

Зоологический журнал, 2021, T. 100, № 8, стр. 938-946
Количественная оценка величины нерезидентной составляющей в локальных популяциях обыкновенных бурозубок (Sorex araneus)
Н. А. Щипанов *
Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: shchipa@mail.ru
Поступила в редакцию 15.06.2020
После доработки 31.08.2020
Принята к публикации 03.09.2020
Аннотация
Проведена количественная оценка нерезидентности (перемещений за пределами домашнего участка) у обыкновенной бурозубки. Использована особенность бурозубок – преимущественное попадание нерезидентных зверьков в ловушки типа “западня” (pitfall). На основании сопоставления обилия животных на одном и том же участке в течение 5 лет получены сравнительные данные по суточной величине потока нерезидентов, выявляемых на линии живоловок и в заборчиках с вкопанными ведрами. Обнаружена синхронность многолетней динамики обилия нерезидентов, учтенных разными способами. Исходя из количества зверьков, пойманных на погонный метр заборчиков, рассчитана суточная плотность нерезидентов. Обнаружено, что в отдельные годы суточная плотность нерезидентов может превышать плотность оседлого населения многократно. Коэффициент, который автор предлагает использовать в методике мечения, позволяет привести количество нерезидентов, выявляемых на линии, в соответствие с количеством нерезидентов, выявляемых в заборчиках.
Мощность “расселения” (dispersal) – одна из ключевых популяционных характеристик (Stenseth, Lidicker, 1992). Строгое содержание термина “расселение” подразумевает перемещение особи от места рождения до места, где она будет, или может, размножаться (Howard, 1960). Но часто этим термином обозначают любые перемещения особи за пределами домашнего участка: собственно расселение, врéменные дальние выходы за пределы участка и вынужденное “бродяжничество” (Lidicker, 1985). Расселение в широком смысле в значительной степени определяет возможность выживания вида в динамичной среде, его генетическую дифференциацию, возможную инвазивность и изменение ареала (Clobert et al., 2012). На практике “расселяющимися” считают всех особей, не давших возврата в пределы наблюдаемой территории (Southwood, Henderson, 2000; Карасева и др., 2008). На основании непредсказуемости перемещений зверьков вне домашнего участка их удобно рассматривать как “нерезидентов” (Щипанов, Купцов, 2004). Как и у многих видов, нерезидентность у обыкновенных бурозубок включает перемещения в ходе собственно расселения, далекие выходы с возвратом на домашний участок – экскурсии и безвозвратные дальние перемещения (Щипанов и др., 2008).
Хорошо известно, что при использовании устройств с ловушками типа “западня” (pitfall) (канавок, “тропинок” или заборчиков, оборудованных ловчими конусами или цилиндрами) улов составляют преимущественно зверьки, по разным причинам находящиеся за пределами домашнего участка, – “нерезиденты” (Наумов, 1955; Кучерук, 1963; Щипанов и др., 2003; Shchipanov et al., 2005; Щипанов и др., 2011). В давилки и живоловки попадаются и оседлые особи, и нерезиденты (Калинин, 2020).
Цель настоящей работы – сравнительная оценка величины потока нерезидентных землероек при отлове в живоловки и в “западни”. В задачи входили:
1) сравнение характеристик потока нерезидентов, полученных двумя способами на одной той же территории,
2) количественная оценка обилия нерезидентных особей в локальных популяциях обыкновенной бурозубки.
МАТЕРИАЛ И МЕТОДЫ
Место работы и животные. Использован материал, собранный в Старицком р-не Тверской обл. в 2006–2007 и 2009–2011 гг. Наблюдения проводили на двух участках 1) на злаково-разнотравном лугу (в дальнейшем “луг”), сформировавшемся в ходе выпаса, и 2) на зарастающем с 1998 г. картофельном поле (в дальнейшем – “поле”). В период наблюдений поле заросло разнотравным молодым березняком с низкими ивняками. В каждом местообитании была выставлена линия из 50 ловушек, с интервалом 7.5 м, длина линии 375 м. Около каждой десятой ловушки, начиная с пятой, на расстоянии 1.5–2 м от нее, в центре 10-метрового заборчика, ориентированного вдоль линии, было вкопано ведро. Вдоль каждой линии было устроено 5 заборчиков общей длиной 50 м. Каждое ведро выстилали слоем мха и ежедневно клали сюда рыбу, геркулес и овощи. Все животные, попавшиеся в ведро, в течение суток оставались живыми. Проверка заборчиков проходила до настораживания линий: зверьков метили и выпускали.
Отлов и мечение зверьков в ловушки проводили в соответствии с протоколом отлова, специально разработанным для землероек-бурозубок (Щипанов и др., 2000). Протокол предусматривает использование специализированной трапиковой ловушки (Щипанов, 1986), проверку через 1.5 ч после настораживания и последующую проверку через следующие 1.5 ч (всего 3 ч работы ловушек в сутки). Приманка – геркулес с нерафинированным подсолнечным маслом. После проверки ловушки оставляют открытыми в нерабочем положении. Для исключения связанных ловов в данной работе учитывалась только первая проверка.
Животных метили ампутацией концевых фаланг пальцев, что не влияет на выживание бурозубок (Shchipanov et al., 2005), в заборчиках и живоловках использовали сквозную нумерацию. Всего за время работы было помечено 582 зверька, из них попала в заборчики 161 особь. В ловушках отмечена 1861 поимка, из них 1269 повторных, в заборчиках отмечены 166 поимок, из них 5 повторных. Периоды работы заборчиков и число рабочих дней на линии ловушек в эти периоды показаны ниже (см. рис. 1, табл. 3).
Рис. 1.
Фактическое и ожидаемое распределение особей с разным числом поимок. По оси ординат – число особей, по оси абсцисс – число поимок. Даты наверху – период одновременной работы живоловок и заборчиков. В скобках: n – число рабочих дней/проверок на линии живоловок; мах – максимальное число поимок, для которого найдено наибольшее совпадение. Столбики – фактическое число особей. а – число нерезидентов, б – число резидентов, в – наиболее совпадающее ожидаемое распределение, г – ожидаемое распределение для максимального числа (мах) минус 1 поимка, д – ожидаемое распределение для максимального числа (мах) плюс 1 поимка.
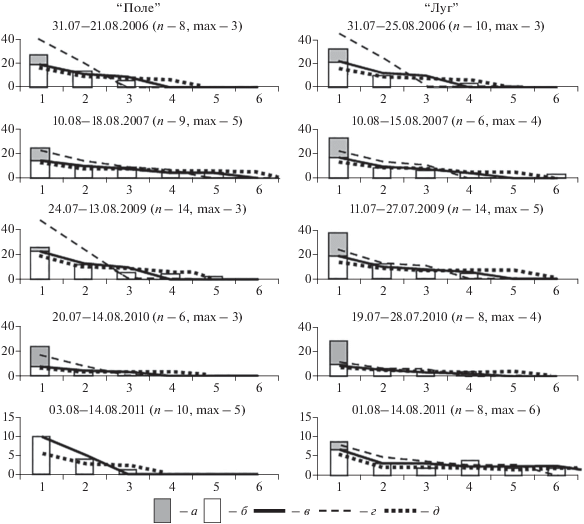
Расчеты на линиях живоловок. Для расчета количества нерезидентов на линиях, на основании анализа частотного распределения особей с разным числом повторных поимок, первоначально определено общее количество оседлых. Расчеты исходят из того, что у среднестатистической особи число поимок максимально в центре участка и убывает к периферии по закону нормального распределения (Shchipanov et al., 2005; Щипанов и др., 2008, 2011; Калинин, 2012). Соответственно максимальное количество поимок может быть получено при попадании ловушки в центр участка, а минимальное – на его периферию. Подробное обоснование и проверка этих методов расчета опубликованы ранее (Щипанов, 2020).
В соответствии с методикой (Щипанов, 2020) расчет основан на подборе фактического распределения числа особей с разным количеством поимок, наиболее близкого к ожидаемому. Распределение ожидаемых долей особей с повторными ловами (N>1) для максимального числа поимок до 10 приведено в табл. 1. Общее число особей с повторными ловами на линии умножено на доли из табл. 1 для ожидаемого максимального числа поимок от 2 до 10 и, по критерию минимальной суммы квадратов отклонений, выбрано распределение, наиболее близкое к фактическому. Число оседлых особей с единичными ловами (N1) получено умножением числа всех особей с повторными поимками на коэффициент K (см. табл. 1) для соответствующего числа максимальных поимок: N1 = KN>1. Общее число резидентов (R) – сумма всех резидентов: R = N>1 + KN>1. Число нерезидентов на линии (Nr) получено при вычитании количества резидентов (R) из общего числа помеченных особей (N): Nr = N – R. Полученные распределения и определяемое число нерезидентов для всех сессий показаны на рис. 1.
Таблица 1.
Доли особей с числом поимок более 1 и коэффициент K, в зависимости от максимального числа поимок (по: Щипанов 2020)
| Число поимок особи | Максимальное число поимок | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| 2 | 1.00 | 0.57 | 0.44 | 0.36 | 0.25 | 0.23 | 0.21 | 0.21 | 0.20 |
| 3 | 0 | 0.43 | 0.33 | 0.27 | 0.25 | 0.23 | 0.21 | 0.14 | 0.13 |
| 4 | 0 | 0 | 0.22 | 0.18 | 0.17 | 0.15 | 0.14 | 0.14 | 0.13 |
| 5 | 0 | 0 | 0 | 0.18 | 0.17 | 0.15 | 0.14 | 0.07 | 0.13 |
| 6 | 0 | 0 | 0 | 0 | 0.17 | 0.15 | 0.07 | 0.14 | 0.07 |
| 7 | 0 | 0 | 0 | 0 | 0 | 0.08 | 0.14 | 0.07 | 0.07 |
| 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0.07 | 0.14 | 0.13 |
| 9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.07 | 0.07 |
| 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.07 |
| K | 2.00 | 1.00 | 0.78 | 0.55 | 0.50 | 0.46 | 0.36 | 0.43 | 0.33 |
Количество нерезидентов, определенное на линиях, отнесено к количеству проверок. Поскольку при расчетах использована только первая проверка, получен среднесуточный улов нерезидентов на линии (Nrt): Nrt = Nr/T, где T – число проверок равное числу рабочих дней.
Популяционная плотность (DR) рассчитана как число оседлых особей в пределах площади возможного обнаружения (S): DR = R/S. Эта площадь получена как произведение длины линии (L) на удвоенный радиус среднестатистического участка (в метрах), на котором особи могут дать хотя бы 1 поимку, с коэффициентом 10 000–1 для перевода в гектары: S = 2Lks/10 000, где k – коэффициент для дистанции, на которой при максимальном числе поимок, определенных для выборки, может быть получена хотя бы 1 поимка (для 1, 2, 3, 4, 5, 6, 7, 8, 9 и 10 поимок k равен 1.1, 1.3, 1.5, 1.6, 1.7, 1.8, 1.8, 1.9 и 1.9 соответственно), s – стандартное отклонение дистанции координат поимок особи (di) от средней координаты ее поимок во всей выборке: s = √Σ$d_{i}^{2}$/(n – 1) (n – общее число поимок в выборке). Дистанция di рассчитана для каждой особи с повторными поимками: di = x – xi, где x – среднее значение координат (номеров ловушек) всех регистраций особи, xi – номер ловушки, в которую пойман зверек.
Расчеты для заборчиков. Всех особей, пойманных в заборчики и не давших повторов (и в заборчики, и на линии), считали нерезидентами (Nf). Работы проводились с использованием заборчика с одним ведром в середине. Выходы в противоположные от ведра стороны оставались свободными. Предполагается, что нерезидентная особь, наткнувшись на заборчик, может двигаться к ведру и в противоположную сторону с равной вероятностью, т.е. величина улова нерезидентов составляет 0.5 от фактического числа нерезидентов (N), натолкнувшихся на заборчик: Nf = 0.5N. В среднем за сутки на одного нерезидента, пересекающего трансект длиной (Lt), приходится некоторая часть – “суточный интервал” (dNf), который равен Lt/2Nf. Этот интервал одинаков в любом направлении, соответственно 0.5 dNf рассматривалось как радиус площади, приходящейся на одного нерезидента в сутки (S): S = π(Lt/4Nf)2, а суточная плотность (DNf) рассчитана как количество таких площадей на 1 гектаре: DNr = = 10 000/π(Lt/4Nf)2. Обращаю внимание на то, что эта формула применима только для расчета улова нерезидентов в заборчики с центральным ведром и двумя свободными выходами.
Статистика. Соответствие динамики оценено непaраметрическим критерием соответствия Спирмeна (Rs), как это рекомендовано для популяций красно-серой полевки (Bjørnstad et al., 1998; Saitoh et al., 1998). Для нормализации распределений все показатели обилия логарифмированы, нуль заменен на 0.1. Нормальными признаны распределения для p > 0.2 в тесте Шапиро-Уилка. Связь нормализованных данных оценена коэффициентом корреляции Пирсона (r). Логарифмы популяционной плотности сравниваются попарным t тестом (в паре – значения одного года). Влияние факторов “место” и “год” оценено двухфакторным ANOVA для основных эффектов. Расчеты сделаны в пакете программ Statistica 7. При оценке динамики обилия использован S-индекс (Si): Si = sd lg 10Dx, где Dx – плотность соответствующей категории населения (Stenseth, Framstad, 1980; Henttonen et al., 1985).
РЕЗУЛЬТАТЫ
Учет живоловками. Максимальное среднесуточное количество нерезидентов отмечено на “поле” в 2010 г. и составило 2.7, минимальное, там же, в 2011 – 0 (табл. 2). Среднесуточный улов нерезидентов в ловушках не зависел от продолжительности учета (r = –0.086, p > 0.81) и максимального числа поимок (r = –0.006, p > 0.98) и не был связан с обилием оседлых особей (r = 0.46, p > 0.17). На среднесуточное обилие нерезидентов фактор “место” не влиял (df = 1, F = 2.2, p > 0.21). Влияние фактора “год” было близко к достоверному (df = 4, F = 5.2, p < 0.07). Среднесуточный улов на участках “поле” и “луг” не различался (t = 1.38, df = 4, p > 0.23), его межгодовая динамика практически совпадала (Rs = 0.9, p < 0.01).
Таблица 2.
Расчет числа оседлых и нерезидентных особей на линии живоловок
| Год | Место | Рабочие дни = = проверки | Число особей, определенных как | s (метры) | max | k | DR (особи/га) | Nrt | |
|---|---|---|---|---|---|---|---|---|---|
| оседлые | нерезиденты | ||||||||
| 2006 | Поле | 8 | 40 | 9 | 10.9 | 3 | 1.3 | 37.6 | 1.1 |
| Луг | 10 | 44 | 11 | 14.3 | 3 | 1.3 | 31.5 | 1.1 | |
| 2007 | Поле | 9 | 41.58 | 10 | 9.5 | 5 | 1.6 | 36.5 | 1.1 |
| Луг | 6 | 38.94 | 12 | 12.6 | 4 | 1.5 | 27.5 | 2.0 | |
| 2009 | Поле | 14 | 46 | 3 | 11.0 | 5 | 1.6 | 35.0 | 0.2 |
| Луг | 13 | 40.71 | 19 | 14.6 | 4 | 1.5 | 24.8 | 1.5 | |
| 2010 | Поле | 6 | 16 | 16 | 10.8 | 3 | 1.3 | 15.2 | 2.7 |
| Луг | 8 | 19.47 | 19 | 14.1 | 4 | 1.5 | 12.3 | 2.4 | |
| 2011 | Поле | 10 | 15 | 0 | 10.8 | 3 | 1.3 | 14.2 | 0.0 |
| Луг | 8 | 19.63 | 2 | 13.5 | 6 | 1.7 | 11.4 | 0.3 | |
Таблица 3.
Расчет обилия нерезидентов по данным отлова в заборчики
| Год | Место | Рабочие дни | Особи | Суточный интервал между Nr м |
S на одного Nr м.кв | DNr особи/га | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| всего | пойманы повторно | нерези-денты (Nr) | Nr за сутки | Nr на 100 м | ||||||
| 2006 | Луг | 9 | 14 | 5 | 9 | 1.0 | 4.0 | 25.0 | 1963.5 | 5.1 |
| Поле | 22 | 23 | 5 | 18 | 0.8 | 3.3 | 30.6 | 2933.1 | 3.4 | |
| 2007 | Луг | 3 | 6 | 0 | 6 | 2.0 | 8.0 | 12.5 | 490.9 | 20.4 |
| Поле | 9 | 18 | 4 | 14 | 1.6 | 6.2 | 16.1 | 811.4 | 12.3 | |
| 2009 | Луг | 5 | 9 | 1 | 8 | 1.6 | 6.4 | 15.6 | 767.0 | 13.0 |
| Поле | 15 | 25 | 3 | 22 | 1.5 | 5.9 | 17.0 | 912.8 | 11.0 | |
| 2010 | Луг | 8 | 25 | 4 | 21 | 2.6 | 10.5 | 9.5 | 285.0 | 35.1 |
| Поле | 8 | 37 | 4 | 33 | 4.1 | 16.5 | 6.1 | 115.4 | 86.7 | |
| 2011 | Луг | 13 | 3 | 2 | 1 | 0.1 | 0.3 | 325.0 | 331 830.7 | 0.03 |
| Поле | 12 | 1 | 0 | 1 | 0.1 | 0.3 | 300.0 | 282 743.3 | 0.04 | |
Максимальная популяционная плотность 37.6 особей на 1 га отмечена в 2006 г. на поле, а минимальная – 11.4 – в 2011 г. на лугу (см. табл. 2). На обилие оседлых влияют факторы “место” (df = 1, F = 71.9, p < 0.002) и “год” (df = 4, F = = 220.3, p < 0.001). Плотность оседлых была достоверно (t = 8.48, df = 4, p < 0.001) выше на “поле”: средняя геометрическая плотность за рассматриваемый период на лугу составила 19.8, на “поле” – 25.3 особи/га. Изменения плотности на участках проходили абсолютно синхронно (Rs = 1).
Учет заборчиками. Всего в заборчики была поймана 161 землеройка. Все они, за исключением пяти, попались однократно. Два зверька были пойманы повторно, на следующий день, и еще один зверек был пойман через день, в те же ведра, из которых все эти зверьки были выпущены. Еще два зверька попались в заборчики повторно на расстоянии около 200 и около 300 м от места первой поимки. Из попавшихся в ведра зверьков 23 особи ловились также и в ловушки на линии, где были определены как оседлые. Таким образом, в улове в заборчики присутствовало около 14% оседлых зверьков. При этом на линиях оседлым был признан 321 зверек, и, следовательно, только 7% оседлых особей попадало в ведра. От всех пойманных в ловушки зверьков (n = 421) особи, пойманные в заборчиках, составляют около 5%, или около 2% от всех поимок (n = 1105) в ловушки.
При расчете нерезидентов особи, попадавшиеся повторно и в ведра, и в ловушки, из выборки исключены. Среднесуточный улов вдоль линии, т.е. суммарно во все 5 заборчиков, дистанция между нерезидентами, площадь, приходящаяся на одного нерезидента в сутки, и суточная плотность нерезидентов представлены в табл. 3. Среднесуточный улов в заборчиках и, соответственно, расчетная плотность нерезидентов не связаны с продолжительностью учета (r = –0.38, p > 0.28) и не коррелируют с плотностью оседлых (r = –0.37, p > 0.37). Фактор “место” не влияет на суточный улов нерезидентов (df = 1, F = 0.001, p > 0.99), а фактор “год” – влияет высоко значимо (df = 4, F = 102.8, p < 0.001). Средняя геометрическая суточной плотности нерезидентов на “лугу” – 17.0, на “поле” 17.1 особей/га в сутки. Динамика обилия нерезидентов на участках синхронна (Rs = 1).
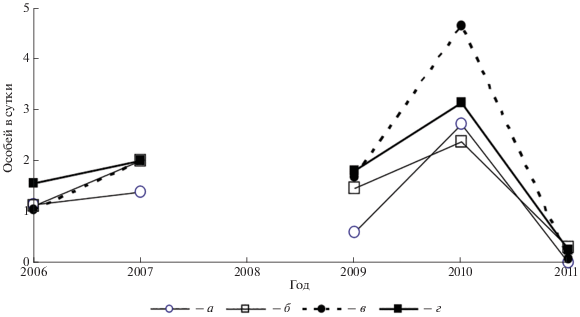
Сравнение данных ловушек и заборчиков. Межгодовая динамика среднесуточного улова нерезидентов в живоловки и заборчики (рис. 2) синхронна (Rs = 0.93, p < 0.01). Число зверьков, попадающихся за сутки вдоль всей линии в ловушки и в заборчики, различается лишь на уровне тенденции (t = 1.9, df = 9, p < 0.092), среднее геометрическое 0.95 и 1.1 особи, но при сравнении улова на погонный метр трансекта обилие нерезидентов, при учете в заборчик, многократно выше. На 100 м линии ловушек среднее геометрическое число нерезидентов в сутки составляет 0.2, а в заборчиках – 4.7 особи. Таким образом, при расчете на единицу длины заборчика среднесуточное количество определяемых нерезидентов в 24 раза больше, чем в ловушках. Однако замечу, что суточное время экспозиции ловушек составляло лишь 1/16 суточного времени работы заборчиков.
Рис. 2.
Динамика улова нерезидентов на линии живоловок и в заборчиках вдоль линии ловушек: а – в ловушки на линии “поле”, б – в ловушки на линии “луг”, в – в заборчики вдоль линии “поле”, г – в заборчики вдоль линии “луг”.
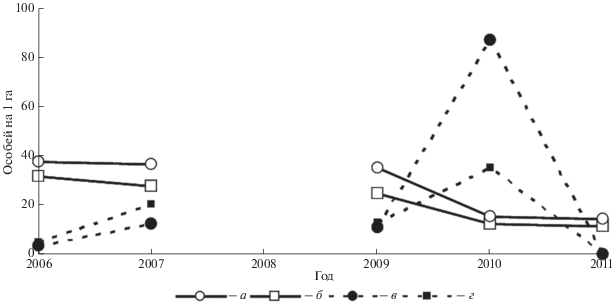
Суточная плотность нерезидентного населения составила в среднем около 20% от плотности оседлых. Плотность оседлых в период наблюдений почти не изменяется, а плотность нерезидентов существенно колеблется и в 2010 г. многократно превышала обилие оседлых (рис. 3). Для плотности оседлых Si равен 0.21 на “поле” и 0.22 на лугу, для суточной плотности нерезидентов за тот же период Si = 1.26 и 1.24 соответственно. Разница плотности предыдущего и последующего годов, по отношению к средней геометрической, для нерезидентов составляла, в среднем, на “поле” 10.1 и на лугу 4.7, а для оседлых, за тот же период, – 0.25 и 0.37 соответственно.
ОБСУЖДЕНИЕ
Как и ожидалось, в заборчики попадаются преимущественно нерезидентные землеройки, а землеройки, жившие вблизи от “западней”–ведер, оседлые, в ведра почти не попадали, однако динамика улова нерезидентов в живоловках и в заборчиках синхронна, а величина улова вполне сравнима. Это подтверждает результаты, полученные ранее для рыжих полевок и землероек: в живоловках и плашках отмечено присутствие заметного количества нерезидентных особей, а динамика нерезидентного населения соответствует динамике улова в ловчие канавки (Калинин, 2012; Калинин и др., 2020).
Если в ловчих устройствах с “западнями”– ведрами получена практически чистая выборка нерезидентов, то в наземных ловушках выборка смешанная. И демографический состав, и поведение, и физиология у нерезидентов и оседлых особей могут существенно различаться (Lidicker, 1985; Щипанов, Купцов, 2004; Щипанов, 2002, 2020а). Оценить долю нерезидентов в популяции a priori невозможно. Величина нерезидентного потока лишь в половине случаев связана с плотностью оседлого населения (Lidicker, 1985; Matthysen, 2005; Rachel et al., 2020). Полученные в настоящей работе данные показывают, что обилие нерезидентного населения может меняться со значительно большей амплитудой, чем плотность оседлого населения, кроме того, обилие нерезидентов в разных местах не различалось, а плотность оседлых различна и, соответственно, вклад нерезидентов в улов также был разный.
Оценка нерезидентной составляющей важна для понимания динамики популяции в целом (Korpimäki et al., 2004; Henttonen et al., 2017). Однако соотношение резидентной и нерезидентной составляющих, даже в классических работах, часто не учитывают (например, Sheftel, 1989; Захаров и др., 2011; Кшнясев, 2014; Кшнясев, Давыдова, 2015; Бобрецов и др., 2020). Вместе с тем дифференцированная оценка населения может коренным образом изменить представления о типе популяционной динамики. В нашем случае амплитуда колебаний обилия землероек в заборчиках соответствует циклической динамике, Si > 1, что значительно превышает критическое значение 0.5 для циклических популяций (Henttonen et al., 1985), в то время как плотность оседлого населения мало изменчива, Si < 0.22.
У обыкновенной бурозубки оценка величины нерезидентной составляющей важна еще и для понимания эколого-эволюционных процессов: благодаря ярко выраженному хромосомному полиморфизму, этот вид представляет удобную модель для таких исследований (Zima, Searle, 2019). Одна из проблем – несоответствие между молекулярной и кариотипической изменчивостью, известными у этого вида (Horn et al., 2012; Yannic et al., 2019) – возможно связана с чрезвычайно высоким генетическим разнообразием локальных популяций (Щипанов, Павлова, 2019). В настоящем исследовании нерезидентная компонента по величине улова сравнима с оседлой. Это, с одной стороны, фокусирует внимание на способе взятия выборки, а, с другой, – заставляет учитывать возможность быстрого изменения локального генетического разнообразия.
Оценка величины нерезидентной части населения важна и для прикладных исследований. Обилие мелких млекопитающих учитывают при мониторинге окружающей среды (Бердюгин, Большаков, 2005; Истомин, 2014; Ильяшенко и др., 2015) и природных очагов инфекций (Мазин и др., 2013; Транквилевский и др., 2014, 2016). Обычно оценивают величину улова в ловушки или в ловчие устройства с “западнями” без разделения на оседлую и нерезидентную составляющие (Карасева и др., 2008; Шефтель, 2018). Вместе с тем и для оценки эпизоотического процесса и для адекватной оценки изменений в окружающей среде важна оценка величины нерезидентной части населения. Увеличение доли нерезидентной составляющей – одна из наиболее быстрых популяционных реакций на дестабилизацию среды (Мухачева, Лукьянов, 1997; Лукьянова, Лукьянов, 2004; Luk’yanov, Luk’yanova, 2002; Горигоркина, Оленев, 2013; Щипанов, 2002, 2020а). О роли динамической плотности в эпизоотическом процессе писал еще Раль (1945).
Обобщая сказанное, можно заключить, что результаты исследования позволяют говорить о валидности оценок потока нерезидентов на основе данных, получаемых при мечении зверьков на линии ловушек. В нашем случае число нерезидентов, обнаруженных в ловушках на линии с коэффициентом 24 (24Nr), соответствует действительному количеству нерезидентов, пересекающих ее по всей длине в сутки. Особенностью нерезидентной части населения является накопление улова, прямо пропорциональное времени учета, это создает сложности в оценке обилия нерезидентов в популяции. Я предлагаю для оценки обилия нерезидентов использовать число нерезидентов, присутствующих на 1 га в сутки. Это число рассчитывается на основании числа особей, пересекающих единицу трансекта.
Список литературы
Бердюгин К.И., Большаков В.Н., 2005. Млекопитающие в экологическом мониторинге // Методы экологического мониторинга: большой специальный практикум: учебное пособие. Екатеринбург: изд-во Уральского университета. С. 192–201.
Бобрецов А.В., Лукьянова Л.Е., Петров А.Н., Быховец Н.М., 2020. Периодические изменения численности средней бурозубки (Sorex caecutiens, Eulipotyphla) в предгорной тайге Северного Урала и факторы ее популяционной динамики // Экология. Вып. 3. С. 224–229.
Григоркина Е.Б., Оленев Г.В., 2013. Миграции грызунов в зоне влияния Восточно-Уральского радиоактивного следа (радиобиологический аспект) // Радиационная биология. Радиоэкология. Т. 53. Вып. 1. С. 76–83.
Захаров В.М., Шефтель Б.И., Дмитриев С.Г., 2011. Изменение климата и популяционная динамика: возможные последствия (на примере мелких млекопитающих в Центральной Сибири) // Успехи современной биологии. Т. 131. Вып. 5. С. 435–439.
Ильяшенко В.Б., Лучникова Е.М., Скалон Н.В., 2015. Мелкие млекопитающие как объект биомониторинговых исследований на территории Кемеровской области // Вестник Кемеровского государственного университета. Т. 3. № 4 (64). С. 25–30
Истомин А.В., 2014. Мелкие млекопитающие в мониторинге лесных экосистем: комплексный подход // Вестник Тверского гос. университета. Сер. Биология и экология. № 4. С. 95–113.
Калинин А.А., 2012. Оседлая и нерезидентная составляющая численности массовых видов мелких млекопитающих по данным учета на линиях живоловок // Зоологический журнал. Т. 91. Вып. 6. С. 759–768.
Калинин А.А., Куприянова И.Ф., Александров Д.Ю., 2020. Вклад плотности оседлого населения и нерезидентной активности мелких млекопитающих в результаты учетов методом безвозвратного изъятия // Сибирский экологический журнал. № 2. С. 233–242.
Карасева Е.В., Телицына А.Ю., Жигальский О.А., 2008. Методы изучения грызунов в полевых условиях. М.: Изд-во ЛКИ. 416 с.
Кучерук В.В., 1963. Новое в методике количественного учета вредных грызунов и землероек // Организация и методы учета птиц и вредных грызунов. М.: Изд-во АН СССР. С. 159–183.
Кшнясев И.А. 2014. Изменение режима динамики популяций мелких млекопитающих: диагностика и симптомы, улики и главные подозреваемые. (Экологический детектив: наброски) // Математические модели в теоретической экологии и земледелии (Полуэктовские чтения). Материалы международного семинара. Санкт-Петербург, 14–16 октября. СПб.: АФИ. С. 88–92.
Кшнясев И.А., Давыдова Ю.А., 2015. Наблюдаемая динамика популяций: детерминизм и стохастика, циклы и хаос // Всероссийский семинар “Гомеостатические механизмы биологических систем” с общей темой “Проблемы популяционной экологии” (6–10 апреля 1915 г., Тольятти, Россия). Под ред. Розенберга Г.С. Тольятти: Кассандра. С. 186–191.
Лукьянова Л.Е., Лукьянов О.А., Экологически дестабилизированная среда: влияние на население мелких млекопитающих // Экология. № 3. С. 210–217.
Мазин Л.Н., Каштанкин М.Н., Румянцева Л.Н., 2013.Териологический и паразитологический мониторинг за популяциями мелких млекопитающих на незастроенной территории Москвы. Сообщение 1. Териологический мониторинг в 2002–2011 гг. // Дезинфекционное дело. № 2. С. 39–52.
Мухачева С.В., Лукьянов О.А., 1997. Миграционная подвижность населения рыжей полевки (Clethrionomys glareolus (Schreber, 1780)) в градиенте техногенных факторов // Экология. № 1. С. 34–39.
Наумов Н.П., 1955. Изучение подвижности мелких млекопитающих с помощью ловчих канавок // Вопросы краевой общей и экспериментальной паразитологии и медицинской зоологии. Т. 9. С. 179–202.
Раль Ю.М., 1945. Динамическая плотность грызунов и методы ее изучения // Бюллетень Московского общества испытателей природы. Т. 50. Вып. 5–6. С. 62–64.
Транквилевский Д.В., Квасов Д.А., Мещерякова И.С., Михайлова Т.В., Кормилицына М.И., Демидова Т.Н., Ананьина Ю.В., Савельева О.В., Малкин Г.А., Мутных Е.С., Коротина Н.А., Дзагурова Т.К., Простаков Н.И., Сурков А.В., Куролап С.А., Клепиков О.В., Стёпкин Ю.И., Чубирко М.И., Жуков В.И., 2014. Вопросы организации мониторинга природных очагов инфекций опасных для человека. Планирование, проведение и анализ результатов полевых наблюдений // Здоровье населения и среда обитания. Вып. 8. С. 38–43.
Транквилевский Д.В., Царенко В.А., Жуков В.И., 2016. Современное состояние эпизоотологического мониторинга за природными очагами инфекций в Российской Федерации // Медицинская паразитология и паразитарные болезни. Вып. 2. С. 19–24.
Шефтель Б.И., 2018. Методы учета численности мелких млекопитающих // Russian journal of ecosystem ecology. Т. 3. С. 1–21.
Щипанов Н.А., 1986. К экологии малой белозубки (Crocidura suaveolens) // Зоологический журнал. Т. 66. Вып. 7. С. 1051–1060.
Щипанов Н.А., 2002. Функциональная организация популяции: возможный подход к изучению популяционной устойчивости. Прикладной аспект (на примере мелких млекопитающих) // Зоологический журнал. Т. 81. Вып. 9. С. 1048–1077.
Щипанов Н.А., 2020. Мечение на линиях живоловок для мониторинга мелких млекопитающих. Способы расчета популяционной плотности и индексов нерезидентности // Зоологический журнал. Т. 99. № 9. С. 1062–1076.
Щипанов НА., 2020а. Популяционная устойчивость мелких млекопитающих. Почему это важно и от чего это зависит? // Поволжский экологический журнал. Т. 4. С. 503–523.
Щипанов Н.А., Калинин А.А., Олейниченко В.Ю., Демидова Т.Б., Гончарова О.Б., Нагорнев Ф.В., 2000. К методике изучения использования пространства землеройками-бурозубками // Зоологический журнал. Т. 79. № 3. С. 362–371.
Щипанов Н.А., Купцов А.В., 2004. Нерезидентность у мелких млекопитающих и ее роль в функционировании популяции // Успехи современной биологии. Т. 124. Вып. 1. С. 28–43.
Щипанов Н.А., Купцов А.В., Калинин А.А., Олейниченко В.Ю., 2003. Конуса и живоловки ловят разных землероек-бурозубок (Insectivora, Soricidae) // Зоологический журнал. Т. 82. № 10. С. 1258–1265.
Щипанов Н.А., Павлова С.В., 2019. Роль популяционного структурирования в формировании кариотипического разнообразия обыкновенной бурозубки Sorex araneus (Lipotyphla, Mammalia) // Экология. № 2. С. 101–111.
Щипанов Н.А., Тумасьян Ф.А., Распопова А.А., Купцов А.В., 2011. Два типа использования пространства оседлыми обыкновенными бурозубками Sorex araneus L. // Известия Российской академии наук. Серия биологическая. Вып. 1. С. 107–112.
Щипанов Н.А., Купцов А.В., Демидова Т.Б., Калинин А.А., Александров Д.Ю., Павлова С.В., 2008. Нерезидентность и расселение у обыкновенных бурозубок (Sorex araneus, Insectivora) // Зоологический журнал. Т. 87. № 3. С. 331–343.
Bjørnstad O.N., Stenseth N.C., Saitoh T., Lingjære O.C., 1998. Mapping the regional transitions to cyclicity in Clethrionomys rufocanus: spectral densities and functional data analysis // Population Ecology. V. 40. P. 77–84.
Clobert J., Baguette M., Benton T.J., Bullock M., 2012. Preface. Clobert J., Baguette M., Benton T.J., Bullock M. (eds). Dispersal ecology and evolution. Oxford University Press. P. xiv-xv.
Henttonen H., Gilg O., Ims R.A., Korpimäk E., Yoccoz N.G., 2017. Ilkka Hanski and small mammals: from shrew metapopulations to vole and lemming cycles // Annales zoologici Fennici. V. 54 (1–4). P. 153–162.
Henttonen H., McGuire A.D., Hansson L., 1985. Comparisons of amplitudes and frequencies (spectral analyses) of density variations in long-term data sets of Clethrionomys species // Annales Zoologici Fennici. V. 22. № 3. P. 221–227.
Horn A., Basset P., Yannic G., Banaszek A., Borodin P.M., Bulatova N.S., Jadwiszczak K., Jones R.M., Polyakov A.V., Ratkiewicz M., Searle J., Shchipanov N.A., Zima J., Hausser J., 2012. Chromosomal rearrangements do not seem to affect the gene flow in hybrid zones between karyotypic races of the common shrew (Sorex araneus) // Evolution. V. 66. P. 882–889.
Howard W.E., 1960. Innate and environmental dispersal of individual vertebrates // American Midland Naturalist. V. 63. P. 152–161.
Korpimäki E., Brown P.R., Jacob J., Pech R.P., 2004. The puzzles of population cycles and outbreaks of small mammals solved? // Bioscience. V. 54 (12). P. 1071–1079.
Lidicker W.Z., Jr., 1985. Dispersal. The American Society of Mammalogists. Special publication. V. 8. P. 420–454.
Luk’yanov O.A., Luk’yanova L.E., 2002. Phenomenology and analysis of dispersal in populations of small mammals // Entomological Review. V. 82 (1). P. 181.
Matthysen E., 2005. Density-dependent dispersal in birds and mammals // Ecography. V. 28 (3). P. 403–416.
Rachel R., Goddard H. J., Shivaji R., Cronin J.T., 2020. Frequency of Occurrence and Population-dynamic consequences of different forms of density-dependent emigration // The American Naturalist. V. 195 (5). P. 851–867.
Saitoh T., Stenseth N.C., Bjørnstad O.N., 1998. The population dynamics of the vole Clethrionomys rufocanus in Hokkaido, Japan // Population Ecology. V. 40. № 1. P. 61–76.
Shchipanov N.A., Kalinin A.A., Demidova T.B., Oleinichenko V.Yu., Aleksandrov D.Yu., Kouptzov A.V., 2005. Population ecology of red-toothed shrews, Sorex araneus, S. caecutiens, S. minutus, and S. isodon, in Central Russia // Advances in the Biology of Shrews II. J. Merrit, S. Churchfield, R. Hutterer, B.I. Sheftel (eds). N.Y. Special publication of the international society of shrew biologists. № 1. P. 201–216.
Sheftel B.I., 1989. Long-term and seasonal dynamics of shrews in Central Siberia // Annales Zoologici Fennici. V. 26. P. 357–369.
Southwood T.R.E., Henderson P.A., 2000. Ecological methods. Third edition. Blackwell Science. 592 p.
Stenseth N.C., Framstad E., 1980. Reproductive effort and optimal reproductive rates in small rodents // Oikos. V. 34. № 1. P. 23–34.
Stenseth N.C., Lidicker Jr W.Z., 1992. Preface. Stenseth N.C., Lidicker Jr W.Z., Lidicker W.Z. (eds). Animal dispersal: small mammals as a model. Springer Science & Business Media. P. xii-xiii.
Yannic G., Basset P., Horn A., Hausser J., 2019. Gene flow between chromosomal races and species. Shrews, Chromosomes and Speciation. Cambridge: Cambridge University Press. P. 313–335.
Zima J., Searle J.B., 2019. Milestones in Common Shrew Chromosomal Research. Shrews, Chromosomes and Speciation. Cambridge: Cambridge University Press. P. 1–18.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал