).
Рис. 2.
Крылья самок (1–5, 9, 10) и самцов (6–8) Bittacus italicus: 1 – балка Песковатка; 2, 6, 7 – дельта р. Самур; 3 – Красная Поляна; 4 – Днепр; 5 – Болград; 8 – Ессентуки; 9, 10 – долина р. Мзымта. А1 – 1-я анальная жилка, А2 – 2-я анальная жилка, Cu2 – 2-я кубитальная жилка, М4 – 4-я ветвь медиальной жилки, Rs – радиальный сектор, Sc – субкостальная жилка, m – поперечная жилка 1-го медиального развилка, pt – птеростигма, rm – радиомедиальная жилка, rs1 – 1-я ячейка радиального сектора.
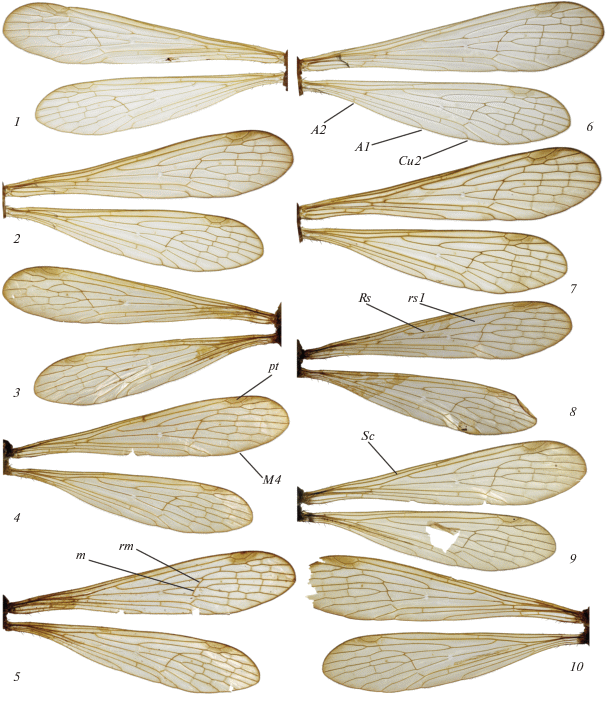
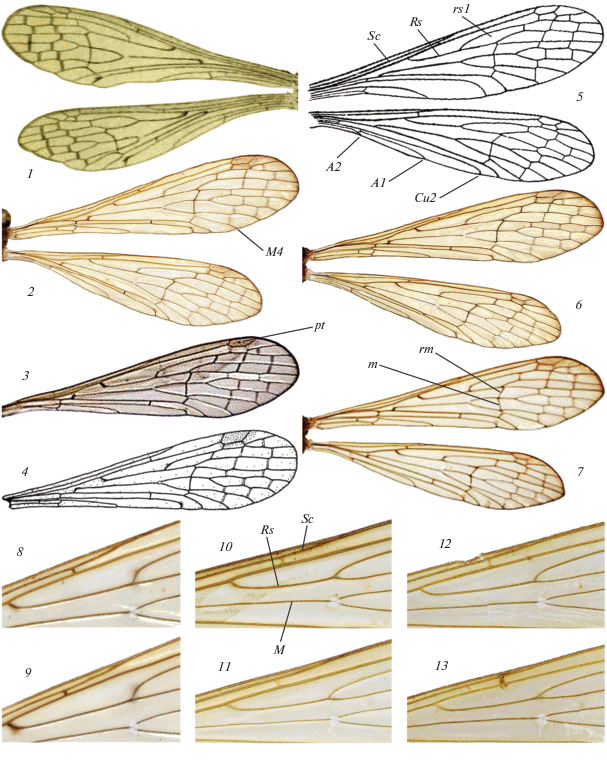
Рис. 3.
Детали строения видов рода Bittacus: 1–9 – B. hageni (по работам разных авторов с изменениями); 10–13 – B. italicus. 1–7 – крылья самок (1–4) и самцов (5–7); 8–13 – участок переднего крыла. 1 – Австрия (по: Brauer, 1860); 2, 6, 8, 9 – Польша (по: Przybyłowicz, 2006); 3 – Франция (по: Tourneur, 2019); 4 – Словения (по: Devetak, 1991); 5 –Чеченская Республика (по: Мартынова, 1959); 7 – Франция (по: Tillier, 2008); 10, 11 – дельта реки Самур; 12 – долина р. Мзымта; 13 – Днепр. М – медиальная жилка; остальные обозначения как на рис. 2.
Для описания структур последних сегментов брюшка и гениталий самца использована терминология, предложенная Тьедером (Tjeder, 1956; 1970).
РЕЗУЛЬТАТЫ
Bittacus hageni Brauer 1860
Распространение. Широко распространен в Южной и Центральной Европе (Vidlička, Kmeťová, 2002; Tillier, 2008; Tillier et al., 2009; Dobosz, Pacuk, 2018).
Для территории России B. hageni достоверно указан только Мартыновой (1959) по единственному самцу. В сухой коллекции ЗИН этот экземпляр отсутствует, в коробке имеется только булавка с этикетками: “Парабоч Кизлярского окр., Терской обл., 16.VIII.1927 Кириченко”, “экземпляр в спирту” и “Bittacus hageni Br. Alexandrova-Martynova det.”. Точность его определения сомнений не вызывает, так как в работе О.М. Мартыновой приведены оригинальные рисунки деталей строения самца B. hageni. В настоящее время Парабоч – это поселок в Шелковском р-не Чеченской Республики, расположенный примерно в 50 км СВВ от города Грозный на левом берегу р. Терек.
Bittacus hageni указан также для Алексеевского и Ровеньского районов Белгородской обл. без точных мест сбора (Присный, 2005). А.В. Присный любезно предоставил нам фотографии экземпляра Bittacus из Алексеевского р-на. Их изучение показало, что этот экземпляр на самом деле относится к B. italicus.
Экология. B. hageni всюду редок, однако известны случаи серийных сборов этого вида (Brauer, 1860, 1871; Vidlička, 2010). Согласно литературным данным это – летний вид, в Западной и Центральной Европе подавляющее большинство экземпляров собрано в июле–августе, самые ранние и самые поздние сборы относятся к середине июня и сентября, соответственно (Tillier, 2008; Tillier et al., 2009; Vidlička, 2010). B. hageni предпочитает затененные густые заросли, влажный подлесок, встречается по берегам рек (Brauer, 1871; Tillier et al., 2009), наиболее массовые сборы были сделаны в дубово-вязовом лесу с помощью ловушки Малеза (Vidlička, 2010). Отметим, что в окрестностях поселка Паробоч в долине р. Терек имеются довольно крупные массивы влажных пойменных дубрав и пойменных берестовых дубрав (Бахтиев, 2011).
Bittacus italicus (Müller 1766)
Материал. Россия: 1 ♀, Волгоградская обл., СВ г. Волгоград, ~9 км СЗ пос. Дубовка, балка Песковатка, 49°07′43″ N, 44°45′27″ E, ~75 м, 14.07.2016 (В.Ю. Савицкий, А.В. Тимохов); 1 ♀, Краснодарский край, пос. Красная Поляна, 27.07.1952 (А.Н. Желоховцев); 1 ♀, окр. пос. Красная Поляна, долина реки Мзымта, 03.08.1951 (А.Н. Желоховцев); 1 ♂, Ставропольский край, г. Ессентуки, 31.07.1919 (М.А. Рябов); 2 ♂♂, 1 ♀, Дагестан, дельта реки Самур близ берега Каспийского моря, ~7 км СВ пос. Самур, 41°52′46″ N, 48°32′33″ E, –25 м, 11.07.2018 (А.В. Тимохов, В.Ю. Савицкий). Украина: 3 ♀♀, Одесская обл., г. Болград, 4–10.08.1953; 1 ♀, Днепропетровская обл., г. Днепр, р-н Игрень, 20.07.1992 (В.Н. Ковтунович); 1 ♀, Полтавская обл., Миргородский р-н, с. Яреськи, 29.07.1919 (И. Фабри); 1 ♂, там же, Гадячский р-н, г. Гадяч, 20–22.07.1911 (Н.М. Книпович).
Распространение. Широко распространен в Южной и Центральной Европе (Dobosz, Hadas, 1999; Tillier et al., 2009).
Мартынова (1959) по материалам коллекции ЗИН указала B. italicus для Крыма и Кавказа без приведения точных мест сбора. В коллекции ЗИН имеются экземпляры этого вида из следующих пунктов с территории России и стран Закавказья: Крым (Артек и Белогорск); Ставропольский край (Ессентуки и Кисловодск); Кабардино-Балкария (Гунделен); Дагестан (Хасавюрт); Грузия (Лагодехи); Азербайджан (село Кабахчол Белоканского района). Для Белгородской обл. B. italicus был указан как B. hageni (Присный, 2005). Согласно устному сообщению А.В. Присного, им были собраны 2 экземпляра B. italicus в следующих пунктах Белгородской обл.: 1 ♀, Алексеевский р-н, 2 км СВ с. Варваровка, 18.08.2004; 1 экз., Ровеньский р-н, западнее пос. Ровеньки, Ровеньский яр, 11–20.08.2001. В Краснодарском крае B. italicus найден в долинах рек Ея, Малая Лаба и Мзымта (Щуров, Макаркин, 2017; коллекция ЗММУ).
Мартынова (1959) ошибочно указала B. italicus для Урала. Это указание основано на 2 экземплярах из коллекции ЗИН, имеющих этикетку: “Ур. Кужа-Тугай Уральск. обл. 15.VI.908 Бородин”. В статье Мартыновой (1959) название этого урочища приведено с ошибками как “Кажи-Тугай (sic!) (Урал)”. Д.Н. Бородин совместно с Б.П. Уваровым и В.М. Шицем летом 1908 г. собирал насекомых в Темирском уезде Уральской обл. (Уваров, Бородин, 1909; Уваров, 1910; 1910а). Согласно Уварову (1910а) урочище Кужа-Тугай находится на р. Эмба, 110 верст (около 117 км) южнее города Темир, у южной оконечности Кок-Джидинских бугристых песков (пески Кокжиде). На доступных картах топоним с таким названием нам найти не удалось, однако непосредственно у южной оконечности песков Кокжиде находится село Кожасай (103 км южнее города Темир, 48°12′52″ N, 57°06′52″ E), для которого в электронных поисковых источниках (https://mapcarta.com/13564062) приводится альтернативное название Kodzha-Tugay. Соответственно данной Б.П. Уваровым привязке урочище Кужа-Тугай, вероятнее всего, находится вблизи села Кожасай Мугалжарского р-на Актюбинской обл. Казахстана.
Таким образом, на основании сборов более вековой давности мы впервые указываем B. italicus для фауны Казахстана. В России этот вид достоверно известен из Крыма, Белгородской и Волгоградской областей, Краснодарского и Ставропольского краев, Кабардино-Балкарии, Чечни и Дагестана. Находки B. italicus в низовьях рек Самур и Терек, окрестностях города Волгоград и в Западном Казахстане, в долине р. Эмба позволяют предположить, что он распространен по всему Северному Прикаспию.
Экология. Вид всюду довольно редок, однако встречается чаще, чем B. hageni. Описан случай массового появления B. italicus в центральной Франции в 1888 г. (Martin, 1892, цит. по: Tillier et al., 2009). Все экземпляры B. italicus, собранные нами в Волгоградской обл. и Дагестане, прилетели ночью на свет ультрафиолетовой лампы. Сравнение дат сборов показывает, что основной период лёта B. italicus в различных точках широкого ареала приходится на июль – первую половину августа (Brauer, 1871; Мартынова, 1959; Щуров, Макаркин, 2017 и др.). Самое раннее появление особей отмечено 28 июня (по григорианскому календарю) в долине р. Эмба в Западном Казахстане и 30 июня в степных районах Краснодарского края.
Брауэр (Brauer, 1871) в результате многолетних наблюдений и экспериментов установил, что этому виду для успешного эмбрионального развития необходимо полное высыхание почвы, в которую были отложены яйца. В противном случае эмбриогенез может продолжаться еще один год. При этом фенология вида определяется не только суммой эффективных температур, ключевым фактором оказывается режим увлажнения почвы в течение года. Обильное увлажнение почвы весной стимулирует ранний выход личинок из яиц и их дальнейшее развитие. Напротив, засушливая весна и последующее обильное увлажнение почвы летом обусловливают более поздний выход личинок и, соответственно, задержку появления имаго на 3–4 месяца по сравнению с обычными годами. Так, в 1856 г. Брауэр (Brauer, 1871) зарегистрировал наиболее позднее появление имаго B. italicus (самца – 25 августа и самки – 10 октября). Эта экологическая особенность, по-видимому, и определяет характер биотопической приуроченности B. italicus, а также встречаемость и распределение его локальных популяций.
Bittacus italicus имеет широкий ареал и распространен как в гумидных, так и в аридных районах. Он приурочен к лесным биотопам в меньшей степени, чем B. hageni, и также встречается на опушках и в открытых биотопах (Brauer, 1871; Tillier et al., 2009; Nidergas, 2014). В аридных районах этот вид встречается по берегам рек и других водоемов, а также в оврагах и балках при наличии в них врéменных водотоков, что обеспечивает необходимое увлажнение почвы в весенний период. Экземпляры из Волгоградской (рис. 5), Белгородской областей и Краснодарского края, для которых имеются соответствующие сведения, были собраны в балках или оврагах (Щуров, Макаркин, 2017; Присный, устное сообщение; наши данные). В Дагестане все особи B. italicus пойманы нами в дельте р. Самур, близ берега Каспийского моря, на участке с разнотравной растительностью и слегка засоленной почвой (рис. 6).
Рис. 4.
Детали строения видов рода Bittacus: 1–5, 8 – B. italicus (Самур); 6, 7 – B. hageni (Франция, по: Petschenka, 2006, с изменениями). 1, 2 – дистальная часть брюшка с вывернутыми пузыревидными образованиями сбоку и сверху; 3 – дистальная часть брюшка сбоку; 4, 5 – вершина брюшка сбоку и сверху; 6, 7 – часть брюшка с вывернутыми пузыревидными образованиями сбоку и сверху; 8 – основание крыла. VI–VIII – 6–8-й тергиты брюшка; пф – пенисфилум; ц – церк; эп – эпиандриум.

Рис. 5.
Местообитание Bittacus italicus в Волгоградской области: 1 – вид сверху на балку Песковатка, 2 – место сбора B. italicus в балке Песковатка.


Особый интерес представляет самая восточная известная точка ареала B. italicus – урочище Кужа-Тугай в долине р. Эмба. Уваров и Бородин (1909) отмечают полное отсутствие здесь и в близлежащих биотопах древесной и кустарниковой растительности. В урочище Кужа-Тугай имеются сырые и болотистые луга, заметно отличающиеся как от окружающих степных и полупустынных биотопов, так и от сухих и солонцеватых лугов долины Эмбы (Уваров, Бородин, 1909; Уваров, 1910а). Благодаря этому здесь, по-видимому, создается градиент экологических условий, пригодных для развития преимагинальных стадий B. italicus.
Диагностика Bittacus hageni и B. italicus
Самцы B. hageni и B. italicus хорошо различаются строением последних сегментов брюшка и эдеагуса, что известно, начиная с работы Брауэра (Brauer, 1860; Esben-Petersen, 1921; Мартынова, 1959; Petschenka, 2006). У B. italicus пенисфилум эдеагуса простой, дуговидно изогнут, доли эпиандриума (видоизмененного 9-го тергита) относительно узкие, на вершине немного расширены и слегка загнуты внутрь, на внутренней стороне примерно у середины с узким отростком, направленным вниз и внутрь, вершина которого с рядом из нескольких плотно расположенных крупных шипиков (рис. 4, 4, 5). У B. hageni пенисфилум эдеагуса спирально закручен, доли эпиандриума широкие, к вершине заметно сужены и сильно загнуты внутрь, на внутренней стороне примерно у середины с широким полусферическим выступом, который покрыт многочисленными шипиками (согласно рисункам в работах: Esben-Petersen, 1921; Мартынова, 1959; Petschenka, 2006; Przybyłowicz, 2006).
Брауэр (Brauer, 1855, 1871) первым описал своеобразные пузыревидные образования сочленовных мембран 6–7-го и 7–8-го тергитов самцов Bittacus, а также отметил их диагностическое значение. У B. italicus они имеют вид непарных поперечных мешковидных образований (рис. 4, 1, 2; Petschenka, 2006: fig. 2, c, d), а у B. hageni – парных пальцевидных структур (рис. 4, 6, 7; Petschenka, 2006: fig. 2, а, b). У B. hageni, как и у B. italicus, эти образования имеются и на сочленовной мембране 7–8-го тергитов, хотя и не показаны на рисунках (Petschenka, 2006).
К настоящему времени подобные образования сочленовных мембран 6–7-го и 7–8-го тергитов описаны для самцов многих видов из разных родов семейства Bittacidae и установлено их функциональное значение (Tjeder, 1956; Bornemissza, 1966; Tan, Hua, 2009; Gao, Hua, 2013 и др.). В гиподерме этих образований находятся феромонные железы (Crossley, Waterhouse, 1969), и самец выворачивает их, привлекая самку (Bornemissza, 1966; Gwynne, 1984; Gao, Hua, 2013).
Ниже рассмотрены признаки, предложенные разными авторами для различения обоих полов B. hageni и B. italicus.
Брауэр (Brauer, 1860) указал значительное число признаков, отличающих самцов и самок B. hageni от B. italicus. Наиболее важные из них, по нашему мнению: у B. hageni более тонкие и короткие ноги, бедра на нижней стороне с 2 рядами щетинок, голени с 3 рядами щетинок; дистальная часть брюшка расширена, 2–7-й тергиты перед задним краем с черными поперечными линиями; крылья относительно шире и короче, на вершине притупленные, поперечная жилка первого медиального развилка (m) перпендикулярна продольной оси крыла и лежит под тупым углом к расположенной непосредственно перед ней прямой радиомедиальной жилке (rm). У B. italicus согласно Ф. Брауэру бедра и голени с бόльшим числом щетинок, образующих по 4 ряда; брюшко почти цилиндрическое, лишь в виде исключения с черными поперечными линиями перед задним краем тергитов; крылья более узкие и длинные, поперечная жилка m наклонена к продольной оси крыла и лежит почти в одном направлении со слегка S-образно изогнутой rm. Также Брауэр (Brauer, 1860) отметил различия в строении яиц этих видов: у B. hageni они серо-желтые, почти сферические, с двумя маленькими углублениями на противоположных концах, а у B. italicus – бочкообразные, темно-коричневые.
Согласно Эсбен-Петерсену (Esben-Petersen, 1921) оба пола B. hageni отличаются от B. italicus более широкими крыльями с более округленной вершиной, более крупным белым пятном в развилке медиальной жилки (М), более короткой 1-й анальной жилкой (А1), отсутствием крупных щетинок на тазиках и наличием крупных щетинок на крае крыла лишь у самого его основания. У B. italicus тазики с несколькими крупными черными щетинками, а край крыла несет крупные черные щетинки на значительном расстоянии от основания крыла.
Дорохова и Мартынова (1987) для различения обоих полов B. italicus и B. hageni приводят лишь признаки жилкования крыльев. По мнению этих авторов, у B. italicus субкостальная жилка (Sc) впадает в костальную (С) перед первым ветвлением радиального сектора (Rs), а А1 оканчивается посередине длины крыла; у B. hageni Sc впадает в С на уровне первого ветвления Rs, а А1 оканчивается проксимальнее середины длины крыла.
У изученных нами экземпляров B. italicus торчащие черные щетинки на бедрах и голенях образуют по 4 ряда, передние тазики с 2–5, средние с 1, задние с 1–3 черными щетинками. Мы не имели возможности изучить эти признаки у B. hageni, но в сравнительном плане изучили их у 2 самцов и 1 самки B. ussuriensis из Южного Приморья. У B. ussuriensis тазики без щетинок, все бедра с 2 рядами щетинок на нижней стороне, передние голени с 3 рядами щетинок, задние лишь с 1–4 щетинками, не образующими явственных рядов, а средние с 3 рядами обычных длинных щетинок и в проксимальной части с рядом коротких сильно утолщенных щетинок. Таким образом, особенности хетотаксии ног у видов рода Bittacus могут значительно различаться. Поэтому непринятые во внимание последующими авторами данные Брауэра (Brauer, 1860) и Эсбен-Петерсена (Esben-Petersen, 1921) о диагностическом значении хетотаксии ног B. hageni и B. italicus заслуживают дополнительного изучения как у этих видов, так и у других видов семейства Bittacidae.
У всех изученных нами экземпляров B. italicus и на рисунке этого вида в работе Брауэра (Brauer, 1860) передний и задний край крыла на некотором протяжении от его основания несет ряд крупных черных щетинок, которые обычно длиннее на заднем крае крыла (рис. 4, 8). На фотографиях крыльев B. hageni и на рисунке в работе Ф. Брауэра такие щетинки незаметны или имеется 1 щетинка в основании переднего края заднего крыла. Поскольку кроме Эсбен-Петерсена (Esben-Petersen, 1921) никто больше не упоминал этот признак, для точного определения его диагностического значения необходимо изучение коллекционных экземпляров B. hageni.
Некоторые изученные нами экземпляры B. italicus имеют отчетливые узкие черные поперечные полосы вдоль переднего края одного или нескольких тергитов. Особенно хорошо они заметны у экземпляров, хранящихся в спирту: у самки из Волгоградской обл. они имеются на 3–5-м, а у самца из Дагестана на 3–6-м тергитах (рис. 4, 1–3: видна черная полоса на 6-м тергите). В обычном состоянии брюшка передний край каждого тергита втянут под задний край предыдущего. Поэтому, если черная полоса просвечивает через задний край предыдущего тергита, создается ложное впечатление, особенно при изучении сухих экземпляров, что она лежит не у переднего края 3–6-го тергитов, а у заднего края 2–5-го тергитов. Таким образом, остается неясным, где именно на тергитах расположены черные поперечные линии, описанные Брауэром (Brauer, 1860), и, соответственно, диагностическое значение этого признака.
Видимо, с этими черными линиями ассоциированы структуры последних тергитов брюшка B. hageni, называемые некоторыми авторами antecostae (Przybyłowicz, 2006; Tillier, 2008; Tourneur, 2019). По мнению этих авторов, наличие antecostae на последних тергитах брюшка позволяет легко различать самок B. hageni и B. italicus. Однако antecostae хорошо различимы только у живых особей, недавно умерщвленных или хранящихся в фиксирующей жидкости, а у сухих экземпляров их наличие или отсутствие оценить сложно (Tillier, 2008; Tourneur, 2019). Это, по нашему мнению, значительно ограничивает диагностическое значение этого признака.
Необходимо отметить, что у изученных нами экземпляров B. italicus черные полосы у переднего края тергитов брюшка по положению соответствуют antecostae в прямом понимании этого термина. Как известно, antecostae образовались на месте первичных межсегментных складок и в виде более или менее явственной бороздки отделяют акротергит от основной части тергита (Snodgrass, 1935). Однако в литературе по таксономии семейства Bittacidae сложилось не вполне правильное использование этого термина. Многие авторы понимают под antecostae только довольно сильно склеротизованные структуры на переднем крае тергита брюшка (Londt, 1972; Tjeder, 1974; Tan, Hua, 2009 и другие). Судя по приведенным в этих работах рисункам, такие структуры часто развиты и на акротергите или в основном на акротертиге. Вместе с тем каждый тергит брюшка имеет и antecostae, и акротергит независимо от степени склеротизации его переднего края. Поэтому морфологически верно писать не о наличии или отсутствии antecostae на тех или иных тергитах, а о степени и характере склеротизации их переднего края.
Крылья B. italicus относительно длиннее и ýже, чем у B. hageni. У B. italicus длина переднего крыла составляет 18–21 мм, длина заднего крыла – 16–19 мм (Esben-Petersen, 1921; наши данные), у B. hageni – 17–19 мм и 15–16.5 мм соответственно (Esben-Petersen, 1921). Согласно нашим промерам у B. italicus длина переднего крыла в 4.6–5.3 раза, а у B. hageni в 3.9–4.4 раза больше ширины, измеренной на уровне проксимального края птеростигмы и места окончания 4-й ветви медиальной жилки (М4).
Белое пятно на крыльях в развилке М может быть одинаково хорошо развито как у B. italicus, так и у B. hageni (рис. 3, 8–13), поэтому этот признак не пригоден для различения этих видов.
У всех изученных нами экземпляров B. italicus, как и указывает Брауэр (Brauer, 1860), поперечная жилка m наклонена к продольной оси крыла (рис. 2). Обычно у B. italicus она лежит почти в одном направлении с жилкой rm, но у некоторых особей (рис. 2, 2, 4, 6) она лежит под тупым углом к rm, как и у B. hageni (рис. 3, 1–7). Жилка rm у B. italicus обычно слабо S-образно изогнута, но иногда почти прямая (рис. 2, 4), как и у большинства B. hageni. В свою очередь, и у B. hageni жилка rm иногда слабо S-образно изогнута (рис. 3, 3). Таким образом, из признаков жилкования крыла, предложенных Ф. Брауэром, для надежного различения B. italicus и B. hageni можно использовать только положение жилки m относительно продольной оси крыла.
Положение вершины Sc относительно 1-го развилка Rs как у B. italicus, так и у B. hageni довольно изменчиво. У обоих видов вершина Sc переднего крыла часто лежит на уровне 1-го развилка Rs или немного дистальнее его (рис. 3, 8–13), а Sc заднего крыла обычно впадает в С заметно проксимальнее 1-го развилка Rs. Таким образом, этот признак непригоден для различения B. italicus и B. hageni, что ранее уже отмечалось (Przybyłowicz, 2006).
Жилка А1 у B. hageni относительно короче, чем у B. italicus, как и указывает Эсбен-Петерсен (Esben-Petersen, 1921). Дорохова и Мартынова (1987) иначе интерпретируют этот же признак: А1 у B. italicus оканчивается посередине длины крыла, а у B. hageni проксимальнее середины длины крыла. Однако в действительности вершина А1 у B. italicus почти всегда лежит заметно проксимальнее середины длины крыла (рис. 2), что затрудняет правильное понимание этого признака в формулировке, предложенной Дороховой и Мартыновой (1987). По нашему мнению, оценивать относительную длину А1 удобнее всего, соотнося место ее впадения в задний край крыла с положением вершин 2-й кубитальной (Cu2) и 2-й анальной (А2) жилок. В этом случае описание этого признака будет следующим: вершина А1 у B. italicus на обоих крыльях лежит гораздо ближе к вершине Cu2, чем к вершине А2, а у B. hageni на переднем крыле – посередине расстояния между вершинами Cu2 и А2 или немного ближе к А2, а на заднем крыле – немного ближе к Cu2.
Дополнительно мы предлагаем следующий признак жилкования крыла для различения рассматриваемых видов: общий ствол Rs у B. italicus в 1.35–1.85 раза короче ячейки rs1, а у B. hageni он относительно длиннее, и отношение длины ячейки rs1 к Rs составляет 0.8–1.3.
Таким образом, для надежного различения обоих полов B. italicus и B. hageni можно использовать особенности жилкования крыла и его форму, а также, вполне вероятно, особенности хетотаксии ног и основания крыла. Ниже мы приводим таблицу для определения B. hageni и B. italicus только по признакам строения крыльев.
Таблица для различения Bittacus hageni и B. italicus по признакам строения крыльев
1(2) Крылья более узкие, длина переднего крыла в 4.6–5.3 раза больше ширины (измеренной на уровне проксимального края птеростигмы и места окончания М4). Общий ствол Rs значительно, в 1.35–1.85 раза, короче ячейки rs1. Вершина А1 на обоих крыльях лежит гораздо ближе к вершине Cu2, чем к вершине А2. Поперечная жилка m лежит под углом к продольной оси крыла (рис. 2) . . . . . . . . . . . . . . . . . . . . B. italicus (Müller 1766)
2(1) Крылья более широкие, длина переднего крыла в 3.9–4.4 раза больше ширины. Общий ствол Rs немного короче, такой же длины или немного длиннее ячейки rs1 (отношение длины rs1 к Rs составляет 0.8–1.3). На переднем крыле вершина А1 лежит посередине расстояния между вершинами Cu2 и А2 или немного ближе к А2, а на заднем крыле – немного ближе к Cu2. Поперечная жилка m лежит почти перпендикулярно продольной оси крыла (рис. 3, 1–7) . . . . . . . . B. hageni Brauer, 1860
Список литературы
Дорохова Г.И., Мартынова О.М., 1987. 26. Отряд Mecoptera – Скорпионовые мухи // В кн.: Определитель насекомых европейской части СССР. Т. IV. Большекрылые, верблюдки, сетчатокрылые, скорпионовые мухи и ручейники. Шестая часть. Л.: Наука. С. 97–106.
Бахтиев А.М., 2011. Особо охраняемые природные территории Чеченской Республики и развитие их оптимальной системы // Рефлексия. № 1. С. 3–13.
Мартынова О.М., 1959. Скорпионницы (Mecoptera) фауны СССР. III. Семейство Bittacidae // Энтомологическое обозрение. Т. 38. Вып. 1. С. 196−199.
Плутенко А.В., 1995. 26. Отряд Mecoptera – Скорпионницы, или скорпионовые мухи // Определитель насекомых Дальнего Востока России. Т. IV. Сетчатокрылообразные, скорпионницы, перепончатокрылые. Ч. 1. СПб.: Наука. С. 68–81.
Присный А.В., 2005. [Биттакус Хагени] Bittacus hageni Br. // Красная книга Белгородской области. Редкие и исчезающие растения, лишайники, грибы и животные. Официальное издание. Белгород: “Белгородская областная типография”. С. 358.
Уваров Б.П., 1910. Материалы по фауне Orthoptera Уральской области // Труды Русского Энтомологического Общества. Т. 39. С. 359−390.
Уваров Б.П., 1910а. К фayне чешуекрылых Зауральской киргизской степи // Русское энтомологическое обозрение. Т. 10. № 3. С. 161–169.
Уваров Б.П., Бородин Д.Н., 1909. Очерк флоры средней Эмбы // Русский ботанический журнал. № 6. С. 94–107.
Щуров В.И., Макаркин В.Н., 2017. Новые данные о сетчатокрылообразных (Neuroptera: Raphidioptera) и скорпионницах (Mecoptera) Северо-Западного Кавказа // Кавказский энтомологический бюллетень. Т. 13. № 1. С. 77–90.
Bornemissza G.F., 1966. Observations on the hunting and mating behaviour of two species of scorpion flies (Bittacidae: Mecoptera) // Australian Journal of Zoology. V. 14. P. 371–382.
Brauer F., 1855. Beiträge zur Kenntniss des inneren Baues und der Verwandlung der Neuropteren // Verhandlungen der kaiserlich-königlichen zoologisch-botanischen Gesellschaft in Wien. Bd. 5. S. 701–726.
Brauer F., 1860. Bittacus hageni, eine neue europäische Art, beschrieben und mit den verwandten Arten verglichen // Verhandlungen der kaiserlich-königlichen zoologisch-botanischen Gesellschaft in Wien. Bd. 10. S. 691–696.
Brauer F., 1871. Beiträge zur Kenntniss der Lebensweise und Verwandlung der Neuropteren (Micromus variegatus Fabr., Panorpa communis L., Bittacus italicus Klg., Bittacus hageni Brau.) // Verhandlungen der kaiserlich-königlichen zoologisch-botanischen Gesellschaft in Wien. Bd. 21. S. 107–116.
Crossley A.C., Waterhouse D.F., 1969. The ultrastructure of a pheromone-secreting gland in the male scorpion-fly Harpobittacus australis (Bittacidae: Mecoptera) // Tissue and Cell. V. 1. № 2. P. 273–294.
Devetak D., 1991. The genus Bittacus Latr. (Bittacidae, Mecoptera) in Yugoslavia and Albania // Zeitschrift der Arbeitsgemeinschaft Österreichischer Entomologen. Jg. 43. № 1/2. S. 51–54.
Dobosz R., Hadas T.B., 1999. Bittacus italicus (O.F. Müller, 1766) (Mecoptera: Bittacidae) w Polsce i na Ukrainie // Wiadomosci Entomologiczne. V. 17. № 3–4. S. 145–150.
Dobosz R., Pacuk B., 2018. Bittacus hageni Brauer, 1860 – a new locality in northern Poland, and a summary of current knowledge of the distribution and biology of this species // Annals of the Upper Silesian Museum in Bytom, Entomology. V. 26. P. 1–4.
Esben-Petersen P., 1921. Mecoptera. Monographic Revision // Collections Zoologiques du Baron Edm. de Selys Longchamps. Catalogue Systématique et Descriptif. Fasc. 5. Part. 2. P. 1–172.
Gao Q., Hua B.Z., 2013. Co-evolution of the mating position and male genitalia in insects: a case study of a hangingfly // PLoS ONE. V. 8. № 12. e80651. P. 1–8.
Gwynne D.T., 1984. Nuptial feeding behaviour and female choice of mates in Harpobittacus similis (Mecoptera: Bittacidae) // Journal of the Australian Entomological Society. V. 23. P. 271–276.
Londt J.G.H., 1972. Mecoptera of Southern Africa // Journal of the Entomological Society of Southern Africa. V. 35. № 2. P. 313–343.
Nidergas V., 2014. Contribution à l’étude des Mécoptères de France. Bittacus italicus (Müller, 1766), une nouvelle espèce pour le département de la Savoie (France, Rhône-Alpes) (Mecoptera Bittacidae) // L’Entomologiste. T. 70. № 3. P. 217–218.
Penny N.D., Byers G.W., 1979. A check-list of the Mecoptera of the World // Acta Amazonica. V. 9. № 2. P. 365–388.
Petschenka G., 2006. On the morphology of Bittacus hageni Brauer, 1860 and Bittacus italicus (Müller, 1766) (Mecoptera: Bittacidae) // Entomologische Zeitschrift. Bd. 116. № 3. S. 124–126.
Przybyłowicz Ł., 2006. Bittacus hageni Brauer, 1860 (Mecoptera: Bittacidae) – new to the fauna of Poland, with some remarks on Polish specimen of Bittacus italicus (O. F. Müller, 1766) // Polskie Pismo Entomologiczne. V. 75. № 3. P. 333–337.
Snodgrass R.E., 1935. Principles of Insect Morphology. London: Comstock Publishing Associates. 667 p.
Tan J.L., Hua B.Z., 2009. Bicaubittacus, a new genus of the Oriental Bittacidae (Mecoptera) with descriptions of two new species // Zootaxa. № 2221. P. 27–40.
Tillier P., 2008. Contribution à lʹétude des Mécoptères de France. Troisième partie: présence de Bittacus hageni Brauer dans le Val-dʹOise (Île-de-France, France). Synthèse des données françaises et européennes (Mecoptera Bittacidae) // L’Entomologiste. T. 64. № 3. P. 131–138.
Tillier P., Danflous S., Giacomino M., Jacquemin G., Maurel J.-P., Mazel R., 2009. Cartographie des Mécoptères de France (Mecoptera: Panorpidae, Bittacidae, Boreidae) // Revue de l’Association Roussillonnaise d’Entomologie. T. 18. № 1. P. 1–27.
Tjeder B., 1956. Zwei neue ost-asiatische Bittacus-arten (Mecoptera: Bittacidae) // Beiträge zur Entomologie. Bd. 6. № 1/2. S. 45–53.
Tjeder B., 1970. 18. Mecoptera // In: Tuxen S. L. (ed.). Taxonomist’s glossary of genitalia in insects. Copenhagen: Munksgaard. P. 99–104.
Tjeder B., 1974. Bittacidae from Burma, collected by R. Malaise (Mecoptera) // Entomologisk Tidskrift. V. 94 (1973). № 3–4. P. 188–193.
Tourneur J., 2019. Redécouverte pour l’ouest de la France d’un insecte rare en Europe: Bittacus hageni Brauer, 1860 (Mecoptera: Bittacidae) // Invertébrés Armoricains. V. 20. P. 8–10.
Vidlička Ľ., 2010. Srpice (Mecoptera) PR Šúr // Majzlan O. & Vidlička, Ľ. (eds). Príroda rezervácie Šúr. Bratislava: Ústav zoológie SAV. P. 277–284.
Vidlička Ľ., Kmeťová I., 2002. First record of Bittacus hageni (Insecta: Mecoptera, Bittacidae) in Slovakia // Biologia (Bratislava). V. 57. № 2. P. 170.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал