

Зоологический журнал, 2021, T. 100, № 9, стр. 1028-1054
Миграции красавки (Anthropoides virgo, Gruiformes): дистанционное слежение на путях пролета и зимовках
Е. И. Ильяшенко a, *, Е. А. Мудрик b, **, Ю. А. Андрющенко c, ***, В. П. Белик d, ****, О. В. Белялов a, М. Викельски e, *****, А. Э. Гаврилов f, ******, О. А. Горошко g, *******, Е. В. Гугуева h, ********, М. В. Корепов i, *********, Р. А. Мнацеканов j, **********, Д. В. Политов b, ***********, К. А. Постельных k, ************, Цао Лей l, *************, В. Ю. Ильяшенко a, **************
a Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Россия
b Институт общей генетики имени Н.И. Вавилова РАН
119991 Москва, Россия
c Азово-Черноморская орнитологическая станция Института зоологии имени И.И. Шмальгаузена НАНУ
72312 Мелитополь, Украина
d Академия биологии и биотехнологии имени Д.И. Ивановского Южного федерального университета
344006 Ростов-на-Дону, Россия
e Институт поведения животных Макса Планка
04103 Радольфцелль, Германия
f Институт зоологии Министерства образования и науки Республики Казахстан
050040 Алматы, Казахстан
g Институт природных ресурсов, экологии и криологии СО РАН
672014 Чита, Забайкальский край, Россия
h Природный парк “Волго-Ахтубинская пойма”
404147 Волжск, Волгоградская область, Россия
i Ульяновский государственный педагогический университет имени И.H. Ульянова
432700 Ульяновск, Россия
j Региональное отделение Всемирного фонда дикой природы “Северный Кавказ”
350020 Краснодар, Россия
k Окский государственный природный биосферный заповедник
391071 Брыкин бор, Рязанская область, Россия
l Исследовательский центр по экологии и окружающей среде Китайской Академии наук,
Университет Китайской Академии наук
38400 Пекин, Китай
* E-mail: eilyashenko@savingcranes.org
** E-mail: mudrik@vigg.ru
*** E-mail: anthropoides73@gmail.com
**** E-mail: vpbelik@mail.ru
***** E-mail: wikelski@ab.mpg.de
****** E-mail: aegavrilov@bk.ru
******* E-mail: oleggoroshko@mail.ru
******** E-mail: elenagugueva@yandex.ru
********* E-mail: korepov@list.ru
********** E-mail: ramnatsekanov@mail.ru
*********** E-mail: dmitri.p17@gmail.com
************ E-mail: kirill_cbc@mail.ru
************* E-mail: leicao@rcees.ac.cn
************** E-mail: valpero53@gmail.com
Поступила в редакцию 25.05.2020
После доработки 27.12.2020
Принята к публикации 31.12.2020
Аннотация
В 2017–2020 гг. с использованием передатчиков GPS-GSM уточнены маршруты миграций красавок (Anthropoides virgo), помеченных на Украине, юге России и в Казахстане. Выявлено восемь миграционных потоков. Журавли из европейской части ареала следуют двумя миграционными потоками к разным местам зимовки, используя осенью и весной один и тот же маршрут. Установлено, что азово-черноморская группировка зимует в Чаде, где нами впервые показана область зимовки, а прикаспийская, волго-уральская и предуральская группировки зимуют в Судане и прилегающих районах Эфиопии и Эритреи. Красавки из азиатской части ареала, за исключением зауральской группировки, совершают кольцевую миграцию. Осенью они используют шесть основных миграционных потоков, прибывая в Северо-Западную Индию с севера, северо-востока и востока, а весной сначала летят узким фронтом до западной оконечности Западного Тянь-Шаня, а затем разлетаются веером в северном, северо-восточном и восточном направлениях. Для каждого миграционного потока выявлены ключевые территории на путях пролета и зимовках. Осенняя миграция проходит в сжатые сроки. При общей протяженности пролетных путей от 2170 до 5600 км, дистанция транзитных перелетов варьирует от 1900 до 4600 км, а длительность от 7 до 13 дней. Весенний пролет у взрослых особей более растянутый, чем осенний, с более короткими дневными перелетами и более длительным отдыхом на транзитных остановках. Некоторые молодые особи весной возвращаются на места рождения с родителями, другие совершают транзитный перелет до первых миграционных остановок, расположенных в пределах миграционного потока. Часть из них остается на этих территориях все лето, остальные постепенно перемещаются, прибывая к местам рождения на 1–1.5 месяца позже родителей. Молодые птицы из Забайкалья и, возможно, Алтая и Хакасии, совершают два транзитных перелета с длительным отдыхом в середине пути.
Красавка (Anthropoides virgo (Linneaus 1758)) – широко распространенный вид. Гнездовую часть ареала с середины прошлого века рассматривали как сплошную полосу, тянущуюся по степной и полупустынной зонам Евразии от Украины до Северо-Восточного Китая (Судиловская, 1951; Johnsgard, 1983; Флинт, 1987; Meine, Archibald, 1996). В середине 20 в. еще существовали изолированные популяции в Северной Африке и Турции, которые, вероятно, исчезли к концу века (Ильяшенко, Ильяшенко, 2011; Akarsu et al., 2013). В настоящее время в европейской части ареала выделяют азово-черноморскую (Северное Приазовье, Приазовская возвышенность, Сиваш, Крымский и Таманский полуострова), средне-донскую (Калачская излучина Дона в Волгоградской обл.), прикаспийскую (Восточное Предкавказье и Прикаспийская низменность) и волго-уральскую (Волго-Уральское междуречье) гнездовые группировки (Андрющенко, 1997, 2015; Андрющенко и др., 2008; Белик, 1999; Белик и др., 2011; Ильяшенко, 2018; Ilyashenko, 2019). Азиатская часть ареала условно разделена на казахстанско-среднеазиатскую (Кыргызстан, Казахстан и районы России, граничащие с Казахстаном) и восточноазиатскую (юг Восточной Сибири, Забайкалье, Монголия и Северо-Восточный Китай) (Ильяшенко, 2018; Ilyashenko, 2019). Эта часть ареала также фрагментирована, однако детальные исследования и картирование для выявления гнездовых группировок здесь не проводили.
В представленной работе на основании данных мечения дополнительно выделены следующие группировки: предуральская (запад Оренбургской обл.) в европейской части ареала и зауральская (восток Оренбургской и Актюбинской областей), хакасская (Республика Хакасия), алтайская (Республика Алтай), восточно-казахстанская (Восточно-Казахстанская обл. и восток Алматинской обл.), тянь-шаньская (горная часть Алматинской обл.) и забайкальская (юго-восток Забайкальского края) в азиатской части.
Известны две основные области зимовки – в Северо-Восточной Африке, куда летят журавли из европейской части ареала, и на п-ове Индостан, где зимуют журавли из казахстанско-среднеазиатской и восточноазиатской частей ареала (Судиловская, 1951; Johnsgard, 1983; Флинт, 1987; Urban, Gichuki, 1991; Meine, Archibald, 1996; Ilyashenko, 2019). Случаи зимовки отмечены в Восточном Китае (Ma, Ma, 2001).
История мечения журавлей началась с красавки, впервые в мире окольцованной в 1892 г. в Аскании-Нова (Херсонская обл., Украина). Эта особь была добыта в ноябре того же года в бассейне среднего Нила в Судане в районе г. Дангола (Андрющенко и др., 2006; Забашта, 2020). С началом использования цветных колец в европейской части ареала с 1986 г. красавок метили в Крыму и на Приазовской возвышенности, а в 1989 г. – в Калмыкии. За этот период получена информация о встрече на Кипре двух окольцованных в Крыму птиц (Андрющенко и др., 2006). В азиатской части ареала стандартными металлическими кольцами метили в Казахстане, где к 2017 г. окольцовано 238 молодых и два взрослых журавля, однако возвратов не получено. Не встречены и особи, помеченные в Индии (Mundkur, 1992). С 1990-х гг. цветными кольцами метили красавок в Забайкалье на Торейских озерах. От одной из них, окольцованной в 1990 г., получен возврат в 1992 г. из Индии, штат Гуджарат (Mundkur, 1992). Мечение цветными кольцами продолжается в Монголии и Китае; меченых там особей регулярно встречают в Индии (Suresh Kumar, Dau Lal Bohra, личн. сообщ.).
В 1995 г. впервые проведено мечение спутниковыми передатчиками в азиатской части ареала красавки в России в Забайкалье, в Западной Монголии и Юго-Восточном Казахстане. Оно позволило проследить весь путь осенней миграции четырех особей – одной из Казахстана и трех из Западной Монголии и отдельные участки путей миграции одной особи из Западной Монголии и трех особей из Забайкалья (Kanai et al., 2000).
В настоящее время активно ведется мечение передатчиками GPS-GSM в Забайкалье и Монголии, результаты которого находятся в обработке (Н. Батбаяр, личн. сообщ.), а также в Китае, где прослежен весь путь миграции трех птиц, помеченных на плато Ордос (Ordos) в автономном районе Внутренняя Монголия (Guo, He, 2017).
Цель нашей работы – уточнение маршрутов миграций, выявление ключевых территорий в предмиграционный период, на путях пролета и зимовках красавок, помеченных в разных географических районах гнездовой части ареала.
МАТЕРИАЛЫ И МЕТОДЫ
Отлов и мечение. Полевые работы по отлову и мечению красавки проведены в 2017–2020 гг. на территории Украины, России и Казахстана.
Для того чтобы отловить нелетных птенцов, к ним подъезжали на автомобиле как можно ближе и затем догоняли. Взрослые особи и молодая трехмесячная особь отловлены с использованием транквилизатора альфа-хлоралозы на гнездовых участках и на месте предмиграционного скопления. Отлов и мечение проведены по разрешениям соответствующих государственных природоохранных органов.
Птенцов в возрасте от 15 до 25 дней метили цветными спиральными кольцами, в возрасте от 25 до 35 дней – стандартными металлическими и цветными пластиковыми кольцами с цифровым или буквенно-цифровым кодом. Птенцов старше 35 дней и взрослых особей, кроме того, метили GPS-GSM передатчиками с солнечными батареями. 127 особей помечено передатчиками производства университета г. Констанца (Германия), прикрепленными к двум ножным цветным пластиковым кольцам ELSA (Германия). Общий вес передатчика и колец составлял 30 г. Передатчики помещали на голень правой ноги, цветные пластиковые кольца – на голень левой. Один взрослый журавль на Тянь-Шане GPS-GSM снабжен передатчиком “Lego” (“Druid”, Китай), прикрепленным к спине (рюкзачковый тип), и помечен желтым немецким пластиковым кольцом ELSA, надетым на левую голень.
Всего в 2017–2020 гг. окольцовано 196 красавок. Из них передатчиками помечены 128 особей, включая 12 взрослых, одна трехмесячная птица и 115 нелетных птенцов в возрасте от 35 до 50 дней. Функционировать начали 104 передатчика, из них 16 продолжали работать до ноября 2020 г. (время завершения работы над статьей). Поступление сигналов от большинства передатчиков было недостаточно регулярным.
Подробная информация о мечении (координаты, номера передатчиков, цвет и номера колец) опубликована ранее (Ильяшенко и др., 2018, 2020).
Анализ данных и терминология. Слежение за журавлями, помеченными передатчиками, вели на сайте www.movebank.org в период с 2017 по 2020 гг. При обработке данных использованы программы Microsoft Office Excel и Google Earth Pro. Биотоп локации определяли в программе Google Earth Pro.
Для анализа данных осенней миграции в качестве количественной единицы рассматривали молодую птицу в возрасте одного-двух лет, взрослую особь или семью, в которой помечены родитель или один-два птенца, так как в этот период семьи держатся вместе.
При анализе данных с мест зимовки, на весенней миграции или во время летнего пребывания в качестве количественной единицы рассматривали особь, так как период отделения птенцов от родителей для большинства семей начинается в зимний период и заканчивается весной, как правило, до отлета с зимовок.
При описании миграции мы, используя терминологию Гаврилова (1979), впервые для журавлей разделили ее на два этапа – трофический и транзитный, имеющих различные значения. Во время трофического этапа мигранты накапливают энергетические ресурсы на определенных, часто традиционных территориях, включающих места трофических предмиграционных скоплений и трофические миграционные остановки. Во время перемещений между трофическими территориями в начальный период этого этапа журавли могут отклоняться от основного направления миграции, а к концу этапа двигаться в направлении мест зимовки. Они проводят на трофических территориях не менее четырех дней – минимум, необходимый для восполнения энергетических ресурсов, что описано для серого журавля (Grus grus) (Hedenström, 2008). Во время транзитного этапа основной задачей птиц является быстрое достижение мест зимовок или летнего пребывания. Этот этап включает перелет на большой высоте по кратчайшему пути и кратковременные транзитные остановки на 1–2 дня, без существенного восполнения энергетических затрат.
Основываясь на данной терминологии, для указанных выше количественных единиц оценена длительность транзитного этапа осенней и весенней миграций. Длительность трофического этапа осенней миграции не определена, так как в разных географических районах и у различных социальных групп (птенцов, их родителей и неполовозрелых особей) он начинается в разное время с началом интенсивной кормежки, которое можно зафиксировать лишь при проведении специальных суточных наблюдений. Длительность трофического этапа весенней миграции также не определена из-за нерегулярного поступления сигналов, за исключением трех птиц прикаспийской гнездовой группировки, одной – зауральской и одной – забайкальской.
Дистанцию миграционного пути рассчитывали, измеряя расстояния между локациями передатчика. Для определения общей дистанции суммировали расстояния между трофическими территориями на пути пролета и расстояние транзитного перелета. При этом учитывали, что осенью транзитный перелет для одних журавлей начинался с места трофического предмиграционного скопления, для других – с места последней трофической миграционной остановки и для всех птиц заканчивался в области зимовки на территории, где они держались более четырех дней. Расстояние весенней транзитной миграции рассчитывали от места зимовки до места гнездования для взрослых особей, а для неполовозрелых птиц – до первой длительной трофической миграционной остановки. Местные перемещения в пределах трофических территорий в протяженность общего миграционного пути не включали.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Маршруты миграции журавлей разных гнездовых группировок представлены на рис. 1. Длительность и дистанция осеннего пролета даны в табл. 1 для всех гнездовых группировок за исключением хакасской из-за недостатка данных. Длительность и дистанция весенней миграции отражены в табл. 2.
Рис. 1.
Осенние и весенние миграционные маршруты красавки из разных частей гнездового ареала: А – азово-черноморской; B – прикаспийской, волго-уральской и предуральской; Ca, Cb – зауральской; D – хакасской; E – алтайской; F – восточно-казахстанской; G – тянь-шаньской; H – забайкальской; 1 – пути осенней миграции, 2 – пути весенней миграции, 3 – весенние транзитные остановки, 4 – весенние трофические миграционные остановки, 5 – осенние транзитные остановки, 6 – осенние трофические предмиграционные скопления и миграционные остановки, 7 – наиболее значимые территории во время осенней миграции, 8 – осенние и весенние трофические миграционные остановки, 9 – наиболее значимые территории во время осенней и весенней миграций, 10 – ключевые места зимовки, 11 – ареал красавки (Ilyashenko, 2019).
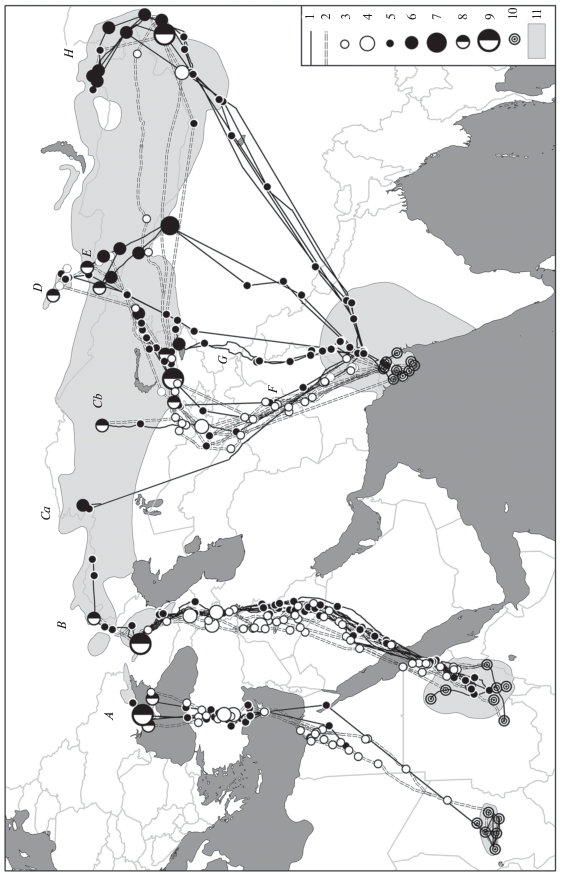
Таблица 1.
Длительность и дистанция осеннего пролетного пути красавок разных гнездовых группировок
| Гнездовая группировка (n) | Длительность и дистанция осеннего пролетного пути | ||||
|---|---|---|---|---|---|
| Трофический этап | Транзитный этап | Общая дистанция, км | |||
| дистанция, км | длительность, дней | дистанция, км | всего | над преградами (% от общей дистанции) | |
| Азово-черноморская (n = 4) | 0–130 | $\frac{{8{\kern 1pt} - {\kern 1pt} 13}}{{11}}$ | $\frac{{3480{\kern 1pt} - {\kern 1pt} 4070}}{{3775}}$ | $\frac{{3480{\kern 1pt} - {\kern 1pt} 4940}}{{3764}}$ | 2900–3000 (80%) |
| Прикаспийская и волго-уральская (n = 8) | 0–570 | $\frac{{8{\kern 1pt} - {\kern 1pt} 13}}{{10}}$ | $\frac{{3300{\kern 1pt} - {\kern 1pt} 4700}}{{4000}}$ | $\frac{{3300{\kern 1pt} - {\kern 1pt} 4700}}{{4000}}$ | 1000–1150 (30%) |
| Зауральская (n = 3) | 0 | $\frac{{6{\kern 1pt} - {\kern 1pt} 8}}{7}$ | 2800–3200* | 2800–3200* | 1800 (57%) |
| Хакасская (n = 2) | Нет данных | Нет данных | Нет данных | $\frac{{3700{\kern 1pt} - {\kern 1pt} 3900}}{{3800}}$ | 1500 (45%) |
| Алтайская (n = 4) | $\frac{{800{\kern 1pt} - {\kern 1pt} 900}}{{850}}$ | $\frac{{7{\kern 1pt} - {\kern 1pt} 9}}{8}$ | 2650 | $\frac{{3435{\kern 1pt} - {\kern 1pt} 3585}}{{3510}}$ | 2700 (90%) |
| Восточно-казахстанская (n = 2) | $\frac{{600{\kern 1pt} - {\kern 1pt} 800}}{{700}}$ | 8 | 2300 | $\frac{{2900{\kern 1pt} - {\kern 1pt} 3100}}{{3000}}$ | 1100 (37%) |
| Тянь-шаньская (n = 1) | 300 | 7 | 1900 | 2200 | 1000 (40%) |
| Забайкальская (n = 5) | $\frac{{1000{\kern 1pt} - {\kern 1pt} 1600}}{{1120}}$ | $\frac{{10{\kern 1pt} - {\kern 1pt} 17}}{{13}}$ | $\frac{{4000{\kern 1pt} - {\kern 1pt} 4600}}{{4480}}$ | $\frac{{5000{\kern 1pt} - {\kern 1pt} 6200}}{{5600}}$ | 2250 (50%) |
Примечания. Над чертой – округленные минимальные и максимальные значения, под чертой – среднее арифметическое; n – количественная единица (семья, взрослая или неполовозрелая особь, совершившие миграцию в течение одного и более сезонов). * Среднее значение не указано, так как транзитный перелет в 2018 и 2019 гг. начинался с разных географических территорий.
Таблица 2.
Длительность и дистанция весеннего пролетного пути красавок разных гнездовых группировок
| Гнездовая группировкa | Длительность и дистанция весеннего пролетного пути | ||||||
|---|---|---|---|---|---|---|---|
| Транзитный этап | Трофический этап | Общая миграция | |||||
| длитель-ность, дней | дистанция, км | длитель-ность, дней | дистанция, км |
длитель-ность, дней | дистанция, км |
над преградами (% от общей дистанции) | |
| Азово- черноморская | 23 (n = 1) |
$\frac{{4000{\kern 1pt} - {\kern 1pt} 4200}}{{4100}}$ (n = 2) |
0 | 0 | 23 | $\frac{{4000{\kern 1pt} - {\kern 1pt} 4200}}{{4100}}$ (n = 7) |
3000 (75%) |
| $\frac{{3{\kern 1pt} - {\kern 1pt} 12}}{7}$ (n = 7) |
$\frac{{2600{\kern 1pt} - {\kern 1pt} 3400}}{{3100}}$ (n = 8) |
Нет данных | $\frac{{850{\kern 1pt} - {\kern 1pt} 1000}}{{930}}$ (n = 5) |
Нет данных | |||
| Прикаспий-ская и волго-уральская | $\frac{{15{\kern 1pt} - {\kern 1pt} 25}}{{20}}$ (n = 5) |
$\frac{{3300{\kern 1pt} - {\kern 1pt} 4700}}{{4000}}$ (n = 5) |
0 | 0 | $\frac{{15{\kern 1pt} - {\kern 1pt} 25}}{{20}}$ (n = 5) |
$\frac{{3300{\kern 1pt} - {\kern 1pt} 4700}}{{3900}}$ (n = 11) |
1000 (25%) |
| $\frac{{9{\kern 1pt} - {\kern 1pt} 17}}{{11.5}}$ (n = 6) |
$\frac{{2700{\kern 1pt} - {\kern 1pt} 3400}}{{2900}}$ (n = 6) |
$\frac{{23{\kern 1pt} - {\kern 1pt} 38}}{{27}}$ (n = 3) |
$\frac{{280{\kern 1pt} - {\kern 1pt} 1380}}{{835}}$ (n = 4) |
Нет данных | |||
| Зауральская | $\frac{{4{\kern 1pt} - {\kern 1pt} 13}}{8}$ (n = 2) |
1200 (n = 1) |
127 (n = 1) |
1050 (n = 1) |
Нет данных | 2250 (n = 1) |
1000 (45%) |
| Хакасская | Нет данных | $\frac{{2100{\kern 1pt} - {\kern 1pt} 2500}}{{2300}}$ (n = 2) |
Нет данных | $\frac{{2700{\kern 1pt} - {\kern 1pt} 2800}}{{2750}}$ (n = 2) |
Нет данных | $\frac{{4800{\kern 1pt} - {\kern 1pt} 5300}}{{5050}}$ (n = 2) |
1400 (30%) |
| Алтайская | $\frac{{9{\kern 1pt} - {\kern 1pt} 11}}{{10}}$ (n = 2) |
$\frac{{2100{\kern 1pt} - {\kern 1pt} 2500}}{{2400}}$ (n = 3) |
Нет данных | $\frac{{2000{\kern 1pt} - {\kern 1pt} 2200}}{{1900}}$ (n = 3) |
Нет данных | $\frac{{4100{\kern 1pt} - {\kern 1pt} 4300}}{{4200}}$ (n = 3) |
1500 (35%) |
| Восточно-казахстанская | 7 (n = 1) |
$\frac{{2100{\kern 1pt} - {\kern 1pt} 2500}}{{2400}}$ (n = 2) |
Нет данных | $\frac{{1300{\kern 1pt} - {\kern 1pt} 1600}}{{1450}}$ (n = 3) |
Нет данных | $\frac{{3400{\kern 1pt} - {\kern 1pt} 3800}}{{3600}}$ (n = 2) |
1000–1200 (27–33%) |
| Тянь- шаньская | 16 (n = 2) |
$\frac{{3150{\kern 1pt} - {\kern 1pt} 3500}}{{3500}}$ (n = 2) |
0 | 0 | 15–16 (n = 2) |
$\frac{{3500{\kern 1pt} - {\kern 1pt} 3900}}{{3700}}$ (n = 2) |
1100 (30%) |
| Забайкальская | 9 + 7* (n = 1) |
3000 + 4300* (n = 1) |
29 (n = 1) |
700 (n = 1) |
45 (n = 1) |
8000 (n = 1) |
2000 (25%) |
Примечания. Над чертой – округленные минимальные и максимальные значения, под чертой – среднее арифметическое; n – количественная единица (семья, взрослая или неполовозрелая особь, совершившие миграцию в течение одного и более сезонов). * Среднее значение не указано, так как данные о длительности и дистанции двух транзитных перелетов имеются для одной птицы.
Европейская часть ареала
Азово-черноморская гнездовая группировка
Для данной группировки известно наиболее западное место крупных летних и предмиграционных скоплений на заливе Сиваш, где мониторинг ведется более 30 лет (Андрющенко, 1997; Андрющенко, Шевцов, 1998; Андрющенко, Горлов, 1999; Андрющенко и др., 2008). Предполагалось, что перед отлетом здесь собираются красавки из Приазовья и Крыма, образуя скопления более тысячи особей, а также, возможно, журавли из более восточных частей ареала (Андрющенко, 1997; Андрющенко, Шевцов, 1998). Летние и предмиграционные скопления численностью до 800 особей известны также на западе Крымского п-ова на Тарханкутской возвышенности на оз. Джарылгач и на юге Керченского п-ова на озерах по периметру Опукского заповедника, где собирается до 200–300 особей (Андрющенко, Шевцов, 1998; Андрющенко, 2015).
Осенью перед пересечением Черного моря красавки летят над Крымскими горами (Костин, 1983; Андрющенко и др., 2006). Кроме пролёта через Чёрное море, незначительное число красавок мигрирует вдоль балканского побережья, останавливаясь в Северо-Западном Причерноморье – в Херсонской, Николаевской, Одесской областях Украины, а также в Румынии и Болгарии (Нанкинов, 2009; Андрющенко, 2015). К этой же ветви пролетного пути относятся встречи в Греции и на западном побережье Турции в проливе Босфор и Измире (Kasparek, 1988). Для Турции также отмечены встречи в период миграций в центре и на востоке страны (Kasparek, 1988; Akarsu et al., 2013). Пролет над Кипром известен по визуальным наблюдениям пролета и отдыха красавок на оз. Акротири (Akrotiri), где численность вида в 1957 г. оценена в 8.4 тыс. особей, а в августе 1971 г., при организации специального учета, отмечено минимум 1468 особей (Flint, 1971, 1972). Кроме того, на Кипре встречены две птицы, окольцованные цветными пластиковыми кольцами на Украине (Андрющенко и др., 2006). В Египте красавку относили к редкому мигрирующему и зимующему виду (Atta, 1996).
Предполагали, что область зимовки журавлей из европейской части ареала тянется полосой от Чада до Эфиопии через Судан. В Чаде красавки отмечены на озерах Чад, Иро и Фитри (Johnsgard, 1983; Urban, Gichuki, 1991; Meine, Archibald, 1996; Beilfuss et al., 2007). Причем одни авторы пишут о больших стаях на оз. Чад (Mackworth-Praed, Grant, 1970), другие отмечают единственную встречу группы из 24 особей в октябре 1991 г. (Scholte, 1996). Одна особь зарегистрирована в Северо-Восточной Нигерии (Urban, Gichuki, 1991). Информация о зимовке в Судане основана на добыче молодой красавки, помеченной в Аскании-Нова в 1892 г. (Андрющенко и др., 2006).
В 2018–2020 гг. проведено дистанционное слежение 24 молодых особей из 15 семей, помеченных на Сиваше, Приазовской и Тарханкутской возвышенностях, Керченском и Таманском полуостровах.
Место трофического предмиграционного скопления на заливе Сиваш в 2018–2020 гг. использовали восемь семей. Из них четыре семьи гнездились на Тарханкутской возвышенности, расположенной в 100–160 км южнее, две – на Сиваше и две семьи – на Приазовской возвышенности. Стартуя с Сиваша, две семьи полетели вдоль западного побережья через озера Джарылгач и Сасык-Сиваш. Из-за нерегулярного поступления сигналов осталось неясным, как пересекали Крым остальные семьи.
Одна семья, гнездящаяся на Тарханкутской возвышенности, начала миграцию с оз. Джарылгач. Две семьи стартовали с юга Керченского п-ова. Одна из них, гнездящаяся на севере полуострова, до отлета держалась на оз. Тобечикское. Вторая семья, гнездившаяся на Таманском п-ове в Краснодарском крае, в предмиграционный период кормилась на полях, прилегающих к гнездовой территории, и ночевала на оз. Маркитанское, расположенном в двух-трех километрах от нее. За 10 дней до отлета на зимовку она переместилась севернее на водохранилище Юзмак на юг Керченского п-ова.
Осенняя миграция семей (n = 8) в 2018–2020 гг. начиналась в период с 17 по 28.08. Данные об ее длительности и дистанции представлены в табл. 1.
Маршрут осенней миграции семи семей проходил над Крымом, центральной частью Турции, над Кипром, Египтом, юго-восточной частью Ливии и Чадом (рис. 1А). Они пересекли такие географические преграды как Крымские горы (30–50 км), Черное море (250–300 км), горный массив Таурус (Taurus) (150 км) на юге Турции, Средиземное море (500 км), дельту Нила или Суэцкий канал и пустыню Сахара (2000 км). Данные дистанционного слежения позволили определить основной путь миграции, но нерегулярное поступление сигналов не позволило выявить все транзитные остановки. Сигнал над Кипром получен только для одной птицы, однако, по-видимому, над островом проходит основной пролетный путь (Flint, 1971, 1972).
Одна молодая птица отклонилась от общего курса. После пересечения Черного моря она ослабла и была поймана на побережье в турецкой провинции Зонгулдак (Zonguldak) (Ильяшенко и др., 2020а). Птицу осмотрели, не нашли никаких повреждений и через два дня выпустили. Она продолжила миграцию на восток вдоль побережья и достигла оз. Ладик (Ladik), известного места миграционной остановки серых журавлей (Burak Tatar, личн. сообщ.). Через 20 дней полетела в юго-западном направлении и прилетела сначала в Республику Нигер, а затем в Нигерию, откуда последние сигналы пришли из г. Даматуру (Damaturu) 15.11.2019. Это известное место зимовки в Северо-Восточной Нигерии (Urban, Gichuki, 1991).
14 молодых журавлей прилетели на зимовку в Чад в регион Батха (Batha), расположенный в зоне сахеля, в поросшей кустарником саванне. Сигналы еще от двух молодых птиц стали поступать только с этого места зимовки.
Выделено несколько ключевых участков, которые красавки использовали по определенной схеме в течение периода зимовки (табл. 3). После прибытия и до начала–середины октября они держались на низменности на севере и северо-западе региона. В середине октября – середине ноября часть птиц переместилась или в долину р. Бахр-эль-Газаль (Bahr El Gazal), вытекающей из оз. Чад, или в долину р. Батха, впадающей в оз. Фитри (Fitri), или на восток региона в долины рек, стекающих с горного массива Ваддаи (Ouddai), западнее пос. Битлин (Bitline). Этот участок расположен на юге охраняемого природного резервата Ouadi Rime-Ouadi Achim. С конца октября – середины ноября журавли (n = 16) переместились на основное место зимовки на заболоченном участке долины р. Батха, в месте слияния двух рек, между поселками Абзиуф (Absiuf) и Ам Сак (Am Sak). Здесь они держались 3.5–4 месяца до начала весенней миграции. Такое перераспределение в пределах области зимовки, охватывающей приблизительно 12.5 тыс. км2, связано, очевидно, с климатическими условиями. Сезон дождей приходится на май–октябрь (http:// www.restbee.ru/world/afrika/chad). Поэтому в сентябре и октябре красавки держались на низменности запада провинции или в долинах рек на востоке провинции, где в этот период года еще сохраняется вода. В ноябре они переместились на болото на р. Батха, которое, видимо, не пересыхает весь период зимовки. Они летали кормиться в саванну или на поля к западу – юго-западу от болота на расстояние 15–20 км.
Таблица 3.
Ключевые территории на миграционных путях и зимовках красавки
| Гнездовые группировки | Ключевые территории по пути миграции и на зимовках |
|---|---|
| Европейская часть ареала | |
| Азово-черноморская | Украина: Сиваш (Л, ТО, ТВ) |
| Крым: 1) оз. Джарылгач (Л, ТО), 2) озера на юге Керченского п-ова (Л, ТО) | |
| Турция: провинция Конья, бассейн оз. Туз (ТВ) | |
| Чад: 1) долина р. Бахр-эль-Газаль, бассейн оз. Чад (З); 2) долина р. Батха в бассейне оз. Фитри (З); 3) долины рек, стекающих с горного массива Ваддаи западнее пос. Битлин (З); 4) долина р. Барха между пос. Абзиуф и Ам Сак (З) | |
| Прикаспийская, волго-уральская, предуральская | Россия: Ростовская обл., Республика Калмыкия, Ставропольский край, долина р. Маныч (Л, ТО, ТВ) |
| Россия, Волгоградская, Саратовская обл., бассейн рек Торгун, Еруслан, Соленая Куба, (Л, ТО) | |
| Грузия: долины рек Иори и Алазани (ТВ) | |
| Азербайджан: Нахичеванская Автономная Республика, г. Карацуг, Араксинское вдхр. (ТВ) |
|
| Турция: провинция Ыгдыр, бассейн р. Аракс (ТВ) | |
| Ирак: п. Байджи, бассейн р. Тигр (ТВ) | |
| Судан: 1) бассейн Голубого Нила, междуречье Диндера и Аль Рахада (З); 2) долина Нила между гор. Бербер и Эль Тиккавин (З); 3) бассейн Белого Нила в районе гор. Эль-Обейд (3); долина р. Шагр эн Нил, 100 км восточнее Хартума | |
| Судан, Эритрея, Эфиопия: долина р. Текезе, в районе городов Омхайер (Эритрея) и Химора (Эфиопия) | |
| Азиатская часть ареала | |
| Зауральская | Россия, Казахстан: Оренбургская и Костанайская обл., озера на Тургайском плато (ТО) |
| Хакасская | Россия, Хакасия: 1) Южно-Минусинская котловина, уроч. Трехозерки (Л, ТО); 2) оз. Белё (ТО) |
| Китай: Синьцзян-Уйгурский автономный район, долина р. Эмель (ТВ) | |
| Монголия: Убсунурский аймак, котловина Больших озер (ТВ) | |
| Алтайская | Россия: Тува, долины рек Хемчик и Чадан (Л) |
| Монголия: 1) аймак Ховд, оз. Хар-Ус (Л, ТО); 2) Убсунурский аймак, котловина Больших озер (ТО); 3) Баян-Улэгейский аймак, бассейн р. Ховд (ТО, ТВ) | |
| Китай, Синьцзян-Уйгурский автономный район: 1) уезд Чингиль (ТО); 2) долина р. Эмель (ТВ); оз. Баркёль (ТО) | |
| Восточно-казахстанская | Китай, Синьцзян-Уйгурский автономный район: долина р. Эмель (Л, ТО, ТВ) |
| Казахстан: 1) Капчагайское вдхр., долина р. Или (ТО, ТВ); 2) Ташуткольское вдхр., долина р. Чу (ТО, ТВ); 3) Терс-Ащибулакское вдхр., долина р. Терс (ТО, ТВ) | |
| Тянь-шаньская | Кыргызстан: оз. Иссык-Куль (ТО, ТВ) |
| Забайкальская | Россия: Забайкальский край, котловина Торейских озер (Л, ТО) |
| Монголия: аймак Дорнод, долина р. Халхин-Гол (ТO) | |
| Китай, Внутренняя Монголия: 1) пос. Хулун-Буир, долина р. Хайлар (ТО); 2) вдхр. Вулагай (ТО); 3) Шилин-Гол, оз. Дали (Л, ТО, ТВ) | |
| Все группировки азиатской части ареала | Узбекистан: оз. Айдаркуль (ТВ) |
| Казахстан: 1) Шимкентская обл., Коксарайское вдхр., пос. Баиркум, долина р. Сыр-Дарьи (ТВ); 2) Шимкентская обл., вдхр. Капшагай, пос. Бирлик (ТВ); 3) Жамбыльская обл., Терс-Ащибулакское вдхр., долина р. Терс (ТО, ТВ); 4) Жамбыльская обл., Ташуткольское вдхр., долина р. Чу (ТО, ТВ); 5) Алматинская обл., Капчагайское вдхр., долина р. Или (ТВ) | |
| Индия, штат Раджастан: 1) окрестности Джодхпура (З); 2) Кичан (З); 3) Ланкарасар и Биканер (3) | |
| Индия, штат Гуджарат: 1) побережье Аравийского моря с наибольшей концентрацией в районе Мияни и Порбандор (З); 2) солончак Большой Качский Ранн (З); северное и южное побережье Качского залива (З) | |
Примечания. Л – летние скопления, ТО – осенние места трофических предмиграционных скоплений и трофические осенние миграционные остановки, ТВ – трофические весенние миграционные остановки, З – места зимовки. Жирный шрифт – отправные пункты транзитной миграции (за исключением зауральской группировки, где отправной пункт не выявлен).
Весенняя миграция в 2019 и 2020 гг. начиналась в период с 07.03 по 20.03 (n = 14), наиболее интенсивно 10–12.03. Пролет проходил по тому же маршруту, что и осенью. Данные о длительности и дистанции представлены в табл. 2.
Восемь молодых особей совершили транзитный перелет до первой трофической миграционной остановки в Турции. Некоторые до прибытия на территорию Турции (n = 4) останавливались на 2–5 дней в Египте в западной части дельты Нила на границе с Сахарой или на южном побережье Средиземного моря. В Турции молодые особи держались до 40 дней в провинции Конья (Konya), главным образом, севернее Чумры (Chumra), и в бассейне оз. Туз (Tuz). В 2019 г. два журавля, помеченные на Тарханкутской возвышенности, прилетели на оз. Джарылгач 24.05 и 9.06. Затем переместились на Сиваш 14 и 23.07 и держались там до отлета на зимовку. Три особи вернулись на места рождения на Керченском и Таманском полуостровах.
Две из 16 молодых красавок не использовали трофическую миграционную остановку в Турции. Одна прибыла к месту рождения на оз. Маркитанское в Краснодарском крае 23.03.2019. Она встречена 24.03.2019 в группе из трех птиц, вероятно, с родителями (Ильяшенко и др., 2020б). Сигналы от второй птицы прекратились после пересечения территории Турции.
Таким образом, наши данные подтвердили значимость залива Сиваш как основного места трофического предмиграционного и миграционного скопления. При этом выявлена некоторая обособленность журавлей, гнездящихся на Керченском и Таманском полуостровах. В отличие от журавлей с Тарханкутской возвышенности, они не переместились перед началом миграции на Сиваш, а держались на юге Керченского п-ова до отлета на зимовку.
Впервые показан весь пролетный путь, который осенью и весной проходил по одному маршруту. Ни один из прослеживаемых журавлей не использовал путь миграции вдоль западного или восточного побережий Черного моря, что можно объяснить ограниченным числом меченых птиц. Выявлены ключевые места зимовки в Чаде в регионе Батха и на реках бассейнов озер Чад и Фитри. Определены места трофических весенних миграционных остановок в Центральной Турции в провинции Конья, включая бассейн оз. Туз, где молодые птицы могут оставаться до начала лета. Некоторые неполовозрелые особи держатся в Турции все лето и начинают оттуда осеннюю миграцию.
Численность азово-черноморской гнездовой группировки оценена в 2000 особей, из которых 1500–1700 перед отлетом концентрируются на Сиваше и 300–500 особей – на Керченском п-ове. Предположение, что на Сиваш перед отлетом могут слетаться журавли из средне-донской гнездовой группировки (Андрющенко, Шевцов, 1998; Андрющенко, Горлов, 1999), не подтверждено, так как в июне 2019 г. в местах ее обитания красавки не обнаружены (Ильяшенко и др., 2020в). Однако эта группировка, численность которой в 1990-е гг. оценена в 200–300 пар (Белик и др., 2011), а также недоучтенные птицы в Северо-Восточном Приазовье (восток Донецкой и запад Ростовской областей) могли повлиять на общую оценку численности всей азово-черноморской группировки.
Прикаспийская и волго-уральская гнездовые группировки
Для журавлей прикаспийской группировки известно традиционное крупное летнее и предмиграционное скопление в долине р. Маныч на стыке границ Республики Калмыкия, Ставропольского края и Ростовской области (Букреева, 2003; Белик и др., 2011). В Калмыкии на озерах и прудах, главным образом, в Сарпинской низменности отмечены летние скопления, которые к августу укрупняются за счет присоединения семей с птенцами, а к концу августа перемещаются в долину Маныча. Предполагали, что перед отлетом на Маныче собираются красавки не только со всей Калмыкии, но и прилегающих районов Ставропольского края и Ростовской области, образуя стаи общей численностью до 40–45 тыс. особей (Букреева, 2003). Для журавлей волго-уральской группировки известны предмиграционные скопления в Волгоградском Заволжье у пос. Золотари численностью до 800 особей (Ильяшенко, 2013) и в Саратовском Заволжье в долинах рек Большой и Малый Узень, где учтено 173 особи (Завьялов и др., 2003). В Западном Казахстане отмечено скопление до тысячи особей в 1938 г. в районе оз. Рыбный Сакрыл, а в конце 1980-х такие скопления встречены на Битикском водохранилище (Шевченко и др., 1993).
Красавка считается редким мигрантом в Западной Грузии и обычным в Восточной Грузии (Abuladze, 2018). Предполагалось, что через Грузию летят красавки из Украины и Калмыкии (Abuladze, 1995). В Восточной Грузии численность мигрирующих птиц оценена в 4–4.5 тыс. осенью и 2.7–3 тыс. весной; известны места кратковременных остановок, главным образом, в долинах рек Иори и Алазани (Abuladze, 2018). В Армении отмечены весенне-осенние и летние встречи (Ляйстер, Соснин, 1942; Даль, 1954; Саркисян, 2011). В Азербайджане пролет идет, главным образом, через западную часть республики по линии Загатала – Самух – Гянджа – Нахичевань (Султанов и др., 2011). В Иране красавки летят через провинции Восточный и Западный Азербайджан (Ranaghad, Ebrahimi, 2007). Известно два места транзитных остановок к западу от оз. Урмия у г. Салмас (Salmas) (270–400 особей) и на границе с Азербайджаном в долине р. Аракс на Араксинском водохранилище (300 особей). Кроме того, около 1000 особей оставались на зимовку вместе с серыми журавлями в провинции Восточный Азербайджан в Иране (Ashtiani, 1999). В Ираке отмечена зимовка вблизи г. Мосул в декабре 1922 г. (Meinertzhagen, 1924 по: Kasparek, 1988). В Саудовской Аравии на участке от г. Джедда (Jedda), расположенном на восточном побережье Красного моря, до г. Хаиль (Ha’il) в местах регулярных наблюдений пролета красавок весной 1992 и 1993 гг. проведены специальные учеты на нескольких обзорных пунктах с привлечением волонтеров (Newton, Simmens, 1993; Newton 1996). Численность оценена в 4.5 и 6 тыс. соответственно, хотя предполагалось, что она занижена из-за неопытности учетчиков и из-за пролета некоторых стай на большой высоте. Ширина пролетного пути определена в 100 км (Newton, 1996).
Зимовка красавки в Судане на р. Белый Нил впервые описана Бергом (Berg, 1930). В 1986 г. определена область, включающая провинции Гезира, Хартум, Кассала и Голубой Нил с наибольшей концентрацией в бассейне Белого и Голубого Нила и р. Атбара (Atbara), а также на реках Диндер и Рахад, притоках Голубого Нила (Nikolas, 1987; Urban, Gichuki, 1991). В Эфиопии в национальном парке Кафта Шераро (Kafta Sheraro National Park) в марте 2009 г. учтено 21.5 тыс. красавок (Gebremedhin et al., 2009), однако зимой 2010/2011 гг. журавли здесь не обнаружены (Gunter Nowald, личн. сообщ., 2011). Область зимовки находится в зоне сахеля в типичных саваннах с разреженным древостоем, преимущественно из акаций (Jonhsgard, 1983). Количество осадков составляет от 100 до 600 мм в год, около 90% которых выпадает в течение двух–трех летних месяцев (http://www.gecont.ru/articles/geo/sudan.htm). Предполагалось, что в Судане зимуют птицы, гнездящиеся на Украине и в прикаспийской низменности к западу от р. Волга (Newton, 1996).
Журавлей, гнездящихся в Западном Казахстане в Волго-Уральском междуречье, относили к использующим Урало-Эмбинский миграционный поток. Это наиболее западный из пяти миграционных потоков, проходящих через территорию Казахстана (Гаврилов, 1979). Он отделен от соседнего Арало-Ишимского потока горами Мугоджарами, простирающимися от Уральских гор с севера на юг.
Это предположение основано на встрече небольших мигрирующих стай в Волго-Уральских песках и в бассейне Верхней Эмбы на юго-востоке Атырауской (Гурьевской) обл. (Климов и др., 1991), а также на наблюдениях за весенней миграцией на юге Казахстана, где из 2854 учтенных особей 0.4% летело в западном направлении (Губин, Скляренко, 1991). Однако точные пути пролета и места зимовки журавлей этой группировки оставались неизвестными (Gavrilov, Van der Ven, 2004).
В 2017–2020 гг. прослежено 50 особей (включая семь взрослых птиц), помеченных в Восточном Предкавказье, Прикаспийской низменности, Волго-Уральском междуречье и в Предуралье. У шести особей сигналы передатчиков поступали около двух лет, что позволило проследить летние перемещения молодых птиц после первой и второй зимовок. Для журавлей прикаспийской и волго-уральской группировок получены наиболее полные данные за счет наибольшего числа помеченных птиц и большего числа передатчиков (n = 8) с регулярно поступавшими сигналами.
Все красавки (семьи с мечеными птенцами, взрослые и неполовозрелые особи) перед началом транзитного этапа миграции собирались в долине р. Маныч. Гнездовые участки семей располагались от места сбора на расстоянии от нескольких сот метров до 1000 км. Журавли с более дальних гнездовий из Заволжья, Западного Казахстана и Предуралья перед прилетом на Маныч использовали другие места трофических предмиграционных скоплений, главным образом, обширную территорию сельскохозяйственных полей в бассейнах рек Еруслан, Торгун, Соленая Куба и Малый Узень на стыке границ Волгоградской, Саратовской областей России и Западно-Казахстанской области Казахстана. С этих территорий одни особи начинали перемещаться в долину Маныча в середине июля–начале августа, другие, уже начав миграцию, прилетали в середине–конце августа за 10–15 дней до начала транзитного перелета.
Осеннюю миграцию красавки начинали в 2017 г. с 1 по 8.09 (n = 8), в 2018 г. – c 4 по 13.09 (n = 9), в 2019 г. – c 23.06 по 11.09 (n = 13). Общий период начала осенней миграции в 2017–2019 гг. длился с 23.08 по 13.09 (n = 29). Начало этого периода с 23.08 по 4.09, окончание – с 8.09 по 13.09. Более растянутый, чем у журавлей азово-черноморской группировки, период начала осеннего пролета может быть связан с большим числом прослеживаемых особей. Данные о длительности и дистанции осенней миграции прикаспийской и волго-уральской группировок представлены в табл. 1.
Маршрут журавлей прикаспийской и волго-уральской группировок проходил над Ставропольским краем, Дагестаном, Чечней, Восточной Грузией, Западным Азербайджаном, Арменией, Ираном, Ираком, Саудовской Аравией к местам зимовки в Северо-Восточной Африке (рис. 1В). По пути они пересекли такие географические преграды как Кавказские горы (Большой и Малый Кавказ) (около 250 км), Сирийскую пустыню и пустыню Большой Нефуд в Ираке и Саудовской Аравии (450–600 км) и Красное море (около 300 км). Первую треть пути от Маныча до оз. Урмия в Иране журавли летели узким фронтом, шириной 70 км. Далее над пустынями пролетный коридор расширился до 150–250 км. Над пустыней Большой Нефуд они летели между городами Хаиль (Hail) и Бурайдах (Buraydah), расположенными на расстоянии 250 км друг от друга, а перед пересечением Красного моря – между городами Медина (Al-Madinah) и Джедда (Jeddah), лежащими в 200 км друг от друга.
Осенний транзитный пролет (n = 7) проходил с транзитными остановками, главным образом, на ночной отдых. В первой трети пути красавки останавливались на одну, реже на две ночи в традиционных местах: 1) оз. Малый Маныч и его окрестности на севере Дагестана, 2) долины рек Алазани, Иори и Кура в Восточной Грузии и Западном Азербайджане после пересечения Главного Кавказского хребта, 3) долина р. Аракс и Араксинское водохранилище на границе Ирана и Нахичеванской Автономной Республики Азербайджана и 4) оз. Урмия в провинции Западный Азербайджан в Иране. Только одна семья не сделала остановку в Дагестане, пролетев до Алазанской долины в Азербайджане более 600 км около 18 ч, в том числе в ночное время. Еще одна семья не остановилась на оз. Урмия, а пролетела от Араксинского водохранилища до р. Сируон (Sirwan) в Ираке. Над Арменией птицы летели над оз. Севан или восточнее него без остановки.
В Ираке и Саудовской Аравии при пересечении пустыни красавки делали транзитные остановки на случайно выбранных территориях, не приуроченных к сельскохозяйственным полям, водоемам или оазисам. После перелета через Красное море в направлении к г. Порт-Судан, 1–2 дня они отдыхали на западном побережье в 15–20 км севернее или южнее этого города, после чего продолжали полет вглубь страны.
Область зимовки в бассейне рек Нил, Белый и Голубой Нил и Атбара охватывает восемь провинций в Судане, северо-запад провинции Тыграй (Tigray) в Эфиопии и юго-запад провинции Гаш-Барка (Gash-Barka) в Эритрее. Она тянется от излучины р. Нил в районе Абу-Хамад (Abu Hamad) на севере до Эд Дамазин (Ed Damazin) на юге на протяжении 900 км в меридиональном направлении и от Эль Обейд (El Obeid) на западе до Химоры (Himora) на востоке протяженностью 700 км в широтном направлении.
По прибытии в Судан журавли держались в провинции Кассала в долине р. Атбара. С сентября по ноябрь они перемещались в пределах обширной территории в междуречье Голубого Нила и его притоков – Диндер (Dinder) и Рахад (Al Rahad) в пределах 200–300 км от Хартума до Сеннара, с наибольшей концентрацией в районе городов Вад Медани (Wad Medani) и Мунира (Munirah). В ноябре часть журавлей переместилась на север в долину Нила между городами Бербер (Berber) и Эт Тиккавин (Et Tikkawin), часть – на запад в провинцию Северный Кордофан в бассейн Белого Нила в район Эль Обейд (El Obeid) и часть – в восточном направлении в долину р. Шагр эн Нил (Shagr en Nile) в 100 км восточнее Хартума. Во второй половине зимы большая часть журавлей перемещалась на самый восточный участок в долину р. Текезе на стыке границ Судана, Эритреи и Эфиопии в районе городов Омхайер (Omhajer) в Эритрее и Химора (Himora) в Эфиопии, в том числе в Национальный парк Кафта Шераро (табл. 3).
Весенняя миграция в 2018–2020 гг. начиналась в период с 11 по 26.03 (n = 19) и для большинства красавок (n = 14) проходила по тому же пролетному пути, что и осенью. Данные по ее длительности и дистанции даны в табл. 2.
Пять особей (взрослая, двухлетняя и три годовалых) в Ираке отклонились от осеннего маршрута и пролетели западнее над восточной частью Турции.
Пять журавлей (три взрослых, один двухлетний и один годовалый) совершили транзитный перелет до мест гнездования без длительных трофических миграционных остановок. Их дневные перелеты были короче и число транзитных остановок – от 10 до 13 (n = 4), немного больше, чем во время осенней миграции. Остальные неполовозрелые красавки (n = 15) в 2018–2020 гг. пролетели транзитом до первых трофических весенних миграционных остановок, расположенных в Ираке, Турции, Грузии и Азербайджане (табл. 3), где держались от 19 до 34 дней, в среднем 25 дней (n = 5). Из четырех трофических территорий каждый из меченых журавлей использовал одну или две. Затем неполовозрелые журавли продолжили перемещаться по пролетному пути на места летних скоплений в долину р. Маныч (n = 8), в бассейны рек Торгун и Еруслан (n = 3) и на запад Калмыкии (n = 3).
Три журавля, помеченные в Калмыкии и Заволжье, держались в долине Маныча все лето до отлета на зимовку. Одна красавка, помеченная в Волгоградском Заволжье, лето после первой зимовки провела в междуречье Торгуна и Еруслана и прилетела на Маныч за 15 дней до отлета на зимовку. Три журавля в 2020 г., после второй зимовки, прилетели на места рождения в Заволжье и Западный Казахстан, затем переместились на место летнего и предмиграционного скопления в бассейны рек Торгун и Еруслан, а в середине августа прилетели на Маныч.
Из 30 прослеживаемых журавлей, использующих данный пролетный путь, один молодой журавль, помеченный в Западном Казахстане, совершил отличные от других осенние и весенние миграции. Зиму 2018/2019 гг. он провел в Судане в бассейне Нила, прилетев туда вместе с журавлями прикаспийской и волго-уральской группировок. Во время весенней миграции в 2019 г. пересек Красное море не в районе г. Порт Судан, как это делали птицы данных группировок, а пролетел вдоль западного побережья на север, перелетел через северную часть моря, затем мигрировал через Иорданию и Сирию в Турцию. Через центр Турции он долетел до южного побережья Черного моря и вернулся в провинцию Сивас, где провел все лето и откуда начал осеннюю миграцию 22.08.2019. На пути он пересек западную часть Сирии, Израиль, Суэцкий залив и Сахару и прибыл в провинцию Батха в Республике Чад, где зимуют журавли азово-черноморской гнездовой группировки. Весной 2020 г. он полетел по пути, используемом красавками этой группировки, и долетел до дельты Нила, где сигналы от передатчиков прекратились (рис. 2).
Рис. 2.
Пути осенней и весенней миграций и места зимовки в 2018/2019 гг. (А) и 2019–2020 гг. (В) неполовозрелого журавля, помеченного птенцом в Западном Казахстане. Условные обозначения как на рис. 1.
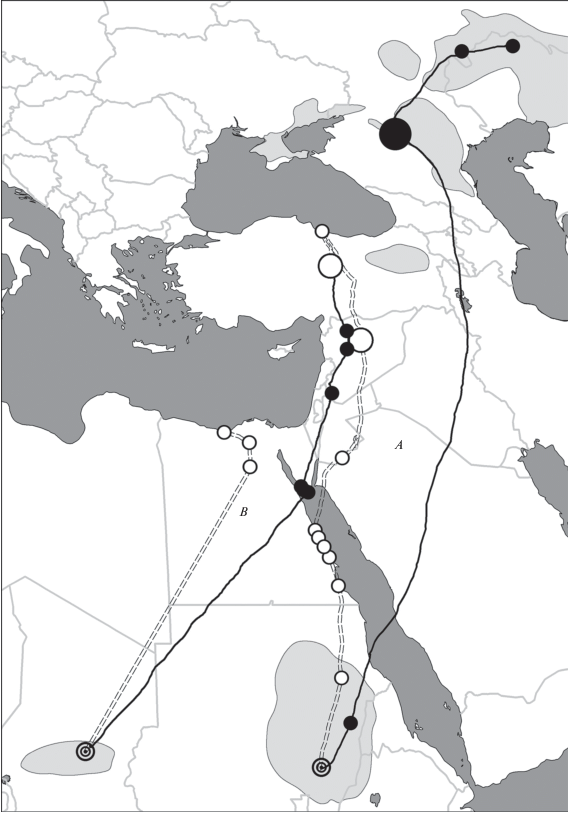
Таким образом, данные дистанционного слежения показали значимость долины р. Маныч не только для журавлей из Восточного Предкавказья и Прикаспия, но и из Волго-Уральского междуречья и Предуралья. Для красавок, которые гнездятся на расстоянии до 400 км от Маныча и держатся здесь до 1.5 месяцев, эта территория является местом летнего и трофического предмиграционного скоплений. Для журавлей с более дальних гнездовий – это место миграционной трофической остановки, где они пополняют энергетические ресурсы перед миграцией.
Все красавки из описываемого региона мигрируют на места зимовки в Судан и прилегающие районы Эфиопии и Эритреи. Осенью и весной они используют один и тот же пролетный путь, лишь некоторые птицы летят весной через Восточную Турцию. Также подтверждено, что в Саудовской Аравии на севере пустыни Большой Нефуд журавли традиционно мигрируют в районе городов Хаиль и Джедда, при этом ширина пролетного пути здесь больше, чем предполагалось. Это, в том числе, объясняет недоучет красавок во время весенней миграции в Саудовской Аравии (Newton, 1996).
Необходимо отметить, что при описании миграции журавлей в Турции предполагалось, что осенью красавки летят через центр страны, а весной – через ее восточную часть (Akarsu, 2013). Наши данные показали, что через центр Турции как осенью, так и весной летят журавли азово-черноморской группировки, а через восток страны весной летит часть журавлей из прикаспийской и волго-уральской группировок.
Выявлены новые места трофических и транзитных миграционных остановок, а для известных – долины рек Кура, Алазани и Иори, Араксинское водохранилище и оз. Урмия, показано их более широкое использование. По данным дистанционного слежения, красавки останавливались на оз. Урмия в пределах 50–60 км, хотя ранее было известно лишь одно место остановки (Ranaghad, Ebrahimi, 2007).
Определена более широкая, чем предполагалось, область зимовки в Северо-Восточной Африке. Выделено несколько ключевых территорий и определен характер перемещений между ними. Зимовка в Ираке и Иране не отмечена.
Численность журавлей, использующих второй миграционный поток, на основании литературных источников и собственных учетов территориальных пар и скоплений неполовозрелых особей оценена в 40–57 тыс. особей, включая прикаспийскую (30–40 тыс.) и волго-уральскую (15–17 тыс.) группировки (Букреева, 2003; Бидашко и др., 2006; Ильяшенко, 2018; Ilyashenko, 2019).
Азиатская часть ареала
В Северном Казахстане наиболее крупное предмиграционное скопление красавки известно в Костанайской обл. на озерах Уркаш и Жарсор, где в предмиграционный период собирается 15–20 тыс. журавлей, 40% которых составляют красавки (Брагин, 2002). Отмечены скопления на оз. Кушмурун (400 особей) и в устье р. Улькаяк на оз. Кызылколь на границе Костанайской и Актюбинской областей (2500 особей) (Брагин, 2011). В Павлодарской обл. в окрестностях оз. Малый Калмакан в начале августа учтено в общей сложности более 1000 особей (Гисцов, 1991). В Центральном Казахстане в Карагандинской обл. в Тенгиз-Кургальджинской впадине отмечены скопления до 10 тыс. особей (Кошкин, 2007). На юге Казахстана в предгорьях Тянь-Шаня на Ташуткольском водохранилище в июле может собираться до тысячи особей, а во время миграции в конце августа–начале сентября здесь учли 1797 красавок (Березовиков, 2002). В Кыргызстан на оз. Иссык-Куль они начинают прибывать в июле и держатся до конца августа. В период осенней миграции на северо-восточной оконечности озера в районе пос. Тюп и Кар-Кара может собираться до 2.5 тыс. особей, а весной – до 9 тыс. особей (Торопова, Еремченко, 1980; Торопова, Кулагин, 2005). В Хакасии предмиграционные скопления от 150 до 250 особей отмечены на озерах Белё и Черное, а также в урочище Сорокаозерки в Южно-Минусинской котловине (Емельянов, Савченко, 1991). В Забайкальском крае наиболее крупное предмиграционное скопление расположено в котловине Торейских озер. Численность здесь колеблется от 400 особей в засушливый период до 42 тыс. в благоприятные годы (Горошко, 2015).
На основе многолетних наблюдений и анализа литературы существуют детальные описания встреч во время осенней и весенней миграций красавки в Казахстане, Кыргызстане, Узбекистане, Туркменистане и Таджикистане (Гаврилов, 1977; Gavrilov, Van der Ven, 2004). В Казахстане, кроме указанного выше Урало-Эмбинского потока, выделены Арало-Ишимский, Срединно-Казахстанский, предгорный и горный миграционные потоки. Отмечено, что осенняя миграция из Северного и Центрального Казахстана проходит широким фронтом и малозаметна. Наиболее выражена миграция по предгорному участку вдоль северных предгорий Западного Тянь-Шаня и по горному участку – в Иссык-Кульской долине. При описании миграционных потоков оставался вопрос – мигрируют ли красавки из Костанайской обл. только в Индию или и в Африку (Gavrilov, Van der Ven, 2004).
В Туркменистане красавки, мигрирующие в Индию, летят через восточную часть пустыни Каракумы и Келифские озера, расположенные в старом русле р. Амударья (Дементьев, 1952).
В Таджикистане осенью и весной мигрируют через Гиссарский, Зеравшанский и Туркестанский хребты Памиро-Алая, оз. Искандеркуль и нижнее течение рек Кафирниган и Вакш, однако численность их здесь относительно невелика – от нескольких сотен до тысячи (Иванов, 1940; Gavrilov, Van der Ven, 2004).
В Афганистане мигрируют через провинции Балх (Balkh), Кундуз (Kunduz) и Тахар (Takhar) на севере страны с наиболее известными миграционными остановками на озерах Аб-и-Эстада (Ab-e-Estada) и Даште-Навар (Dasht-e-Nawor) (Хан, 2010).
В Пакистане летят через провинции Белуджистан (Balochistan) и Хайбер-Пахтунхва (Khyber Pakhtunkwa) вдоль долин рек Зоб (Zhob), Куррам (Kurram), Гамбила (Gambilla), Кеч (Kech) и Инд (Indus) (Ahmad, Kurshid, 1991; Jan, Ahmad, 1995).
Весной важное значение для отдыха и кормежки красавок имеют озера Айдаркуль и Тузкан в Айдар-Арнасайской системе озер в Узбекистане (Лановенко, Крейцберг, 2006), а также долины рек и образованные на них водохранилища в северных предгорьях Западного Тянь-Шаня в Казахстане (Губин, Скляренко, 1991; Ковшарь, Березовиков, 1991).
На основе визуальных наблюдений отмечено, что на осеннем пролете, проходящем в Казахстане в северных предгорьях Западного Тянь-Шаня в западном направлении, красавка встречается в 23 раза реже, чем на весеннем пролете, который имеет восточное направление (Гаврилов, Гисцов, 1985). Более интенсивная, чем осенняя, весенняя миграция отмечена в Пакистане (Jan, Ahmad, 1995), Афганистане (Хан, 2010), Узбекистане (Лановенко и др., 2011) и Кыргызстане (Торопова, Кулагин, 2011). На этом основании Гаврилов (1977) впервые предположил, что журавли, гнездящиеся восточнее Казахстана, осенью и весной используют разные пролетные пути. Позднее это подтверждено для трех красавок, помеченных передатчиками GPS-GSM на плато Ордос (Ordos) в автономном районе Внутренняя Монголия в Китае в июле 2015 г. (Guo, He, 2017). Осенью они мигрировали над Гималаями и долиной р. Ганг в штат Гуджарат в Индии, а весной возвратились вдоль северных предгорий Тянь-Шаня через Монголию или северо-запад Китая.
Известно, что зимой красавки встречаются почти по всему Индостану, с наибольшей концентрацией в штатах Гуджарат (Gujarat) и Раджастан (Rajasthan) и редкими небольшими группами в остальных штатах – Карнатака (Karnataka), Махараштра (Maharashtra), Мадхья-Прадеш (Madhya Pradesh), Андхра-Прадеш (Andhra Pradesh) и Уттар-Прадеш (Uttar-Pradesh) (Perennou, Mundkur, 1991), а также зимой и ранней весной в долинах на юге Непала (Ripley, 1950).
В Гуджарате журавли зимуют на п-ове Катхиявар (Kathiawar) (другое название Саураштра (Saurashtra), расположенном между Качским и Камбейскими заливами Аравийского моря (Basalingappa et al., 1989). Сельскохозяйственные поля и большое число естественных и искусственных водоемов благоприятны для зимующих журавлей (Banerjee, Gopakumar, 1986). В штате Гуджарат в январе 2002 г. численность красавки составляла 211 900 особей, из них 103 180 держались в районе Порбандар (Porbandar) (Teli, Tatu, 2004).
В штате Раджастан в окрестностях Джодхпура (Jodhpur) журавли широко распределяются по водохранилищам и сельскохозяйственным полям, с наибольшей концентрацией в деревне Кичан (Khichan). Здесь местные жители начали подкармливать красавок зимой 1983/1984 гг., в результате чего численность увеличилась с 200–250 особей до 6 тыс. в 1995/1996 гг. и 11.5 тыс. в 2004/2005 гг. (Pfister, 1996; Jain et al., 2005) и, вероятно, продолжает расти.
Данные дистанционного слежения для 30 журавлей из 24 семей, помеченных в Зауралье, Республиках Алтай и Хакасия, Восточном Казахстане, на Тянь-Шане и в Забайкалье, существенно дополнили информацию из литературных источников.
Для зауральской гнездовой группировки (восток Оренбургской обл.) прослежены осенняя миграция трех семей в 2018 г., а также осенние и весенние миграции одной молодой птицы в 2019 и 2020 г. В предмиграционный период две семьи в течение августа (22 и 28 дней) держались на озерах на Тургайском плато на западе Костанайской обл., расположенных в 55–70 км восточнее мест гнездования и в 60 км от известного места скопления на озерах Уркаш и Жарсор. Третья семья осталась в районе гнездования у оз. Коскуль. Возможно, это озеро также является местом трофического предмиграционного скопления.
В 2018 г. семьи начали осеннюю миграцию 28, 30 и 31.08. Данные ее длительности и дистанции представлены в табл. 1. Маршрут проходил в южном направлении по Арало-Ишимскому миграционному потоку между Аральским морем и хребтом Каратау в Казахстане, через пустыню Кызылкумы в Узбекистане и восточную часть пустыни Каракумы в Туркменистане. Здесь журавли поменяли направление на юго-восточное и пересекли Бактрийскую равнину и горный массив Гиндукуш в Афганистане. В Пакистане пролетели вдоль Сулеймановых гор в провинции Белуджистан, пересекли долину Инд и пустыню Тар и прибыли в индийский штат Раджастан (рис. 1Ca). Из-за нерегулярных сигналов передатчиков места остановок не выявлены, но, судя по продолжительности миграции (6–8 дней), все они были транзитные. В 2019 г. одна молодая птица начала осеннюю миграцию из Карагандинской обл. 12.09. Она полетела в юго-западном направлении, используя Срединно-Казахстанский миграционный поток, через пустыню Бетпак-Дала и над горами Каратау в Казахстане, восточнее пустыни Кызылкумы через Айдар-Арнасайскую систему озер в Узбекистане и Гиссарским хребтом в западной части Памиро-Алая в Таджикистане (рис. 1Сb). Здесь поменяла направление на юго-восточное, и остальной участок пути над Гиндукушем в Афганистане, долиной р. Инд в Пакистане и пустыней Тар в Индии был таким же, как во время осенней миграции в 2018 г.
Отметим, что длительность транзитного перелета в 2018 и 2019 гг. составляла семь дней, хотя дистанция в 2019 г. была на 400 км меньше (2800 км).
Весеннюю миграцию молодая птица начинала 02.04.2019 и 22.03.2020 г. Ее длительность и дистанция представлены в табл. 2. В 2019 г. она совершила транзитный перелет до первой трофической миграционной остановки на оз. Айдаркуль в Узбекистане, где провела 93 дня, с 6.04 по 8.07, после чего переместилась в северном направлении до следующей трофической миграционной остановки в долине р. Сыр-Дарья у пос. Бирлик в Кызыл-Ординской обл. Казахстана, где держалась 19 дней. Она продолжила полет 31.07 на север еще на 100 км и достигла с. Щербаковское на севере Карагандинской обл., расположенного в 750 км северо-восточнее места рождения, откуда через две недели начала осеннюю миграцию. Весной 2020 г. эта молодая птица совершила транзитный перелет до трофической миграционной остановки в долине р. Сыр-Дарья, где оставалась 12 дней, с 27.04 по 8.05. Далее продолжила миграцию на север и достигла запада Карагандинской обл., после чего сигналы прекратились.
Для хакасской гнездовой группировки частично прослежены осенняя миграция двух семей в 2019 г., а также весенняя и осенняя миграции двух молодых птиц в 2020 г. Покинув гнездовую территорию, одна из семей держалась 21 день на месте трофического предмиграционного скопления на степном соленом оз. Белё, в 20 км южнее места рождения. Вторая семья сначала кормилась в урочищe Трехозерки в Южно-Минусинской котловине, где летом 2019 г. отмечены летние скопления неполовозрелых и негнездящихся красавок и серых журавлей (Ильяшенко и др., 2020б), а затем также переместилась севернее на оз. Белё. Отсюда обе семьи начали осеннюю миграцию 3 и 6.09, однако путь проследить не удалось. В 2020 г. одна из молодых птиц также начала осеннюю миграцию с оз. Белё 8.09, вторая 10–12.09 с запада Красноярского края, где держалась в течение августа. Маршрут осенней миграции молодых птиц в 2020 г. проходил над Западным Саяном на западе Тывы, западной частью Джунгарской равнины, Восточным Тянь-Шанем и западной частью пустыни Такла-Макан в Синьцзян-Уйгурским АР и горной цепью Каракорум в Тибетском АР Китая и пустыней Тар в индийском штате Раджастан (рис. 1D).
Из-за нерегулярности и непродолжительности сигналов места трофических остановок, время пребывания на них и длительность осенней миграции не выявлены, а общая дистанция представлена в табл. 1.
Весеннюю миграцию две молодые птицы начали 22 и 24.03.2020 г. Ее длительность и дистанция приведены в табл. 2. Они полетели на север через Пакистан и Афганистан и сделали первую трофическую миграционную остановку на оз. Айдаркуль в Узбекистане, а затем в долине р. Чу в северных предгорьях Западного Тянь-Шаня в Казахстане, где держались в общей сложности 40–50 дней. Далее одна птица достигла котловины Больших озер в Монголии, поменяла направлении на северное и в начале июня прилетела в Хакасию. Вторая птица, полетев от предгорий Западного Тянь-Шаня в северо-восточном направлении, достигла места длительной трофической миграционной остановки в долине р. Эмель (Emel, Emin) в окрестностях пос. Дурбульджин (Dorbiljin) Синьцзян-Уйгурского АР Китая. В Хакасию она прилетела в конце июня.
Летом один из журавлей держался в урочище Трехозерки и затем переместился на оз. Белё, второй журавль – в бассейне р. Абакан, после чего полетел на северо-восток, на запад Красноярского края.
В Республике Алтай в Чуйской степи (алтайская гнездовая группировка) в 2018 г. передатчиками помечены два птенца из двух семей, сигналы от которых продолжали поступать ко времени подготовки статьи в ноябре 2020 г.
В предмиграционный период 2018 г. обе семьи держались в районе гнездовых территорий до 30.08. Далее они совершили перемещения с двумя остановками. Первая находилась в 300 км от мест гнездования, в Западной Монголии на пресноводном оз. Хар-Ус, где в июле 1995 г. на скоплении спутниковыми передатчиками помечено девять журавлей (Kanai et al., 2000). Таким образом, это озеро является как местом миграционной трофической остановки для журавлей, гнездящихся севернее, так и местом летнего и предмиграционного трофического скопления красавок, гнездящихся в Западной Монголии. Вторая трофическая миграционная остановка была на оз. Баркёль в Синьцзян-Уйгурском АР Китая, в 500 км от оз. Хар-Ус. Это бессточное горько-соленое озеро расположено в межгорной пустынной котловине, на восточной оконечности Тянь-Шаня. Наличие пресных водоемов и сельскохозяйственных полей, прилегающих к озеру на северо-востоке, делают эту остановку благоприятной для пополнения энергетических ресурсов.
Данные по длительности и дистанции осеннего пролетного пути представлены в табл. 1. В 2019 и 2020 гг. молодые птицы начинали трофический этап осенней миграции в период с 12 по 30.08 с мест летнего пребывания в Республиках Алтай и Тыва и места трофической миграционной остановки в Западной Монголии. До достижения оз. Баркёль каждый из меченых журавлей в разные годы использовал одну-две из четырех выявленных трофических миграционных остановок (табл. 3), где эти птицы держались 10–15 дней. Во время трофического этапа пролетный путь проходил над Алтайскими горами в Республике Алтай и Западной Монголии и Синьцзян-Уйгурским АР Китая.
В течение трех осенних сезонов журавли начинали транзитный перелет с оз. Баркёль (табл. 1), как и красавки, помеченные в 1995 г. (Kanai et al., 2000). Они стартовали в период с 13 по 23.09. Во время транзитного этапа маршрут проходил в Китае над восточной частью Тянь-Шаня и Кашгарией к востоку от пустыни Такла-Макан в Синьцзян-Уйгурском АР, Гималаями в Тибетском АР и пустыней Тар в штате Раджастан в Индии (рис. 1, Е).
Весной 2019 г. два молодых журавля начинали весеннюю миграцию 18 и 26.03, в 2020 г. – 27 и 26.03. Ее длительность и дистанция представлены в табл. 2. Обе птицы полетели в северном направлении, но нерегулярность сигналов не позволила проследить полностью их путь. Одна из них в 2019 и 2020 гг. делала трофические миграционные остановки в Казахстане в северных предгорьях Западного Тянь-Шаня в окрестностях Терс-Ашибулакского водохранилища в Жамбыльской обл., а затем на Капчагайском водохранилище в Алматинской обл. Она держалась на них до середины мая в 2019 г. и до конца апреля в 2020 г. Сигналы от второй птицы в 2019 г. во время весенней миграции отсутствовали. В 2020 г. она сделала трофическую остановку на оз. Айдаркуль в Узбекистане, где оставалась около месяца. Сигналы от обоих журавлей на участке от северных предгорий Тянь-Шаня и до запада Синьцзян-Уйгурского автономного района также отсутствовали. Но, судя по опубликованным данным (Анненков, 1991; Ковшарь, 2007), можно предположить, что от предгорий Тянь-Шаня они пролетели на северо-восток вдоль западных склонов Джунгарского Алатау, далее между озерами Алаколь и Сасыколь и затем вдоль долины р. Эмель в восточном направлении в Синьцзян-Уйгурский АР. Здесь они поменяли направление на северное – северо-восточное и достигли мест рождения.
Один из молодых журавлей после первой зимовки в 2019 г. провел лето на месте мечения в Чуйской степи, в 2020 г., после второй зимовки – в бассейне р. Ховд в Баян-Улэгейском аймаке в Монголии. Вторая молодая птица после первой зимовки в 2019 г. провела лето в 400 км северо-восточнее места рождения, в Тыве, в долинах рек Хемчик и Чадан и у Саяно-Шушенского водохранилища, а в 2020 г., после второй зимовки – в Чуйской степи в Республике Алтай, в месте своего рождения.
Для восточно-казахстанской гнездовой группировки получены данные от семи птенцов из шести семей, помеченных в Восточно-Казахстанской и Алматинской областях в 2018 и 2019 гг., но передатчики работали с перерывами. Четыре семьи, гнездившиеся на восточном и западном побережьях оз. Алаколь, в конце августа переместились на 100 км на восток в Китай, на место трофического предмиграционного скопления в Синьцзян-Уйгурском АР в долине р. Эмель между пос. Юйминь (Yumin) и Дурбульджин. Здесь сигналы от одной птицы прекратились, а остальные три держались в этом районе от 15 до 30 дней и 21, 25.08 и 1.09 полетели в западном направлении, сделав трофические миграционные остановки на Капчагайском водохранилище на р. Или и Ташуткольском водохранилище на р. Чу. Пятая семья с птенцом, помеченным в Ушаральском районе Алматинской обл., 30.08 прилетела на Ташуткольское водохранилище, однако из-за отсутствия сигналов неясно, держалась ли она до этого еще на каком-либо месте трофического предмиграционного скопления. Шестая семья с птенцом, помеченным в Илийском р-не Алматинской обл., 17.08 переместилась из района гнездования южнее в окрестности г. Алматы, затем 6.09 полетела в западном направлении вдоль северных предгорий Тянь-Шаня и достигла Ташуткольского водохранилища. Из пяти семей, достигших Ташуткольского водохранилища, прослежена только одна, которая начала отсюда транзитный перелет 12.09.2019 и достигла места зимовки в штате Раджастан 25.09. Сигналы от трех семей на Ташуткольском водохранилище прекратились, от одной семьи – возобновились в середине периода зимовки.
В 2020 г. частично прослежена осенняя миграции двух годовалых особей. Они использовали Ташуткольское водохранилище в качестве трофической миграционной остановки. Отсюда полетели в западном направлении, используя предгорный миграционный поток, через Терс-Ащибулакское водохранилище, далее поменяли направление на южное и мигрировали через западную оконечность Тянь-Шаня в Казахстане, Памиро-Алайскую горную систему в Таджикистане, западную часть Гиндукуша в Афганистане, вдоль Сулеймановых гор и долину р. Инд в провинции Пенджаб в районе г. Раджанпур (Rajanpur) в Пакистане. Здесь 8.09 найден передатчик от одной из птиц, информация о гибели которой в результате охоты поступила в Институт поведения животных Макса Планка. Вторая птица продолжила миграцию и достигла пустыни Тар в штате Раджастан в Индии (рис. 1F). Длительность и дистанция их осенней миграции представлены в табл. 1.
Весной 2020 г. две молодые особи начали миграцию 18 и 28.03. в северном направлении. Затем от одной из них сигналы пришли 29.04 с оз. Сысыколь, расположенном севернее оз. Алаколь, места ее рождения. Далее с начала мая и до начала трофического этапа миграции 22.08 она держалась западнее Алаколя в бассейнах рек Аксу, Лепсы и Тентек. Вторая красавка совершила транзитный перелет до Узбекистана, где держалась до 2 мая в предгорьях хребта Нуратау севернее оз. Айдаркуль. Далее вдоль северных предгорий Тянь-Шаня полетела на восток и достигла 4.05 водохранилища на р. Каш (Kash), притока р. Или в Синьцзян-Уйгурском АР Китая. 16.05 она вернулась в Алматинскую обл. в междуречье Аксу и Лепсы, а в июне вновь полетела в Синьцзян-Уйгурский автономный район, где держалась в долине р. Эмель в окрестностях Дурбульджина до 31.08, когда начался трофический этап миграции. Из-за недостатка данных определена лишь дистанция весенней миграции (табл. 2).
На Тянь-Шане (тянь-шаньская гнездовая группировка) на высокогорном оз. Тузколь у стыка границ Казахстана, Киргизии и Китая в мае 2017 г. на гнезде помечен самец, регулярные сигналы от которого поступали около двух лет. Он встречен в год мечения 9 и 10.08 в семье с двумя птенцами (Ильяшенко и др., 2018). В предмиграционный период держался на сельскохозяйственных полях в окрестностях Тузколя, а 16.08 совершил однодневный перелет на запад на северо-восточную оконечность оз. Иссык-Куль, где 21 день кормился на полях между пос. Тюп и Кар-Кара. В 2018 г., судя по широким перемещениям в окрестностях оз. Тузколь, пара не гнездилась. 26.07 он полетел на оз. Иссык-Куль, где держался 38 дней. С этой трофической остановки в оба сезона начинал транзитный перелет на место зимовки 6.09.2017 и 2.09.2018. Длительность и дистанция его осенней миграции представлены в табл. 1. Маршрут проходил над Тянь-Шанем в Кыргызстане, над западной частью пустыни Такламакан в Синьцзян-Уйгурском АР Китая, восточнее Памиро-Алая, над горной системой Каракорум и Гималаями в Тибетском АР Китая, над долиной р. Инд в Пакистане и пустыней Тар в штате Раджастан (рис. 1G). Оба сезона он делал по пути по семь кратковременных транзитных остановок для ночного отдыха на горных озерах или в межгорных долинах, а в долине р. Инд – на сельскохозяйственных полях.
Весеннюю миграцию этот журавль начинал 21.03.2018 и 10.03.2019. Он совершил транзитный перелет, длительность и дистанция которого даны в табл. 2. Как и другие красавки, полетел в северном направлении, достиг западной оконечности Тянь-Шаня и далее вдоль северных предгорий полетел на восток. Сделав остановку на Ташуткольском водохранилище на пять дней в 2018 г. и на 10 дней в 2019 г., достиг мест гнездования на оз. Тузколь 5.04 и 26.03 соответственно.
Для забайкальской гнездовой группировки в 2018 г. частично прослежена осенняя миграция пяти семей, в 2019 г. – весенняя и осенняя миграции двух годовалых особей, а в 2020 г. – весенняя и частично осенняя миграции одной годовалой и одной двухгодовалой особей.
В предотлетный период семьи кормились на местах трофических предмиграционных скоплений в котловине Торейских озер. В конце августа– начале сентября переместились на места трофических миграционных остановок, расположенных в 300–1000 км юго-восточнее мест гнездования (табл. 3). Каждый из меченых журавлей использовал две-три из этих четырех трофических остановок в течение 25–46 дней.
Три семьи в 2018 г. и две молодые особи в 2019 г. начинали транзитный перелет из округа Шилин Гол 21, 26, 27.09 и 2.10. Длительность и дистанция их осенней миграции представлены в табл. 1. Маршрут проходил через Монгольское плато в автономном районе Внутренняя Монголия, Тибетское плато и Гималаи в Тибетском АР в Китае, Индо-Гангскую низменность в Непале и индийский штат Уттар-Прадеш и закончился в штатах Раджастан и Гуджарат (рис. 1H). Этим же маршрутом летели годовалая и двухгодовалая особи осенью 2020 г. Нерегулярность сигналов позволила выявить лишь некоторые транзитные остановки в долине р. Хуанхэ севернее Баотоу (Baotou) в Нинься-Хуэйском АО вблизи г. Нинься (Китай); в бассейне оз. Цинхай одноименной провинции, где прекратились сигналы от трех красавок, помеченных в 1995 г. (Kanai et al., 2000); в горной долине на Тибетском плато и в округе Джонгба в северных предгорьях Гималаев. Некоторые из них совпадают с остановками журавлей, помеченных в 2015 г. на плато Ордос в автономном районе Внутренняя Монголия, расположенном на 1000 км южнее Шилин Гол (Guo, He, 2017).
В 2019 г. два годовалых журавля начали весеннюю миграцию 19 и 31.03, в 2020 г. годовалый и двухгодовалый – 20 и 26.03. Оба весенних сезона они летели с места зимовки в северном направлении и, достигнув западной оконечности Тянь-Шаня, меняли направление на восточное. В 2019 и 2020 гг. останавливались на длительный отдых на трофических миграционных остановках в Узбекистане и Казахстане (табл. 3), где держались в течение апреля до 29 дней. Затем совершили еще один транзитный перелет до места летнего пребывания в Китае, Монголии или Забайкалье в России. В Алматинской обл. птицы (n = 3) пересекали границу Казахстана и Китая вдоль долины р. Или. В 2019 г. один журавль далее полетел через хребет Борохоро, южную часть Джунгарской впадины в Синьцзянь-Уйгурском АР в Китае, Гобийский Алтай, вдоль южных предгорий Хангая и северной границы пустыни Гоби в Монголии и достиг округа Шилин Гол в автономном районе Внутренняя Монголия Китайской Народной Республики. Сигналы от второго на этом участке пути отсутствовали. Этим же путем в 2020 г. летел годовалый журавль. Двухлетний журавль в 2020 г., после пересечения границы Казахстана и Китая, полетел южнее, чем в 2019 г., вдоль южной границы пустыни Гоби и достиг округа Шилин Гол.
Длительность и дистанция весенней миграции одной особи в 2019 г., для которой получены наиболее полные данные, даны в табл. 2.
В 2019 г. оба годовалых журавля все лето до отлета на зимовку держались на довольно обширной территории в округе Шилин Гол, в 1000 км северо-западнее мест рождения. В 2020 г. двухгодовалый журавль, достигнув 24.05 округа Шилин-Гол, поменял направление на северное и 30.05 сигналы поступили уже из Забайкальского края, где он держался, главным образом, в долине р. Ингода в окрестностях пос. Дарасун, в 200 км от места рождения, до отлета в конце августа. Годовалый журавль в 2020 г. провел лето в Хэнтийском аймаке в Монголии.
Все журавли из азиатской части ареала от Зауралья до северо-востока Китая прибыли на зимовку в Северо-Западную Индию. Прилет проходил с севера, северо-востока и востока в период с 5.09 по 17.10 (42 дня), в том числе особей из Зауралья и Восточного Казахстана – с 5 по 23.09 (n = 5), из Республики Алтай и Хакасии – с 24.09 по 1.10 (n = 4), из Забайкалья – с 1 по 13.10 (n = 7).
Слежение в течение двух зимних сезонов показало, что из 21 журавля две птицы (9.5%) сразу полетели в штат Гуджарат, а 19 остановились в штате Раджастан, где держались, главным образом, в окрестностях Джодхпура на полях между пустыней Тар и хребтом Аравали. Осенью здесь, вероятно, еще сохранилась вода в реках, стекающих с гор после летних муссонных дождей. С приходом более засушливого сезона 14 журавлей (66%) в период с 17.10 по 10.11 переместились в штат Гуджарат. Подобная стратегия описана для журавля, помеченного в 1995 г. в Копе в Казахстане, который после почти месячного пребывания в штате Раджастан переместился в Гуджарат (Kanai et al., 2000). В Гуджарате красавки держались, главным образом, вдоль побережья Аравийского моря, с наибольшей концентрацией на водохранилищах у пос. Мияни (Miyani) и Порбандар (Porbandar), на солончаке Большой Качский Ранн, а также на северном и южном побережьях Качского залива. Журавль с белым пластиковым кольцом, помеченный на востоке Оренбургской обл., встречен 24.12.2018 на водохранилище Вадла (Vadla) недалеко от орнитологического заказника Нал Cаровар (Nal Sarovar) (Ильяшенко и др., 2020).
Пять из 21 (23.5%) журавлей оставались в штате Раджастан всю зиму. Журавль, помеченный на оз. Тузколь на Тянь-Шане, встречен 30.10.2019 в Ланкарансар (Lunkaransar), Биканер (Bikaner) в штате Раджастан, уже после прекращения работы передатчика (Бохра, 2020). Кроме того, журавль, помеченный в Республике Алтай белым пластиковым кольцом, встречен в деревне Кичан на севере пустыни Тар в феврале 2019 г. (Ильяшенко и др., 2020).
Таким образом, дистанционное слежение подтвердило известную ранее информацию, что ключевые территории зимовки в Индии лежат в окрестностях Джодхпура в штате Раджастан и на п-ове Катхиявар в штате Гуджарат. Ни одна из 21 меченой особи не использовала места зимовки в других штатах.
Весной отлет с мест зимовки журавлей азиатской части ареала проходил с 10.03 по 3.04 (24 дня). Стартуя с мест зимовки, красавки летели единым потоком шириной 100–200 км на протяжении 700–800 км до западной оконечности Тянь-Шаня. Их общая численность на этом участке пути на основании литературных данных может быть оценена в 125–160 тыс. особей (Ильяшенко, 2018; Ilyashenko, 2019). Меченые особи летели транзитом через долину р. Инд над районом Зхоб в провинции Белуджистан в Пакистане, горный массив Гиндукуш в Афганистане, Келифские озера в восточной части Туркменистана или долину р. Амударья в районе г. Термез в Узбекистане. Они делали транзитные остановки в долине р. Инд перед пересечением Гиндукуша, на берегах рек и озер в Пакистане и Афганистане без выбора определенных территорий. При облете с запада Памиро-Алая и Западного Тянь-Шаня ширина миграционного потока увеличивалась до 300 км. Такое расширение соответствует наблюдениям весенней миграции журавлей в южной части Узбекистана в Сурхандарьинской, Самаркандской, Кашкадарьинской, Джизакской и Ташкентской областях (Лановенко и др., 2011).
Достигнув западной оконечности и затем северных предгорий Тянь-Шаня, журавли делали трофические миграционные остановки в Узбекистане и Казахстане, причем из шести выявленных трофических территорий (табл. 3) каждый из меченых журавлей использовал по две-три.
По достижении западной оконечности Тянь-Шаня, начиная с оз. Айдаркуль и далее с других трофических миграционных остановок в северных предгорьях Тянь-Шаня, проходил разлет веером на север, северо-восток и восток. Часть журавлей продолжали миграцию в северном направлении, остальные, поменяв в Южно-Казахстанской обл. направление с северного на восточное, летели вдоль северных предгорий Тянь-Шаня и на этом пути меняли направление на северо-восточное на места гнездования в южной Сибири и Западной Монголии или продолжали движение в восточном направлении на места гнездования в Забайкалье, Монголии и Северо-Восточном Китае. При этом красавки следовали в восточную часть ареала довольно широким фронтом, как показали три особи, помеченные нами в Забайкалье, и три красавки, помеченные в 2015 г. в Китае на плато Ордос во Внутренней Монголии (Guo, He, 2017).
ЗАКЛЮЧЕНИЕ
Представленные материалы обобщают и значительно дополняют данные визуальных наблюдений о предмиграционных скоплениях, пролетных путях, миграционных остановках, зимовках и местах летнего пребывания красавок из разных гнездовых группировок.
Выделено восемь миграционных потоков.
В европейской части ареала существуют два потока (рис. 1А, 1B).
Первый поток образуют журавли азово-черноморской группировки, летящие осенью из Приазовской возвышенности, Крыма и Таманского п-ова в бассейн озер Чад и Фитри в Республике Чад. По второму потоку журавли прикаспийской и волго-уральской группировок, гнездящиеся от Восточного Предкавказья до Предуралья, летят на зимовку в бассейн Нила и Атбары в Судане и прилегающих районах Эфиопии и Эритреи. Весной журавли обоих потоков летят теми же путями, но некоторые особи прикаспийской и волго-уральской группировок могут пролетать западнее через Восточную Турцию.
Перемещения журавлей между областями зимовки в Чаде и Судане не отмечено. Эти территории разделены горными массивами Дарфур на западе Судана и Эддени, Эрди и Вадаи на востоке Чада.
Некоторые красавки из европейской части ареала могут использовать оба места зимовки и оба пути пролета, пролетая по промежуточной ветви через Израиль, Сирию и Иорданию, что объясняет встречи красавок в этих странах в период миграций (Howes, 1989; Azar et al., 2007).
В азиатской части ареала выявлено шесть осенних миграционных потоков, ведущих к общей зимовке в Северо-Западной Индии, где все меченые нами журавли держались на территории только двух штатов – Гуджарата и Раджастана.
Журавли из зауральской гнездовой группировки следуют маршрутом, по которому, возможно, летят также птицы из Северного и Центрального Казахстана. В Казахстане он идет широким фронтом по Арало-Ишимскому и Срединно-Казахстанскому пролетным путям через пустыни Кызылкумы и восточную часть Каракумов. При этом часть журавлей огибает Памиро-Алай, а часть летит над его хребтами. Далее поток сужается, и красавки мигрируют по межгорным долинам через горный массив Гиндукуш, долину р. Инд и пустыню Тар (рис. 1Са, 1Сb).
Второй поток выявлен для журавлей хакасской гнездовой группировки (рис. 1D). Третий поток, которым следуют журавли алтайской гнездовой группировки (рис. 1Е), вероятно, используют и красавки, гнездящиеся в Тыве и Западной Монголии. Этот же маршрут использовали три журавля, помеченные на оз. Хар Ус в Западной Монголии, при этом они пересекали пустыню Такла-Макан как над восточной, так и над центральной ее частями (Kanai et al., 2000).
Учитывая небольшое число меченых особей в Республиках Хакасия и Алтай, можно предположить, что красавки из этих географических регионов могут использовать как второй, так и третий миграционные потоки. Кроме того, журавли из этого региона могут лететь и по маршруту четвертого миграционного потока (см. ниже). Так, красавка, помеченная в Западной Монголии, полетела в юго-западном, а затем в западном направлении по горному миграционному потоку через оз. Иссык-Куль, вдоль Ферганской долины до Исфаны в Таджикистане, где сигналы прекратились (Kanai et al., 2000).
Четвертым миграционным потоком следуют журавли восточно-казахстанской гнездовой группировки (рис. 1E). Они используют сначала предгорный миграционный поток в западном направлении, а затем меняют его на юго-западное или южное направления. Этим же путем мигрировала молодая красавка, помеченная на р. Копа в Казахстане в 1995 г. (Kanai et al., 2000).
Миграционные потоки журавлей зауральской и восточно-казахстанской группировок могут сливаться при пересечении Памиро-Алая и Гиндукуша, после чего журавли следуют единым путем до места зимовки в Индии.
Пятый миграционный горный поток используют журавли тянь-шаньской гнездовой группировки, гнездящиеся в горных долинах Тянь-Шаня (рис. 1G). Этот путь могут использовать и красавки из Восточного и Юго-Восточного Казахстана, которые стартуют с северных предгорий Тянь-Шаня и летят в южном направлении по межгорным долинам Центрального и Внутреннего Тянь-Шаня (Gavrilov, Van der Ven, 2004).
Шестой поток используют журавли забайкальской гнездовой группировки, а также особи, обитающие в Северо-Восточной Монголии и Северо-Восточном Китае (рис. 1H). Этим путем мигрировали и три красавки, помеченные на плато Ордос в автономном районе Внутренняя Монголия в Китае в июле 2015 г. (Guo, He, 2017).
Все осенние миграционные потоки, за исключением первого, большую часть пути проходят над горами, в том числе Гималаями, пересекая их в различных местах на высоте до 7 тыс. м над ур. м., откуда имеется ряд прямых наблюдений (Martens, 1971; Bijlsma, 1991; Kanai et al., 2000).
Весной все журавли азиатской части ареала летят узким фронтом до западной оконечности Тянь-Шаня по тому же пути, каким осенью следуют журавли из Зауралья, Северного и Центрального Казахстана. Далее часть продолжает миграцию на север, часть меняет направление на восточное и от предгорий Западного Тянь-Шаня веером разлетается на северо-восток и восток. Журавли забайкальской и, возможно, алтайской и хакасской группировок совершают транзитную миграцию двумя этапами, разделенными перемещениями между трофическими миграционными остановками.
За исключением журавлей зауральской группировки, все красавки азиатской части ареала совершали кольцевую миграцию, используя осенью и весной разные пролетные пути. Разница в их дистанции варьирует от 600–700 км для алтайской и восточно-казахстанской группировок, 1300–1500 км для хакасской и тянь-шаньской группировок и до 2400 для забайкальской группировки.
Практически для каждого миграционного потока, за исключением используемого красавками зауральской группировки, выявлены или подтверждены наиболее значимые территории, которые журавли используют в качестве летних скоплений, осенних трофических предмиграционных скоплений, осенних и весенних трофических миграционных остановок и зимовок (табл. 3). Такие территории расположены в зонах зернового земледелия, где птицы могут накапливать энергетические ресурсы. Некоторые осенние трофические территории являются “отправными” пунктами транзитной миграции и обычно расположены перед географическими преградами.
Для журавлей из Зауралья, Северного и Центрального Казахстана мест крупных трофических предмиграционных скоплений не выявлено. Обилие озер на Тургайском плато, окруженных обширными сельскохозяйственными полями, обеспечивает питание и отдых, и красавки мигрируют небольшими группами широким фронтом.
Как на длительных трофических, так и на кратковременных транзитных остановках красавки используют довольно обширную территорию, в пределах которой могут останавливаться на ее разных участках. В горных долинах рек такие территории небольшого размера.
Периоды начала и окончания осенней и весенней миграций меченых журавлей в целом соответствуют опубликованным данным (Гаврилов, 1977; Кашкаров и др., 1977; Гаврилов, Гисцов, 1985; Матюхин и др., 1991; Губин, Скляренко, 1991; Kanai et al., 2000; Букреева, 2003; Guo, He, 2017). Нами выявлено, что миграции журавлей в европейской и азиатской частях ареала различаются по фенологическим параметрам. В европейской части ареала осенняя миграция проходит в довольно сжатые сроки, в то время как весенняя миграция более растянута. В азиатской части ареала сроки осенней миграции более растянуты, сроки весенней – более сжаты.
В целом осенью отмечено смещение дат начала транзитной миграции с запада на восток: наиболее ранние для журавлей азово-черноморской группировки (с 17 по 24.08) и наиболее поздние для птиц из Забайкалья (26.09–2.10) (рис. 3). Очевидно, это связано с аналогичным смещением сроков гнездования с запада на восток, что, в свою очередь, зависит от климатических факторов и протяженности весеннего миграционного пути. Вследствие этого птенцы из восточных гнездовых группировок к концу августа еще физически не готовы к транзитному перелету и сначала совершают перемещения между трофическими остановками в направлении миграции. Отмечено также небольшое смещение с запада на восток дат начала весенней миграции: с 1 по 20.03 для азово-черноморской группировки и 10.03–3.04 для журавлей из азиатской части ареала.
Рис. 3.
Средние величины дистанции и сроков осенней миграции красавок из различных частей гнездового ареала.
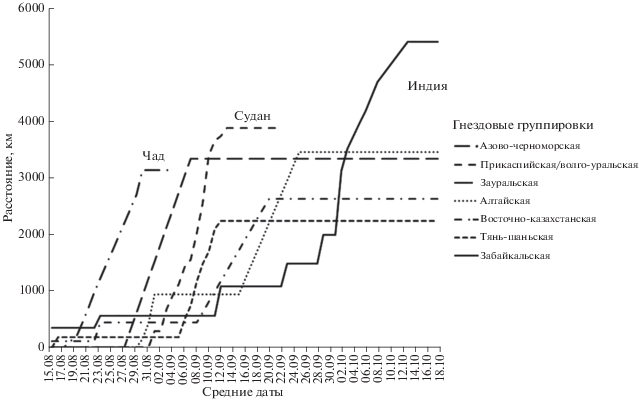
В некоторых публикациях данные по срокам осенней миграции красавки в Грузии (Abuladze, 2018), Азербайджане (Султанов и др., 2011) и Египте (Atta, 1996) ошибочны, что связано с трудностью отличия красавки от серого журавля в полете по внешнему виду и крикам, особенно на большой высоте. О сложности отличия подробно описано при наблюдениях за мигрирующими стаями на о-ве Кипр (Flint, 1971) и в Средней Азии и Казахстане (Гаврилов, 1977; Gavrilov, Van der Ven, 2004; Лановенко и др., 2011; Торопова, Кулагин, 2011). Согласно визуальным наблюдениям (Букреева, 2003) и данным дистанционного слежения, миграция журавлей прикаспийской и волго-уральской группировок начинается после двадцатых чисел августа и заканчивается к середине сентября, с наиболее интенсивным стартом в конце августа – начале сентября. А так как журавли осенью летят транзитом, не останавливаясь на длительный срок, с продолжительностью миграции в среднем 10 дней, то и сроки пролета проходят в указанный период с небольшим сдвигом в датах. Поэтому вышеуказанные встречи в конце сентября – начале ноября очевидно относятся к пролету серых журавлей, мигрирующих именно в эти даты (Белик, 2006).
Слежение за осенними миграциями красавок показали, что транзитный перелет проходит очень быстро, с остановками лишь на короткий отдых, без восполнения энергетических ресурсов. При общей протяженности пролетного пути от 2170 до 5600 км, транзитные перелеты варьировали от 1900 до 4600 км, а их продолжительность – от 7 до 13 дней. Очевидно, это тот период, который красавка способна преодолевать без восполнения энергетических затрат, практически только за счет ресурсов, накопленных перед началом транзитного перелета. Подобная продолжительность транзитного этапа миграции получена и по результатам предыдущих исследований (Kanai et al., 2000; Guo, He, 2017).
Длительность весенней миграции более растянутая, в том числе у взрослых особей, за счет большего числа кратковременных транзитных остановок, меньшей скорости и более коротких дневных перелетов. Такой характер миграции, очевидно, связан с экономией энергии, необходимой для гнездования, а также с более бедной, чем осенью кормовой базой.
Некоторые молодые особи весной летят вместе с родителями. Случаи, когда птенцы прошлого года рождения держались с семьей вплоть до появления у родителей нового потомства, отмечены на Приазовской возвышенности (Андрющенко и др., 2006). Большая часть неполовозрелых особей вместе со взрослыми птицами совершает транзитную миграцию до первых мест длительного пребывания. Далее они продолжают перемещения на север на места летних скоплений в пределах гнездовой части ареала, прибывая туда на 1–1.5 месяца позже взрослых (рис. 4). При этом следует отметить, что в азиатской части ареала, где маршруты осенней и весенней миграций различны, молодые особи после длительных трофических остановок в середине пути продолжают транзитную миграцию самостоятельно по незнакомому для них маршруту. Некоторые неполовозрелые особи весной не достигают гнездовой части ареала и остаются на все лето в пределах миграционного коридора, откуда начинают осеннюю миграцию. Об этом свидетельствуют как наши данные о летнем пребывании молодых меченых птиц, не достигших районов гнездования родителей, в Турции, Казахстане и Китае, так и сообщения о летних встречах красавки в Армении (Саркинсян, 2011), Иране (Ashtiani, 1999), Турции (Akarsu et al., 2013) и Узбекистане (Лановенко, Крейцберг, 2006).
Рис. 4.
Средние величины дистанции и длительности весенней миграции взрослых и молодых особей из различных частей гнездового ареала.
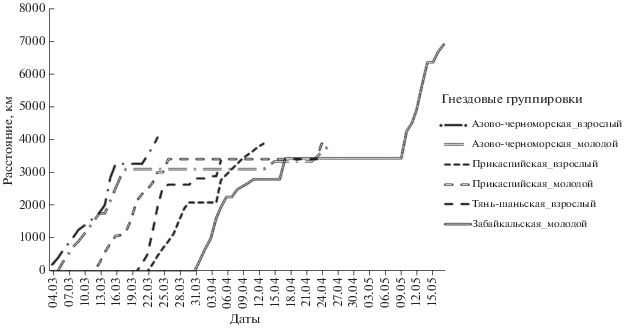
Некоторые молодые особи посещают район рождения только после второй зимовки. Одна из птиц, помеченных в Республике Алтай, после первой зимовки провела лето в Тыве, в 500 км от места рождения, а после второй – на месте рождения. Птенец, помеченный в Заволжье, после первой зимовки провел лето в долине Маныча, а после второй – прилетел на место рождения. Журавль, помеченный в Западном Казахстане, после первой зимовки летом держался на границе Волгоградской и Саратовской областей, а после второй – прилетел на место рождения на Камыш-Самарские озера в окрестности пос. Жангалы. Случаи, когда молодые журавли только на второй год посещают место рождения, отмечены и для серых журавлей в европейской части России (Ю.М. Маркин, личн. сообщ.).
Дистанционное слежение показало некоторую изолированность гнездовых группировок. Разрыв между азово-черноморской и прикаспийской/ волго-уральской группировками обусловлен антропогенной трансформацией территории, особенно после массовой кампании по освоению целинных земель в середине XX века. Деградация гнездовых местообитаний в последние десятилетия усугубились зарастанием пастбищ высокотравьем вследствие олуговения степей и сокращения площади выпаса скота из-за кризиса в животноводстве и перевода крупного рогатого скота на стойловое содержание (Ильяшенко, 2019; Белик и др., 2020). Вероятно, среди 8.4 тыс. красавок, учтенных на оз. Акротири на Кипре в 1957 г. (Flint, 1971), были не только птицы азово-черноморской группировки, но и журавли из соседних районов в бассейне Дона. Более низкая оценка численности (около 1500 особей) в 1971 г. (Flint, 1972) указывает не только на возможный недоучет птиц, но и на сокращение численности красавки, отмеченное в те годы в гнездовой части ареала (Ильяшенко, 2018). Использование в разные годы неполовозрелой особью, помеченной в Западном Казахстане, обоих африканских мест зимовки, указывает на возможности обмена птицами азово-черноморской и прикаспийской/ волго-уральской группировок. Очевидно, что ранее, при большей численности красавки на юге европейской части России и Украине, такой обмен мог быть более интенсивным. Возможно, этим объясняется добыча красавки на зимовке в Судане, помеченной в 1892 г. в Аскании-Нова (Андрющенко и др., 2006).
Последние данные показали также относительную изолированность журавлей из европейской части ареала, мигрирующих в Северо-Восточную Африку, и журавлей из азиатской, следующих в Индию. Вероятно, их разделяют южные предгорья Уральских гор, Мугоджары и степи Урало-Эмбинского междуречья. Обследования в 2018 и 2019 гг. степей в центральной части юга Оренбургской и Актюбинской областей не дали положительных результатов (Ильяшенко и др., 2020в). Красавки не были отмечены и во время обследования этой территории в начале 2000-х годов (Ковшарь и др., 2007; Ерохов, 2009), что, очевидно, связано с долговременной засухой. Однако в более благоприятные годы на этой территории регистрировали пары и небольшие группы (Варшавский и др., 1991; Ковшарь, Давыгора, 2004). Из этого следует, что полной изоляции между журавлями из европейской и азиатской частей ареала не существует. Таким образом, несмотря на относительную изолированность журавлей из географических популяций, использующих три области зимовки, между ними возможен поток генов, что подтверждается невысокой генетической дифференциацией красавки от азово-черноморского побережья до Забайкалья (Mudrik et al., 2018). При этом генетический обмен более интенсивен в азиатской части ареала, где находится одна область зимовки, по сравнению с европейской, где их две.
Выделение гнездовых группировок красавки актуально для мониторинга их состояния и перераспределения, а также для природоохранных целей.
Список литературы
Андрющенко Ю.А., 1997. Положение украинской группировки журавля-красавки в пределах мирового ареала вида // Беркут. № 6 (1–2). С. 33–46.
Андрющенко Ю.А., 2015. Ревизия ареала азово-черноморской популяции красавки // Журавли Евразии (биология, распространение, разведение). Вып. 5. М. С. 153–167.
Андрющенко Ю.А., Шевцов А.А., 1998. Летние скопления журавля-красавки на Сиваше // Бранта: сборник научных трудов Азово-Черноморской орнитологической станции. № 1. С. 92–102.
Андрющенко Ю.А., Горлов П.И., 1999. Состояние журавля-красавки и серого журавля на Сиваше // Размещение околоводных птиц на Сиваше в летне-осенний период. Бранта: Мелитополь – Сонат: Симферополь. С. 83–88.
Андрющенко Ю.А., Винтер С.В., Стадниченко И.С., 2006. Мечение красавки на юге Украины // Журавли Евразии (биология, охрана, разведение). Вып. 2. М. С. 104–108.
Андрющенко Ю.А., Мнацеканов Р.А., Динкевич М.А., 2008. Современное состояние красавки на Керченском и Таманском полуостровах // Журавли Евразии (биология, распространение, миграции). Вып. 3. М. С. 141–147.
Анненков Б.П., 1991. Миграции журавля-красавки в Алакольской котловине // Журавль-красавка в СССР. Алма-Ата: изд-во “Гылым”. С. 149–151.
Белик В.П., 1999. Современное состояние донских популяций журавля-красавки // Журавли Украины. Мелитополь. С. 38–53.
Белик В.П., 2006. Миграции серого журавля на юге России // Журавли Евразии: биология, охрана, разведение. Вып. 2. М. С.117–133.
Белик В.П., Гугуева Е.В., Ветров В.В., Милобог Ю.В., 2011. Красавка в Северо-Западном Прикаспии: распространение, численность, успешность размножения // Журавли Евразии (биология, распространение, миграции, управление). Вып. 4. М. С. 157–174.
Белик В.П., Ильяшенко В.Ю., Ильяшенко Е.И., 2020. Журавль-красавка – индикатор экологического благополучия в степях России // Природа. № 4. С. 3–15.
Березовиков Н.Н., 2002. Новые данные о распространении и миграциях журавля-красавки в южных, центральных и восточных регионах Казахстана // Журавли Евразии (распространение, численность, биология). Вып. 1. М. С. 151–161.
Бидашко Ф.Г., Пак М.В., Сатыбаев С.М., Парфенов А.В., Буханько Г.А. и др., 2006. О распространении и состоянии численности журавля-красавки (Anthropoides virgo L.) в северо-западной части Республики Казахстан // Selevinia. Казахстанский орнитологический ежегодник. С. 201.
Бохра Д.Л., 2020. Лимитирующие факторы для журавлей на местах зимовки в Индии // Информационный бюллетень Рабочей группы по журавлям Евразии. № 15. С. 162–165.
Брагин Е.А., 2002. Современное состояние журавлей в Кустанайской области, Казахстан // Журавли Евразии (распределение, численность, биология). Вып. 1. М. С. 168–174.
Брагин Е.А., 2011. Серый журавль и красавка в Тургайском регионе (Казахстан) на рубеже 20 и 21 веков // Журавли Евразии (биология, распространение, миграции, управление). Вып. 4. М. С. 190–202.
Букреева О.М., 2003. Состояние популяций журавля-красавки и серого журавля в Калмыкии // Стрепет: фауна, экология и охрана птиц Южной Палеарктики. № 2. С. 42–62.
Варшавский С.Н., Варшавский Б.С., Гарбузов В.К., 1991. Важнейшие особенности распространения и численности журавля-красавки в Актюбинско-Мугоджарском районе и Северном Приаралье // Журавль-красавка в СССР. Алма-Ата: изд-во “Гылым”. С. 24–31.
Гаврилов Э.И., 1977. О численности и путях пролета журавля-красавки в Казахстане // Редкие и исчезающие звери и птицы Казахстана. Алма-Ата: изд-во “Наука” КазССР. С. 167–175.
Гаврилов Э.И., 1979. Сезонные миграции птиц на территории Казахстана. Алма-Ата: изд-во “Наука” КазССР. 256 с.
Гаврилов Э.И., Гисцов А.П., 1985. Сезонные перелеты птиц в предгорьях западного Тянь-Шаня. Алма-Ата: Наука. 223 с.
Гисцов А.П., 1991. Послегнездовые скопления журавлей-красавок в Павлодарской области // Журавль-красавка в СССР. Алма-Ата: изд-во “Гылым”. С. 148–149.
Горошко О.А., 2015. Динамика и современное состояние популяций журавлей в Даурии (Забайкалье, Монголия) // Журавли Евразии (биология, распространение, разведение). Вып. 5. М. С. 116–134.
Губин Б.М., Скляренко С.Л., 1991. Весенний пролет журавля-красавки у восточной кромки песков Кызылкум // Журавль-красавка в СССР. Алма-Ата: Наука. С. 138–139.
Даль С.К., 1954. Животный мир Армянской ССР. Т. 1. Позвоночные животные. Ереван: Академия наук Армянской ССР. 415 с.
Дементьев Г.П., 1952. Птицы Туркменистана. Ашхабад: Академия наук Туркменской ССР. 347 с.
Емельянов В.И., Савченко А.П., 1991. Журавль-красавка на юге Красноярского края // Журавль-красавка в СССР. Алма-Ата: изд-во “Гылым”. С. 51–55.
Ерохов С.Н., 2009. Орнитологические наблюдения в Актюбинской области летом 2008 г. // Казахстанский орнитологический бюллетень 2008. Алматы: “Tethys”. С. 37–40.
Забашта А.В., 2020. О времени добычи первого журавля, помеченного в Аскании-Нова // Русский орнитологический журнал. № 29 (экспресс-выпуск 1906). С. 1528–1531.
Завьялов Е.В., Шляхтин Г.В., Хрустов И.А., Якушев Н.Н., 2003. Современное распространение и особенности экологии журавля-красавки в Саратовском Заволжье // Беркут. № 12 (1–2). С. 76–82.
Иванов А.И., 1940. Птицы Таджикистана. М.-Л.: Издательство Академии наук СССР. 299 с.
Ильяшенко Е.И., 2013. Учеты красавки в Волгоградском Заволжье, Россия, в 2011 г. // Информационный бюллетень Рабочей группы по журавлям Евразии. № 12. С. 37–41.
Ильяшенко Е.И., 2018. Динамика численности и ареала красавки в XX и начале XXI веков // Орнитология. № 42. С. 5–23.
Ильяшенко Е.И., 2019. Влияние изменений в агроландшафтах на распределение и численность красавки (Anthropoides virgo Linnaeus, 1758) (Gruidae, Aves) в гнездовой части ареала с середины XX века // Поволжский экологический журнал. Вып. 2. С. 222–236.
Ильяшенко Е.И., Ильяшенко В.Ю., 2011. Поиски красавки в Марокко // Информационный бюллетень Рабочей группы по журавлям Евразии. № 11. С. 38–40.
Ильяшенко Е.И., Белялов О.В., Ильяшенко В.Ю., Гаврилов А.Э., Зарипова С.Х. и др., 2018. Результаты мечения журавлей на оз. Тузколь, Казахстан, в 2017 г. // Информационный бюллетень Рабочей группы по журавлям Евразии. № 14. С. 89–101.
Ильяшенко Е.И., Ильяшенко В.Ю., Андрющенко Ю.А., Белик В.П., Гаврилов А.Э. и др., 2020. Предварительные результаты мечения красавки в 2018 и 2019 гг. // Информационный бюллетень Рабочей группы по журавлям Евразии. № 15. С. 112–134.
Ильяшенко Е.И., Ильяшенко В.Ю., Белик В.П., Корепов М.В., Арюлина И.П., Мнацеканов Р.А., 2020а. Обследование мест обитания красавки на юге азово-черноморской группировки красавки в 2018 и 2019 гг. // Информационный бюллетень Рабочей группы по журавлям Евразии. № 15. С. 31–38.
Ильяшенко Е.И., Ильяшенко В.Ю., Шуркина В.В., Постельных К.А., Мудрик Е.А., 2020б. Обследование мест обитания журавлей в Республике Хакасия в 2019 г. // Информационный бюллетень Рабочей группы по журавлям Евразии. № 15. С. 56–62.
Ильяшенко Е.И., Ильяшенко В.Ю., Белик В.П., Корепов М.В., Гугуева Е.В. и др., 2020в. Обследование мест обитания красавки в южных степях России и Западном Казахстане в 2018 и 2019 гг. // Информационный бюллетень Рабочей группы по журавлям Евразии. № 15. С. 38–50.
Кашкаров Д.Ю., Третьяков Г.П., Лановенко Е.Н., 1977. Наблюдения за весенней миграцией журавлей на юге Казахстана // Редкие и исчезающие звери и птицы Казахстана. Алма-Ата. С. 187–188.
Климов А.С., Самарин Е.Г., Сараев Ф.А., 1991. Распространение и места обитания журавля-красавки в Гурьевской области // Журавль-красавка в СССР. Алма-Ата: изд-во “Гылым”. С. 16–24.
Ковшарь А.Ф., 2007. Журавль-красавка // Птицы Средней Азии. Т. 1. Алматы. С. 343–348.
Ковшарь А.Ф., Березовиков Н.Н., 1991. Весенние скопления и перемещения журавля-красавки в долине р. Копа (Алма-Атинская область) // Журавль-красавка в СССР. Алма-Ата: изд-во “Гылым”. С. 130–135.
Ковшарь А.Ф., Давыгора А.В., 2004. Результаты российско-казахстанской экспедиции в Мугоджары и верхнюю часть бассейна Эмбы // Казахстанский орнитологический бюллетень 2003. Алматы. С. 32–34.
Ковшарь А.Ф., Давыгора А.В., Карпов Ф.Ф., 2007. Орнитологические наблюдения в Урало–Эмбинском междуречье (Эмба, Темир, Сагиз, Уил, Б. Хобда) в июне 2006 г. // Selevinia 2006: Казахстанский зоологический ежегодник. Алматы. С. 63–81.
Костин Ю.В., 1983. Птицы Крыма. М.: Наука. 241 с.
Кошкин А.В., 2007. Орнитологические наблюдения в Тенгизском регионе в 2006 г. // Казахстанский орнитологический бюллетень 2006. Алматы: “Tethys”. С. 41–44.
Лановенко Е.Н., Крейцберг Е.А., 2006. Ключевые территории для журавлей в Узбекистане и пути их охраны // Журавли Евразии (биология, охрана, разведение). Вып. 2. М. С. 175–176.
Лановенко Е.Н., Филатова Е.А., Филатов А.К., Шерназаров Э., 2011. О миграции журавлей в Южном Узбекистане в начале 21 века // Журавли Евразии (биология, распространение, миграции, управление). Вып. 4. М. С. 344–351.
Ляйстер А.Ф., Соснин Г.В., 1942. Материалы по орнитофауне Армянской ССР. Ереван. 402 с.
Матюхин А.В., Любущенко С.Ю., Мороз А.В., Ковальский А.И., 1991. Пролет журавля-красавки на крайнем юге Казахстана весной 1988 г. // Журавль-красавка в СССР. Алма-Ата: изд-во “Гылым”. С. 136–138.
Нанкинов Д.Н., 2009. История красавки Anthropoides virgo в Болгарии // Русский орнитологический журнал. № 18 (экспресс-выпуск 480). С. 695–704.
Саркисян А., 2011. Встречи серых журавлей и красавок в Армении // Информационный бюллетень Рабочей группы по журавлям Евразии. № 11. С. 46.
Судиловская А.М., 1951. Отряд журавли // Птицы Советского Союза. Т. 2. М. С. 97–138.
Султанов Э.Г., Керимов Т.А., Мамедов А.Ф., 2011. Результаты исследования журавлей в Азербайджане в начале XX века // Журавли Евразии: (биология, распространение, миграции, управление). Вып. 4. М. С. 312–316.
Торопова В.И., Еремченко В.К., 1980. Миграции птиц в урочище Кар-Кара (Киргизия) // Миграции птиц в Азии. Душанбе: Академия наук Таджикской ССР. С. 119–125.
Торопова В.И., Кулагин С.В., 2005. Миграционные скопления красавки в Кыргызстане // Информационный бюллетень Рабочей группы по журавлям Евразии. № 9. С. 40.
Торопова В.И., Кулагин С.В., 2011. Миграции журавлей в Кыргызстане // Журавли Евразии: (биология, распространение, миграции, управление). Вып. 4. М. С. 384–387.
Флинт В.Е., 1987. Семейство Журавлиные // Птицы СССР: Курообразные, журавлеобразные. Л.: Наука. С. 266–335.
Хан А., 2010. Орнитологический заказник Аб-и-Эстада // Атлас ключевых территорий для стерха и других околоводных птиц Западной и Центральной Азии. Барабу, США: Международный фонд охраны журавлей. С. 93–95.
Шевченко В.Л., Дебело П.В., Э.И. Гаврилов, Наглов В.А., Федосенко А.К., 1993. Об орнитофауне Волжско-Уральского междуречья // Фауна и биология птиц Казахстана. Алма-Ата. С. 7–10.
Abuladze A., 1995. Seasonal migrations of Demoiselle Cranes in Georgia // Crane Research and Protection in Europe. Proceedings of the Second European Crane Workshop. Germany, Halle-Wittenberg: Martin-Luther Univrsitat. P. 302–304.
Abuladze A., 2018. On the migration of Demoiselle Crane (Anthropoides virgo) across Georgia // IXth European Crane Conference. Abstracts. 3–7 December 2018, Arjuzanx. France, Arjuzanx. P. 3.
Ahmad A., Khurshid N., 1991. Observations of migration and migratory routes of cranes through Baluchistan // Natura. № 8. P. 9–11.
Akarsu F., ed., 2013. Biyo-Kültürel Açıdan Turna (Grus grus) Araştırması 2013. Doğa Derneği, Ankara, Türkiye. 56 p. (In Turkish).
Akarsu F., Ilyashenko E., Hays Branscheid C., 2013. Current status of cranes in Eastern Turkey // Proceedings of the VIIth European Crane Conference. Crane Conservation Germany, Groß Mohrdorf. P. 69–76.
Ashtiani M.A., 1999. Distribution and conservation of cranes in the Islamic Republic of Iran // Proceedings of the Third European Crane Workshop. Germany, Halle-Saale, Martin-Luther–Univ. P. 211–217.
Atta G.A.M., 1996. Status of cranes in Egypt // Proceedings 1993 African Crane & Wetland Training Workshop, Maun, Botswana. Baraboo, Wis., International Crane Foundation. P. 71–73.
Azar J.F., Al Hasani I., Nishimura K., 2007. The Demoiselle Crane, Anthropoides virgo (Linnaeus, 1758) (Aves: Gruidae), new to Jordan // Zoology in the Middle East. № 41. P. 109–110.
Banerjee A.K., Gopakumar G., 1986. Effect of drought on the wintering cranes in Rajkot, Saurashtra // Cheetal. № 27 (3). P. 31–37.
Basalingappa S., Reddy M.S., Mathapathi S.B., Muralidhar K.S., 1989. Some observations on the appearance and foraging of Demoiselle Cranes (Anthropoides virgo) in Dharwad and other parts of Karnataka, India // Workshop on Cranes and Wetland Research. Abstracts, Pune, India, 21–24 January 1989. P. 3.
Beilfuss R.D., Dodman T., Urban E.K., 2007. The status of cranes in Africa in 2005 // Ostrich. № 78 (2). P. 1–4.
Berg B., 1930. To Africa with the Migratory Birds. NY: G.P. Putnam’s Sons. P. 1–274.
Bijlsma R.G., 1991. Migration of raptors and Demoiselle Cranes over central Nepal // Birds of Prey Bulletin. № 4. P. 73–80.
Flint P., 1971. A reappraisal of the relative status of the Common Crane Grus grus and the Demoiselle Crane Anthropoides virgo on autumn passage through Cyprus // Cyprus Ornithological Society: First Bird Report. P. 84–91.
Flint P., 1972. Observations of the autumn crane passage at Akrotiri, Cyprus // Cyprus Ornithological Society: 2nd Bird Report. P. 123–128.
Gavrilov E., Van der Ven J., 2004. About migration routes of Demoiselle Crane (Anthropoides virgo) in Asia // Миграции птиц в Азии. Вып. 12. С. 6–28.
Gebremedhin B., Demeke Y., Atsebeha T., Meressa B., 2009. Notable records of a wintering site of the Demoiselle Crane in Kaftasheraro National Park // African cranes, wetlands and communities. № 5. P. 9–15.
Guo Yumin, He Fenqi, 2017. Preliminary results of satellite tracking on Ordos Demoiselle Cranes // Chinese Journal of Wildlife. № 38 (1). P. 141–143.
Hedenström A., 2008. Adaptations to migration in birds: behavioral strategies, morphology and scaling effects // Philosophical Transactions of the Royal Society. № B 363. P. 287–299.
Howes A.R.M., 1989. Demoiselle Cranes in Israel // Bulletin of the Ornithological Society of the Middle East. № 22. P. 21.
Ilyashenko E.I., 2019. Demoiselle Crane (Anthropoides virgo) // Crane Conservation Strategy. Mirande C.M., Harris J.T., editors. Baraboo, Wisconsin, USA: International Crane Foundation. P. 383–396.
Jain P., Jeenagar B., Rajpurohit S.N., 2005. Demoiselle Crane (Anthropoides virgo): A Study of its Management and Conservation at Kheechan in Rajasthan State of India // WWF India and Nature & People Research and Support Group. New Delhi, India. 30 p.
Jan A., Ahmad A., 1995. Cranes in Pakistan // Crane Research and Protection of Europe. Proceedings of the Palearctic Crane Workshop in Tallinn. Halle–Wittenberg. P. 337–352.
Johnsgard P.A., 1983. Demoiselle Crane (Anthropoides virgo Linnaeus, 1758) // Cranes of the World. University of the Nebraska – Lincoln. P. 95–102.
Kanai Y., Minton J., Nagendran M., Ueta M., Auyrsana B., et al., 2000. Migration of Demoiselle Cranes in Asia based on satellite tracking and fieldwork // Global Environ Research. № 4 (2). P. 143–153.
Kasparek M., 1988. The Demoiselle Crane, Anthropoides virgo, in Turkey: distribution and population of a highly endangered species // Zoology in the Middle East. № 2. P. 31–38.
Ma J., Ma Y., 2001. The status and conservation of cranes in China // Cranes in East Asia: Proceedings of the Symposium Held in Harbin, People’s Republic of China, 9–18 June 1998. Fort Collins, Colorado, USA: Geological Survey, Midcontinent Ecological Science Center. P. 3‒9.
Mackworth-Praed C.W., Grant C.H.B., 1970. Demoiselle Crane // Birds of West Central and Western Africa. V. 1. London. P. 231.
Martens V.J., 1971. Zur Kenntnis des Vogelzuges im nepalischen Himalaya // Die Vogelwarte. № 26 (1). P. 113– 128.
Meine C.D., Archibald G.W., 1996. The Cranes: Status Survey and Conservation Action Plan. Gland, Cambridge: IUCN. 294 p.
Mudrik E.A., Ilyashenko E.I., Goroshko O.A., Kashentseva T.A., Korepov M.V., et al., 2018. The Demoiselle crane (Anthropoides virgo) population genetic structure in Russia // Vavilovskii Zhurnal Genetiki i Selektsii = Vavilov Journal of Genetics and Breeding. V. 22. № 5. P. 586–592. https://doi.org/10.18699/VJ18.398
Mundkur T., 1992. Observation of a colour banded Demoiselle Crane in Gujarat, India // Newsletter for Birdwatchers. № 32 (7–8). P. 8.
Newton S.F., 1996. Establishing a monitoring program for the African wintering population of the Demoiselle Cranes during their spring migration through Saudi Arabia // Proceedings 1993 African Crane and Wetland Training Workshop. Baraboo, Wisconsin: International Crane Foundation. P. 83–88.
Newton S.F., Simens P., 1993. A survey of African Demoiselle Cranes Anthropoides virgo migrating through Saudi Arabia in spring 1992 // Proceedings of the Eight Pan-African Ornithological Congress. P. 595–596.
Nikolas G., 1987. Distribution atlas of Sudan’s bids with notes of habitats and status. Bonn, Germany. P. 64.
Perennou C., Mundkur T., 1991. Special feature: Distribution of the Demoiselle crane in India // Newsletter for Birdwatchers. № 31. P. 9–10.
Pfister O., 1996. The Demoiselle Cranes in Khichan // Oriental Bird Club. № 24. P. 22–24.
Ranaghad H., Ebrahimi A.M., 2007. The status of Common Crane Grus grus and Demoiselle Crane Anthropides virgo in West Azerbaijan, northwest Iran // Podoces. West & Central Asian Ornithological Journal. № 2 (2). P. 154–155.
Ripley S.D., 1950. Birds from Nepal 1947–1949 // Journal of the Bombay Natural History Society. № 49. P. 355–417.
Scholte P., 1996. Conservation status of cranes in North Cameroon and Western Chad // Proceedings 1993 African Crane & Wetland Training Workshop, Maun, Botswana. Baraboo, Wis., International Crane Foundation. P. 153–156.
Teli J., Tatu K., 2004. Migratory crane survey in Gujarat State – 2002 // Proceedings of the Workshop “Cranes and community in Gujarat – holistic conservation strategy for future”. Gujarat, India. P. 15–18.
Urban E.K., Gichuki N.N., 1991. Recent Research and Conservation Activities with cranes in Africa // Proceedings of the 1987 International Crane Workshop. Baraboo, Wisconsin. P. 351–355.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал