

Зоологический журнал, 2022, T. 101, № 6, стр. 679-686
Особенности вокализации и поведения пустынной славки (Sylvia nana, Sylviidae, Aves) в связи с ее положением в роде Sylvia
А. С. Опаев a, *, А. В. Матюхин a
a Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: aleksei.opaev@gmail.com
Поступила в редакцию 01.04.2021
После доработки 03.08.2021
Принята к публикации 04.08.2021
- EDN: BYNYPV
- DOI: 10.31857/S0044513422040092
Аннотация
Согласно молекулярно-генетическим данным, пустынная славка занимает обособленное положение в роде славок Sylvia. Чтобы подтвердить или не подтвердить это, мы описали вокализацию и некоторые особенности поведения пустынной славки, а затем сопоставили эти данные с материалами по славкам-завирушкам и серой славке. Оказалось, что пение пустынной славки своеобразно. У этого вида (1) маленький репертуар (1 тип песни на всех изученных фонограммах), (2) песни занимают узкий частотный диапазон, т.к. состоят из удлиненных тоновых звуков со слабовыраженной частотной модуляцией, (3) многие песни предваряет позывка. Все эти особенности отличают данный вид от других славок и подтверждают его обособленное положение в роде Sylvia. Некоторым своеобразием, возможно, характеризуется также территориальное поведение. А вот брачное поведение очень похоже на таковое у других славок.
Пустынная славка (Sylvia nana (Hemprich et Ehrenberg 1833)) распространена преимущественно в Средней и Центральной Азии, от Каспийского моря (включая дельту Волги и северо-восточный Иран) до центральной Монголии и северо-восточного Китая (Волчанецкий, 1954; Козлова, 1975; Cramp, 1992; Ковшарь, 2012; del Hoyo, Collar, 2016). Ранее в состав этого вида включали сахарскую славку (Sylvia deserti), оседло живущую на северо-западе Сахары. Сейчас эти два аллопатричных вида обычно считают отдельными, хотя и очень близкими (del Hoyo, Collar, 2016; Schweizer, 2020).
Положение пустынной славки в роде Sylvia понятно не до конца. В роде выделяют две группы видов (Blondel et al., 1996; Böhning-Gaese et al., 2003; Voelker, Light, 2011; Abdilzadeh et al., 2019). К первой относятся славка-черноголовка (S. atricapilla), садовая славка (S. borin) и несколько близких к ним африканских видов. Ко второй – все прочие славки, включая пустынную. Козлова (1975) сближала интересующий нас вид со славкой-завирушкой (S. curruca sensu lato). Данные по ДНК-ДНК гибридизации (Blondel et al., 1996) также говорят о некотором (но не очень близком) родстве со славкой-завирушкой, а также с певчей славкой (S. hortensis) и родственными им видам. По результатам секвенирования ряда генов, как митохондриальной, так и ядерной ДНК, пустынная славка не имеет близких родственников (за исключением сахарской славки) и занимает базальное положение во второй группе видов рода Sylvia (Böhning-Gaese et al., 2003; Voelker, Light, 2011; Abdilzadeh et al., 2019). Однако в одной работе анализ последовательности гена цитохрома b также сблизил пустынную славку со славкой-завирушкой (Abdilzadeh et al., 2019).
Таким образом, пустынная славка, вероятнее всего, обособленна относительно других славок. А значит она должна обладать теми или иными уникальными, отличными от других Sylvia особенностями. В этой связи полезно обратить внимание на вокализацию и поведение – ведь это группы признаков, которые у птиц эволюционируют быстрее других, т.е. по этим группам признаков родственные виды потенциально различаются в наибольшей степени (Панов, 1978). Наличие таких особенностей подтвердит обособленность пустынной славки, выявленную молекулярно-генетическими методами.
Обзор основных особенностей биологии пустынной славки сделал Ковшарь (2012). Однако ее поведение изучено недостаточно полно (Cramp, 1992; Ковшарь, 2012), а вокализация описана лишь в самых общих чертах (Козлова, 1975; Cramp, 1992; Wallschläger, 1994; Ковшарь, 2012; Boesman, 2016).
В данном сообщении мы приводим сведения по вокализации и поведению пустынной славки. Наша задача состояла в том, чтобы выявить и подчеркнуть уникальные особенности этого вида, подтверждающие ее обособленное положение в роде Sylvia. Для этого мы провели пилотное сопоставление некоторых из представленных данных с аналогичными материалами по славкам-завирушкам (как, предположительно, относительно близким к интересующему нас виду), а также по серой славке (S. communis). Славок-завирушек мы при этом понимали как комплекс видов, включающий собственно славку-завирушку, или славку-мельничка (S. curruca sensu stricto), горную славку (S. althaea) и кашгарскую славку (S. minula) (Olsson et al., 2013; Votier et al., 2016).
МАТЕРИАЛ И МЕТОДЫ
В основу работы положены записи разных вокализаций пустынной славки с сайта xeno-canto.org. Всего проанализировано 11 фонограмм (от девяти разных птиц) общей длительностью около 8 мин. Эти записи содержат 41 песню (собственно песню и песню-“колокольчик”: см. ниже) и 36 позывок-трелей (см. ниже) (табл. 1). Для сравнения мы использовали записи пения трех видов славок: серой, горной и славки-мельничка (2 подвида с заметно различным пением). Эти записи сделаны Опаевым в Ростовской обл. (село Кагальник, 8–10 июня 2008 г.: 3 самца серой славки), Таджикистане (ущелье Кондара, 3–20 июня 2010 г.: 4 самца горной славки) и Казахстане (Алматинская обл., 1–4 мая 2015 г.: 4 самца туркестанской славки-мельничка Sylvia curruca halimodendri). Записи пения трех самцов европейского подвида славки-мельничка (S. c. curruca) загружены с сайта xeno-canto.org. Эти фонограммы получены в Нидерландах (запись XC140038) и Польше (записи XC236803 и XC562898).
Таблица 1.
Каталог записей пустынной славки с сайта xeno-canto.org, использованных в работе
| Дата, место | Номер | Колич. песен | Колич. позывок | Длительность, с | Автор |
|---|---|---|---|---|---|
| Oman, Muntasar oasis, 29.01.2010 | XC44450 | 0 | 3 | 9 | Herman van Oosten |
| Kazakhstan, Uygur Distict, Almaty Province, 15.05.2002* | XC49767 | 6 | 0 | 40 | D. Farrow |
| Oman, Barr al Hikman, 17.11.2012 | XC118140 | 1 | 27 | 68 | Thijs Fijen |
| Kazakhstan, Shygan, 29.05.2011* | XC120526 | 6 | 0 | 71 | Patrick Franke |
| Iran, Golestan, 01.03.2001 | XC329367 | 3 | 5 | 84 | Manuel Schweizer |
| Iran, Khar Turan National Park, Shahrud, Semnan, 18.04.2017 | XC405207 | 5 | 0 | 28 | Patrick Aberg |
| Iran, Khar Turan National Park, Shahrud, Semnan, 18.04.2017 | XC405208 | 3 | 0 | 24 | Patrick Aberg |
| Saudi Arabia, Al Madinah Province, 12.11.2018 | XC446982 | 1 | 1 | 13 | David Darrell-Lambert |
| Mongolia, Sevrei, Umnugovi, 28.05.2019* | XC481529 | 5 | 0 | 37 | Frank Lambert |
| Mongolia, Sevrei, Umnugovi, 28.05.2019* | XC481530 | 9 | 0 | 76 | Frank Lambert |
| Saudi Arabia, Judah, Al Ahsa, Eastern Province, 04.12.2020 | XC607066 | 2 | 0 | 12 | Gregory Askew |
Для визуализации и измерения песен и звуков использовали программу Syrinx PC version 2.6. При построении спектрограмм использовали окно Блэкмана и длину быстрого преобразования Фурье, равную 256 точкам. Для каждой песни и позывки пустынной славки измеряли длительность (с точностью до 0.01 с), нижнюю и верхнюю границы частотного диапазона (с точностью до 0.01 кГц) (разница последних двух параметров – это частотный диапазон песни). Названные частотные параметры являются не очень хорошими в том смысле, что в значительной степени зависят от качества записи и громкости сигнала (Zollinger et al., 2012) – т.е. погрешность измерения может быть довольно высока. Тем не менее в описательных целях они, несмотря на погрешность, по-нашему мнению подходят даже лучше других, т.к. позволяют получить общее представление о частотных параметрах песни того или иного вида.
У песен трех других видов славок измеряли только частотный диапазон. Для каждого самца измерено по 5 песен, а затем рассчитана медиана. Всего измерены частотные диапазоны 70 песен – серой славки, горной славки и двух подвидов славки-мельничка (суммарно – 14 самцов всех видов).
Полевые наблюдения за пустынной славкой проведены в Волго-Уральских песках, а именно – в северо-западном Казахстане (Уральская обл., Джангалинский р-н – ныне Жангалинский р-н Западно-Казахстанской обл.) в марте–мае 1977 г. в урочище Айбас и в мае 1984 г. в окрестностях стационара Кзыл-Капкан. Здесь пустынная славка занимает закрепленные пески, заросшие разреженной кустарниковой растительностью, в основном тамариском Tamarix sp. и джузгуном Calligonum sp. В понижениях местами растет пустынное высокотравье, где славки любят устраивать гнезда. Мы находили гнезда этого вида в зарослях песчаного овса, или волосенца кистистого (Leymus racemosus), а Гаврилов с соавторами (1968) указывают на песчаную полынь (Artemisia arenaria) как наиболее частое место гнездования в районе исследования.
Биология и фенология гнездования пустынной славки в Волго-Уральских песках неплохо изучены (Гаврилов и др., 1968; Матюхин и др., 2000; Матюхин, Пыхов, 2005). В 1960–1980 гг. она являлась здесь обычной птицей (Гаврилов и др., 1968; Матюхин, Пыхов, 2005). Впоследствии численность, судя по всему, сократилась. В 2007 г. Матюхин с коллегами проводил учеты птиц в песках Жангалинского р-на, в том числе с автомобиля. Пустынная славка была встречена лишь однажды: 12 июня 2007 г. в урочище Коктау (Матюхин и др., 2010). А наблюдая за птицами в том же урочище 9–13 мая 2010 г., Опаев (неопубл. данные) не отметил эту птицу вовсе.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Вокализация
В ходе наших наблюдений мы отметили у пустынной славки 4 основные вокализации: (1) позывка-трель, (2) песня, (3) песня-“колокольчик” и (4) песня во время токового полета. Три из них (кроме пения во время токового полета) имеются в интернет-коллекции сайта xeno-canto.org (рис. 1). Каждая из этих трех вокализаций была записана как на зимовках, так и на местах гнездования.
Рис. 1.
Вокализация пустынной славки: песня со вступительной позывкой (а1 и б1) и без нее (а2); песня-“колокольчик” со вступительной позывкой (в2 и г1) и без нее (г2); позывки (б2 и в1). Вокализации каждой строки, разделенные пунктирной линией, взяты из непрерывных фонограмм (с сайта xeno-canto.org) из: а – Казахстана (XC120526, 29.05.2011), б – Ирана (XC329367, 1.03.2001), в – Саудовской Аравии (XC446982, 12.11.2018), г – Монголии (XC481529, 28.05.2019). Фонограммы б и в получены на зимовке, а и г – на местах гнездования.
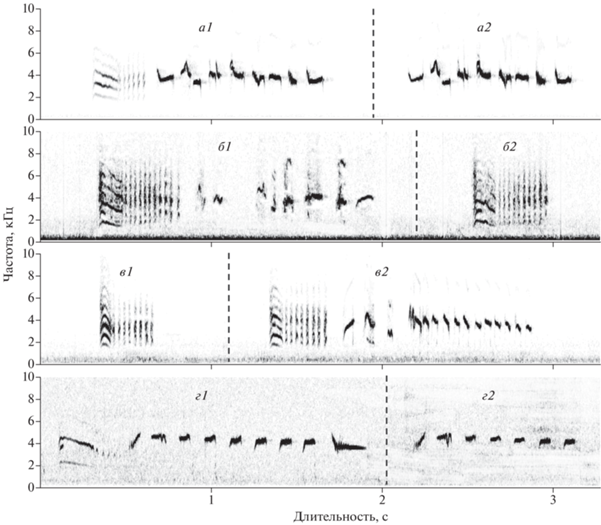
Позывка-трель – плотная секвенция коротких (около 20–30 мс) широкополосных звуков с гармоническим спектром заполнения. Медианная длительность у разных самцов (n = 4) нашей выборки 0.3–0.7 с, медианная нижняя граница частотного диапазона 1.4–1.7 кГц, а верхняя 6.9–7.9 кГц (всего измерено 36 позывок у четырех птиц). Во многих, но не во всех случаях первый звук трели удлинен (до 150 мс) и отличается по структуре от последующих (рис. 1, б2, в1).
Аналогичные позывки имеются в репертуарах других палеарктических славок (в т.ч. из группы славок-завирушек: наши наблюдения). Но обычно в них отсутствует удлиненный вступительный звук. Ситуации, в которых славки, в том числе пустынные, издают этот звук, многообразны. У пустынных славок позывку-трель чаще можно услышать при тревоге, а в некоторых случаях – также при взаимодействии самца и самки. Вероятно, это сигнал, указывающий на общий подъем “возбуждения” самца. Самки также издают позывку-трель, но значительно реже. Похожим образом используют позывку-трель и другие палеарктические Sylvia.
Особенность же пустынной славки в том, что у этого вида позывкой-трелью часто начинается песня, в том числе – песня-“колокольчик” (см. ниже). Для других славок (например, серой, славки-мельничка и горной) это не характерно.
Песня пустынной славки (рис. 1, а1, а2, б1) часто состоит из двух частей: позывки-трели (почти всегда – с удлиненным первым звуком) и основной части, представляющей собой секвенцию тоновых звуков (рис. 1, а1, б1). Нередки песни, состоящие только из основной части (рис. 1, а2). В нашей выборке с сайта xeno-canto.org они составили 59% (n = 17) и наблюдались у двух самцов из четырех (песни с позывкой-трелью зафиксированы у трех птиц). Медианная длительность песен с позывкой-трелью у разных самцов 1.1–1.6 с (n = 3), а без нее – 1.1–1.3 с (n = 2). Таким образом, пустынная славка может исполнять позывку-трель и основную часть песни порознь, а может объединять их в одну конструкцию. Данная особенность отличает этот вид от других славок.
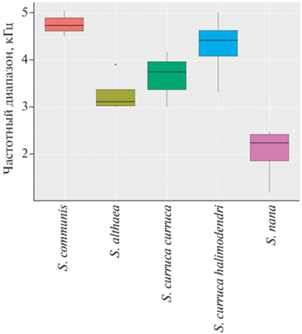
Еще одна особенность рассматриваемого вида кроется в структуре основной части песни. Она состоит обычно из 10–15 тоновых узкополосных звуков, большинство из которых незначительно модулированы по частоте. За счет такой структуры вся песня в целом занимает относительно узкий частотный диапазон. У четырех самцов медианная нижняя граница диапазона варьировала в пределах 2.2–3.1, а верхняя 4.3–5.5 кГц. По-иному устроена песня у других видов. В пении многих из них заметную долю составляют глубоко-модулированные по частоте тоновые звуки, а также широкополосные посылки с шумовым либо гармоническим спектром заполнения (рис. 2). Поэтому частотный диапазон их песен существенно шире (рис. 3). Судя по спектрограммам, приведенным в сводке Крэмпа (Cramp, 1992), сказанное справедливо и для других европейским Sylvia, а не только для славок-завирушек и серой славки.
Рис. 2.
Песни разных видов славок – для каждого вида представлено по две песни разных самцов: а – пустынная славка (приведены для примера две песни, представленные также на рис. 1, а2 и г2); б – славка-мельничек (по фонограммам из Нидерландов и Польши с сайта xeno-canto.org); в – туркестанская славка-мельничек (по фонограммам Опаева из южного Казахстана); г – горная славка (по фонограммам Опаева с Памиро-Алая, Таджикистан); д – серая славка (по фонограммам Опаева из Ростовской обл.).
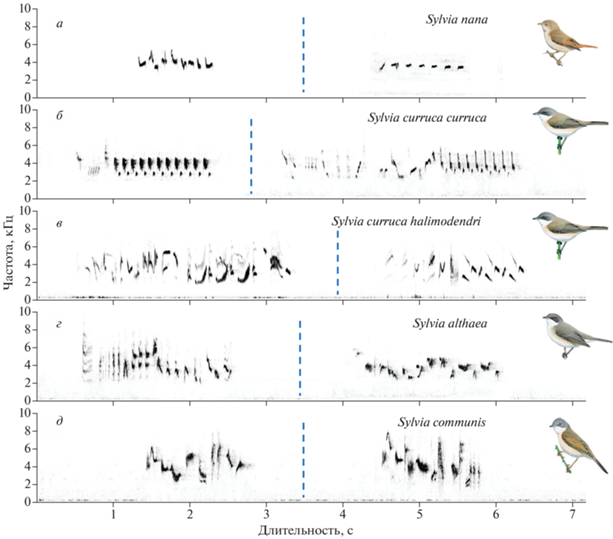
Рис. 3.
Частотный диапазон песен четырех видов славок. Показаны медианы, квартили (цветные четырёхугольники) и разброс значений. График построен по медианным значениям для каждого индивидуума: всего 3–4 самца для каждого вида/подвида.
Наконец, третьей особенностью пения пустынной славки является его структурная простота. Мы имеем в виду, что на всех изученных фонограммах выявлен только один стереотипный тип песни (хотя могут быть варианты: например, с позывкой-трелью и без нее). Конечно, данных недостаточно: самая длинная из изученных фонограмм содержала всего 14 песен. Однако такую простоту отмечают и другие авторы (Ковшарь, 2012). Что же касается других славок, то их репертуары обширнее, и на фонограммах это сразу заметно. Дело в том, что самцы серой славки, славки-мельничка и горной славки если и повторяют одну и ту же песню, то обычно не более 2–5 раз (наши наблюдения). Поэтому ни одна из просмотренных фонограмм этих видов не содержала песни только одного типа, как это было отмечено на всех записях пустынной славки. В литературе имеются довольно подробные сведения о пении серой славки и славки-мельничка (Balsby, 2000; Klit, 1999), а также очковой славки (Sylvia conspicillata) (Palmero et al., 2014). Манера пения этих видов дискретная, но стереотипные типы песен выделить не удается: в песне имеется вариабельная и более консервативная часть. Каждый самец имеет репертуар из многих типов звуков, которые он комбинирует в отдельные песни в соответствии с определенными правилами и собственными предпочтениями. Тем не менее изменчивость песен одной птицы может быть довольно высока. Конкретные размеры репертуаров звуков составляют: 33–111 типов у славки-мельничка (в среднем 66 типов: речь идет о “щебечущей” части песни; помимо нее имеется однообразный заключительный “стукоток”) (Klit, 1999), 33–67 типов у очковой славки (Palmero et al., 2014) и несколько сотен типов у серой славки (Balsby, 2000).
Песня-“колокольчик” напоминает звон маленького колокольчика (Волчанецкий, 1954). Этот звук очень характерен для пустынной славки, и его отмечают многие орнитологи (Ковшарь, 2012). Эта песня также может исполняться с позывкой-трелью (рис. 1, в2, г1) и без нее (рис. 1, г2). У трех самцов (из четырех) все песни-“колокольчики” начинались позывкой-трелью, а у одного были песни как с этой позывкой, так и без. Основная часть песни-“колокольчика” (т.е. песня без позывки-трели) – это секвенция сходных по структуре тоновых звуков. Секвенция нисходящая: каждый последующий звук обычно ниже по частоте, чем предыдущий. В ряде случае основная часть песни-“колокольчика” может начинаться или заканчиваться звуками иной структуры (например: рис. 1, в2). Медианная длительность основной части песни-“колокольчика” у разных самцов (n = 4) нашей выборки 1.3–1.7 с, медианная нижняя граница частотного диапазона 2.4–3.3, а верхняя 5.1–5.3 кГц (всего измерено 24 песни четырех птиц). Таким образом, по сравнению с собственно песней, частотный диапазон песни-“колокольчика” ýже, кроме того, она смещена в более высокочастотную область. Явных аналогов песни-“колокольчика” у других славок мы не отметили.
Песню-“колокольчик” мы зафиксировали на четырех фонограммах. При этом на одной фонограмме ни разу не были отмечены одновременно и собственно песня и песня-“колокольчик”. Судя по нашим наблюдениям в природе, в репертуаре каждого самца пустынной славки имеется по меньшей мере один тип песни и один тип песни-“колокольчика”. Эти вокализации могут чередоваться в составе одной последовательности, но некоторые ситуативные различия между ними, вероятно, есть (поэтому в нашей выборке фонограмм они не встречены вместе в одной записи). Так, по нашим наблюдениям, песня-“колокольчик” чаще используется самцами при патрулировании границ.
По нашим наблюдениям, для пустынной славки характерны токовые полеты [по данным Ковшаря (2012), они отмечаются редко]. Представление о частоте токовых полетов дают следующие данные. 10–13 мая 1984 г. мы проводили наблюдения за гнездом пустынной славки, у которого держались самец и самка. Откладка яиц, судя по всему, была начата 14 мая: 15 мая в гнезде было 2 яйца. Наблюдения вели из скрадка в течение всего дня, подсчитывая песни, спетые самцом неподалеку от гнезда. Всего зафиксировано 597 песен, из них 11 (1.8%) были исполнены во время токового полета.
Токовой полет пустынной славки происходит так. Самец плавно набирает высоту, а затем с пением снижается. Траектория полетов изменчива: высота колеблется от 5–6 до 20 м (чаще – около 10 м), а длина полета (точнее – ее проекция на землю) – от 3 до 40 м. Самец начинает петь перед самой верхней точкой траектории и продолжает вокализировать при снижении. Записями пения при токовом полете мы не располагаем. По данным визуальных наблюдений, основу составляет удлиненный вариант обычной песни. Но часто, особенно в конце, можно услышать песню-“колокольчик”. Подобные токовые полеты известны и у ряда других палеарктических славок (Cramp, 1992). Так, они очень характерны для серой славки (Cramp, 1992; наши наблюдения), отмечены у туркестанской славки-мельничка (наши наблюдения в Алматинской обл. Казахстана в мае 2015 г.), но при этом крайне редки у европейской славки-мельничка (Cramp, 1992).
Особенности территориального и брачного поведения
Пустынная славка – территориальный моногамный вид (Ковшарь, 2012). В начале периода гнездования нами отмечено поведение, напоминающее патрулирование границ: самец перемещался по территории по одному и тому же маршруту. Конфронтации соседних самцов близ границ их участков мы наблюдали в мае 1984 г. Такие конфронтации могут происходить в виде драк (наблюдали 4 мая (дважды), 5 и 8 мая) либо ритуализованных конфликтов (отмечали 5 и 8 мая). В последнем случае при встрече двух самцов на границе они перемещались один возле другого по песку в течение 5–7 мин в одинаковых позах. Поза это своеобразна: хвост, перпендикулярный поверхности земли, периодически опускается вниз и затем снова забрасывается вверх; голова и шея вытянуты вверх параллельно хвосту, отмечаются постоянные поклоны; крылья слегка приспущены и периодически вздрагивают. Интересно, что подобная поза однажды отмечена у славки-мельничка – при этом в целом поза не характерна для этого хорошо изученного вида (Cramp, 1992). У других славок указаний на такую мы не нашли, не наблюдали ее и мы сами. Отметим, что столь выразительные позы вообще редки у славок, моторика сигнального поведения которых выглядит довольно бедной. Таким образом, территориальное поведение пустынной славки возможно характеризуется некоторым своеобразием, но данных недостаточно.
В то же время некоторые черты брачного поведения пустынной славки очень похожи на таковые у других Sylvia. Как и у многих других видов, первыми прилетают самцы. Прежде чем пытаться привлечь самку, самец выстраивает на своем участке так называемое сигнальное гнездо. Сигнальное гнездо (обычно только одно) строят холостые самцы и серой славки и славки-мельничка. У всех этих трех видов такое гнездо отличается от готового гнезда только отсутствием выстилки лотка, которой занимается уже самка (Музаев, 1980, 1980а; Ковшарь, 2012). Во время строительства сигнального гнезда самец пустынной славки поет, часто довольно активно – как сразу же после укладки в гнездо очередной порции материала, так и в перерывах между строительством (Волчанецкий, 1954; Ковшарь, 2012). В целом аналогично ведут себя в такой ситуации и самцы серой славки. Любопытно, что, по нашим наблюдениям, они могут иногда петь даже с гнездовым материалом в клюве. У этого вида строительство сигнального гнезда тесно переплетено с демонстративным поведением, а при появлении самки такое строительство становится важным компонентом ухаживания (неопубл. наблюдения Опаева в Ростовской обл. в мае 2001–2002 гг.). Судя по всему, у пустынной славки в общих чертах все происходит таким же образом (Ковшарь, 2012).
Насиживают у пустынной славки оба партнера, причем у самца (как и у самки) образуется наседное пятно (Рустамов, 1954; Волчанецкий, 1954; Елисеев, 2003; Ковшарь, 2012). Явное наседное пятно образуется также у самцов славки-мельничка (и некоторых других видов), но не у самцов серой славки, которые также принимают участие в насиживании (Ефремов, Паевский, 1973).
ЗАКЛЮЧЕНИЕ
В данном сообщении мы провели пилотное сопоставление вокализации и поведения пустынной славки с аналогичными признаками нескольких родственных видов. Оказалось, что заметным своеобразием отличается структура пения этого вида. Уникальные черты свойственны, возможно, также территориальному поведению.
В систематике воробьиных птиц, наряду с морфологией, широко используется анализ ДНК и вокализации – главным образом, пения (Martens et al., 2011; del Hoyo, Collar, 2016). Согласно современным представлениям в русле “интегральной таксономии” (“integrative taxonomy”: Sangster, 2018), систематическое положение таксона определено более четко тогда, когда разные группы признаков (например, молекулярно-генетические и акустические) соответствуют друг другу. А использование только одной группы признаков повышает вероятность ошибки. Мы проверили, подтверждают ли акустические и этологические данные обособленное положение пустынной славки в роде Sylvia, выявленное при анализе ДНК. По нашему мнению – подтверждают (главным образом, речь идет о структуре пения). Таким образом, пустынная славка, вероятно, действительно обособлена относительно других славок.
Список литературы
Волчанецкий И.Б., 1954. Род Славка // Птицы Советского Союза. Т. VI. М.: Советская наука. С. 330–387.
Гаврилов Э.И., Наглов В.А., Федосенко А.К., Шевченко В.Л., Татаринова О.М., 1968. Об орнитофауне Волжско-Уральского междуречья (воробьиные) // Новости орнитологии Казахстана. Труды Института зоологии. Т. 29. Алма-Ата. С. 153–207.
Елисеев Д.О., 2003. К биологии пустынной славки (Sylvia nana) в северном Приаралье // Функциональная морфология, экология и жизненные циклы животных. Санкт-Петербург. Вып. 3. С. 116–123.
Ефремов В.Д., Паевский В.А., 1973. Поведение насиживания и наседные пятна самцов у пяти видов птиц рода Sylvia // Зоологический журнал. Т. 52. Вып. 5. С. 721–728.
Козлова Е.В., 1975. Птицы зональных степей и пустынь Центральной Азии. Л.: Наука. 250 с.
Ковшарь А.Ф., 2012. Пустынная славка [Sylvia nana (Hemprich et Ehrenberg, 1833)] в Казахстане и Средней Азии: распространение, численность, биология // Орнитологический выпуск Казахстана и Средней Азии. Вып. 1. С. 139–164.
Матюхин А.В., Танитовский В.А., Шевченко В.Л., 2000. К биологии бормотушки (Hippolais caligata) и пустынной славки (Sylvia nana) в степях Западного Казахстана // Материалы междунар. симпозиума “Степи Северной Азии: стратегия сохранения природного разнообразия и степного природопользования в ХХI веке”. Оренбург. С. 253.
Матюхин А.В., Пыхов С.Г., 2005. О возможных последствиях влияния газодобывающей промышленности на фауну региона (на примере орнитофауны Астраханской области) // Экологические проблемы Полесья и сопредельных территорий. Материалы VII Междунар. науч.-практ. конф. Гомель. С. 120–125.
Матюхин А.В., Бидашко Ф.Г., Парфенов А.В., Верейкин В.И., 2010. К орнитофауне Волго-Уральского междуречья // Орнитология в Северной Евразии. Материалы XIII Междунар. орнитологической конф. Северной Евразии. Оренбург. С. 213–214.
Музаев В.М., 1980. Сравнительная экология, территориальное поведение и годовые циклы некоторых представителей рода Sylvia (Aves). Автореф. дис. … канд. биол. наук. Ленинград. 21 с.
Музаев В.М., 1980а. Явление бигамии у серой славки // Доклады Высшей школы. Биологические науки. № 12. С. 38–42.
Панов Е.Н., 1978. Механизмы коммуникации у птиц. М.: Наука. 306 с.
Рустамов A.К., 1954. Птицы пустыни Кара-Кум // Ученые записки Туркменского университета. Вып. 2. С. 7–344.
Abdilzadeh R., Aliabadian M., Olsson U., 2019. A molecular assessment of the taxonomy of Iranian Sylvia warblers (Aves; Sylviidae) // Journal of Genetic Resources. V. 5. P. 149–162.
Balsby T.J.S., 2000. Song activity and variability in relation to male quality and female choice in whitethroats Sylvia communis // Journal of Avian Biology. V. 31. P. 56–62.
Blondel J., Catzeflis F., Perret P., 1996. Molecular phylogeny and the historical biogeography of the warblers of the genus Sylvia (Aves) // Journal of Evolutionary Biology. V. 9. P. 871–891.
Boesman P., 2016. Notes on the vocalizations of Desert Warbler (Sylvia nana) // HBW Alive Ornithological Note 260. In: Handbook of the Birds of the World Alive. Barcelona. http://www.hbw.com/node/1251590
Böhning-Gaese K., Schuda M.D., Helbig A.J., 2003. Weak phylogenetic effects on ecological niches of Sylvia warblers // Journal of Evolutionary Biology. V. 16. P. 956–965.
Cramp S. (ed.), 1992. The birds of the Western Palearctic. Handbook of the birds of Europe, the Middle East and North Africa. V. VI. Warblers. Oxford. 728 p.
del Hoyo J., Collar N.J., 2016. HBW and BirdLife International Illustrated Checklist of the Birds of the World. V. II. Passerines. Barcelona. 743 p.
Klit I., 1999. The function of song forms in the Lesser Whitethroat Sylvia curruca // Bioacoustics. V. 10. P. 31–45.
Martens J., Tietze D.T., Päckert M., 2011. Phylogeny, biodiversity, and species limits of passerine birds in the Sino-Himalayan region – a critical review // Ornithological Monographs. V. 70. P. 64–94.
Olsson U., Leader P.J., Carey G.J., Khan A.A., Svensson L., Alström P., 2013. New insights into the intricate taxonomy and phylogeny of the Sylvia curruca complex // Molecular Phylogenetics and Evolution. V. 67. P. 72–85.
Palmero A.M., Espelosín J., Laiolo P., Illera J.C., 2014. Information theory reveals that individual birds do not alter song complexity when varying song length // Animal Behaviour. V. 87. P. 153–163.
Sangster G., 2018. Integrative taxonomy of birds: the nature and delimitation of species // Tietze D.T. (ed.). Bird species. How they arise, modify and vanish. Basel. P. 9–38.
Schweizer M., 2020. Diversification and taxonomy of birds characteristic of the Palearctic desert belt // Dutch Birding. V. 42. P. 191–202.
Voelker G., Light J.E., 2011. Palaeoclimatic events, dispersal and migratory losses along the Afro-European axis as drivers of biogeographic distribution in Sylvia warblers // BMC Evolutionary Biology. V. 11 (163). P. 1–13.
Votier S.C., Aspinall S., Bearhop S., Bilton D., Newton J., Alström P., Olsson U., 2016. Stable isotopes and mtDNA reveal niche segregation but no evidence of intergradation along a habitat gradient in the Lesser Whitethroat complex (Sylvia curruca; Passeriformes; Aves) // Journal of Ornithology. V. 157. P. 1017–1027.
Wallschläger D., 1994. On the song of Sylvia curruca, Sylvia nana and Sylvia communis in Mongolia // Bioacoustics. V. 6. P. 80.
Zollinger S.A., Podos J., Nemeth E., Goller F., Brumm H., 2012. On the relationships between, and measurement of, amplitude and frequency in birdsong // Animal Behaviour. V. 84. P. e1–e9.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал