

Зоологический журнал, 2022, T. 101, № 11, стр. 1255-1267
Структура и динамика численности городской популяции хохлатой чернети (Aythya fuligula, Anseriformes, Anatidae) в Москве
К. В. Авилова *
Биологический факультет Московского государственного университета
имени М.В. Ломоносова
119234 Москва, Россия
* E-mail: wildlife@inbox.ru
Поступила в редакцию 22.11.2021
После доработки 12.01.2022
Принята к публикации 13.01.2022
- EDN: YHPRUP
- DOI: 10.31857/S0044513422090033
Аннотация
Исследовали структуру и численность городской популяции хохлатой чернети (Aythya fuligula) в Москве. Подсчитывали зимующих птиц в течение 36 сезонов, 1985−2021 гг., на свободных ото льда водоемах, а также самок с выводками и число утят в каждом выводке в течение 24 сезонов, 1998−2021 гг., на 150 прудах и 23 реках Москвы. Методом обратного отсчета, исходя из возраста птенцов, определяли дату начала откладки яиц самками. Размер зимующей группировки с 1985 по 2009 гг. составлял в среднем 32 (1–75) птицы. За 2010–2021 г. этот показатель вырос в среднем до 174 (111–332) птиц. Число гнездящихся самок с выводками колебалось от 11 до 45, в среднем ежегодно встречалось 27 выводков. Размер зимующей группировки коррелировал с длиной морозного периода и со среднесуточной температурой воздуха в ноябре и декабре, как и у кряквы и гоголя. Зимняя численность хохлатых чернетей коррелировала с числом взрослых самок следующим летом. Сходство цикла размножения чернети, гоголя и кряквы заключалось в упорядоченности сроков начала кладки относительно перехода среднесуточной температуры воздуха через 0°С. Чем раньше устанавливалась положительная температура воздуха, тем больше времени проходило с этого дня до начала кладки у самок. С 1998 по 2009 гг. летом росла численность взрослых чернетей, выводков и птенцов, с 2010 по 2021 гг. число выводков и утят снижалось, число взрослых птиц значимо не менялось. Соотношение числа молодых и взрослых птиц за период наблюдений сокращалось. В популяции чернети слабее, чем у кряквы, проявлялись последствия переуплотнения: удлинение периода откладки яиц и сокращение размера выводка с ростом числа самок. Динамика пространственного распределения гнездящихся хохлатых чернетей в Москве на ранних этапах определялась размещением колоний озерных чаек, с которыми они образовывали совместные поселения. Впоследствии такая зависимость ослабла.
Широкомасштабная урбанизация и техногенное преобразование больших территорий сопровождаются все более заметными проявлениями микроэволюцонных процессов в городских популяциях животных, в том числе птиц (Морозов и др., 2016; Схилтхёйзен, 2021; Baratti et al., 2015; van Dongen et al., 2015; Johnson, Munshi-South, 2017; Minias et al., 2017). Обособление таких популяций начинается с изменения их пространственных и временны́х характеристик. Многолетний мониторинг городской популяции кряквы (Anas platyrhynchos) и гоголя (Bucephala clangula) в Москве выявил основные направления трансформации жизненного цикла, компенсаторные и регуляторные явления, ведущие к более рациональному использованию ресурсов, а также к определенным трендам динамики численности популяции (Авилова, 2016, 2018, 2019). В то же время Москва служит ареной освоения жизненного пространства еще для ряда видов водоплавающих птиц. Это может быть вызвано смещением мест зимовки к северо-востоку (Švažas et al., 2001; Lehikoinen et al., 2013) и сокращением миграционных путей (Viksne et al., 2010; Gunnarson et al., 2012; Guillemain et al., 2013), мероприятиями по обогащению городской фауны (Благосклонов, 1967; Кудрявцев, 1967; Остапенко и др., 1989), активным освоением птицами урбанизированной среды обитания (Фридман, Еремкин, 2009; Скилтхёйзен, 2021). Один из гнездящихся в Москве видов – хохлатая чернеть (Aythya fuligula), широко распространенная на территории Палеарктики. Ее ареал занимает всю лесную зону умеренного пояса и прилежащие участки соседних зон. Это самая многочисленная нырковая утка в России. В европейской части России обитает 1.8 млн особей, или 21% от общего числа. Основными местообитаниями служат крупные мезотрофные озера, а также долины рек (Кривенко, Виноградов, 2008). Инвентаризация орнитофауны 16 крупных городов Европы от Лиссабона до Санкт-Петербурга (Kelcey, Rheinwald, 2005) показала, что хохлатая чернеть гнездится в девяти городах, включая Москву и Санкт-Петербург. Мы поставили целью с помощью анализа данных многолетних исследований (1985–2021) оценить структуру популяции и особенности динамики численности чернети в Москве, сравнить их с таковыми у других видов и выявить факторы, определяющие особенности освоения городской среды обитания.
МАТЕРИАЛЫ И МЕТОДЫ
В качестве показателя зимней численности использовали число чернетей обоего пола, зимующих в Москве, а в качестве показателя летней численности – число самок с утятами и взрослых птиц без выводков на городских акваториях. Подсчет птиц зимой проводили с 1985 по 2021 гг. в третьей декаде января на всех незамерзающих водных объектах Москвы. Летний подсчет проводили в 1998–2021 гг. на большинстве освоенных водоплавающими птицами акваторий в границах МКАД с конца июня до конца июля. Это более 150 замкнутых водоемов (озер, прудов, отстойников), а также 23 участка рек и каналов Москвы. В качестве показателей плотности населения использовали количество гнездящихся самок, успешно пришедших с утятами на водоем (Михельсон и др., 1977, 1986). Подсчитывали число утят в каждом выводке и определяли их возраст по шкале, разработанной для уток и основанной на показателях развития оперения (Gollop, Marshall, 1954; Pirkola, Högmander, 1974). Время начала первых кладок − традиционная точка отсчета возникновения благоприятной для гнездования ситуации (Артемьев, 2008; Соколов, 2010). Сроки вылупления птенцов и дату откладки первого яйца вычисляли методом обратного отсчета (Онно, 1974; Oja, Poysa, 2007). Исходили из возраста птенцов, среднего срока насиживания (25 дней) и среднего срока откладки яиц (8 дней) при среднем размере кладки в 8 яиц (Исаков, Птушенко, 1952; Птушенко, Иноземцев, 1968; Мальчевский, Пукинский, 1983; Пыжьянов, Березовская, 2010; Hill, 1984).
При анализе влияния климатических факторов на основные этапы жизненного цикла использовали данные по ходу среднесуточных температур воздуха из базы главной агрометеостанции Москвы (ВВЦ) и Метеорологической обсерватории МГУ. Длину морозного периода вычисляли как число дней между устойчивым переходом температур через 0°С весной (Садоков и др., 2012) и осенью. Значения индекса североатлантического колебания САК (Нестеров, 2013; Холопцев и др., 2015) взяты из базы данных Климатического прогностического центра США (CPC/NCEP/ NOAA). Для удобства анализа связи дат с другими показателями в расчетах использовали число дней, прошедших с фиксированной даты (01.02) до даты становления положительной среднесуточной температуры воздуха, а также с 1.04 до даты откладки первого яйца. Статистическую значимость связи различных показателей оценивали методами непараметрической статистики, факторного анализа и множественной регрессии в пакете программ Statistica 10, подбор наилучшей модели для многолетних учетных данных проводили в программе Curveexpert 1.
РЕЗУЛЬТАТЫ
Хотя в Москве ежегодно образуется более 30 незамерзающих акваторий, пригодных для зимовки, несколько группировок зимующих чернетей встречаются только на Москве-реке вблизи центра и ниже по течению, несколько летных птиц зимует в зоопарке. Зимующие чернети единично и нерегулярно появлялись на городской акватории Москвы-реки с конца 1980-х гг. С 1991 года число зимующих птиц превысило 20, с 1997 по 2009 гг. зимовало уже в среднем 45–47 птиц, а с 2010 г. – более ста. В 2014 г. численность зимующих чернетей превышала 300 птиц (рис. 1). Общая численность нарастала (rs = 0.90, p < 0.05, n = 36), но с 2014 г. она стала постепенно снижаться (rs = = –0.71, p < 0.05, n = 8). От четырех до 19 чернетей зимует также на Москве-реке ниже границы города, рост их числа не отмечен (Зубакин и др., 2016–2021).
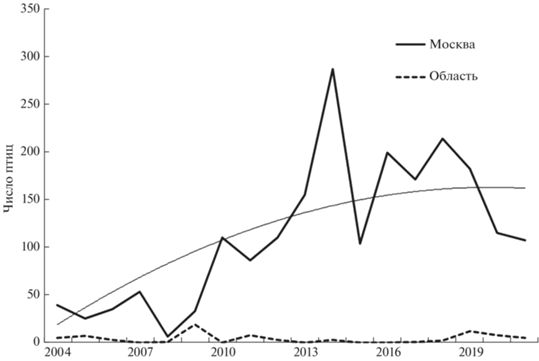
С 2004 г. кривая динамики численности приобрела форму купола, которую можно описать степенной зависимостью (рис. 1)
r – коэффициент корреляции ожидаемого согласно формуле и наблюдаемого числа птиц.
Скопления зимующих птиц формируются в ноябре, когда замерзают стоячие водоемы, и достигают максимальной численности в декабре. Число зимующих птиц остается примерно на одном уровне до марта, а затем чернети постепенно отлетают с Москвы-реки к местам гнездования (Зубакин и др., 2016–2021).
Продолжительность морозного периода в Москве, т.е. число дней с момента перехода среднесуточной температуры через 0°С осенью до перехода через 0°С весной, за 37 лет значимо сократилась. В первой половине этого периода продолжительность в среднем составила 135 дней, во второй половине – 105 дней (Z = 4.1, p = 0.0001, n = 37). В тот же период росла средняя температура воздуха ноября и декабря (rs = 0.53 и rs = 0.44, p < 0.05, n = 37).
Численность зимующих в Москве чернетей коррелирует с длиной морозного периода (rs = –0.65, p < 0.001, п = 36), а также со среднесуточной температурой ноября (rs = 0.51, p < 0.05, п = 36) и декабря (rs = 0.41, p < 0.05, п = 36), т.е. в короткие зимы и в более теплые периоды конца осени и начала зимы в городе зимует больше птиц. Связь с температурами января не выявлена. Также не проявилась связь с характеристикой крупномасштабных циркуляций атмосферы Северного полушария в холодное время года − Индексом Северо-Атлантического Колебания, в ноябре, декабре и январе.
Весной после освобождения ото льда замкнутых водоемов на прудах Москвы появляются перезимовавшие чернети, к которым на время присоединяются стаи пролетных. К июню численность снижается, но группы и пары птиц без выводков встречаются на прудах в течение всего гнездового сезона. Число взрослых чернетей без выводков на водоемах значимо выше в годы с ранней весной (rs = –0.45, p < 0.05, п = 24).
Зимняя численность хохлатых чернетей статистически связана с числом гнездящихся следующим летом самок с 1998 по 2021 гг. (rs = 0.45, p < < 0.05, п = 24). В то же время, чем выше зимняя численность, тем раньше самки приступают к гнездованию (rs = –0.42, p < 0.05, п = 24).
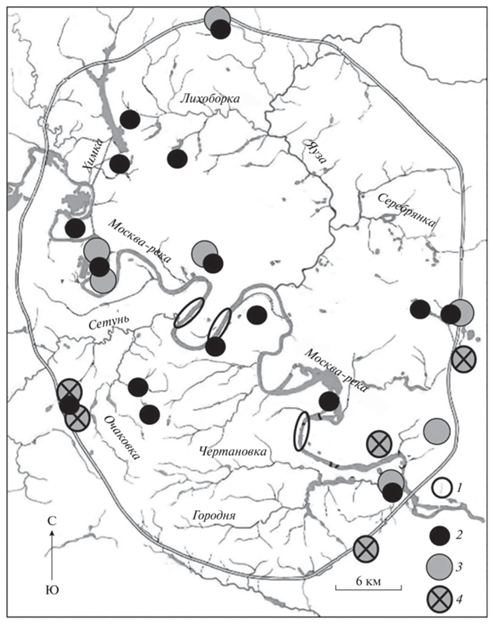
Гнездовая популяция в период наблюдений была сосредоточена на нескольких водно-болотных комплексах запада, северо-запада и, в меньшей степени, востока Москвы. В центре сформировались лётная группа Московского зоопарка и поселение на острове Большого Голицынского пруда ЦПКиО им. Горького. Хохлатые чернети гнездились и на урбанизированных акваториях за МКАД: Долгих прудах, у пл. Марк Савеловской ж.д., на Яузских болотах, в Новой Москве и др. Места гнездования часто были сконцентрированы возле колоний чайковых птиц, чаще всего – озерных чаек (Larus ridibundus), реже – сизых чаек (L. canus), речных крачек (Sterna hirundo) (рис. 2).
Рис. 2.
Летнее и зимнее размещение хохлатых чернетей и локализация колоний чаек в Москве в 1998–2021 г.: 1 – места зимовки чернетей, 2 – места гнездования чернетей, 3 – существующие колонии озерных чаек, 4 – исчезнувшие колонии озерных чаек.
Чернети гнездятся в траве и в зарослях кустарников по берегам водоемов, особенно на островах. В зоопарке гнездятся в домиках для уток (Авилова и др., 2007). В настоящее время размножению хохлатой чернети и других уток в зоопарке препятствует хищничество серебристых чаек. В 2017 и 2018 гг. было установлено гнездование чернетей в пределах Садового кольца на Патриаршем пруду, причем в 2017 г. в одном домике гнездились сразу две самки, впоследствии водившие на пруду сдвоенный выводок. С 1998 по 2009 гг. летом росла численность взрослых птиц (rs = 0.75, p < 0.01, п = 12), выводков (rs = 0.67, p < 0.05, п = 12) и птенцов (rs = 0.70, p < 0.05, п = 12), затем рост прекратился (рис. 3). Число взрослых чернетей без выводков, в основном самок, на городских водоемах колебалось в широких пределах от единиц до скоплений из более 150 птиц, число выводков достигало 45, а число птенцов – почти трехсот. После 2009 г. численность выводков и суммарное число утят снижалось (rs = –0.75, rs = = 0.82, p < 0.05, п = 12), но число взрослых птиц на водоемах значимо не изменялось. Это свидетельствует об ухудшении условий гнездования при сохранении общего размера группировки.
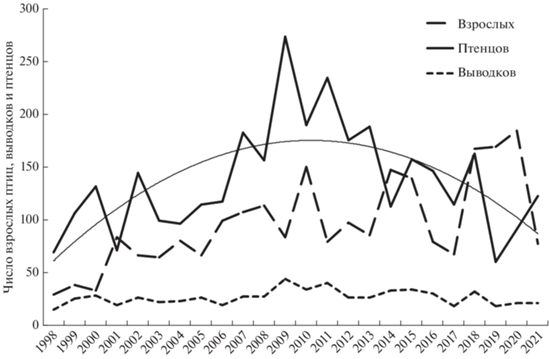
Динамика численности выводков за 24 сезона описывается полиномом 3-й степени:
По нашим подсчетам за 23 сезона самая ранняя дата откладки первого яйца хохлатой чернетью в Москве пришлась на период с 9 по 18 апреля. Это было как в 2014, так и в 2020 гг., то есть в годы с ранней весной. Самая поздняя дата пришлась на период с 7 по 12 августа 2020 г. Интервал между самыми ранними и самыми поздними кладками составлял от 10 в 2010 г. до 116 дней в 2020 г. Этот интервал сильно колебался (С = = 26.5), но возрастал за период 1998−2021 гг. (rs = = 0.53, p < 0.01, п = 24).
Несмотря на ключевое значение для гусеобразных устойчивого весеннего потепления, к которому приурочено освобождение водоемов ото льда, дата откладки первого яйца не коррелировала с датой перехода средних температур воздуха через 0°С. Влияние сроков начала весны проявлялось в том, что чем позже температура воздуха устойчиво переходила через 0°С и позже оттаивали водоемы, тем короче был промежуток со дня этого перехода до откладки первого яйца чернетью (rs = –0.86, p < 0.001, n = 24).
За весь период исследований переход средних температур через 0°С состоялся между 2-й декадой февраля и 2-й декадой марта в ранние весны (12 лет) либо между 3-й декадой марта и 1-й декадой апреля в поздние (12 лет). В раннюю весну с перехода через 0°С до откладки 1-го яйца проходило 54 ± 4.5 дней, в позднюю – 30 ± 2.5 дня, эти показатели (рис. 4, 5) значимо различались (Z = = 3.67, p < 0.001, n = 24).
Рис. 4.
Даты откладки первого яйца у хохлатой чернети относительно дат перехода среднесуточной температуры воздуха через 0°С: 1 – декада, когда произошел переход среднесуточной температуры воздуха через 0°С; 2 – декада, когда было отложено первое яйцо.
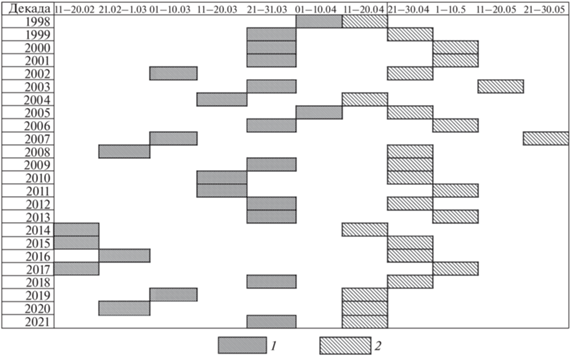
Рис. 5.
Сроки перехода средней суточной температуры воздуха через 0°С (число дней с 1 февраля) и начала кладки у чернети в раннюю (сверху) и позднюю весну.
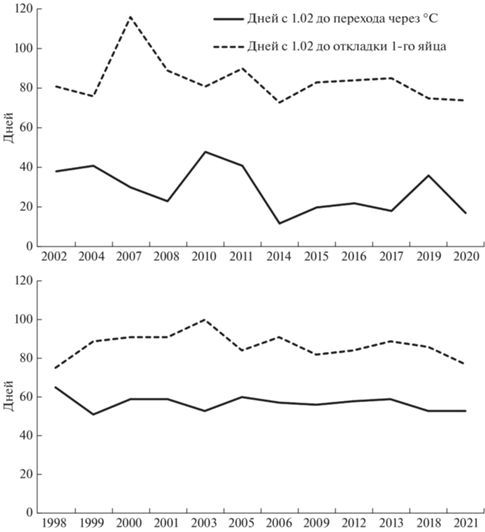
Средние суточные температуры в день начала кладки в годы с ранней и поздней весной значимо не различались. Частота наступления ранних весен в последние годы возрастает (rs = –0.51, p < < 0.05, п = 24). Срок перехода средней суточной температуры через 0°С в течение изучаемого периода достоверно не зависит от условий предыдущих сезонов, так как автокорреляция с аналогичными сроками один, два или три года назад не проявляется.
Для выявления факторов, значимо влияющих на сроки начала гнездования, мы применили метод главных компонент к группе показателей: зимней и летней численности, дате установления положительных температур воздуха, числом дней с этой даты до начала гнездования. Сроки размножения определяются группой факторов, среди которых год, численность размножающихся самок, а также скрытая переменная с биполярной факторной нагрузкой, т.е. с наличием положительных и отрицательных показателей в одном факторе. Эта переменная объединяет дату перехода средних температур через 0°С и отрезок времени между переходом температур через 0°С и датой откладки первого яйца. Таким образом, последняя связана с ранним или поздним наступлением весны в текущем сезоне и численностью гнездящихся самок. Метод множественной корреляции показал, что почти 75% дисперсии промежутка времени от перехода через 0°С до начала гнездования определяется сроком перехода средних температур через 0°С (F = 14.3, R2 = 74.6, p < < 0.0001), т.е. ранним или поздним наступлением весны.
В ранние весны разброс сроков начала гнездования, т.е. стандартное отклонение даты начала кладки, больше (С = 11.5), чем в поздние (С = 8.5). Тем не менее чернети даже при позднем наступлении потепления начинали кладку не сразу после перехода среднесуточных температур через 0°С, а со сдвигом во времени (рис. 4). Диапазон сроков перехода средних температур через 0°С (С = 16.7) шире, чем диапазон начала первых в сезоне кладок (С = 10.6). Сроки размножения хохлатой чернети, таким образом, упорядочены относительно более изменчивых сроков наступления устойчиво теплой погоды.
Вылупление птенцов хохлатой чернети в Москве происходило в основном в июле. Появление выводков в мае за 24 года отмечено 9 раз (1.32%), а в конце июля–августе – 5 раз (0.75%). В сроках появления выводков выявляются два пика, первый из которых приходится на июнь, а второй, как правило, более выраженный, – на июль (рис. 6). Растянутость сезона гнездования растет с годами (rs = 0.53, p < 0.05, n = 24) и коррелирует с числом гнездящихся самок (rs = 0.40, p < < 0.05, n = 24). Значимых сдвигов в сроках вылупления за 24 года не выявлено, хотя происходило некоторое сокращение доли июльских выводков (rs = –0.49, p < 0.05, n = 24).
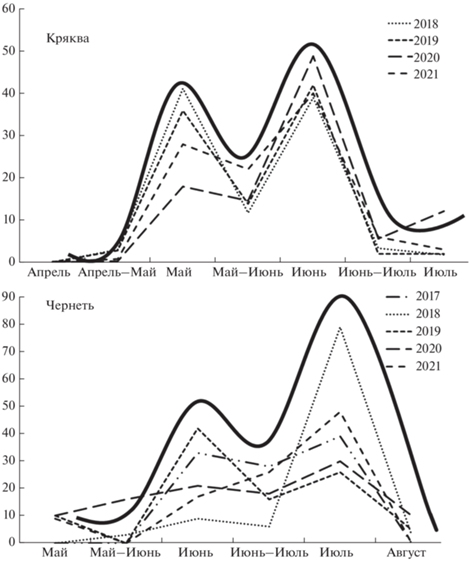
Изменение размера выводка за период наблюдений, а также по мере роста и развития птенцов не выявлено. Не выявлена и его связь с числом гнездящихся самок. Число утят в выводке незначимо увеличивалось в годы с ранней весной.
В отличие от размера выводка соотношение числа молодых и взрослых птиц за период наблюдений сокращалось (rs = –0.44, p < 0.05, n = 24) независимо от сроков наступления весны. Это соотношение числа молодых и взрослых птиц уменьшалось в годы высокой численности самок и возрастало в годы с низкой (rs = –0.40, p < 0.05, n = 24). При высокой плотности населения водоплавающих птиц, например на прудах ЦПКиО, отмечали смешанные выводки чернети и кряквы, обычно сопровождаемые самками крякв.
В Москве летом хохлатые чернети вместе с кряквами и гоголями охотно берут подкормку у горожан. При этом между самками чернетей и крякв регулярно возникают конфликты, победителями из которых выходят, как правило, кряквы.
ОБСУЖДЕНИЕ
В начале и середине прошлого века хохлатая чернеть была обычной пролетной и спорадично гнездящейся птицей Московского региона (Птушенко, Иноземцев, 1968). Во второй его половине она стала довольно обычной в средней полосе на гнездовании благодаря развитию, прежде всего, сети рыборазводных прудов (Мищенко, 1983) и других искусственных водоемов (карьеров, очистных сооружений). Формирование системы рыбхозов в европейской части России произошло в основном в 1950–1960-е годы и стало одной из предпосылок начала гнездования уток на техногенных водоемах, в том числе – в пригородах. На рыборазводных прудах в 1980-е годы благодаря оптимальному сочетанию условий – мелководности, изобилию кормов, – гнездилось более половины подмосковной популяции хохлатой чернети (Суханова, 2005; Суханова, Мищенко, 2008). Не в последнюю очередь аттрактантом для нырковых уток – хохлатой чернети и красноголового нырка (Aythya ferina) − стали формирующиеся на рыборазводных прудах колонии чайковых птиц, как правило – озерных чаек. Под их защитой от хищников эти утки охотно гнездятся (Формозов, 1981; Глушенков, 1997; Пыжьянов, Березовская, 2010; Liordos, Lauder, 2015; Väänänen, 2000; Väänänen et al., 2016).
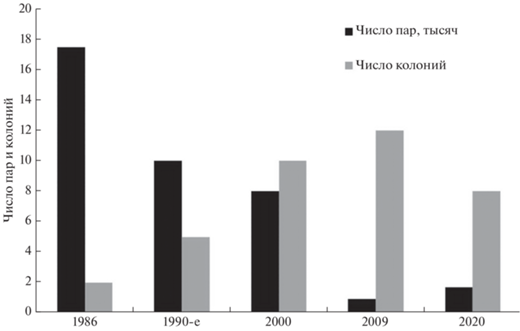
Второй предпосылкой образования городской популяции стали искусственные водоемы и иловые площадки на очистных сооружениях юго-востока Москвы (Авилова, Чичев, 1997). В 1980-х годах там сформировалось крупное поселение озерных чаек, достигавшее численности в 15 тысяч пар (Попова-Бондаренко, 1986; Еремкин, 1997; Зубакин, 2018). На иловых площадках Люблинской станции аэрации рядом с озерными чайками размножалось от 20 до 50 пар хохлатых чернетей (Еремкин, 1997). Они стали активно расселяться по городу в конце 1990-х после начала реализации решения о ликвидации очистной станции и строительстве на ее месте жилого района Марьино (рис. 7). Динамика пространственного распределения определялась на ранних этапах распределением колоний озерных чаек. Одним из следствий уничтожения колоний на старом месте стало образование рассеянных по городу новых, более мелких группировок, рядом с которыми и появлялись выводки чернетей: в Мневниковской и Братеевской поймах, на Большом Крылатском карьере, на р. Пустая Наверашка, на Косинских озерах и в других местах (рис. 1). Затем чернети стали занимать и другие водоемы, где чайки отсутствовали (Кузиков, 2020). Наличие колоний чаек оказалось для чернетей не столь необходимым условием, как для красноголовых нырков, более тесно привязанных в Москве к поселениям чаек (Авилова и др., 2018).
Рис. 7.
Примерная численность и структура населения озерных чаек в Москве за период 1986–2020 гг. (с привлечением данных В.А. Зубакина, Г.С. Еремкина и И.М. Панфиловой).
В динамике численности и сроках основных явлений годового цикла у трех исследованных видов (хохлатая чернеть, гоголь и кряква) выявляются как сходные особенности, так и различия.
Средняя зимняя и летняя численность хохлатых чернетей (111–112 птиц) на городских водоемах не различаются. У кряквы средняя зимняя превосходит среднюю летнюю в 5.4 раза, а у гоголя – в 12.5 раз. Группировки чернетей, в отличие от двух других видов, компактные и размещены на реке локально скоплениями до 80 птиц, в основном недалеко от центра. За пределами города зимует 4–19 чернетей, в то время как у гоголя суммарно в среднем 500–700 птиц, у кряквы – 1760 птиц. Тем не менее, численность зимующих в границах города чернетей так же, как гоголей и крякв, увеличивается с сокращением морозного периода и с ростом среднесуточных позднеосенних температур. Влияния североатлантического колебания не выявлено, хотя численность гнездящихся уток зависит от этого явления на территории Западной Сибири (Михантьев, Селиванова, 2018).
Увеличение числа гнездящихся самок в годы с ранним наступлением весны у кряквы достигало 40%, а у гоголя было статистически незначимо. У чернети этот показатель почти значимо возрастал на 2–3%. Рост говорит об улучшении условий размножения при раннем наступлении устойчивого потепления, но он труднее улавливается при небольших объемах выборки. Об этом свидетельствуют и значимо более высокая численность самок на водоемах в годы с ранней весной, и некоторое увеличение размера выводка. Таким образом, упорядоченность сроков размножения относительно срока наступления устойчивой теплой погоды и увеличение продуктивности при раннем наступлении весны отчетливо проявляются у чернети, кряквы и гоголя (Авилова, 2016, 2019). Очевидно, это характеризует общую адаптацию к географическим и климатическим характеристикам местности (Ларионов, 1953, 1956, 1959) и связано не только с ее широтой, но и с ходом температур конкретного сезона в месте гнездования. Упорядоченность становится особенно заметной в ходе изменения климата и увеличения частоты наступления ранних весен. Несмотря на сокращение морозного периода, городские чернети ни разу за 24 сезона не приступали к кладке до перехода температуры через 0°С или одновременно, что связано, по-видимому, с типом гнездования чернети, которая, в отличие от кряквы и особенно гоголя, реже занимает дупла, домики и другие укрытия. В Московской обл. хохлатая чернеть, как и кряква, приступает к кладке позднее, чем в Москве, с 12 мая по 1 июня (Птушенко, Иноземцев, 1968). Общая продолжительность сезона гнездования у чернети, как и у кряквы, удлиняется с ростом числа гнездящихся самок (rs = 0.40, p < 0.07, n = 24), чего не отмечено в популяции гоголя. Более раннему гнездованию чернети, как и кряквы, способствует увеличение числа зимующих птиц (rs = 0.43, p < 0.05, n = 24).
Сезон откладки яиц у хохлатой чернети короче, чем у кряквы (Z = –5.21, p < 0.01, n = 24), но длиннее, чем у гоголя (Z = 2.1, p < 0.05, n = 24). В Великобритании сезон по продолжительности не отличался от сезона в Москве (Hill, 1984), а на Байкале в стабильных условиях был значительно короче и без ярко выраженных пиков (Пыжьянов, Березовская, 2013). Первый условный “пик” авторы относят к первичным кладкам, а второй “пик” – к компенсаторным и поздним первичным. В Москве, где условия далеки от стабильных, пики размножения выражены в разные годы по-разному, часто с преобладанием второго пика в сезоне. Очевидно, это связано с частой потерей гнезд и кладок. Вылупление утят у кряквы происходит в основном в мае и июне, у чернети – в июне и в июле (рис. 6). У гоголя вылупление птенцов происходит преимущественно в мае. Это усиливает конкуренцию выводков на плотно заселенных городских водоемах, так как чернети приводят утят на пруды, уже занятые выводками других видов. Размер выводка у чернети, как и у гоголя, не изменяется в зависимости от числа самок, что ярко проявляется у кряквы, но соотношение молодых и взрослых птиц уменьшается как в течение всего периода наблюдений, так и в годы более высокой летней численностью самок.
Выявленные особенности позволяют говорить о возможной оседлости городской популяции хохлатой чернети: сходство размеров зимней и летней группировки, локальное размещение зимних скоплений, отличное от других водоплавающих. Определенное значение в этом имело создание летной группы в Московском зоопарке. Для подтверждения оседлости необходимо располагать данными мечения, которых пока нет. Часть особей размножается в зоопарке и пополняет городскую группировку (Остапенко, Виноградов, 1984; Остапенко и др., 1989; Авилова и др., 2007). Летом 2021 г. в зоопарке гнездилось минимум две самки, осенью и весной собирались стаи по 20 птиц (И. Сметанин, личное сообщение).
Более раннее гнездование при более высокой зимней численности, увеличение продолжительности сезона размножения и сокращение соотношения молодые/взрослые с ростом числа самок могут быть интерпретированы как адаптации, компенсирующие потери кладок и выводков, которые неизбежны в городе с постоянным фактором беспокойства. Такие адаптации отчетливо выражены в городской популяции кряквы (Авилова, 2016). Это признаки прогрессирующей интеграции группировки в городскую среду обитания.
В 2000–2010 гг. выделилось около 30 мест регулярного гнездования чернетей в Москве (Авилова, Самойлов, 2011). В дальнейшем большинство их сохранилось (Голицынский пруд в ЦПКиО, пруды в Покровском-Стрешневе, Большой Крылатский карьер и др.), хотя используются эти участки не ежегодно. Чернеть нерегулярно гнездится также на Большом Садовом пруду МСХА, на Москве-реке выше и ниже Строгинского моста, на прудах р. Наверашка, на оз. Бездонное в Серебряном бору, на Андреевских прудах Воробьевых гор. К 2009 г. численность восстановилась практически до уровня 1980–1990-х годов. Главной причиной постепенного последующего ее снижения стало, по нашему мнению, увеличение антропогенной нагрузки в связи с масштабным благоустройством природных территорий и повсеместная экспансия кряквы (Авилова, 2016, 2018), с которой чернети трудно конкурировать, особенно при сокращении площади местообитаний в результате благоустройства. Обилие лодок и катамаранов на прудах приводит к постоянному беспокойству гнездящихся водоплавающих птиц, к перемешиванию и гибели выводков.
Филопатрия, которая у самок гусеобразных выражена сильнее, чем у самцов (Savard, 1985), хорошо проявляется у репродуктивно успешных птиц в стабильных местообитаниях (Соколов, 1991). У самок хохлатой чернети в природе отмечен рост числа возвратов к месту гнездования после удачного сезона размножения (Михельсон и др., 1986; Blums et al., 2002). Сокращение числа гнездящихся самок после неудачных попыток гнездования ведет к ослаблению филопатрии и росту дисперсии, к нарушению устойчивости популяции. Изменение численности и размещения колоний озерных чаек в Москве (Зубакин, 2018) также играет свою роль. Высокая уязвимость гнездящихся птиц и низкий успех размножения даже при удачном гнездовании не дают полной уверенности в дальнейшем благополучии популяции.
Хохлатая чернеть внесена в Красную книгу Москвы (Авилова, Самойлов, 2011) как уязвимый вид и в Приложение 1 Красной книги Московской области (2018) как популяция, нуждающаяся в постоянном контроле и наблюдении. Следует принять во внимание рекомендации международной группы ученых по сохранению условий обитания колоний озерных чаек в ходе реабилитации существующих и создания новых водоемов (Kauppinen, 2012; Väänänen et al., 2016).
ЗАКЛЮЧЕНИЕ
Главными источниками формирования городской популяции хохлатой чернети в Московском регионе стали искусственные местообитания – пруды рыборазводных хозяйств и полигоны депонирования осадка сточных вод.
Динамика пространственного распределения гнездящихся хохлатых чернетей в мегаполисе определялась на ранних этапах распределением колоний озерных чаек, с которыми они образовывали совместные поселения, впоследствии такая зависимость ослабла.
В гнездовых группировках чернети, как и двух других видов гусеобразных Москвы, выявляется упорядоченность сроков начала гнездования относительно сроков весеннего потепления, вслед за которым следует вскрытие водоемов, но не проявляются отрицательные последствия зимних погодных аномалий, которые стали ключевыми по отношению к городской группировке гоголя.
В немногочисленной городской популяции хохлатой чернети не столь выражены те компенсаторные и регуляторные явления, которые обнаружены в более крупной и стабильной популяции кряквы: удлинение периода размножения, более раннее начало кладки, уменьшение успеха размножения с ростом числа самок. Тем не менее, их слабые проявления свидетельствуют о постепенной интеграции хохлатой чернети в городскую среду обитания при сохранении подходящих биотопов.
Список литературы
Авилова К.В., 2016. Жизненный цикл и динамика численности городской популяции кряквы (Anas platyrhynchos) в Москве // Зоологический журнал. Т. 95. № 12. С. 1427–1440.
Авилова К.В., 2018. Структура и долговременная динамика зимнего населения кряквы (Anas platyrhynchos, Anseriformes, Anatidae) в Москве // Зоологический журнал. Т. 97. № 3. С. 309–320.
Авилова К.В., 2019. Структура и динамика численности городской популяции гоголя (Bucephala clangula, Anseriformes, Anatidae) в Москве // Зоологический журнал. Т. 98. № 3. С. 1–13.
Авилова К.В., Поповкина А.Б., Сметанин И.С., 2007. Роль Московского зоопарка в поддержании городской группировки гусеобразных птиц // Орнитологические исследования в зоопарках и питомниках. Вып. 2. М.: Московский зоопарк. С. 21–30.
Авилова К.В., Зубакин В.А., Еремкин Г.С., Лыков Е.Л., Панфилова И.М., 2018. Пути освоения водоплавающими птицами городской среды обитания // Актуальные проблемы охраны птиц. Материалы Всероссийской науч.-практ. конференции, посвященной 25-летию Союза охраны птиц России (Москва, 10–11 февраля 2018 г.). Отв. ред. А.В. Салтыков. Москва. Махачкала: АЛЕФ (ИП Овчинников). С. 170–176.
Авилова К.В., Самойлов Б.Л., 2011. Хохлатая чернеть // Красная Книга города Москвы. 2-е издание. Отв. ред. Б.Л. Самойлов, Г.В. Морозова. М.: Изд-во Департамента природопользования и охраны окружающей среды г. Москвы. С. 126–128.
Авилова К.В., Чичёв А.В., 1997. Сукцессионные смены растительных сообществ и группировок птиц полигона депонирования осадка сточных вод Москвы // Бюллетень МОИП, отд. биол. Т. 102. Вып. 3. С. 34–38.
Артемьев А.В., 2008. Популяционная экология мухоловки-пеструшки в северной зоне ареала. М.: Наука. 268 с.
Благосклонов К.Н., 1967. Птицы города Москвы // Животное население Москвы и Подмосковья, его изучение, охрана и направленное преобразование. М.: Изд-во МГУ. С. 79–82.
Глушенков О.В., 1997. Формирование орнитоценозов околоводных и водоплавающих птиц в пригородных зонах // Птицы техногенных водоемов Центральной России. М.: Изд-во МГУ. С. 86–99.
Еремкин Г.С., 1997. Очерк орнитологической фауны Люблинских полей фильтрации // Птицы техногенных водоемов Центральной России. М.: Изд-во МГУ. С. 7–24.
Зубакин В.А., 2018. Урбанизированная популяция озерных чаек города Москвы и ближнего Подмосковья: история и закономерности формирования пространственной структуры // Орнитология: история, традиции, проблемы и перспективы. Материалы Всероссийской конференции, посвященной 120-летию со дня рождения Г.П. Дементьева. М.: Товарищество научных изданий КМК. С. 169–175.
Зубакин В.А., Ерёмкин Г.С., Бащинская С.В., Бондарева Н.А., Варламов А.Е. и др., 2016. Результаты учета зимующих водоплавающих и околоводных птиц на реках Москве и Оке в столице и Подмосковье в сезон 2015/2016 гг. // Московка. Новости программы Птицы Москвы и Подмосковья. № 23. С. 9–27.
Зубакин В.А., Ерёмкин Г.С., Бащинская С.В., Бондарева Н.А., Воронов Д.А. и др., 2017. Итоги учета зимующих водоплавающих и околоводных птиц на реках Москве и Оке в столице и Подмосковье в сезон 2016/2017 гг. // Московка. Новости программы Птицы Москвы и Подмосковья. № 26. С. 13–32.
Зубакин В.А., Ерёмкин Г.С., Бащинская С.В., Бондарева Н.А., Варламов А.Е. и др., 2018. Водоплавающие и околоводные птицы, зимовавшие на реках Москве и Оке в столице и Подмосковье в сезон 2017/2018 гг. // Московка. Новости программы Птицы Москвы и Подмосковья. № 27. С. 10–36.
Зубакин В.А., Ерёмкин Г.С., Бащинская С.В., Бондарева Н.А., Варламов А.Е. и др., 2019. Итоги учета зимующих водоплавающих и околоводных птиц на реках Москве и Оке в столице и Подмосковье в сезон 2018/2019 гг. // Московка. Новости программы Птицы Москвы и Подмосковья. № 30. С. 12–28.
Зубакин В.А., Ерёмкин Г.С., Бащинская С.В., Бондарева Н.А., Воронов Д.А. и др., 2020. Численность зимующих водоплавающих и околоводных птиц на реках Москве и Оке в Московском регионе в сезон 2019/2020 гг. // Московка. Новости программы Птицы Москвы и Подмосковья. № 31. С. 13–36.
Зубакин В.А., Шведко М.А., Ерёмкин Г.С., Антоновский Т.Р., Бащинская С.В. и др., 2021. Результаты учета водоплавающих и околоводных птиц на реках Москве и Оке в столице и Подмосковье в сезон 2020/2021 гг. // Московка. Новости программы Птицы Москвы и Подмосковья. № 33. С. 9–33.
Исаков Ю.А., Птушенко Е.С., 1952. Хохлатая чернеть // Птицы Советского Союза. Т. 4. М.: Советская наука. С. 497–506.
Красная книга Московской области, 2018. Отв. ред. Т.И. Варлыгина, В.А. Зубакин, Н.Б. Никитский, А.В. Свиридов. Изд. 3-е, перераб. и доп. Московская обл.: Верховье. 810 с.
Кривенко В.Г., Виноградов В.Г., 2008. Птицы водной среды и ритмы климата Северной Евразии. М.: Наука. 588 с.
Кудрявцев С.М., 1967. Утки Московского зоопарка, живущие на полной свободе // Животное население Москвы и Подмосковья, его изучение, охрана и направленное преобразование. М.: Изд-во МГУ. С. 86–89.
Кузиков И.В., 2020. Водоплавающие и околоводные птицы Большого Строгинского залива Москвы-реки и его окрестностей // Русский орнитологический журнал. Т. 29. Вып. 1915. С. 1839–1869.
Ларионов В.Ф., 1953. Географическая изменчивость размножения кряквы // Перелеты птиц в европейской части СССР. Рига. С. 155–171.
Ларионов В.Ф., 1956. О годовых колебаниях в сроках размножения кряквы // Зоологический журнал. Т. 35. № 1. С. 89–95.
Ларионов В.Ф., 1959. Соотношение времени прилета и размножения кряквы // Орнитология. № 2. С. 32–34.
Мальчевский А.С., Пукинский Ю.Б., 1983. Хохлатая чернеть // Птицы Ленинградской области и сопредельных территорий. Т. 1. Л.: Изд-во ЛГУ. С. 100–104.
Михантьев А.И., Селиванова М.А., 2018. Связь динамики численности гнездящихся на юге Западной Сибири уток с глобальными климатическими колебаниями // Первый Всероссийский орнитологический конгресс. Тезисы докладов. Тверь. С. 224–225.
Михельсон Х.А., Меднис А.А., Блум П.Н., 1977. Изучение демографии гнездовых популяций методом кольцевания // Методы изучения миграций птиц. Материалы Всесоюзной школы-семинара. М. С. 46–61.
Михельсон Х.А., Меднис А.А., Блум П.Н., 1986. Популяционная экология мигрирующих уток Латвии. Ред. Виксне Я.А.. Рига: Зинатне. 111 с.
Мищенко А.Л., 1983. Значение рыборазводных прудов для сохранения орнитофауны в условиях антропогенного ландшафта Московской области // Бюллетень МОИП. Отд. биол. Т. 83. № 4. С. 15–25.
Морозов Н.С., Худяков В.В., Панфилова И.М., 2016. Рябинник Turdus pilaris в большом городе: особенности размножения и репродуктивные потери из-за хищников // Русский орнитологический журнал. Т. 25. № 1267. С. 1114–1115.
Нестеров Е.С., 2013. Североатлантическое колебание: атмосфера и океан. М.: Триада-лтд. 144 с.
Онно С., 1974. Время гнездования у водоплавающих и прибрежных птиц в Матсалуском заповеднике (Эстонская ССР) // Сообщения Прибалтийской комиссии по изучению миграций птиц. № 8. Тарту: Изд. АН ЭССР. С. 107–155.
Остапенко В.А., Виноградов С.И., 1984. Состояние и перспективы работы Московского зоопарка по обогащению фауны Москвы некоторыми видами гусеобразных // Современное состояние ресурсов водоплавающих птиц. Тезисы Всесоюзного семинара 20–23 октября 1984 г. С. 303–305.
Остапенко В.А., Виноградов С.И., Березина М.Ф., Курилович Л.Я., 1989. Свободноживущие утки Московского зоопарка // Экология и охрана диких животных. Межвузовский сборник научных трудов. Госагропром СССР. МВА им. К.И. Скрябина. М. С. 39–48.
Попова-Бондаренко Е.Д., 1986. Гнездование чайковых птиц в Москве // Изучение птиц СССР, их охрана и рациональное использование. Тезисы IX Всесоюзной орнитологической конференции. Ч. 2. Л. С. 161–162.
Птушенко Е.С., Иноземцев А.А., 1968. Хохлатая чернеть // Биология и хозяйственное значение птиц Московской области и сопредельных территорий. М.: Изд-во МГУ. С. 84–85.
Пыжьянов С.В., Березовская А.О., 2010. Особенности гнездовой биологии хохлатой чернети Aythya fuligula L., 1758 в стабильных условиях среды // Байкальский зоологический журнал. № 4. С. 56–64.
Садоков В.П., Козельцева В.Ф., Кузнецова Н.Н., 2012. Определение весенних дат устойчивого перехода средней суточной температуры воздуха через 0°, +5°С, их прогноз и оценка // Труды гидрометеорологического НИЦ РФ. Под. ред. Докт. физ.-мат. наук М.А. Толстых. Вып. 348. С. 144–152.
Соколов Л.В., 1991. Филопатрия и дисперсия птиц // Труды Зоологического института АН СССР. Т. 230. Л. ПО-3 Ленуприздата. 232 с.
Соколов Л.В., 2010. Климат в жизни растений и животных. СПб.: Изд-во ТЭССА. 344 с.
Суханова О.В., 2005. Тенденции динамики численности уток на рыборазводных прудах в изменившихся социально-экономических условиях // Гусеобразные птицы Северной Евразии. Тез. докл. 3-го Международ. симп. СПб.: 251–252.
Суханова О.В., Мищенко А.Л., 2008. Тенденции динамики численности уток на рыборазводных прудах в изменившихся социально-экономических условиях // Казарка: бюллетень Рабочей группы по гусеобразным Северной Евразии, 11-2. С. 115–123.
Схилтхёйзен М., 2021. Дарвин в городе: как эволюция продолжается в городских джунглях. М.: ЭКСМО. 352 с.
Формозов А.Н., 1981. Взаимоотношения водоплавающей дичи, чаек, куликов и некоторых хищников на гнездовьях в открытых ландшафтах СССР // Проблемы экологии и географии животных. М.: Наука. С. 312–318.
Фридман В.С., Еремкин Г.С., 2009. Урбанизация “диких” видов птиц в контексте эволюции урболандшафта. М.: Книжный дом ЛИБРОКОМ. 240 с.
Холопцев А.В., Астафьева Д.А., Никифорова М.П., 2015. Атмосферная циркуляция в северном полушарии и североатлантическое колебание // Scientific Journal “ScienceRise”. № 8/1 (13). С. 37–46.
Baratti M., Baccetti N., Cordaro M., Mori A., Dessì-Fulgheri F., 2015. Investigating the puzzling genetic structure of mallard populations (Anas platyrhynchos L.) in Italy // European journal of wildlife research. V. 1. № 1. P. 81−89.
Blums P., Nichols J., Hines J., Mednis A., 2002. Sources of variation in survival and breeding site fidelity in three species of European ducks // Journal of Animal Ecology. V. 71. № 3. P. 438–450.
Gollop J.B., Marshall W.H., 1954. A guide to aging duck broods in the field // Mississippi Flyway Council Technical Section. Northern Prairie Wildlife Research Center Online. [Электронный ресурс] URL: http://www.npwrc.usgs.gov/resource/birds/ageduck/ index.htm (Version 14NOV97).
Guillemain M., Poysa H., Fox A.D., Arzel C., Dessborn L. et al., 2013. Effects of climate change on European ducks: what we know and what do we need to know? // Wildlife Biology. V. 19. P. 404–419.
Gunnarson G., Waldenstrom J., Fransson T., 2012. Direct and indirect effects of winter harshness on the survival of Mallards Anas platyrhynchos in northwest Europe // Ibis. V. 154. № 2. P. 307–317.
Hill D.A., 1984. Laying date, clutch size and egg size of the Mallard Anas platyrhynchos and Tufted Duck Aythya fuligula // Ibis. V.126. № 4. P. 484–495.
Johnson M.T., Munshi-South J., 2017. Evolution of life in urban environments // Science. V. 358. P. 607–618.
Kauppinen J., 2012. Pochard and tufted duck in Central Finland – from winners to losers // Suomen Riista. V. 58. P. 42–49. (In Finnish with English summary)
Kelcey J.G., Rheinwald G. (eds), 2005. Birds in European Cities. St. Katharinen. Ginster Verlag. 486 p.
Lehikoinen A., Jaatinen K., Vahatalo A.V., Clausen P., Crowe O. et al., 2013. Rapid climate driven shifts in wintering distributions of three common waterbird species // Global change biology. V. 19. № 7. P. 2071–2081.
Liordos V., Lauder A.W., 2015. Factors Affecting Nest Success of Tufted Ducks (Aythya fuligula) Nesting in Association with Black-Headed Gulls (Larus ridibundus) at Loch Leven, Scotland // Waterbirds. V. 38. № 2. P. 208–213.
Minias P., Włodarczyk R., Minias A., Dziadek J., 2017. How birds colonize cities: genetic evidence from a common waterbird, the Eurasian coot // Avian biology. V. 48. № 8. P. 1095–1103.
Oja H., Pöysä H., 2007. Spring phenology, latitude, and the timing of breeding in two migratory ducks: implications of climate change impacts // Annales Zoologici Fennici. V. 44. P. 475−485.
Pirkola M.K., Högmander J., 1974. Sorsapoikueiden iänmääritys [The age determination of duck broods in the field] // Suomen Riista. V. 25. P. 50–55. [In Finnish with English summary]. Цит пo: Oja, Pöysä, 2007.
Savard J.-P.L., 1985. Evidence of long-term pair bonds in barrow’s goldeney (Bucephala islandica) // Auk. V. 102. № 2. P. 389−391.
Švažas S., Dagys M., Sydelis R., Raudonikis L., 2001. Changes in numbers and distribution of wintering waterfowl populations in Lituania in the 20th century // Acta Zoologica Lituanica. V. 11. № 3. P. 243−254.
Väänänen V.M., 2000. Predation risk associated with nesting in gull colonies by two Aythya species: observations and an experimental test // Journal of Avian Biology. V. 31. № 1. P. 31–35.
Väänänen V.M., Pöysä H., Runko P., 2016. Nest and brood stage association between ducks and small colonial gulls in boreal wetlands // Ornis Fennica. T. 93. № 1. P. 47–54.
Van Dongen W.F.D., Robinson R.W., Weston M.A., Mulder R.A., Guay P.J., 2015. Variation at the DRD4 locus is associated with wariness and local site selection in urban black swans // Evolutionary Biology. V. 15. P. 253–263.
Viksne J., Švažas S., Czajkowski A., Janaus M., Mischenko A. et al., 2010. Atlas of Duck population In Eastern Europe. Vilnius: “Akstis”. 199 p.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал