

Зоологический журнал, 2021, T. 100, № 9, стр. 984-1001
К систематике и экологии рода Leptostenus (Crustacea, Amphipoda) из озера Байкал, с описанием двух новых видов
a Иркутский государственный университет
664003 Иркутск, Россия
b Байкальский музей Иркутского научного центра СО РАН
664520 пос. Листвянка, Иркутской обл., Россия
* E-mail: zoozhurn@mail.ru
Поступила в редакцию 10.06.2020
После доработки 31.08.2020
Принята к публикации 31.08.2020
Аннотация
В результате ревизии монотипического рода Leptostenus из состава эндемичной фауны амфипод оз. Байкал детально переописан типовой вид L. leptocerus (Dybowsky 1874) и описаны два новых вида, L. westschevi sp. n. и L. solontsovii sp. n. Для видов этого рода характерно тонкое стройное тело с шипами на сегментах метасомы и уросомы, очень длинные антенны; стержни антенн 1 не менее чем в 2 раза толще стержней антенн 2, 3-й членик стержня очень короткий. В уроподах 3 ветви почти равной длины, с густыми перистыми щетинками, наружная двучленистая. Вид L. westschevi отличается крупными темными глазами, наличием на уроподах 1 и 2 только шипов (у других видов имеются перистые щетинки); задний край базиподитов переоподов 5–7 образует спускающуюся вниз большую закругленную лопасть, в 5-й паре достигающую конца исхиоподита. L. solontsovii имеет темные, но более узкие глаза; в базиподите переопода 5 задний край образует небольшую лопасть, спускающуюся лишь до середины исхиоподита. Для L. leptocerus характерны крупные неправильной формы глаза с рассеянным светлым пигментом; в базиподитах переоподов 5–7 крыловидный край сильно суживается в нижней части, спускающейся вниз, лопасти не образует. По образу жизни все три вида – нектобентические. L. leptocerus – наиболее глубоководный вид; особенно многочислен в местах выхода нефтяных, метановых и гидротермальных сипов на дне озера. L. westschevi найден в южной котловине Байкала в составе ночного миграционного комплекса возле крутого подводного склона и над обширной мелководной отмелью. L. solontsovii обнаружен при облове придонного слоя воды над банкой Дриженко в северной котловине. С учетом вновь описанных видов таксономическое богатство фауны амфипод оз. Байкал составляет 278 видов и 78 подвидов. Многие находки новых и редко встречающихся таксонов могут быть сделаны в участках озера с необычными геологическими и ландшафтными условиями.
Глубочайшее и древнейшее на планете оз. Байкал является настоящим биосферным феноменом, учитывая очень высокую степень эндемизма в ряде групп населяющей его фауны и уникальную для пресноводных водоемов структуру экосистемы, устроенную по “океаническому” типу. Из всех групп беспозвоночных, не считая инфузорий, амфиподы подверглись наиболее бурному автохтонному видообразованию, по числу эндемиков и субэндемиков превосходят все другие группы; из озера их известно 278 видов и 78 подвидов (всего 356 таксонов низшего ранга, включая два описанных в настоящей статье). При этом, по нашему предположению, по меньшей мере несколько десятков видов, обитающих в озере, еще неизвестны науке.
На протяжении многих десятилетий, после выхода последнего полного определителя по байкальским амфиподам (Базикалова, 1945), таксономическим ревизиям подвергались полиморфные роды амфипод, включающие достаточно большое число видов (здесь не имеются в виду имевшиеся предложения просто разделить “крупные” роды на более “мелкие”; см.: Тахтеев, 2000; Takhteev, 2019). Исключением стало описание рода Koshovia Bazikalova 1975, включавшего один вид.
Статья посвящена ревизии одного из монотипических байкальских родов – Leptostenus. Согласно типологической классификации амфипод Байкала, разработанной автором (Takhteev, 2019), этот род относится к семейству Pallaseidae Tachteew 2000. Он включал единственный вид L. leptocerus (Dybowsky 1874), глубоководный, нектобентический, встречавшийся в сборах достаточно редко. Морфологически он наиболее близок к глубоководным представителям рода Poekilogammarus Stebbing 1899 (Tachteew, 1995). Ниже приведены расширенный диагноз рода, полное переописание типового вида на основе синтипов, коллекционных и оригинальных материалов, описания двух новых видов, ключ для определения видов рода и анализ биотопической приуроченности таксонов рода Leptostenus.
Сокращения названий организаций, в которых хранятся коллекции: ZMH – Зоологический институт и Зоологический музей Гамбургского университета, Гамбург, Германия; ZMK – Научно-природоведческий музей Национальной академии наук Украины, Киев, Украина (Зоологический музей Киева); ЛИН СО РАН – Лимнологический институт Сибирского отделения РАН, Иркутск, Россия; КЗБ ИГУ – кафедра гидробиологии и зоологии беспозвоночных Иркутского государственного университета, Иркутск, Россия.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Род Leptostenus Bazikalova 1945
Типовой вид – Gammarus leptocerus Dybowsky 1874 (по первоначальной монотипии).
Диагноз. Тело тонкое, стройное, гладкое, не более 25 мм в длину. Сегменты метасомы с зачаточными шипиками или заметными шипами по заднему краю. Сегменты уросомы также с тонкими шипами на заднем крае. Межантеннальные лопасти короткие и высокие. Глаза крупные. Антенны очень длинные, в 1-й паре длиннее тела, не менее чем в 2 раза длиннее антенн 2. Стержни антенн 1 значительно короче таковых антенн 2; базальный членик стержня антенны 1 цилиндрический, с суженными концами; 3-й членик стержня очень короткий, с вогнутой дорсальной поверхностью. Членики проксимальной части жгута антенны 1 также очень короткие. В придаточном жгутике не менее 4–5 члеников. Стержни антенн 2 не менее чем в 2 раза тоньше стержней антенн 1, жгут их также тоньше, кальцеолы отсутствуют. Ротовые придатки заметно выдаются вниз. Концевой членик пальпуса мандибулы с сильно вогнутым краем, заострен к концу, имеет вид лезвия косы. Гнатоподы слабые, у самки ладони во 2-й паре очень узкие. Коксальные пластинки 1–4 пар короткие, закругленные, с редкими щетинками по краю. Переоподы длинные и тонкие, в особенности карпоподиты и проподиты в 5–7 парах. Уроподы 1 и 2 также длинные и тонкие, особенно их ветви. В уроподах 3 обе ветви с перистыми щетинками, наружная ветвь двучленистая, с длинным концевым члеником, почти равна по длине внутренней. Тельсон рассечен как минимум на 2/3 длины, ветви удлиненные и суженные к концам, несут по 2–3 апикальных шипа.
Ключ для определения видов рода Leptostenus
1 (4) Глаза темные (черные или темно-бурые), с плотно сомкнутыми фасетками, почковидные. В базиподите переопода 5 задний (крыловидный) край образует более или менее развитую, спускающуюся вниз лопасть (рис. 5, 8).
2 (3) На уроподах 1 и 2 имеются редкие перистые щетинки, на наружном крае ветвей во 2-й паре они образуют ряд (рис. 8). В базиподите переопода 5 крыловидный край развит в нижней части, образует небольшую спускающуюся вниз лопасть, доходящую до середины исхиоподита; в переоподах 6 и 7 он снизу плавно закруглен, лопасть едва заметная (рис. 8) …… L. solontsovii sp. n.
3 (2) Уроподы 1 и 2 только с шипами (рис. 6). В базиподитах переоподов 5–7 крыловидный край развит в нижней части, образует спускающуюся вниз закругленную лопасть во всех трех парах; в базиподите 5 эта лопасть достигает конца исхиоподита (рис. 5) ……………… L. westschevi sp. n.
4 (1) Глаза с рассеянным пигментом (расставленными фасетками), светлые у фиксированных экземпляров, широкие, неправильно-округлой формы (рис. 2). Уроподы 1 и 2 с довольно многочисленными перистыми щетинками на стержнях и ветвях (рис. 2). В базиподитах переоподов 5–7 крыловидный край сильно суживается в нижней части, спускающуюся вниз лопасть не образует (рис. 2, 3) …….….. L. leptocerus (Dybowsky 1874).
Рис. 1.
Leptostenus leptocerus (Dybowsky), самец 9.5 мм, из коллекции ZMH. C – голова, A2 – антенна 2, L – верхняя губа, PMD – пальпус (щупик) мандибулы, MX1 – максилла 1, MX2 – максилла 2, MXP – максиллипед, GN1 – гнатопод 1, SGN2 – ладонь гнатопода 2 (проподит, subchaela), EP – эпимеральные пластинки 2 и 3. Масштаб произвольный.
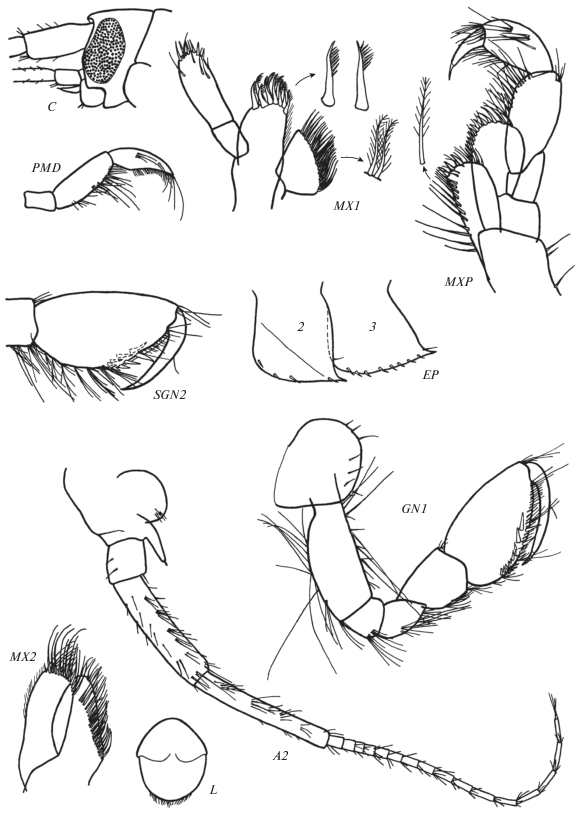
Leptostenus leptocerus (Dybowsky 1874) (рис. 1–3)
Gammarus leptocerus et G. leptocerus var. nematocerus. Dybowsky, 1874, S. 27–28, 85–86; Taf. VIII, fig. 2.
Echinogammarus leptocerus. Stebbing, 1899, p. 429. Stebbing, 1906, p. 490.
Abyssogammarus leptocerus. Совинский, 1915, с. 228–231, рис. 220–223 ; табл. XXXI , рис. 15, 16 .
Leptostenus leptocerus leptocerus et L. leptocerus nematocerus. Камалтынов, 2001, с. 672–673.
Leptostenus leptocerus. Базикалова, 1945, с. 133–134; табл. XV , фиг. 5. Barnard, Barnard, 1983, p. 473. Тахтеев, 2000, с. 332. Takhteev et al., 2015, p. 358.
Материал. Синтипы, 2 ♂♂. 1871 г. Оз. Байкал; предположительно – зал. Култук (Б.И. Дыбовский, В.И. Годлевский). Хранятся в ZMH, № 21 650 (Andres, Lott, 1977, S. 59).
4 ♂♂. 1871 г. Оз. Байкал (Б.И. Дыбовский, В.И. Годлевский). Хранятся в ZMH, № 37 960. Обнаружены мною в 1995 г. под № 21 696 с маркировкой “Carinogammarus rhodophthalmus var. micophthalma Dybowsky” и карандашной припиской “Paratypen”, пробирка была помечена “type”. В опубликованном каталоге типовых экземпляров (Andres, Lott, 1977) не значились. В пробирке оказались 3 экз. (♂♂) Echiuropus rhodophthalmus rhodophthalmus (Dybowsky 1874) и 4 экз. Leptostenus leptocerus.
Серия экземпляров, определенных В.К. Совинским из материалов экспедиции А.А. Коротнева (1900–1902 гг.) на оз. Байкал. Хранятся в ZMK, № 128, 275, 400 (просмотрены мною в 1990 г.).
2 экз. 25.10–03.11.1949. Оз. Байкал, напротив пос. Листвянка, глубина 200–300 м (А.Я. Базикалова). Хранятся в ЛИН СО РАН, № 414 (Механикова и др., 2010–2011).
1 экз. 17.11.1948. Оз. Байкал, напротив пади Баранчик, глубина 100 м (А.Я. Базикалова). Хранился в ЛИН СО РАН; в изданном каталоге (Механикова и др., 2010–2011) не значится; возможно, утерян.
Серия множества экземпляров. 14.08.1949. Оз. Байкал, напротив пос. Листвянка, глубина 200 м (А.Я. Базикалова). Хранились в ЛИН СО РАН; в изданном каталоге (Механикова и др., 2010–2011) не значатся; возможно, утеряны.
3 экз. 25.08.1991. Оз. Байкал, напротив бух. Фролиха, глубина 420 м, ил с примесью песка и растительного детрита (район рассеянного гидротермального высачивания). Сбор с глубоководного обитаемого аппарата (ГОА) “Pisces”. Хранятся на КЗБ ИГУ, № 21/101.
5 экз. 02.09.1991. Там же, глубина 410 м, черный ил с примесью растительного детрита (район рассеянного гидротермального высачивания). Сбор с ГОА “Pisces” с поверхности бактериальных матов. Хранятся на КЗБ ИГУ, № 21/102.
Описание. Длина тела особей с признаками пола 7–24 мм. Тело тонкое, гладкое. Все сегменты мета- и уросомы с шипами и единичными короткими щетинками по заднему краю. Голова сверху умеренно выпуклая; рострум короткий, покатый. Межантеннальная лопасть короткая и высокая, ее передний край прямой. Глаза очень крупные, неправильно-овальные, близко сходятся на дорсальной стороне головы и занимают большую часть ее боковой поверхности. Задний край глаза может быть с выемками (не всегда).
Антенны 1 не менее чем в 2–3 раза длиннее тела, по данным Механиковой и Ситниковой (Mekhanikova, Sitnikova, 2014) (концы жгутов у фиксированных особей почти всегда оторваны). При анализе видеозаписей эти же авторы отмечают, что антенны в 1–4 раза длиннее тела. В антенне 1 базальный членик стержня довольно толстый, цилиндрический, с суженными концами, равен длине головы или немного короче ее; 2-й членик стержня на 2/5 короче базального; 3-й членик очень короткий, сверху слегка вогнутый, его длина лишь немного превышает ширину. Жгут состоит более чем из 65 члеников, очень коротких в проксимальной части и более длинных в дистальной. В придаточном жгутике 5–10 члеников.
В антеннах 2 стержни значительно (более чем на треть) длиннее стержней антенн 1 и как минимум вдвое тоньше их. Антеннальный конус равен длине 3-го членика стержня, направлен прямо вперед; 5-й членик стержня немного длиннее 4-го или равен ему по длине, оба несут короткие щетинки. В жгуте 15–35 члеников, кальцеолы отсутствуют.
Верхняя губа плавно округленная. Пальпус мандибулы крупный, его концевой членик заостренный, серповидный, с выемкой по краю; щетка занимает более половины его длины; средний членик широкий; базальный членик без щетинок. В максилле 1 наружная пластинка снабжена косо-гребневидными иглами, “гребенка” хорошо развита, с многочисленными зубцами; внутренняя пластинка широкая, с многочисленными (не менее 20–22) перистыми щетинками. В максилле 2 внутренняя пластинка также с длинным косым рядом щетинок. В максиллипедах пальпус с длинным коготком, немного короче концевого членика; наружная пластинка с рядом зубчиков на внутреннем крае, на верхнем крае переходящих в щетинки; внутренняя пластинка с тремя зубцами на верхней стороне и перистыми щетинками на боковой.
Гнатоподы самца с ладонями умеренной ширины, толстыми в проксимальной части, с длинными тонкими когтями (дактилоподитами). Ладони гнатоподов 1 миндалевидные, достигают максимальной ширины в проксимальной трети; пальмарный край сильно скошен и плавно переходит в нижний. Ладони гнатоподов 2 наиболее широки в средней части, с более выраженным углом пальмарного края. Вдоль последнего в обеих парах по два ряда шипиков.
У самки ладони гнатоподов 1 узко-миндалевидные, со слабо выраженным углом пальмарного края. Ладони гнатоподов 2 бокаловидные – узкие, удлиненные, с коротким пальмарным краем, расположенным почти под прямым углом к нижнему краю.
Коксальные пластинки 1–4 пар короткие, с плавно округленными нижне-передними углами и с редкими, но не короткими щетинками.
Переоподы 3 и 4 с тонкими и длинными члениками, несущими немногочисленные редко расставленные щетинки; меро-, карпо- и проподиты почти равны по длине; когти длинные, тонкие, слабо изогнутые, достигают половины длины проподита.
В переоподах 5–7 пар базиподиты одинаковой формы, но последовательно увеличиваются в длину; их передний край несет шипики. Крыловидный край (утонченная задняя часть членика) неширокий, сужается книзу, не образует оттянутой вниз лопасти ни в одной из пар, может варьировать по ширине (см. рис. 2, 3). Прочие членики несут шипы, проподиты – еще и единичные очень длинные щетинки. Когти длинные, тонкие, равны 1/2–1/3 длины проподита. Длина переоподов 6 и 7 составляет 80–90% длины тела (Mekhanikova, Sitnikova, 2014), при анализе видеозаписей эти же авторы указывают, что “длина переоподов в 2 раза больше длины тела”, оговаривая, что это может быть лишь визуальным эффектом (с. 1509–1511).
Эпимеральные пластинки широкие, во 2-й и 3-й парах (либо только в 3-й) с оттянутыми в тонкое острие нижне-задними углами.
В уроподах 1 и 2 и стержни, и ветви вооружены шипами и довольно многочисленными перистыми щетинками, на стержне могут иметься также простые щетинки. Ветви уроподов 1 равной или почти равной длины, в уроподах 2 наружная на 1/3–1/4 короче внутренней. В уроподах 3 наружная ветвь двучленистая, ее второй членик длинный и тонкий; внутренняя ветвь немного длиннее основного членика наружной, стержень несет шипы на наружной стороне и щетинки на внутренней. Обе ветви вооружены шипами и перистыми щетинками по обоим краям, на концах ветвей сидят простые щетинки.
Тельсон рассечен до основания (по: Базикалова, 1945, на 3/4), ветви конические, несут шипы и щетинки на концах и часто по наружному или внутреннему краю; выемка широкая. У части экземпляров внутренние края ветвей вогнутые, и тельсон имеет клешневидную форму.
Окраска тела красновато-желтая (Dybowsky, 1874) или белая (Базикалова, 1945). На подводных видеозаписях из района мыса Горевой Утес (Mekhanikova, Sitnikova, 2014) видны только белые особи.
Примечание. Дыбовский (Dybowsky, 1874), помимо номинативной формы, выделял глубоководную разновидность “Gammarus leptocerus var. nematocerus”, отмечая у нее беловатую окраску и более тонкие придатки тела. Валидность ее в качестве подвида (Leptostenus leptocerus nematocerus) принимал Камалтынов (2001, с. 673), ссылаясь на неназванные различия в рисунках Дыбовского для форм номинативной и var. nematocerus. Однако мелко выполненные рисунки Дыбовского оказываются не всегда точными. Так, применительно к рассматриваемому виду, стержни антенн 1 и 2 на его рисунках (Dybowsky, 1874, Taf. VIII, fig. 2a, 3a) почти равны по толщине, что противоречит приводимому там же на с. 85 описанию. На основании этих рисунков и кратких замечаний Дыбовского нет возможности составить дифференциальный диагноз подвида L. l. nematocerus, и я воздерживаюсь от его признания в этом статусе. Различие же в окраске в зависимости от глубины обитания проявляется у разных видов байкальских амфипод, более глубоководные особи одного и того же вида обычно светлее.
Распространение и экология. Долгое время род Leptostenus считался не только монотипическим, но и крайне редко встречающимся. Дыбовский (Dybowsky, 1874) указывал глубины нахождения 150–670 м – скорее всего, для района пос. Култук, где он занимался исследованиями, отбывая политическую ссылку. Базикалова (1945) упоминала местонахождение у с. Лиственничного (ныне пос. Листвянка), глубину 877 м и илистый грунт. В дальнейшем на протяжении нескольких десятилетий не отмечалось ни одной находки. На этом основании автор данной статьи даже выдвигал предположение о том, что Leptostenus leptocerus является гибридным псевдовидом (Tachteew, 1995, p. 62).
Однако в дальнейшем вид был определен мною из сборов в районе подводного метансодержащего источника напротив бухты Фролиха (Северный Байкал) (Takhteev, 1997). Материал собран в 1991 г. манипулятором ГОА “Pisces”. На глубинах 410–425 м обнаружены целые скопления Leptostenus leptocerus (Тахтеев, 2009). В этом районе происходит рассеянный выход термальных вод, просачивающихся через илы, и формирование бактериальных матов, состоящих из бактерий-хемосинтетиков (Кузнецов и др., 1991; Гебрук и др., 1993).
Наконец, в 2008–2009 гг. исследования с ГОА “Мир-1” и “Мир-2” нефтяных и метановых сипов в районе мыса Горевой Утес (Средний Байкал, в интервале глубин 800–900 м) показали, что Leptostenus leptocerus является характерным и многочисленным видом в местах подводной разгрузки метана и нефти и может служить индикатором таких участков (Mekhanikova, Sitnikova, 2014). Пищей амфиподам, очевидно, служат бактериальные маты, которые также формируются в таких участках, и придонная взвесь. Наблюдения из ГОА показали, что этот вид является хорошим пловцом, однако для отдыха и питания ему требуется твердый субстрат (Mekhanikova, Sitnikova, 2014). На фото, приводимом авторами, а также на имеющихся видеозаписях видно, что особи L. leptocerus в состоянии покоя сидят в разных положениях практически на вертикальных поверхностях битумных построек. Вид в массе отмечен и плавающим над постройками (Mekhanikova, Sitnikova, 2014).
Таким образом, L. leptocerus обитает во всех трех котловинах Байкала: северной (напротив бухты Фролиха и мыса Фролова), средней (напротив мыса Горевой Утес) и южной (Култук, Листвянка), на глубинах от 150 до 900 м. Образ жизни вида – нектобентический. Спаривающиеся особи и самки с яйцами в выводковой камере визуально отмечены в ходе погружений на ГОА в августе 2008 и 2009 гг. (Mekhanikova, Sitnikova, 2014), однако период размножения может быть круглогодичным, как у ряда других глубоководных амфипод.
Leptostenus westschevi Tachteew sp. n. (рис. 4–6)
Рис. 4.
Leptostenus westschevi Tachteew sp. n.: GV – общий вид, без уроподов 3, жгут A1 показан частично (рис. Д.А. Батранина); C – голова, A1 – антенна 1 (жгут изображен частично), A2 – антенна 2 (жгут изображен частично), L – верхняя губа, MD – левая мандибула, MX1 – максилла 1, GN1 – гнатопод 1, CP + SGN1 – карпоподит и ладонь гнатопода 1. GV, C – паратип самец 13.5 мм, остальные элементы – голотип самка 13.3 мм. Масштаб: GV, C, A1, A2 – 1 мм, остальные элементы – 0.5 мм.
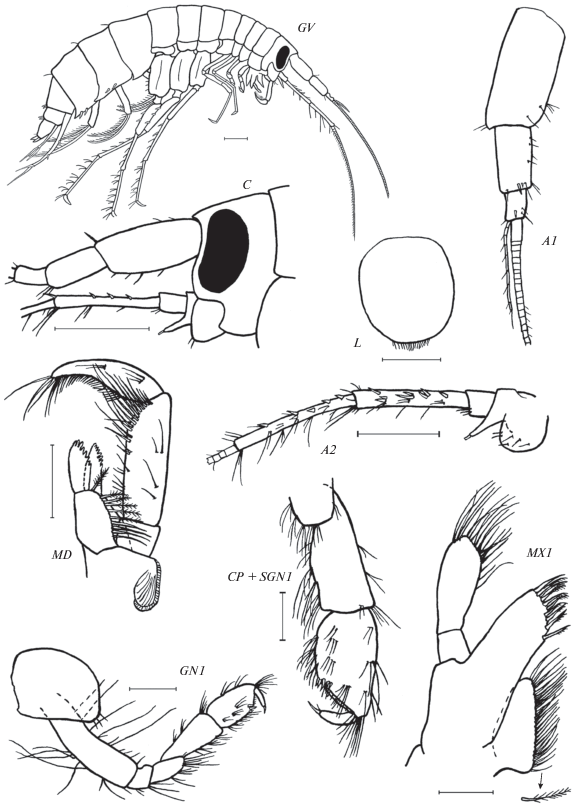
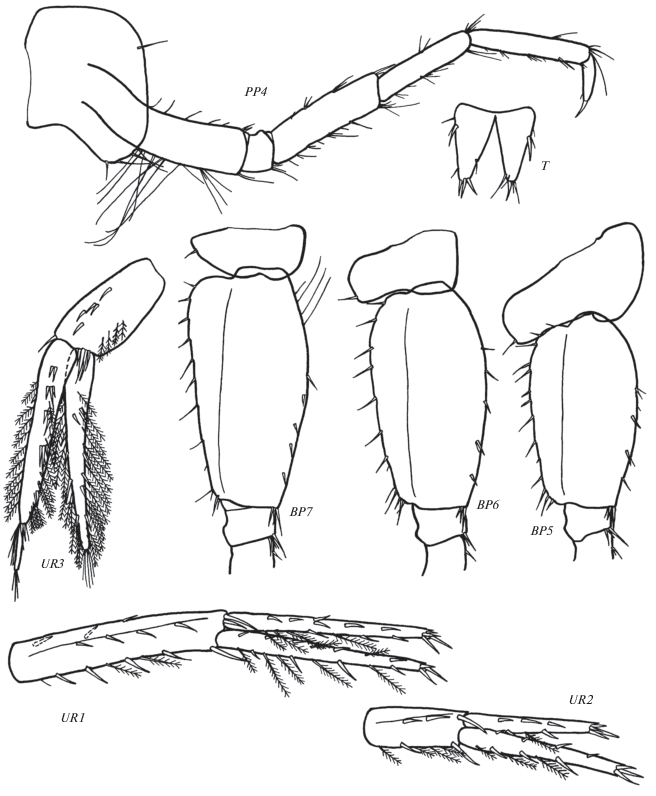
Рис. 5.
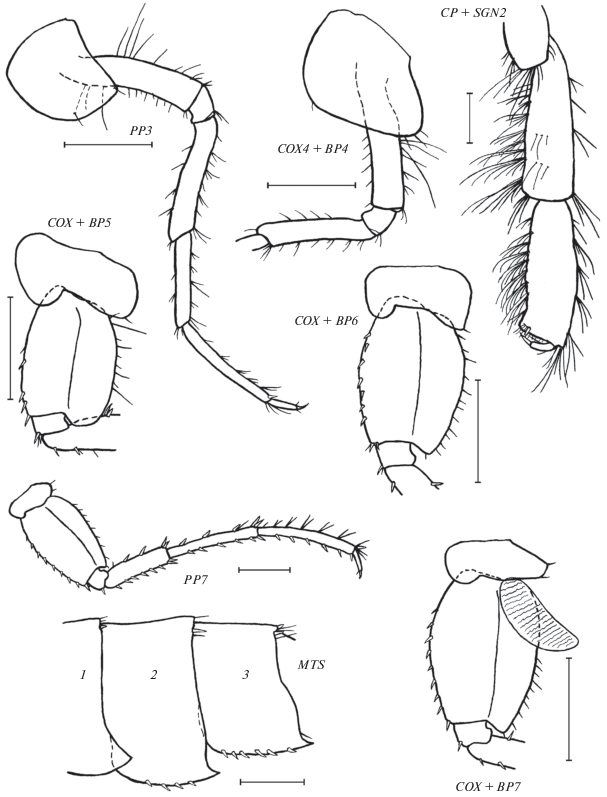
Leptostenus westschevi Tachteew sp. n., голотип самка 13.3 мм. CP + SGN2 – карпоподит и ладонь гнатопода 2, PP3 – переопод 3, COX4 + BP4 – коксальная пластинка и базиподит переопода 4, COX5 + BP5 – коксальная пластинка и базиподит переопода 5, COX6 + BP6 – коксальная пластинка и базиподит переопода 6, PP7 – переопод 7, COX7 + BP7 – коксальная пластинка и базиподит переопода 7, MTS – сегменты метасомы. Масштаб: CP + SGN2 – 0.5 мм, остальные элементы – 1 мм.
Материал. Голотип самка 13.3 мм, 1-й стадии развития, согласно классификации Базикаловой (1941), с зачаточными оостегитами. 02.03.2009. Оз. Байкал, бухта Большие Коты, перед падью Жилище, над уступом обрывистой скалы. Сеть Джеди, тотальный подледный лов, глубина от 38 до 0 м. Время 00:35 (В.В. Тахтеев, Н.А. Поляков, А.С. Мишарин). Хранится на КЗБ ИГУ, № 21/104.
Паратип ♂ 13.5 мм. 20.03.2014. Оз. Байкал, в том же месте. Сеть Джеди, тотальный подледный лов, глубина от 32 до 0 м. Время 07:25, начало рассвета (В.В. Тахтеев, Д.Ю. Карнаухов). Хранится на КЗБ ИГУ, № 21/105.
Паратип 1♂ 9.0 мм. 21.03.2014. Оз. Байкал, в том же месте. Сеть Джеди, тотальный подледный лов, глубина от 32 до 0 м. Время 01:30 (В.В. Тахтеев, Д.Ю. Карнаухов, И.А. Махов). Хранится в ЛИН СО РАН, № 1131.
Паратип 1♀ 19.0 мм, 2-й стадии зрелости, по классификации Базикаловой (1941) – с нормально развитыми оостегитами без щетинок. 14.07.2017. Оз. Байкал, напротив пос. Клюевка. Сеть Джеди, тотальный лов от дна до поверхности, глубина от 15 до 0 м, над пологой мелководной платформой. Грунт – слабо заиленный рыжий песок с примесью гальки, встречаются немногочисленные крупные глыбы (по данным видеосъемки и дночерпательной пробы). Время 23:35. НИС “Профессор Кожов” (В.В. Тахтеев, И.О. Еропова, Е.Б. Говорухина). Хранится на КЗБ ИГУ, № 21/106.
Описание. Длина тела особей с признаками пола 9.0–19.0 мм. Тело гладкое, стройное. Все сегменты мета- и уросомы по заднему краю несут редкие щетинки и короткие шипы, иногда едва заметные.
Голова короткая, ее дорсальный профиль прямой или слабо выпуклый, рострум очень короткий. Глаза крупные, черные, высокие, широко-почковидные или неправильно овальные, могут быть немного сужены в верхней либо нижней части, их задний край не изрезан.
Длина антенн 1 не менее чем вдвое превышает длину тела (концы жгутов оторваны у трех экземпляров из четырех имеющихся). Стержни антенн 1 примерно в 1.5 раза короче стержней антенн 2 и в 3 раза (базальный членик) превышают их по толщине. Членики стержня последовательно уменьшаются в длину и толщину, снабжены редкими короткими и тонкими щетинками; 2-й и 3-й членики могут иметь единичные слабые шипы. Базальный членик стержня утолщенный, цилиндрический, суженный к концам; 2-й членик на 1/3 короче базального, 3-й вдвое короче 2-го, слабо вогнутый сверху, его длина не более чем в 2 раза превышает ширину. В основном жгуте у самого крупного экземпляра более 82 члеников; в проксимальной части жгута членики очень короткие (длина меньше ширины), в дистальной части – длинные и тонкие. В придаточном жгутике от 5 до 8 длинных и тонких члеников, и короткий, рудиментарный концевой членик.
В антеннах 2 стержень тонкий, почти нитевидный. Антеннальный конус равен длине 3-го членика стержня или чуть короче, направлен вперед и немного вниз. 3-й членик умеренной длины, 4-й членик равен по длине 5-му или немного короче, несет шипы на верхней стороне и редкие щетинки на нижней. 5-й членик также с шипами на верхней стороне и пучками длинных слабых щетинок на нижней. В жгуте до 29 члеников, членики и щетинки на них удлиняются к дистальному концу.
Верхняя губа почти круглая, с тонкими волосками (сетулами) на дистальном крае. Левая мандибула с 8 зубцами на режущем крае и с 8 на режущей пластинке; в зубном ряду – 10 щетинок; пальпус с серповидно изогнутым концевым члеником, его нижний край сильно вогнут. В максиллах 1 пальпус слегка длиннее наружной пластинки, немного расширяется дистально, несет серию длинных щетинок; наружная пластинка с 11 иглами, имеющими одностороннее оперение; на внутренней пластинке 26 перистых щетинок. Максиллы 2 с густыми щетинками, в диагональном ряду на внутренней пластинке 27 перистых щетинок. В максиллипедах наружная пластинка достигает половины длины пальпуса, внутренняя на 1/3 короче. Коготок пальпуса равен 1/2 длины последнего членика.
Коксальные пластинки недлинные, с единичными короткими щетинками по краю; в 1–3-й парах с закругленным нижне-передним углом; в 4-й паре расширенные в нижней части, задний край закругленный, нижний прямой.
Гнатоподы слабые у самцов и самок, ладони в 1-й паре миндалевидные, во 2-й длинные и тонкие (“бокаловидные”), со слабыми граничными шипами. Карпоподиты в обеих парах удлиненные. Коготь 2-й пары слабый.
Переоподы 3 и 4 с удлиненными тонкими члениками и пучками недлинных щетинок по заднему краю; длинные щетинки имеются только в верхней части базиподита. Карпоподиты немного короче меро- и проподитов, равных по длине. Когти тонкие, слабо изогнутые.
Переоподы 5–7 длинные и тонкие, немного короче длины тела. В базиподитах крыловидный край широкий, образует хорошо развитую, спускающуюся вниз лопасть, особенно широкую в 5-й паре; задние края с рядом щетинок средней длины, 5-я пара с единичными длинными щетинками. Мероподиты немного короче карпо- и проподитов, равных по длине; меро- и карпоподиты несут шипы по обоим краям, могут иметься немногочисленные щетинки; проподиты снабжены шипиками по переднему краю и пучками длинных или средней длины щетинок по заднему. Когти удлиненные, острые, почти прямые.
Эпимеральные пластинки широкие, с заостренным нижне-задним углом, 2-я и 3-я пары с рядом шипиков вдоль нижнего края.
Уроподы 1 и 2 тонкие, стержни и ветви только с острыми шипами, щетинки отсутствуют (рис. 11 ). Ветви в 1-й паре почти равной длины, во 2-й наружная на 1/3–1/4 короче внутренней. Уроподы 3 хорошо развиты; ветви равной длины либо наружная слегка короче внутренней; наружная ветвь двучленистая, второй членик удлиненный, конический. Стержень несет единичные шипы, обе ветви – густые перистые щетинки по обоим краям, среди которых встречаются шипы и простые щетинки.
Тельсон удлиненный, рассечен почти до основания, ветви дистально сужены, выемка широкая. На концах ветвей по 2–3 шипа и 2–3 щетинки; единичные слабые шипы сидят по обоим краям ветвей.
Прижизненная окраска светлая, грязно-оранжевая или красновато-коричневая, задняя половина сегментов окрашена темнее передней; сегменты уросомы могут быть темно-красными.
Изменчивость. У экземпляра из района пос. Клюевка некрупные шипы сидят не только по заднему краю, но и на дорсальной поверхности сегментов метасомы в их задней половине. Уроподы 1 и 2 у него с гораздо бóльшим количеством шипов, чем у особей из района Больших Котов, сами шипы более длинные.
Дифференциальный диагноз. В отличие от L. leptocerus, L. westschevi имеет крупные широкие ярко-черные глаза. В отличие от L. solontsovii, 3-й членик стержня антенны 2 умеренной длины, в 1.5–2 раза превышающей ширину. В отличие от обоих названных видов, базиподиты переоподов 5–7 с широким крыловидным краем, который в нижней части образует оттянутую вниз лопасть, в 5-й паре достигающую конца исхиоподита. Щетинки на уроподах 1 и 2 совершенно отсутствуют. Тельсон не клешневидный, выемка с прямыми краями.
Распространение и экология. Все имеющиеся экземпляры отловлены в южной части Байкала, в бухте Большие Коты (северо-западный берег, в подледный период) и напротив пос. Клюевка (юго-восточный берег, летом), на глубинах от 15 до 38 м. Все они добыты планктонной сетью Джеди в темное время суток из состава ночного миграционного комплекса. Этот комплекс состоит из облигатных пелагобионтов и бентосных видов (в основном амфипод), поднимающихся в темное время суток в пелагиаль. При этом особи L. westschevi из района Больших Котов отловлены на одной и той же точке рядом с почти отвесным подводным обрывом, имеющим узкие “ступени”. Особь из района пос. Клюевка поймана над пологой мелководной платформой. Однако на указанной выше точке путем подводных видеонаблюдений и сетного лова мы неоднократно отмечали присутствие в ночных миграционных скоплениях облигатно пелагического вида амфипод Macrohectopus branickii (Dybowsky 1874) (Karnaukhov et al., 2016; Тахтеев и др., 2019; Батранин и др., 2019). Это свидетельствует о том, что в ходе суточных вертикальных миграций обитатели бóльших глубин способны проникать на меньшие. Вполне возможно, основная зона обитания L. westschevi, в которой он находится в дневное время, располагается глубже, чем были сделаны его ночные находки. Как и другие виды рода, он, очевидно, ведет нектобентический образ жизни, в пользу которого свидетельствуют длинные и тонкие переоподы 5–7.
Этимология. Вид назван в честь Сергея Николаевича Вещева, многолетнего капитана НИС “Профессор Кожов”, знатока научной навигации. В экспедициях с С.Н. Вещевым мы с коллегами исследовали самые труднодоступные участки дна Байкала и неоднократно добывали новые для науки таксоны фауны (амфипод, моллюсков, губок).
Leptostenus solontsovii Tachteew sp. n. (рис. 7, 8)
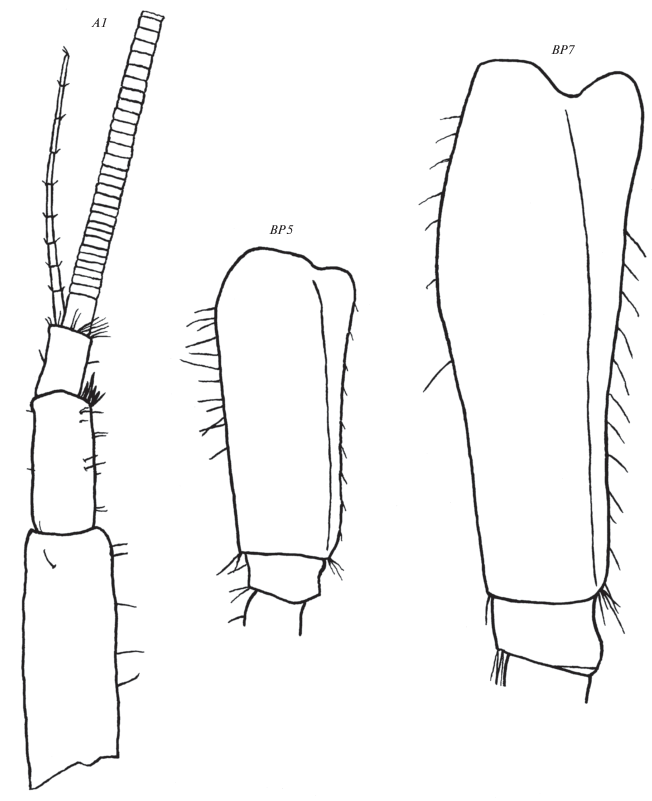
Рис. 6.
Leptostenus westschevi Tachteew sp. n., голотип самка 13.3 мм. UR1 – уропод 1, UR2 – уропод 2, UR3 – уропод 3, T – тельсон. Масштаб: UR1, UR2 – 1 мм, UR3, T – 0.5 мм.
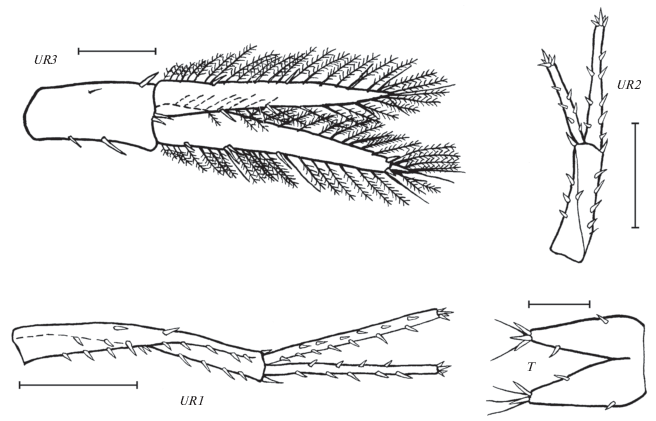
Рис. 7.
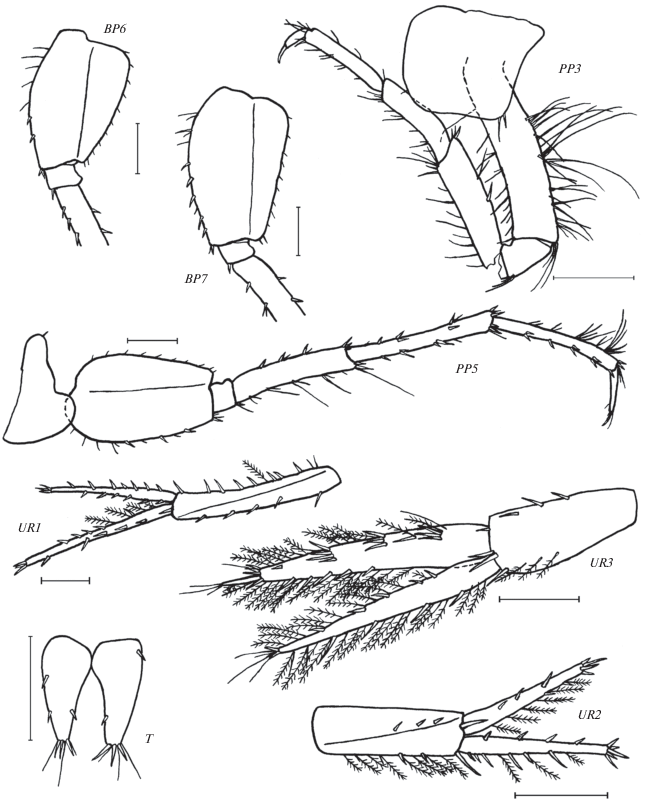
Leptostenus solontsovii Tachteew sp. n., голотип самец 13.5 мм. Жгуты антенн показаны частично. C – голова, A1 – антенна 1, A2 – антенна 2, MD – мандибула, MX1 – максилла 1, GN1 – гнатопод 1, CP + SGN2 – карпоподит и ладонь гнатопода 2. Масштаб: C – 1 мм, MX1 – 0.25 мм, остальные элементы – 0.5 мм.
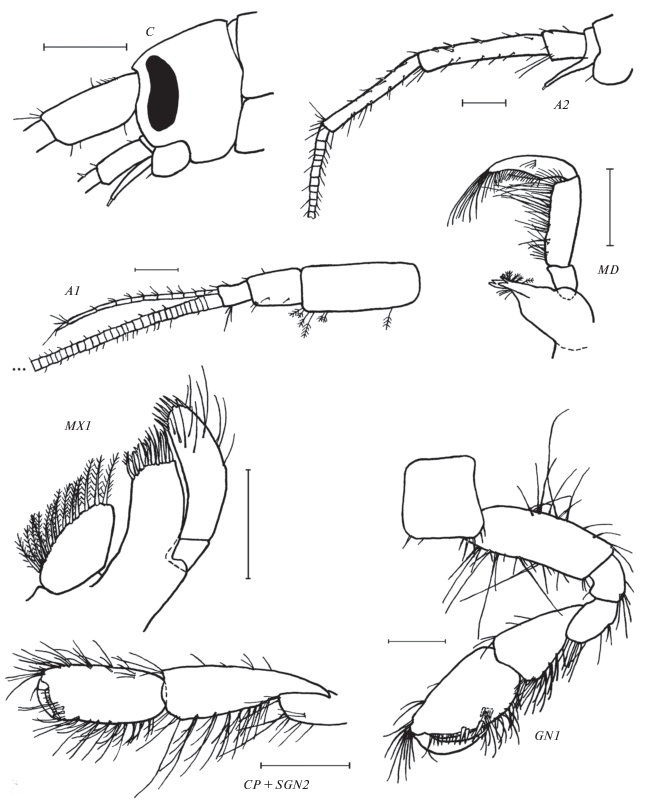
Рис. 8.
Leptostenus solontsovii Tachteew sp. n., голотип самец 13.5 мм. PP3 – переопод 3 (надломлен по мероподиту), PP5 – переопод 5, BP6 – базиподит переопода 6, BP7 – базиподит переопода 7, UR1 – уропод 1, UR2 – уропод 2, UR3 – уропод 3, T – тельсон. Масштаб 0.5 мм.
Материал. Голотип самец 13.5 мм. 24.06.2002. Оз. Байкал, банка Дриженко напротив мысов Большой Солонцовый и Саган-Морян, глубина 78–120 м. Бимтрал. НИС “Профессор Тресков” (В.В. Тахтеев). Хранится на КЗБ ИГУ (№ 21/103). Вид известен по одному экземпляру.
Описание (по голотипу). Длина тела 13.5 мм. Тело гладкое, слабое. Все сегменты метасомы с рядом недлинных, но хорошо заметных шипов по заднему краю, среди которых имеются единичные щетинки, не превышающие по длине шипы. Сегменты уросомы 1 и 2 также с шипами, но объединенными в 3 группы, одну срединную и две боковых, без щетинок. 3-й сегмент уросомы несет лишь 2 латеральных шипа.
Голова сверху слабо выпуклая, рострум едва заметный, межантеннальные лопасти короткие и высокие. Глаза темно-бурые у фиксированного экземпляра, высокие, почковидные, их задние края не изрезаны.
В антенне 1 базальный членик стержня толстый, не менее чем вдвое толще стержня антенны 2, длиннее головы, сужен к концам. 2-й членик стержня почти в 2 раза тоньше базального и вдвое короче его. 3-й членик стержня короткий, чуть длиннее половины 2-го членика, немного тоньше его, сверху слегка вогнутый. Членики стержня без шипов. Основной жгут (конец его оборван) с несколькими десятками члеников, очень коротких в проксимальной части, далее постепенно удлиняющихся. Придаточный жгутик из 11 удлиненных члеников.
В антенне 2 антеннальный конус длинный и тонкий, направлен вперед и лишь слегка вниз, немного длиннее 3-го членика стержня. Сам 3-й членик длинный, длина в 2.5 раза превышает ширину. 4-й и 5-й членики практически равной длины, несут редкие короткие щетинки, по верхней стороне также слабые шипы. Конец жгута также оторван, но жгут, несомненно, длиннее 5-го членика стержня; членики жгута короткие.
Длина верхней губы меньше ее ширины; на дистальном крае группа тонких волосков. Пальпус мандибулы с концевым члеником немного короче среднего, серповидным, заостренным, щетка занимает 2/3 его длины. В максилле 1 пальпус на 1/3 длиннее наружной пластинки, почти равен ей по ширине. На наружной пластинке 11 игл, среди которых имеются неразветвленные, двуветвистые и односторонне оперенные. Внутренняя пластинка неправильно-яйцевидная, с 14 перистыми щетинками. В максилле 2 пластинки удлиненные, внутренняя с 12 перистыми щетинками в диагональном ряду. В максиллипедах пальпус с расширенным в средней части концевым члеником, коготь на 1/3 короче него; наружная пластинка длиннее половины среднего членика пальпуса, внутренняя пластинка с 4 шипиками на вершине.
Гнатоподы самца слабые; ладонь в 1-й паре миндалевидная, почти равна по ширине карпоподиту, с двумя парами тонких граничных шипов и несколькими шипиками на внутренней стороне ближе к нижнему краю. В гнатоподах 2 ладонь узкая, почти прямоугольная, с едва заметной парой граничных шипов; коготь короткий.
Коксальные пластинки 1 почти квадратные, во 2-й паре передний угол плавно срезан, в 3-й паре длина превышает высоту, в 4-й паре задний край в нижней половине образует выдающуюся назад плавно округленную лопасть. В угловых частях коксальных пластинок сидят немногочисленные щетинки.
Переоподы 3 и 4 тонкие, несут щетинки (длинные – только по заднему краю базиподита) и единичные тонкие слабые шипы; мероподиты немного длиннее карпо- и проподитов, почти равных по длине; когти слабые, заостренные.
Переоподы 5–7 длинные и тонкие; базиподиты недлинные, умеренной ширины, задний край в 5-й паре слегка выпуклый, в 6-й и 7-й парах почти прямой, с короткими щетинками, в нижней части во всех трех парах имеется короткая лопасть, не достигающая середины исхиоподита; мероподиты немного короче карпо- и проподитов. Когти очень длинные, тонкие, почти прямые.
Эпимеральные пластинки метасомы в 1-й паре со слегка заостренным нижне-задним углом, во 2-й и 3-й – с резко заостренным, оттянутым. 2-я и 3-я пары широкие, с несколькими шипиками вдоль нижнего края.
Уроподы 1 и 2 тонкие, наружная ветвь в 1-й паре на 1/3, во 2-й паре слегка короче внутренней. 1-я пара вооружена шипами и редкими перистыми щетинками на стержне и внутренней ветви (по наружному краю); во 2-й паре перистые щетинки более многочисленные на стержне и ветвях, на обеих ветвях сидят только по внутренним краям.
В уроподах 3 стержень длинный, вооружен шипами на конце и по наружному краю, несколькими перистыми щетинками по внутреннему. Ветви равномерно суживаются дистально, наружная двучленистая, немного длиннее внутренней, 2-й членик длинный и тонкий, на конце с пучком простых щетинок. Оба края внутренней ветви и основного членика наружной с довольно густыми перистыми щетинками и с единичными простыми, а также с шипами, на внешнем крае наружной ветви образующими группы.
Тельсон удлиненный, с широкой выемкой. Ветви с вырезанными внутренними краями (из-за чего форма тельсона клешневидная), несут по 3 апикальных шипа и 3 щетинки; единичные мелкие шипики беспорядочно сидят и на поверхности ветвей.
Особенности морфологии самок и прижизненная окраска неизвестны.
Дифференциальный диагноз. От L. leptocerus вид отличается хорошо пигментированными и узкими глазами, более короткими базиподитами переоподов, крыловидный край которых образует в 5–7 парах в нижней части слабо развитую закругленную лопасть. От L. westschevi отличается большей длиной головы; более узкими глазами; более длинным 3-м члеником стержня антенны 2 (длина в 2.5 раза превышает ширину); слабым развитием лопасти крыловидного края базиподитов переоподов 5–7, не достигающей даже середины исхиоподита; наличием перистых щетинок на уроподах 1 и 2.
Распространение. Известен только из типового местонахождения. Поскольку трал, судя по очень небольшому улову, шел в придонном слое воды, лишь зацепляя глыбы на гребне банки Дриженко, сделан вывод о том, что вид ведет нектобентический образ жизни.
Этимология. Вид назван по месту нахождения (напротив м. Большой Солонцовый).
ОБСУЖДЕНИЕ
При обобщении имеющегося материала, как оригинального, так и из публикации Механиковой и Ситниковой (Mekhanikova, Sitnikova, 2014), можно констатировать, что все три вида рода Leptostenus по своему образу жизни – нектобентические, активно плавают в придонном слое воды. Вид L. westschevi входит в состав ночного миграционного комплекса в литоральной и сублиторальной зонах. Пока неизвестно, где он проводит дневное время – на дне или в придонном пространстве.
L. leptocerus характеризуется редко расставленными фасетками глаз, с неправильными их очертаниями, что говорит об его глубоководности. Это массовый обитатель глубоководных районов с гидротермальными, нефтяными и метановыми высачиваниями, в том числе с бактериальными матами. По-видимому, ими он и питается; хищничество с активным захватом жертвы маловероятно, так как у рачков слабые гнатоподы, особенно у самок, а ротовой аппарат с многочисленными щетинками, чем отчасти напоминает таковой фильтрующего типа. Морфологически L. leptocerus имеет ряд сходств с глубоководными представителями рода Poekilogammarus – P. (Bathygammarus) semenkewitschi (Sowinsky 1915) и P. (B.) unguisetosus (Sowinsky 1915). С двумя этими видами у него сходное устройство переоподов, в особенности удлиненные базиподиты 7-й пары. Сходны они также по наличию короткого покатого рострума, шипов на задних краях сегментов метасомы, по устройству гнатоподов и уроподов 1 (но щетинки на них у представителей Poekilogammarus, в отличие от L. leptoceus, простые). Также у упомянутых видов Poekilogammarus стержни антенн 2 тоньше таковых антенн 1, хотя и не настолько сильно, как у видов Leptostenus, и к тому же 3-й членик стержня антенны 1 длинный (у Leptosenus – короткий) (Tachteew, 1995). Материал для молекулярно-генетического анализа пока добыть не удалось, а указанные сходства могут оказаться конвергентными.
Большинство местонахождений видов Leptostenus привязаны к необычным по своим геологическим особенностям условиям обитания: гидротермальным, нефтяным и метановым высачиваниям, изолированным подводным поднятиям с твердыми грунтами и постоянной окислительной обстановкой; к крутым и скалистым подводным склонам.
Одним из таких своеобразных мест является банка Дриженко, расположенная в северной части Байкала в 6 км от западного берега, напротив мыса Большой Солонцовый и бухты Солонцовая, и обследованная нами в 2017 г. с помощью подводной видеотехники (Тахтеев, 2018). В историческом плане банка была официально открыта недавно – в 1984 г., во время работы гидрографической экспедиции Военно-Морского Флота СССР (Колотило, 2001), и до сих пор отсутствует на многих издаваемых картах. Вершина банки находится на глубине 56 м, за ней немного глубже (80–82 м) располагается вытянутый гребень. Как удалось установить по видеозаписи и материалу, принесенному бентосной драгой, грунт на гребне банки представлен щебнем, острыми неокатанными валунами, разбросанными глыбами. Поверхность камней со стороны воды имела бурый налет из гидроксидов железа и марганца, что свидетельствует об окислительной обстановке (Тахтеев, 2018).
На гребне банки Дриженко, кроме описываемого в данной статье L. solontsovii sp. n., обнаружены два других новых для науки вида из рода Eulimnogammarus (подрод Eurybiogammarus) (неопубликованные данные), редчайший вид Corophiomorphus gracilicornis (Bazikalova 1945) (Тахтеев, 2018), прежде известный всего из трех местонахождений с крутым подводным склоном (Базикалова, 1945).
Ранее оригинальный состав фауны амфипод нами выявлен для глубоководной части северного подводного склона острова Большой Ушканий (глубина 470–600 м), на котором, по данным минералогического анализа донных отложений и другим косвенным признакам, имеет место гидротермальная активность (Бухаров, Фиалков, 1996; Тахтеев и др., 2001) и открыты действующие и потухшие грязевые вулканы (Хлыстов и др., 2017). В частности, из амфипод идентифицирован еще один редчайший вид Lobogammarus latus Bazikalova 1945, его единственная прежняя находка сделана в первой половине XX столетия.
Таким образом, значительную часть еще не выявленного таксономического разнообразия байкальских амфипод, а также многие виды, считающиеся редкими, в будущем следует искать именно в участках озера с необычными геологическими условиями (Хлыстов и др., 2009; Zemskaya et al., 2012, и др.), многие из которых до сих пор ни разу не предстали взору исследователя.
Список литературы
Базикалова А.Я., 1941. Материалы по изучению размножения байкальских амфипод // Известия АН СССР. Отд. биол. наук. № 3. С. 407–426.
Базикалова А.Я., 1945. Амфиподы озера Байкала // Труды Байкальской лимнологической станции. Т. 11. С. 1–440.
Батранин Д.А., Тахтеев В.В., Еропова И.О., Говорухина Е.Б., 2019. Структура ночного миграционного комплекса гидробионтов в различных районах оз. Байкал летом 2017 г. и проблема изменения трофического статуса озера // Известия Иркутского госуд. ун-та. Сер. Биология. Экология. Т. 27. С. 62–86. https://doi.org/10.26516/2073-3372.2019.27.62
Бухаров А.А., Фиалков В.А., 1996. Геологическое строение дна Байкала: Взгляд из “Пайсиса”. Новосибирск: Наука. Сиб. издат. фирма РАН. 118 с.
Гебрук А.В., Кузнецов А.П., Намсараев Б.Б., Миллер Ю.М., 1993. Роль бактериальной органики в питании глубоководных донных животных в бухте Фролиха (оз. Байкал) в условиях повышенного теплового потока // Известия РАН. Серия биологическая. № 6. С. 903–908.
Камалтынов Р.М., 2001. Амфиподы (Amphipoda: Gammaroidea) // Аннотированный список фауны озера Байкал и его водосборного бассейна. Т. I: Озеро Байкал, кн. 1. Новосибирск: Наука. С. 572–831.
Колотило Л.Г., 2001. Проблемы физико-географического изучения озера Байкал: гидрографический аспект. СПб.: Русское географическое общество. 309 с.
Кузнецов А.П., Стрижов В.П., Кузин В.С., Фиалков В.А., Ястребов В.С., 1991. Новое в природе Байкала. Сообщество, основанное на бактериальном хемосинтезе // Известия АН СССР. Серия биологическая. № 5. С. 766–772.
Механикова И.В., Ситникова Т.Я., Петряшов В.В., Пензина М.М., Тимошкин О.А., 2010–2011. Каталог коллекции амфипод (включая типовые экземпляры), хранящейся в Лимнологическом институте СО РАН (г. Иркутск) // Аннотированный список фауны озера Байкал и его водосборного бассейна. Т. II: Водоемы и водотоки юга Восточной Сибири и северной Монголии, кн. 2. Новосибирск: Наука. С. 1270–1325.
Совинский В.К., 1915. Amphipoda оз. Байкал // Зоологические исследования Байкала. Т. 9. Вып. 1. Киев. 381 с. 37 табл. рис.
Тахтеев В.В., 2000. Очерки о бокоплавах озера Байкал (систематика, сравнительная экология, эволюция). Иркутск: Изд-во Иркутского университета. 355 с.
Тахтеев В.В., 2009. Глубоководные амфиподы озера Байкал, собранные с подводных обитаемых аппаратов “Пайсис” // Байкальский зоологический журнал. № 2. С. 13–16.
Тахтеев В.В., 2018. Банка Дриженко в озере Байкал // Природа. № 5. С. 62–67.
Тахтеев В.В., Бухаров А.А., Провиз В.И., Ситникова Т.Я., Галкин А.Н., 2001. Своеобразие донной фауны в необычных геологических условиях северного подводного склона Большого Ушканьего острова (оз. Байкал) // Исследования фауны водоемов Восточной Сибири: Сб. науч. тр. Иркутск: Иркутский университет. С. 3–8.
Тахтеев В.В., Карнаухов Д.Ю., Говорухина Е.Б., Мишарин А.С., 2019. Суточные вертикальные миграции гидробионтов в прибрежной зоне оз. Байкал // Биология внутренних вод. № 2. С. 50–61. https://doi.org/10.1134/S0320965219020141
Хлыстов О.М., Земская Т.И., Ситникова Т.Я., Механикова И.В., Кайгородова И.А., Горшков А.Г., Тимошкин О.А., Шубенкова О.В., Черницына С.М., Ломакина А.В., Лихошвай А.В., Сагалевич А.М., Москвин В.И., Пересыпкин В.И., Беляев Н.А., Слипенчук М.В., Тулохонов А.К., Грачев М.А., 2009. Донные битумные постройки и населяющая их биота по данным обследования озера Байкал с глубоководных обитаемых аппаратов “Мир” // Доклады Академии наук. Т. 428. № 5. С. 682–685.
Хлыстов О.М., Минами Х., Хачикубо А., Ямашита С., Де Батист М., Наудс Л., Хабуев А.В., Ченский А.Г., Губин Н.А., Воробьева С.С., 2017. Возраст грязевой брекчии грязевых вулканов Академического хребта озера Байкал // Геодинамика и тектонофизика. Т. 8. № 4. С. 923–932. https://doi.org/10.5800/GT-2017-8-4-0324
Andres H.G., Lott N., 1977. Verzeichnis der Typen aus der Sammlung Crustacea des Zoologischen Institut und Zoologischen Museums der Universität Hamburg. Amphipoda, Gammaridae // Mitt. Hamb. Zool. Mus. und Inst. 1977. Bd. 74. S. 53–64.
Barnard J.L., Barnard C.M., 1983. Freshwater Amphipoda of the world. Virginia: Mt. Vernon. 830 p.
Dybowsky B.N., 1874. Beiträge zur näheren Kenntnis der in dem Baikal-See vorkommenden niederen Krebse aus der Gruppe der Gammariden / Herausgegeben von der Russ. Entomol. Gesellsch. zu St. Petersburg. St. Petersburg: Buchdr. Von W. Besobrasoff und Comp. 190 S. 14 Taf.
Karnaukhov D.Yu., Bedulina D.S., Kaus A., Prokosov S.O., Sartoris L., Timofeyev M.A., Takhteev V.V., 2016. Behaviour of Lake Baikal amphipods as a part of the night migratory complex in the Kluevka settlement region (South-Eastern Baikal) // Crustaceana. V. 89. № 4. P. 419–430. https://doi.org/10.1163/15685403-00003530
Mekhanikova I.V., Sitnikova T.Ya., 2014. Amphipods (Amphipoda, Gammaridea) at the Gorevoy Utes oil and methane seep, Lake Baikal // Crustaceana. V. 83. № 13. P. 1500–1520. https://doi.org/10.1163/15685403-00003367
Stebbing T.R.R., 1899. Amphipoda from the Copenhagen Museum and other sources. Part II // Transactions Linn. Soc. London. 2nd Ser. Zoology. V. 7. Part 8. P. 395–432. Plates 30–35.
Stebbing T.R.R., 1906. Amphipoda I. Gammaridea // Das Tierreich. Eine Zusammenstellung und Kennzeichnung der rezenten Tierformen. 21. Lieferung. P. I–XXXIX, 1–806.
Tachteew V.V., 1995. The gammarid genus Poekilogammarus Stebbing, 1899, in Lake Baikal, Siberia (Crustacea Amphipoda Gammaridea) // Arthropoda Selecta. V. 4. № 1. P. 7–64.
Takhteev V.V., 1997. The gammarid genus Plesiogammarus Stebbing, 1899, in Lake Baikal, Siberia (Crustacea Amphipoda Gammaridea) // Arthropoda Selecta. V. 6. № 1/2. P. 31–54.
Takhteev V.V., 2019. On the current state of taxonomy of the Baikal Lake amphipods (Crustacea: Amphipoda) and the typological ways of constructing their system // Arthropoda Selecta. V. 28. № 3. P. 374–402. https://doi.org/10.15298/arthsel.28.3.03
Takhteev V.V., Berezina N.A., Sidorov D.A., 2015. Checklist of the Amphipoda (Crustacea) from continental waters of Russia, with data on alien species // Arthropoda Selecta. V. 24. № 3. P. 335–370.
Zemskaya T.I., Sitnikova T.Y., Pogodaeva T.V., Mekhanikova I.V., Naumova T.V., Shubenkova O.V., Chernitsina S.M., Kotsar O.V., Khlystov O.M., Kiyashko S.I., Kalmychkov G.V., Chernyaev E.S., 2012. Faunal communities at sites of gas- and oil-bearing fluids in Lake Baikal // Geo-Marine Letters. V. 32. № 5–6. P. 437–451. https://doi.org/10.1007/s00367-012-0297-8
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал