

Зоологический журнал, 2021, T. 100, № 9, стр. 1007-1018
Ревизия малоизученной видовой группы жуков-листоедов Chrysolina (sahlbergiana) подрода Pezocrosita (Coleoptera, Chrysomelidae)
Ю. Е. Михайлов a, b, *, С. В. Решетников c
a Уральский федеральный университет
620002 Екатеринбург, Россия
b Уральский государственный лесотехнический университет
620100 Екатеринбург, Россия
c Русское энтомологическое общество
630111 Новосибирск, Россия
* E-mail: yuemikhailov@gmail.com
Поступила в редакцию 07.06.2020
После доработки 20.07.2020
Принята к публикации 24.07.2020
Аннотация
Подрод Pezocrosita, первоначально выделенный Г.Г. Якобсоном в роде Crosita, позже был перенесен в род Chrysolina и в настоящее время включает 38 видов. Pezocrosita разделен на 7 видовых групп, среди которых группа Chrysolina (sahlbergiana) остается самой малоизученной, хотя включает типовой вид подрода. У Ch. sahlbergiana нами показано существование двух подвидов. Дано переописание локального эндемика гор Северного Урала в Свердловской области Ch. hyperboreica Mikhailov, ранее известного по единственному самцу. Из Западного Алтая описан новый вид Ch. korgonica sp. n. Приведен определительный ключ видов группы, обсуждаются реликтовый характер ее разорванного ареала, его связь с тундростепными сообществами.
Подрод Pezocrosita был установлен Якобсоном (Jacobson, 1901) с типовым видом Crosita kuznetzowi Jacobson 1901 по первоначальному обозначению и монотипии. В другой работе Якобсон (Jacobson, 1901a) описал еще один вид, Crosita sahlbergiana Jacobson 1901, и поместил его в этот же подрод, ясно указав, что его типовым видом является C. kuznetzowi (с. 120). Обе работы вышли в разных журналах примерно в одно время, но для вида Crosita kuznetzowi и подрода Pezocrosita долгое время допускался разброс года описания с 1900 до 1902 (Mohr, 1966; Лопатин, 1970; Bienkowski, 2001). Точную дату указанной публикации в журнале “Труды Русского энтомологического общества”/ “Horae Societatis Entomologicae Rossicae” (в данном случае не всего тома, а оттисков этой статьи, согласно ст. 21.8 МКЗН) можно установить благодаря работе Кержнера (1984). Это декабрь 1900 г., что из-за смены календарного стиля и согласно ст. 21.3.1 МКЗН следует считать как 31 января 1901 г. Журнал “Öfversigt af Finska Vetenskaps-Societetens Förhandlingar”, где была описана C. sahlbergiana, вышел в 1901 г. Отдельные оттиски из него тоже существуют (Bienkowski, 2019), но для этого журнала данных о точных датах публикации нет, поэтому согласно ст. 21.3.1 МКЗН (Международный кодекс …, 2004) датой выхода тома нужно считать 31.12.1901 г.
Сам Якобсон (Jacobson, 1901, 1901a) отмечал, что оба описанных им вида по окраске и строению голеней стоят особняком в роде Crosita Motschulsky 1860. В этом роде все виды имеют металлическую окраску, а C. kuznetzowi и C. sahlbergiana внешне напоминают Chrysolina limbata (Fabricius 1775). Контканен (Kontkanen, 1957) на основании изучения формы эдеагуса предложил перенести этот подрод из рода Crosita в род Chrysomela Linnaeus 1758. Хотя намного раньше перенес свой вид C. sahlbergiana в род Chrysomela сам Якобсон (1925). Странно только, что в той же работе он никак не упомянул C. kuznetzowi. Перенос подрода Pezocrosita в род Chrysomela, окончательно оформленный Мором (Mohr, 1966), вызвал вторичную омонимию Chrysomela kuznetzowi (Jacobson 1901) с другим, ранее описанным тем же автором, видом Chrysomela kuznetzowi Jacobson 1897. На это обратил внимание Лопатин (1970) и дал Chrysomela kuznetzowi (Jacobson 1901) новое замещающее название C. jacobsoni Lopatin 1970. Оно в дальнейшем не пригодилось, т.к. Медведев (1982) синонимизировал C. kuznetzowi с C. sahlbergiana, впрочем, никак не пояснив свое решение.
Добавлять уже известные и вновь описанные виды в подрод Pezocrosita начали Мор (Mohr, 1966) и Лопатин (1970), а Беньковский (Bienkowski, 2001) довел число видов до 48, разделив на 9 видовых групп. Позже часть этих видов была перенесена в другие подрода – Allohypericia Bechyne 1950, Chalcoidea Motschulsky 1860, Jeanclaudia Mikhailov 2009 и Hypochalcoidea Bourdonné 2012. По мнению Бурдонэ (Bourdonné, 2012), подрод Pezocrosita должен включать всего 7 видов. В недавно вышедшем мировом обзоре рода Chrysolina Беньковский (Bienkowski, 2019) оставил в составе Pezocrosita sensu lato 38 видов и 7 видовых групп, указав на гетерогенность и возможный парафилетический характер этого подрода. Действительно, это единственный подрод Chrysolina, где обозначены видовые группы, ряд признаков в диагнозе подрода характерен только для отдельных групп, а в определительный ключ видовых групп (Bienkowski, 2019) пришлось включить представителей близких подродов Allohypericia и Jeanclaudia. Но при любом взгляде на подрод в нем присутствует группа наиболее близких к C. sahlbergiana видов, представляющая фактически Pezocrosita s. str.
В видовую группу C. (sahlbergiana) первоначально было включено 7 видов (Bienkowski, 2001), 5 из которых позже были перенесены в подрод Jeanclaudia. Остались в группе только C. sahlbergiana и С. medvedevi Lopatin 1970. Второй вид описан из низкогорного массива Кент в Центральном Казахстане, что на 1200 км юго-западнее типового местонахождения первого вида (Лопатин, 1970). Третьим видом группы стал C. hyperboreica Mikhailov 2002, единственный самец которого был найден одним из авторов Ю.Е. Михайловым в горах Северного Урала в 2000 г. Во время последующих экспедиций на этот же горный массив попытки снова найти этот вид не увенчались успехом, и лишь в 2016 г. была собрана серия жуков. Поскольку первоописание вида (Mikhailov, 2002) было сделано по единственному самцу, и в нем не был упомянут ряд признаков, необходимых для точного отнесения к подроду или видовой группе (Bienkowski, 2019), здесь будет дано переописание C. hyperboreica.
Дополнительным стимулом к написанию работы стала находка на Западном Алтае в 2019 г. экспедицией новосибирских энтомологов, в состав которой входил один из авторов С.В. Решетников, еще не описанного вида. Этот вид оказался очень близок к предыдущим, и тем самым стал четвертым видом группы.
МАТЕРИАЛ И МЕТОДИКА
В июле 2016 г. при обследовании вершин Кытлымского массива в горной части Свердловской обл. в рамках международной программы GLORIA (Global Observation Research Initiative in Alpine Environments) впервые с момента находки в 2000 г. единственного типового экземпляра C. hyperboreica этот вид был пойман снова. В этот раз уже было предположение, что кормовым растением является полынь норвежская (Artemisia norvegica), и при тщательном обследовании ее куртинок были найдены еще жуки и личинки C. hyperboreica.
Особенность видов этой группы состоит в том, что они активны в ночное время, поэтому наиболее эффективным оказался сбор при наступлении темноты с помощью фонаря.
Упомянутые в работе экземпляры, включая типовые, хранятся в следующих коллекциях: ЗИН – Зоологический институт РАН (Санкт-Петербург), ИСиЭЖ – Институт систематики и экологии животных СО РАН (Новосибирск), ЗМУХ – Зоологический музей Университета Хельсинки (Финляндия), КЛМ – коллекция Л.Н. Медведева (ныне передана в ЗИН), КСР – коллекция С.В. Решетникова (Новосибирск), КХК – коллекция Х. Киппенберга (Герцогенаурах, Германия), КЮМ – коллекция Ю.Е. Михайлова (Екатеринбург).
Изучение и измерения коллекционных экземпляров проводились под бинокулярным микроскопом МБС-10 с помощью окуляр-микрометра.
РЕЗУЛЬТАТЫ
Подрод Pezocrosita Jacobson 1901
Типовой вид Crosita kuznetzowi Jacobson 1901 [Chrysolina sahlbergiana jacobsoni (Lopatin 1970)], по первоначальному обозначению.
Chrysolina (Pezocrosita) sahlbergiana (Jacobson 1901a: 120)
Crosita kuznetzowi была описана Якобсоном (Jacobson, 1901) по единственному самцу из Узунжула (Uzun-shul), теперь в Аскизском р-не Хакасии, где находились золотой прииск и дача Петра Ивановича Кузнецова (Калеменева, 2018), в честь которого был назван этот вид.
Crosita sahlbergiana Jacobson 1901 была описана по серии экземпляров (7♂♂, 3♀♀) из окрестностей Минусинска (Jacobson, 1901a). Т.е. оба вида описаны из Минусинской котловины, но разделены долиной р. Енисей, представляющей значимую зоогеографическую границу. При описании второго вида Якобсон (Jacobson, 1901a) указал на его отличия от первого, оговорившись, что C. kuznetzowi может оказаться уродливым экземпляром. Хотя различия двух видов были указаны достаточно четкие, после того, как Медведев (1982) свел C. sahlbergiana и C. kuznetzowi в синонимы, к этому вопросу последующие авторы больше не возвращались, приняв предложенную трактовку (Bienkowski, 2001, 2019; Mikhailov, 2002; Kippenberg, 2010).
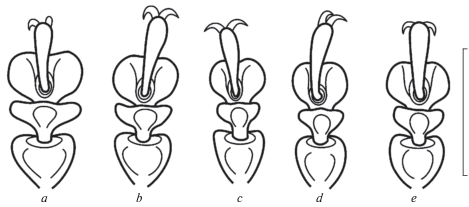
Однако в нашем распоряжении оказались экземпляры, которые являются топотипами обоих таксонов, описанных Якобсоном, и они имеют те же различия (см. определительный ключ ниже). А именно, самец из левобережной (хакасской) части Минусинской котловины (соответствует C. kuznetzowi) имеет явственно блестящие покровы, более правильную пунктировку надкрылий (рис. 1a), сильнее выраженную бороздку на передних голенях и очень сильно расширенные передние лапки (рис. 2a). В то же время, самцы из правобережной части Минусинской котловины (включая синтипы C. sahlbergiana) имеют слабо или шелковисто-блестящие покровы, менее правильную пунктировку надкрылий (рис. 1b), слабую бороздку на передних голенях и заметно расширенные передние лапки (рис. 2b). Самки обоих таксонов сверху матовые, но и у них обнаружились различия (самка C. kuznetzowi Якобсону известна не была). У самки из Хакасии голая линия снизу на первых члениках лапок широкая, а у самок из окрестностей Минусинска – более узкая. Т.е. оба описанных Якобсоном таксона хорошо различимы. Но поскольку по окраске и по форме эдеагуса значимых различий у экземпляров из Хакасии и Красноярского края не выявлено, их можно рассматривать в ранге подвидов.
Рис. 1.
Внешний вид представителей видовой группы Chrysolina (sahlbergiana): а – C. sahlbergiana jacobsoni, самец (Хакасия, окрестности п. Бирикчуль); b – C. sahlbergiana sahlbergiana, самец (Красноярский край, окрестности Минусинска); с – C. sahlbergiana sahlbergiana, самка (Красноярский край, окрестности Минусинска); d – C. medvedevi, голотип, самец (Казахстан, горы Кент ЮВ Каркаралинска); е – C. hyperboreica, голотип, самец (Свердловская обл., г. Серебрянский Камень); f – C. hyperboreica, типичная форма, самка; g – C. hyperboreica, красная форма, самец; h – C. hyperboreica, красная форма, самка (все Свердловская обл., Тылайско-Конжаковско-Серебрянский горный массив); i – C. korgonica, голотип, самец; j – C. korgonica, паратип, самка; k, l – C. korgonica, паратипы, самцы (все Алтайcкий край, Коргонский хребет). Масштаб 1 мм.

Chrysolina (Pezocrosita) sahlbergiana sahlbergiana (Jacobson 1901a)
Crosita (Pezocrosita) sahlbergiana Jacobson 1901a: 120 (“Fl. Jenissej: Ajatschinskaja, ad ripam Taganskoi protok et Minussinsk”, синтипы в ЗИН, ЗМУХ, КЛМ, КХК, изучены).
Chrysomela sahlbergiana: Якобсон, 1925: 26.
Pezocrosita sahlbergiana: Kontkanen, 1957: 89.
Chrysomela (Pezocrosita) sahlbergiana: Mohr, 1966: 90.
Chrysolina (Pezocrosita) sahlbergiana (partim): Медведев, 1982: 245; Медведев, Дубешко, 1992: 100; Bienkowski, 2001: 182 (nota); Mikhailov, 2002: 70; Kippenberg, 2010: 410 (nota); Bienkowski, 2019: 133 (nota), 503–504.
Материал. Синтипы, 2♂♂, 2♀♀, с этикетками: “Minusinsk”, “K. Ehnberg” или “Hammarstr.”, “Crosita (Pezocrosita) sahlbergiana Jacobs. typ. G. Jacobson det.” (ЗИН, ЗМУХ, КЛМ, КХК);
2♂♂, 2♀♀, Красноярский край, 5 км СЗ Минусинска, остепненные сопки, под куртинками Potentilla sericea, 31.VII.2002, Ю.Е. Михайлов (КЮМ).
1♀ без географической привязки с этикетками: “Coll. Mannh.”, “cruenta Gebler. ex Sibir”, “Ny för Samlingen” (ЗМУХ).
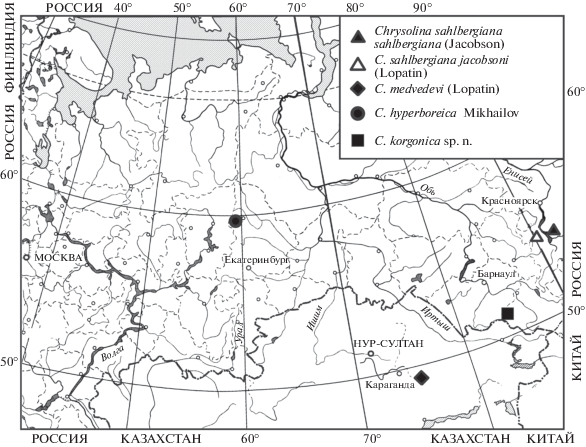
Распространение (рис. 3). Правобережная часть Минусинской котловины в Красноярском крае (окрестности Минусинска). Указания из Дархатской котловины в Северной Монголии (Медведев, 1982) требуют подтверждения.
Биотопы – каменистая степь на сопках периферии котловин, под куртинками Potentilla sericea и камнями (Михайлов, 2006).
Кормовое растение – полынь (Artemisia spp.) (Asteraceae). Питание жуков и личинок подтверждено в садках (Михайлов, 2006).
Этимология. Не указана автором (Jacobson, 1901a), однако из введения к статье с описанием ясно, что название дано в честь известного финского энтомолога Йона Зальберга (John Sahlberg) (1845–1920).
Chrysolina (Pezocrosita) sahlbergiana jacobsoni (Lopatin 1970) stat. resurr., comb. n.
Crosita (Pezocrosita) kuznetzowi Jacobson 1901: 78 (Uzun-shul).
Chrysomela (Pezocrosita) kuznetzovi: Mohr, 1966: 90 (неправильное последующее написание).
Chrysolina kusnetzovi: Kontkanen, 1957: 89; Медведев, 1982: 245 (неправильное последующее написание).
Chrysomela (Pezocrosita) jacobsoni Lopatin, 1970: 183 nom subst. pro Chrysomela kuznetzowi Jacobson, 1901a non Chrysomela kuznetzowi Jacobson, 1895.
Chrysolina jacobsoniana Medvedev & Korotyaev 1976: 243 (неправильное последующее написание).
= Chrysolina (Pezocrosita) sahlbergiana: Медведев, 1982: 245.
Chrysolina (Pezocrosita) sahlbergiana (partim): Медведев, 1982: 245; Медведев, Дубешко, 1992: 100; Bienkowski, 2001: 182 (nota); Mikhailov, 2002: 65, 66 (рисунки переднеспинки и передней лапки), 70; Kippenberg, 2010: 410 (nota); Bienkowski, 2019: 133 (nota), 503–504.
Материал. Голотип (?), 1♂ с этикетками: “1150”, “Cr. (Pezocrosita) rufolimbata G. Jacobs. typ. G. Jacobson det.” (кЛМ).
1♂, 1♀, Респ. Хакасия, Аскизский р-н, 8 км ЮВ п. Бирикчуль, степные склоны, 18–19.VII.1990, С.Э. Чернышев, Д.В. Логунов (ИСиЭЖ).
Распространение (рис. 3) – левобережная часть Минусинской котловины (Аскизский р-н Хакасии).
Биотопы – остепненные склоны сопок.
Кормовые растения – вероятно, полынь (Artemisia spp.).
Этимология. Замещающее название дано в честь Г.Г. Якобсона. Первоначально таксон был назван в честь Петра Ивановича Кузнецова, у которого во время экспедиции по Южной Сибири в 1897 г. останавливался брат Г.Г. Якобсона Алексей.
Chrysolina (Pezocrosita) medvedevi (Lopatin 1970)
Chrysomela (Pezocrosita) medvedevi Lopatin 1970: 186 (Казахстан: Карагандинская обл.: горы Кент на юго-запад от Каркаралинска, 1464 м, голотип в ЗИН, изучен); Арнольди, Медведев, 1969: 413 (nomen nudum); Арнольди, Медведев, 1975: 1636, 1639 (экология).
Chrysolina (Pezocrosita) medvedevi: Лопатин, 1977: 152; Лопатин, Куленова, 1986: 105; Bienkowski, 2001: 182 (nota); Mikhailov, 2002: 67 (рис.), 70; Kippenberg, 2010: 410 (nota); Bienkowski, 2019: 133 (nota), 503–504.
Материал. Голотип, самец, с этикетками: “г. Кент SO Каркаралинска Караг., Г. Медведев, 21 VII 962” [печатн.], “Chrysom. (Pezocrosita) medvedevi sp.n. [рукописн.] I.K. Lopatin det. 1967 [напечат.]”, “Holotypus [напечат.] Chrysomela (Pezocrosita) medvedevi Lopatin [рукописн.]”, Chrysolina (Pezocrosita) medvedevi (Lopatin 1970), Yu. Mikhailov det., 2010 (ЗИН).
Распространение (рис. 3). Локальный эндемик горного массива Кент (Каркаралинско-Кентский горный узел) в Карагандинской обл. Казахстана. Известен только из типового местонахождения (Арнольди, Медведев, 1969, 1975; Лопатин, 1970).
Биотоп – скалистые и щебнистые вершинные плато в наиболее возвышенной юго-восточной части горного массива на высотах 1300–1464 м (Арнольди, Медведев, 1975).
Кормовые растения – вероятно, полынь Artemisia spp.
Этимология (Лопатин, 1970). Вид назван в честь известного специалиста по жукам-чернотелкам, сотрудника ЗИН РАН, профессора Глеба Сергеевича Медведева (1931–2009).
Chrysolina (Pezocrosita) hyperboreica Mikhailov 2002
Chrysolina (Pezocrosita) hyperboreica Mikhailov, 2002: 64 (Северный Урал: горный массив Конжаковский Камень, г. Серебрянский Камень, голотип в ИСиЭЖ); Kippenberg, 2010: 413 (nota); Bienkowski, 2019: 132 (nota), 503–504.
Материал. Голотип, самец с этикетками: “Сев. Урал, массив Конжак. Камень, юж. отрог г. Серебрянский Камень, г. тундра, 9.VIII.2000, Ю.Е. Михайлов leg.”, “HOLOTYPE Chrysolina hyperboreica n. sp. Yu. Mikhailov det., 2000” (ИСиЭЖ).
1♀, Свердловская обл., Тылайско-Конжаковско-Серебрянский массив, вершинное плато г. Трапеция, h = 1200 м, горная тундра, почвенные ловушки, 25.07.2016, Ю.Е. Михайлов leg. (КЮМ); 2♂♂, 3♀♀, там же, западный склон горы Трапеция, h = 1150 м, горная тундра, в куртинках Artemisia norvegica, 25.07.2016, Ю.Е. Михайлов leg. (КЮМ); 2♂♂, 3♀♀, там же, восточный отрог горы Трапеция, h = 1150 м, горная тундра, в куртинках Artemisia norvegica, 29.07.2016, Ю.Е. Михайлов leg. (КЮМ).
Описание. Самцы удлиненно-овальные, шелковисто блестящие, длина тела (мм) 6.2–7.1, ширина 4.0–4.3. Самки продолговато-яйцевидные, матовые, длина тела (мм) 6.9–8.3, ширина 4.2–5.0. Надкрылья бронзовые, голова, переднеспинка и щиток бронзово-зеленые (бронзовая форма) (рис. 1e–1f) или надкрылья кирпично-красные с зачерненным швом и точками пунктировки, переднеспинка темно-бурая (красная форма) (рис. 1g–1h). Ноги черные, усики, челюстные щупики и лапки темно-бурые, 1-й и 2-й членики усиков снизу и коготки рыжие.
Наличник густо и мелко пунктирован, лоб в более редкой пунктировке. Фронтальный шов слабо прочерчен, углублен только по краям, эпикраниальный шов намечен только в верхней половине (у самок часто вообще не выражен). Последний членик челюстных щупиков у самцов поперечный, топоровидно расширенный, с прямо срезанной вершиной, в 1.2 раза шире длины, в 1.6 раза длиннее и в 1.5 раза шире предыдущего членика; у самок – продолговато-овальный, с прямо срезанной вершиной, в 1.3 раза длиннее ширины, в 1.7 раза длиннее и в 1.1 раза шире предыдущего членика.
Антенна прикреплена в 1.4 раза ближе к наличнику, чем к глазу. Отношение длины 1–3 члеников усиков как 8 : 4 : 6. 10-й членик в 1.2 раза длиннее ширины, 11-й членик – в 2 раза. Орбитальные линии выражены и углублены только в верхней четверти глаза.
Переднеспинка в 1.8–1.9 раза шире длины, с наибольшей шириной перед серединой, диск слабо равномерно выпуклый, заметно шагренированный, густо и мелко пунктированный, кроме гладкой средней линии (размер пунктировки диска не отличается у самцов и самок). Бока округлены только в дистальной части, а в основной части почти прямолинейно сужаются, редко у самок могут быть слегка выемчатые. Ширина между передними углами в 1.3 раза меньше ширины в основании у обоих полов. Передние углы умеренно выдающиеся, округло-треугольные, базальные углы почти прямые или тупоугольные у самцов, а у самок могут быть приострены. Передний край окаймлен и широко вырезан, покрыт многочисленными короткими щетинками; задний край посредине заметно дуговидно выступающий. Передние шетинконосные поры отсутствуют. Боковые валики широкие, пунктированы несколько мельче, но так же густо как диск; у обоих полов в основании составляют от 1/4 до 1/3 ширины переднеспинки. Боковые вдавления дуговидные, непрерывные и отчетливые по всей длине, состоят из полностью слившихся грубых точек, а в базальной четверти или трети имеют отвесные стенки и сильно углублены.
Проплевры продольно выпуклые, снаружи с сетью поперечных и продольных морщинок, без наружного окаймления, базальная складка неглубокая. Межтазиковый отросток простернума у обоих полов со слабым продольным углублением, покрытым крупными грубыми точками. Длины простернума и метастернума равны. Метастернум глубоко окаймлен на переднем крае, он в 1.4 раза короче первого стернита брюшка.
Щиток треугольный с округленной вершиной, одинаковой длины и ширины, либо в 1.2 раза шире длины, пунктирован так же, как и диск переднеспинки. Надкрылья в основании едва шире основания переднеспинки, максимально расширены перед серединой, без плечевого бугорка, в 2.2–2.3 раза длиннее ширины (у обоих полов). Первичная пунктировка надкрылий мелкая, у самцов несколько крупнее, чем у самок, образует попарно сближенные ряды, которые частично маскируются густой вторичной пунктировкой; вторичная пунктировка лишь в 1.5 раза мельче первичной, широкие четные междурядья пунктированы заметно чаще узких нечетных. Почти все точечные ряды в разной степени спутанные, особенно ряды 4–7 за серединой, где ряды 5 и 6 могут даже не прослеживаться. Прищитковый ряд короткий, из 7–9 точек. Междурядья плоские, изредка только третье междурядье слабо выпуклое. Точки боковой бороздки крупнее и реже, чем в рядах надкрылий. Пришовная бороздка отсутствует. Эпиплевры наклонные, видны по всей длине, в мелких морщинках. Густые, короткие щетинки расположены лишь в дистальной части эпиплевры. Крылья рудиментарные, узкие, лишь едва заходят за задний край метастернума.
Передние лапки самцов умеренно расширены (рис. 2d), в 2.3 раза длиннее ширины, отношение ширины 1–3 члеников передних лапок как 3.1 : 2.8 : 3.4. Волосяная щетка на подошве всех лапок самцов цельная. Лапки самок узкие, в 2.5–2.7 раза длиннее ширины, отношение ширины 1–3 члеников передних лапок как 2.4 : 2.0 : 2.7, 1-е членики всех лапок снизу с узкой голой линией.
Пигидий со слабо углубленным широким вдавлением в базальной трети у обоих полов. Первый стернит брюшка широко окаймлен на переднем крае, выступ между задними тазиками покрыт крупной грубой и морщинистой пунктировкой, остальная поверхность пунктирована мелко и редко. Последний стернит брюшка самца посредине очень слабо выпуклый, почти плоский, пунктирован рядом точек среднего размера только вдоль заднего края, по бокам слегка вдавлен и покрыт морщинками; его вершина окаймлена и слегка дуговидно вырезана. Последний стернит брюшка самки равномерно выпуклый, редко пунктирован в задней части точками среднего размера, в передней части – мелкими; его вершина прямо срезана. Эдеагус (рис. 4с) латерально слабее изогнут, его вершина коротко сужена.
Рис. 4.
Эдеагус сверху (слева) и сбоку (справа): а – Chrysolina sahlbergiana sahlbergiana, топотип (окрестности Минусинска); b – C. medvedevi, голотип; с – C. hyperboreica, голотип; d – C. hyperboreica (гора Трапеция); e, f – C. korgonica, голотип и паратип. Масштаб 1 мм.
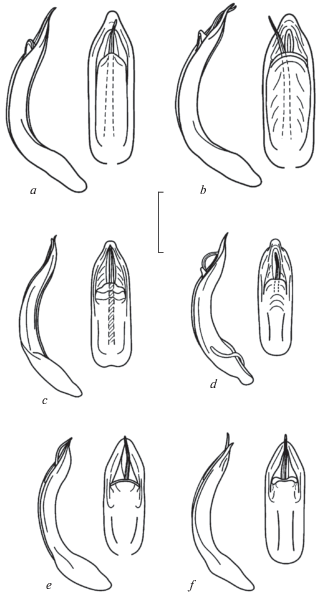
Дифференциальный диагноз. От C. sahlbergiana и C. medvedevi отличается более стройным (узким) телом и полностью металлической окраской (у типичной формы); боковые валики переднеспинки широкие, отделены от диска сплошными неглубокими бороздками, которые в базальной четверти сильно углублены и имеют отвесные стенки (рис. 1e–1h). Эдеагус имеет общую для группы характерную форму и флагеллум, но заметно меньше и ýже, отличается слабым латеральным изгибом и формой вершины (рис. 4с–4d).
Распространение (рис. 3). Североуральский горно-тундровый вид. Локальный эндемик Кытлымского (Тылайско-Конжаковско-Серебрянского) горного массива в Свердловской обл.
Биотопы – мохо-травяно-кустарничковая горная тундра.
Кормовое растение – Artemisia norvegica (Asteraceae). В куртинках этого растения были собраны жуки и личинки, питание подтверждено в садках.
Этимология (Mikhailov, 2002). Название происходит от Гиперборейских гор, древнего названия северной части Уральского хребта.
Chrysolina (Pezocrosita) korgonica Mikhailov et Reshetnikov sp. n.
Материал. Голотип, самец с этикетками: “Алтай, Коргонский хр., верх. р. Кума, 10 км ЮЗ с. Усть-Кумир, 2150–2200 м, 6.07.2019, А.А. Гурина, Р.Ю. и Е.Р. Дудко”, “HOLOTYPE Chrysolina korgonica n. sp. Yu. Mikhailov et S. Reshetnikov design., 2019” (ИСиЭЖ); паратипы: 14♂♂, 1♀, там же, 6.07.2019, А.А. Гурина, Р.Ю. и Е.Р. Дудко leg. (КЮМ); 13♂♂, 8♀♀, там же, 6.07.2019, С.В. Решетников leg. (КСР).
Описание. Самец (голотип) продолговато-овальный, длина тела 7.4 мм, ширина 4.7 мм. Надкрылья темно-зеленые, голова, переднеспинка и щиток темно-бронзовые. Ноги черные, усики, челюстные щупики и лапки темно-бурые, 1-й и 2-й членики усиков снизу и коготки рыжие.
Наличник густо и крупно пунктирован, лоб в такой же крупной пунктировке. Фронтальный шов слабо прочерчен, углублен только по краям, эпикраниальный шов намечен только в верхней половине. Последний членик челюстных щупиков поперечный, топоровидно расширенный, с прямо срезанной вершиной, в 1.3 раза шире длины, в 1.6 раза длиннее и в 1.3 раза шире предыдущего членика.
Антенна прикреплена в 1.4 раза ближе к наличнику, чем к глазу. Отношение длины 1–3 члеников усиков как 9 : 5 : 7. 10-й членик длиннее ширины в 1.2 раза, 11-й членик – в 1.8 раза. Орбитальные линии узкие, углублены только в верхней четверти глаза.
Переднеспинка в 2.0 раза шире длины, с наибольшей шириной перед серединой, диск равномерно выпуклый, мелко шагренированный, густо пунктированный точками среднего размера. Бока широко и правильно округлены (рис. 1i). Ширина между передними углами в 1.4 раза меньше ширины в основании. Передние углы сильно выдающиеся, округленные, базальные углы слабо тупоугольные, почти прямые. Передний край широко окаймлен и покрыт щетинками, задний край посредине заметно дуговидно выступающий. Передние щетинконосные поры отсутствуют. Боковые валики широкие, пунктированы намного мельче, чем диск; в основании составляют 1/4 ширины переднеспинки. Боковые вдавления дуговидные, непрерывные и отчетливые по всей длине, состоят из полностью слившихся грубых точек, а в базальной трети имеют отвесные стенки. Проплевры продольно выпуклые, снаружи с неправильными поперечными морщинками, без наружного окаймления, базальная складка неглубокая. Межтазиковый отросток простернума покрыт крупными грубыми точками, без выраженного продольного углубления. Длины простернума и метастернума равны. Метастернум глубоко окаймлен на переднем крае, он в 1.25 раза короче первого стернита брюшка.
Щиток треугольный с заостренной вершиной, в 1.1 раза шире длины, гладкий, не пунктированный. Надкрылья в основании едва шире основания переднеспинки, максимально расширены за серединой, без плечевого бугорка, в 2.3 раза длиннее ширины. Первичная пунктировка надкрылий средней величины, плотная, образует попарно сближенные, слегка спутанные ряды, которые маскируются густой вторичной пунктировкой. Вторичная пунктировка ненамного мельче первичной, широкие четные междурядья пунктированы чаще и крупнее узких нечетных, покрыты морщинками. Прищитковый ряд короткий, плохо заметный, из 5–6 точек. Все междурядья плоские. Точки боковой бороздки крупнее и реже, чем в рядах надкрылий. Пришовная бороздка отсутствует. Эпиплевры наклонные, видны по всей длине, в мелких морщинках. Густые, короткие щетинки расположены лишь в дистальной части эпиплевры. Крылья рудиментарные, короткие, округленные, достигают лишь заднего края метастернума.
Все лапки умеренно расширены (рис. 2e), передние в 2.4 раза длиннее ширины, отношение ширины их 1–3 члеников как 3.2 : 3.0 : 4.0. Волосяная щетка цельная на подошве всех лапок.
Пигидий со слабо углубленным широким вдавлением в базальной трети. Последний стернит брюшка поперечно выпуклый, крупно и грубо пунктирован, его вершина окаймлена и слегка дуговидно вырезана. Эдеагус (рис. 4e–4f) латерально сильнее изогнут, не сужается дуговидно к вершине, которая широко округлена и имеет треугольную форму.
Изменчивость. Самцы. Длина тела (мм) 6.8–7.5, ширина 4.2–4.7. Преобладает окраска как у голотипа, реже весь верх фиолетовый с зеленым отливом или черно-бронзовый (рис. 1k–1l).
Самки (рис. 1j) продолговато-овальные или яйцевидные, матовые, длина тела (мм) 7.4–8.0, ширина 4.5–4.9. Надкрылья темно-зеленые, голова, переднеспинка и щиток темно-бронзовые. Реже весь верх фиолетовый с зеленым отливом или черно-бронзовый. Лапки узкие, 1-е членики всех лапок с узкой голой линией (пробором). Последний стернит брюшка слабо поперечно выпуклый, пунктирован точками среднего размера, его вершина очень слабо дуговидно вырезана.
Этимология. Название происходит от Коргонского хребта на Алтае, где был найден этот вид.
Среди бескрылых (с редуцированными крыльями) жуков-листоедов Chrysolina группа видов C. (sahlbergiana) в подроде Pezocrosita габитуально ближе всего к видовой группе C. (convexicollis), а в роде Chrysolina – к подроду Jeanclaudia Mikhailov 2006. Различия даны в ключе:
1(2) Надкрылья в спутанной пунктировке, иногда лишь со следами точечных рядов. Переднеспинка с сильно углубленной и расширенной боковой бороздкой лишь в основной трети. Лапки самцов слабо расширены …………Подрод Pezocrosita (видовая группа С. convexicollis)
2(1) Надкрылья с точечными рядами
3(4) Пигидий с глубокой продольной бороздкой в апикальной половине. Переднеспинка с передними щетинконосными порами, ее боковая бороздка узкая, слабо углубленная и непрерывная по всей длине, у некоторых видов – только в основной трети. Эдеагус без флагеллума либо флагеллум короткий и не выступающий …… Подрод Jeanclaudia
4(3) Пигидий без глубокой продольной бороздки в апикальной половине. Передние шетинконосные поры на переднеспинке отсутствуют. Боковая бороздка переднеспинки узкая, непрерывная по всей длине, умеренно или сильно углубленная. Флагеллум эдеагуса узкий, бичевидный, всегда выступает из вершинного отверстия.………………подрод Pezocrosita (видовая группа C. sahlbergiana)
Определительная таблица видов группы C. sahlbergiana
1(6) Переднеспинка черная, надкрылья с широкой красной каймой или почти полностью красные (рис. 1a–1d). Брюшко красное. Боковые валики переднеспинки широкие, выпуклые, отделены от диска умеренно углубленной бороздкой с отвесной наружной стенкой. У самок 1-й членик всех лапок снизу с более или менее широкой голой линией
2(5) Тело более продолговатое. Надкрылья черные, их широкое базальное и постепенно суживающееся кзади боковое окаймление, а также эпиплевры рыжевато-красные (рис. 1a–1c). Лапки самца сильно расширенные. Переднеспинка с правильно округленными боковыми краями. Эдеагус более узкий, параллельносторонний, плавно сужается к сильнее оттянутой вершине (рис. 4а)
3(4) У самцов верх шелковисто-блестящий, передние лапки сильно расширенные (рис. 2b). Голая линия снизу на первых члениках лапок самок средняя. Щиток полуовальный. Ряды пунктировки надкрылий более спутанные, у самцов точки в рядах менее крупные. Длина 6.0–7.5 мм (самцы), 7.5–8.2 мм (самки), ширина 4.1–4.9 мм (самцы), 5.0–5.5 мм (самки). Минусинская котловина в Красноярском крае, (?) Дархатская котловина в Северной Монголии ……………. C. sahlbergiana sahlbergiana (Jacobson 1901)
3(4) У самцов верх блестящий, передние лапки очень сильно расширенные (рис. 2а). Голая линия снизу на первых члениках лапок самок широкая. Щиток треугольный. Ряды пунктировки надкрылий правильные, у самцов точки в рядах более крупные. Длина 7.4–8.0 мм (самцы), 8.0 мм (самки), ширина 4.5–5.0 мм (самцы), 5.2 мм (самки). Минусинская котловина в Хакасии ……C. sahlbergiana jacobsoni (Lopatin 1970)
5(2) Тело более округлое. Надкрылья одноцветные, от кирпично-красных до красновато-коричневых, с узким черным окаймлением основания, боковых краев и шва (рис. 1d). Лапки у самца умеренно расширенные (рис. 2c). Боковые края переднеспинки слабо округленные, в средней части почти прямые. Щиток широко-треугольный (в 1.5 раза шире длины), блестящий, в редких точках, его вершина остроугольная. Первичная пунктировка надкрылий средняя, собрана в попарно сближенные ряды. Эдеагус более массивный и широкий, слабее сужается к менее оттянутой вершине (рис. 4b). Длина 7.0–8.0 мм (самцы), 7.0–9.0 мм (самки), ширина 5.4 (самцы). Центральный Казахстан: Карагандинская обл.: горный массив Кент ………..…………..C. medvedevi (Lopatin 1970)
6(1) Верх полностью металлической окраски, если же надкрылья кирпично-красные, то переднеспинка коричневая. У самок 1-й членик всех лапок снизу с узкой голой линией
7(8) Мельче, тело более стройное (узкое): длина 6.2–7.0 мм (самцы), 6.9–8.3 мм (самки), ширина 4.0–4.3 мм (самцы), 4.2–5.0 мм (самки). Переднеспинка ýже (в 1.8–1.9 раза шире длины), ее бока в основной части почти прямые, и к основанию слегка сужаются, а у самок могут быть слегка выемчатые. Надкрылья бронзовые, переднеспинка и щиток бронзово-зеленые (типичная форма) или надкрылья кирпично-красные с зачерненным швом и точками пунктировки, переднеспинка темно-бурая (красная форма). Боковые валики переднеспинки широкие, отделены от диска сплошными неглубокими бороздками, которые в базальной четверти сильно углублены. Эдеагус (рис. 4с–4d) латерально слабее изогнут, его вершина коротко сужена. Свердловская обл.: Тылайско-Конжаковско-Серебрянский горный массив ……………….. C. hyperboreica Mikhailov 2002
8(7) Крупнее (длина 6.8–7.5 мм (самцы),7.4–8.0 мм (самки), ширина 4.2–4.7мм (самцы), 4.5–4.9 мм (самки)), переднеспинка шире (в 2.0 раза шире длины), ее бока широко и правильно округленные. Надкрылья темно-зеленые, голова, переднеспинка и щиток черно-бронзовые; реже весь верх фиолетовый с зеленым отливом или черно-бронзовый. Эдеагус (рис. 4e–4f) латерально сильнее изогнут, его вершина треугольной формы и широко округлена. Алтайский край: Коргонский хребет ……..…........ Ch. korgonica sp. n.
ОБСУЖДЕНИЕ
Виды в группе Chrysolina (sahlbergiana) очень близки по строению эдеагуса (рис. 4) и ряду важных систематических признаков, но при этом легко различаются по окраске и форме тела (рис. 1). Представители видовой группы также экологически близки, хотя Chrysolina sahlbergiana встречается в каменистой степи на склонах невысоких сопок в Минусинской котловине, а C. medvedevi – на вершинных плато горного массива Кент в Центральном Казахстане на высоте около 1400 м над ур. м. При этом нужно учесть, что в горах Кент выражена высотная поясность и только в наиболее возвышенных частях есть каменистые полынные степи. Географически ближе к первым двум видам оказался описанный в данной работе C. korgonica, найденный в горах Западного Алтая на высоте 2200 м над ур. м. А наиболее удаленная от всех находка – это североуральский вид C. hyperboreica. Однако и эти виды предпочитают каменистые вершинные плато, разница лишь в том, что в горах Алтая и Урала это пояс горных тундр.
Впрочем, различия между современными горно-тундровыми и степными ландшафтами нивелируются, если учесть, что на обширных пространствах Голарктики в течение четырех холодных периодов (стадиалов) плейстоцена существовали криоаридные растительные сообщества, сочетавшие элементы тундровой и степной биоты. Во время сартанского стадиала (МИС-2) в конце плейстоцена так называемые тундростепи распространились почти по всей территории России (Матросова, 2009). В настоящее время от обширного в прошлом зонального биома остались лишь небольшие фрагменты, а виды жесткокрылых, которые в плейстоценовых отложениях северо-востока или юго-запада Сибири встречались совместно, теперь приурочены к различным, и даже весьма удаленным территориям (Жерихин, 2003; Gurina et al., 2019).
Классическими примерами тундростепных видов листоедов являются Chrysolina (Allohypericia) arctica L. Medvedev 1980 и C. (Chalcoidea) brunnicornis vrangeliani Voronova 1982 с о-ва Врангеля (Хрулева, 2009). Они относятся к другим подродам Chrysolina, чем рассматриваемая здесь видовая группа, но при этом также питаются на различных видах полыни.
Виды группы C. (sahlbergiana) также можно рассматривать в качестве реликтовой тундростепной группировки жуков-листоедов. Типовой вид группы описан из Аскизского р-на Хакасии, где по отрогам Абаканского хребта в Минусинской котловине как раз сохранились участки реликтовых типчаково-дриадовых тундростепей (Ершова, Маскаев, 1998–2000). Помимо этого, все известные виды группы – локальные эндемики, ареал которых ограничен одним горным массивом или межгорной котловиной, а расстояния между точками находок разных видов могут составлять более 1000 км (рис. 3). Характер столь сильно фрагментированного ареала (в Минусинской котловине, Казахском мелкосопочнике и Алтае) нельзя объяснить только сложностью сбора, но можно привлечь данные палеоэнтомологии. Ведь оказалось, что у значительной части видов жесткокрылых из позднеплейстоценовых комплексов юга Западно-Сибирской равнины в настоящее время ареалы смещены в северном, южном или восточном направлении. И виды, ареалы которых сместились на юг и восток, обитают сейчас в горных степях Центрального и Восточного Казахстана либо в межгорных котловинах Алтае-Саянской горной страны (Gurina et al., 2019).
Несколько особняком стоит C. hyperboreica из горных тундр Северного Урала. Но состав высокогорной флоры массивов Кытлымский и Денежкин Камень указывает на то, что в плейстоцене и на этих массивах существовали тундростепные сообщества, от которых в современных условиях сохранились только отдельные виды растений, чему способствовали особенности термического и водного режима ультраосновных пород (Куликов, 2002). А целый ряд видов членистоногих из реликтовых тундростепей на северо-востоке Азии в настоящее время отмечен в горах не только Южной Сибири, но и Урала (Хрулева, 2009). Среди жесткокрылых к тундростепным видам в горах Урала, кроме C. hyperboreica, можно отнести также листоеда Chrysolina (Lithopteroides) exanthematica gemmifera Motschulsky 1860 и жужелицу Carabus sibiricus Fischer von Waldheim 1822.
Список литературы
Арнольди Л.В., Медведев Л.Н., 1969. Состав и анализ фауны Западной части Центрально-Казахстанского мелкосопочника и пустыни Бетпакдала. Листоеды (Chrysomelidae) // Растительные сообщества и животное население степей и пустынь Центрального Казахстана. Ч. 1. Л.: Наука. С. 408–417.
Арнольди Л.В., Медведев Л.Н., 1975. Эколого-фаунистический обзор фауны листоедов (Coleoptera, Chrysomelidae) Центрального Казахстана // Зоологический журнал. Т. 54. Вып. 11. С. 1634–1642.
Ершова Э.А., Маскаев Ю.М., 1998–2000. Типчаково-дриадовые тундростепи (Festuca pseudovina + Dryas oxyodonta) // Электронный атлас “Биоразнообразие животного и растительного мира Сибири” [Электронный ресурс]. Режим доступа: http:// www.nsc.ru/win/elbib/bio/green/26.html. Дата обращения: 04.04.2020
Жерихин В.В., 2003. Избранные труды по палеоэкологии и филоценогенетике. М.: Товарищество научных изданий КМК. 548 с.
Калеменева Н., 2018. В Минусинскую степь // Газета “Хакасия”. № 133–134 от 19 июня 2018 г. [Электронный ресурс]. Режим доступа: http://gazeta19.ru/index.php/pechatnaya-versiya/item/14139-v-minusinskuyu-step. Дата обращения: 04.04.2020
Кержнер И.М., 1984. Даты публикации издания “Труды Русского энтомологического общества” и “Horae Societatis Entomologicae Rossicae”, 1861–1932 // Энтомологическое обозрение. Т. 63. Вып. 4. С. 849–856.
Куликов П.В., 2002. Эндемичные и реликтовые элементы во флоре массива Денежкин Камень и их значение для познания генезиса высокогорной флоры Урала // Экологические проблемы горных территорий. Материалы междунар. науч. конф. Екатеринбург: Академкнига. С. 89–93.
Лопатин И.К., 1970. Обзор видов рода Chrysomela, подрода Pezocrosita Jcbs. (Coleoptera: Chrysomelidae) из Средней Азии и Казахстана // Энтомологическое обозрение. Т. 49. Вып. 1. С. 183–196.
Лопатин И.К., 1977. Жуки-листоеды (Chrysomelidae) Средней Азии и Казахстана. Л.: Наука. 270 с.
Лопатин И.К., Куленова К.З., 1986. Жуки-листоеды (Chrysomelidae) Казахстана: определитель. Алма-Ата: Наука. 200 с.
Матросова Т.В., 2009. Изменение растительности и климата Северной Чукотки за последние 350 тыс. лет (по палинологическим данным осадков оз. Эльгыгытгын) // Вестник СВНЦ ДВО РАН. № 2. С. 23–30.
Медведев Л.Н., 1982. Листоеды МНР: определитель. М.: Наука. 304 с.
Медведев Л.Н., Дубешко Л.Н., 1992. Определитель листоедов Сибири. Иркутск: Изд-во Иркутск. гос. ун-та. 224 с.
Международный кодекс зоологической номенклатуры, 2004. Изд. четвертое: Пер. с англ. и фр. Второе, исправленное изд. русского перевода. М.: Товарищество научных изданий КМК. 223 с.
Михайлов Ю.Е., 2006. Преимагинальные стадии и экология Chrysolina (Pezocrosita) sahlbergiana (Coleoptera, Chrysomelidae) // Зоологический журнал. Т. 85. № 7. С. 906–909.
Хрулева О.А., 2009. Группировки членистоногих острова Врангеля в контексте особенностей современных и плейстоценовых тундростепных энтомофаун северо-востока Азии // Виды и сообщества в экстремальных условиях. Сборник, посвященный 75-летию академика Юрия Ивановича Чернова. Отв. ред. Бабенко A.Б. и др. Москва-София: Товарищество научных изданий КМК и PENSOFT. С. 124–167.
Якобсон Г.Г., 1925. Жуки-листоеды Южного Енисея // Ежегодник Гос. Музея им. Н.М. Мартьянова. Т. 3. Вып. 1. С. 1–52.
Bienkowski A.O., 2001. A study on the genus Chrysolina Motschulsky, 1860, with a checklist of all the described subgenera, species, subspecies, and synonyms (Coleoptera: Chrysomelidae: Chrysomelinae) // Genus. V. 12. Fasc. 2. P. 105–235.
Bienkowski A.O., 2019. Chrysolina of the world – 2019 (Coleoptera: Chrysomelidae). Taxonomic review. Livny: Mukhametov G.V. Publ. 919 p.
Bourdonné J.-C., 2012. Révision du genre Chalcoidea Motschulsky, 1860 avec description d’une espèce nouvelle, Chalcoidea (s. str.) hollandei (1ère partie) (Coleoptera, Chrysomelidae, Chrysomelinae) // Nouvelle Revue d’Entomologie (N.S.). V. 28. Fasc. 2. P. 99–181.
Gurina A.A., Dudko R.Yu., Prosvirov A.S., Tshernyshev S.E., Legalov A.A., Zinovyev E.V., 2019. Coleoptera assemblages from the Quaternary deposits of Kizikha river, the southernmost late Pleistocene insects of the West Siberian Plain // Invertebrate Zoology. V. 16. № 2. P. 165–182.
Jacobson G., 1901. Chrysomelidae Sibiriae occidentalis // Труды Русского энтомологического общества / Horae Societatis Entomologicae Rossicae. Т. 35. № 1–2. С. 73–102.
Jacobson G., 1901a. Symbola ad Cognitionem Chrysomelidarum Rossiae asiaticae // Öfversigt af Finska Vetenskaps-Societetens Förhandlingar. V. 43. P. 99–147.
Kippenberg H., 2010. Chrysomelinae // Catalogue of Palaearctic Coleoptera. Löbl I., Smetana A. (eds). V. 6. Chrysomeloidea. Stenstrup: Apollo Books. P. 390–443.
Kontkanen P., 1957. Zur Kenntnis der Gattung Crosita Motsch. (Coleoptera, Chrysomelidae) // Annales Entomologici Fennici. V. 23. № 2. P. 87–90.
Mikhailov Yu.E., 2002. Contribution to the knowledge of the subgenus Chrysolina (Pezocrosita Jacobson, 1901): three new species from Tuva and the Urals (Coleoptera, Chrysomelidae) // Nouvelle Revue d’Entomologie (N.S.). V. 19. P. 57–71.
Mohr K.-H., 1966. Bestimmungstabelle der Untergattung Pezocrosita Jacobson 1902 (Allohypericia Bechyne 1950) // Reichenbachia. V. 8. № 13. P. 89–104.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал