

Зоологический журнал, 2022, T. 101, № 6, стр. 623-640
Годичный цикл жизни муравейника Formica s. str. (Hymenoptera, Formicidae)
А. А. Захаров a, Р. А. Захаров a, *
a Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: rzakharov@inbox.ru
Поступила в редакцию 24.12.2020
После доработки 20.12.2021
Принята к публикации 22.12.2021
- EDN: RNDNFT
- DOI: 10.31857/S0044513422060137
Аннотация
Предложена схема периодизации годичного цикла жизни социума рыжих лесных муравьев. Схема включает 4 основных и 4 переходных периода, границы которых согласуются с календарем природы европейской части России. Рассмотрена функциональная структура годичного цикла. Каждая из 5 форм деятельности социума – репродуктивная, обеспечения энергоресурсами, гнездовая, демографическая, социоструктурная – представляет собой циклический пул событий, которые в совокупности и образуют годичный цикл его жизни. Периоды облигатно сменяют друг друга, и в каждом предшествующем периоде формируются предпосылки и ресурсы для выполнения следующего. В годичном цикле происходит закономерная смена общих приоритетных задач, для выполнения которых необходимы участие больших масс муравьев и последовательная смена функций у рабочих особей. Эти функции меняются не только в соответствии с возрастным или кастовым полиэтизмом, но и подчиняются приоритетным задачам, что можно определить как сезонный полиэтизм. Показано наличие в семьях рыжих лесных муравьев сезонных функциональных групп рабочих, описаны пути их формирования на примере зимней структуры, состоящей из трех функциональных групп: свиты самок, хранителей запасов и мобильных рабочих. Обосновывается различная продолжительность интервалов формирования зимних и летних функциональных групп у рыжих лесных муравьев.
Освоение муравьями средних и северных широт потребовало от них целого ряда физиологических и поведенческих адаптаций, необходимых для выживания и успешного существования в условиях с холодной зимой и длительным периодом неактивного существования (Heinze, 1993; Heinze, Hölldobler, 1994). Важнейшей спецификой жизни муравьев в условиях умеренного климата является жесткая регламентация годичного цикла, касающаяся всех сторон жизни муравейника. Принимая, что самосохранение – это первостепенная цель любой биосистемы (социума) с момента ее возникновения, полную реализацию свойственного данному виду годичного цикла активности (далее “годичный цикл”) можно считать основным способом достижения этой цели.
Муравьи постоянно заняты решением масштабных и трудоемких задач, от успешности выполнения которых зависят благополучие и само существование всего социума. Большинство таких задач находится в компетенции сферы обеспечения (Захаров, 1991, 2015). Эти задачи связаны с различными сторонами жизни семьи: а) сооружением и поддержанием жилища; б) сохранением и самоидентификацией социума; в) воспроизводством населения; д) обеспечением пищей и сохранением кормовых ресурсов, необходимых для жизни семьи; е) реструктуризацией поселения и расселением; ж) взаимодействием с другими муравейниками в рамках общего комплекса гнезд или многовидового сообщества муравьев.
Цель исследования – описание социума Formica s. str. как биосоциальной системы, жизнедеятельность которой претерпевает специфические изменения в годичном цикле. В соответствии с этим основные задачи статьи следующие:
а) Произвести структуризацию годичного цикла с выделением составляющих его постоянных и переходных периодов.
б) Дать описание годичного цикла жизни муравейника как набора взаимосвязанных форм деятельности семьи, которые в совокупности и обеспечивают сбалансированное существование и развитие муравьиной общины.
в) Охарактеризовать годичный цикл как конвейер приоритетных дел с описанием специфики форм деятельности и их взаимодействия в различные периоды.
г) Описать механизмы, обеспечивающие преемственность периодов в годичном цикле семьи модельной группы муравьев.
Актуальность такого исследования вытекает из обусловленных сезонным климатом закономерных цикличных изменений всего хода жизни муравейника в течение года. Вся его жизнь в условиях умеренного климата протекает в жестких сезонных рамках (Feller, 1989; Gösswald, 1989; Heinze, 1993), касающихся всех сторон жизни муравейника. Детальное исследование годичного цикла жизни и соответствующего фенологического контроля жизни муравейников позволят повысить надежность планирования и проведения полевых экспериментов, мониторинговых и инвентаризационных работ, а также биотехнических мероприятий по сохранению и переселению муравьев (Захаров, 2015).
Феномен годичной цикличности обсуждается в данной работе на примере представителей подрода Formica s. str., в основном это рыжие лесные муравьи группы Formica rufa: F. aquilonia Yarrow, F. lugubris Zett., F. rufa L., F. polyctena Förster., а также близкие к ним виды – луговой муравей F. pratensis Retz. и красноголовый муравей F. truncorum Fabr. Такой выбор определяется двумя моментами. Во-первых, муравьи делятся на две основные экологические группировки – зимующие с расплодом и без него, а Formica s. str. являются наиболее продвинутыми по уровню социальной организации представителями второй из этих групп. И во-вторых, именно Formica s. str. – основная модельная группа в многолетних мирмекологических исследованиях обоих соавторов.
Подходы и методы, использованные в описании жизнедеятельности отдельных муравейников и поселений, основаны на концепции годичного цикла активности социума муравьев (Stearns, 1992).
В работе использованы собственные (Захаров, 1991, 2015; Захаров, 2008; Захаров, Захаров, 2013, 2014, 2018; и др.) и литературные (Длусский, 1967, 2001; Kneitz, 1967, 1969; Feller, 1989; Rissing S.W., Pollock G.B., 1988; Сейма, 2008; Радченко, 2016; Дьяченко, 2017; и др.) данные по биологии муравьев Formica, а также информация о годичных циклах у этих и других муравьев (Heinze, 1993; Heinze, Hölldobler, 1994; Кипятков, Лопатина, 2007; и др.). Привлечены данные многолетнего мониторинга комплексов муравейников в мирмекологическом заказнике “Верхняя Клязьма” Московской обл. (40–45 лет) и в Заповеднике “Пинежский” Архангельской обл. (более 15 лет). Использованы предварительные результаты видеоконтроля фотоловушками, работающими в круглогодичном режиме с 2014 г. в заповеднике “Пинежский” (Захаров, Рыков, 2018). Кроме того, текущие сезонные наблюдения за группами муравейников проводились авторами в различных районах Подмосковья. Параметры и категории состояния отдельных муравейников и их комплексов, а также приемы их описания детально изложены в недавних работах (Захаров и др., 2013, 2015).
В годичном цикле природы Подмосковья выделены 14 периодов (Галахов, 1959), что согласуется с периодизацией, принятой для лесной и лесостепной зон европейской части России. Границы сезонов и фенологических периодов приняты в соответствии с таковыми, установленными на основании метеорологических, зоофенологических, фитоценологических и ландшафтно-физиономических границ (Шульц, 1981; Соловьев, 2005). Минимум ключевых событий, означающих начало или завершение определенных периодов годичного цикла муравейника, можно учесть при периодических внешних осмотрах муравейников, их дорог и кормовых участков (Захаров, 2015; Захаров, Захаров, 2018). Важно, что существенная детализация событийного ряда возможна без значимых вторжений в сами гнезда.
СТРУКТУРА ГОДИЧНОГО ЦИКЛА ЖИЗНИ МУРАВЬЕВ
Годичный цикл имеет четкую структуру, т.е. в течение одного года в жизни муравейника сменяют друг друга качественно различающиеся этапы или периоды – пулы взаимосвязанных событий, имеющих определенную временну́ю привязку. Представить годичный цикл жизни муравейника можно с разной степенью дробности и с разной степенью приближения всего процесса к таковому в природе. Так, в совсем недавно вышедшей статье (Захаров, Захаров, 2018) мы пытались описать жизнь муравейника в течение года как цикл, состоящий из пяти этапов (периодов). При этом подразумевалось, что каждый из них, помимо основного отрезка, включает еще два – начальный и конечный отрезки, в ходе которых и происходит смена трендов в жизни семьи. Этап оказывается событийно неоднородным, что снижает степень адекватности описания процесса реальному ходу событий. Такое деление достаточно и удобно для общего описания процесса, но усложняет его детализацию.
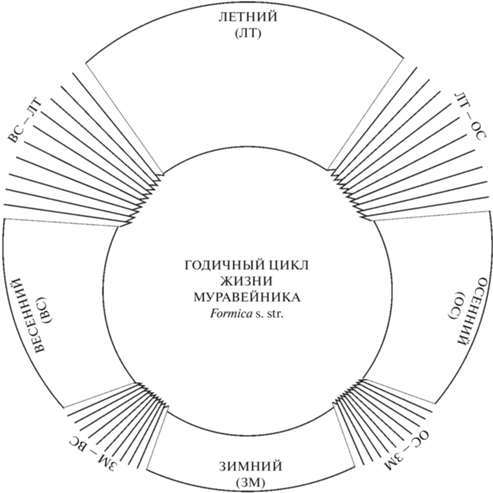
Поэтому в данной публикации исходная схема была трансформирована: периоды были разделены на основные и переходные. Основные периоды характеризуются постоянством общей ситуации в муравейнике и большей продолжительностью. В переходные, достаточно краткосрочные периоды (10–15 дней), проходит радикальная смена векторов развития событий (рис. 1). При этом формируется система из четырех основных периодов и четырех переходных, которые по срокам хорошо согласуются с сезонными сменами в календаре русской природы (Галахов, 1959; Шульц, 1981), что можно расценивать как подтверждение естественности новой схемы.
Рис. 1.
Годичный цикл жизни муравейника Formica s. str. Основные периоды: весенний, летний, осенний, зимний. Переходные периоды: ЗМ–ВС (зимний–весенний), ВС–ЛТ (весенний–летний), ЛТ–ОС (летний–осенний), ОС–ЗМ (осенний–зимний).
Подходы к периодизации годичного цикла у хозяйственно или экономически значимых объектов (например, медоносная пчела) имеют преимущественно утилитарный характер. В зависимости от задач исследования или использования число периодов (этапов) варьирует – от четырех по данным Лаврехина и Панновой (1983) и до восьми по данным Радченко и Песенко (1994).
Основные периоды годичного цикла
Основные, устойчивые периоды (периоды 1–4) качественно отличаются друг от друга по своим характеристикам и имеют достаточную продолжительность, не менее полутора месяцев:
1) весенний период – от пробуждения до вылета крылатых особей (в Южной Европе март–апрель; в Южной тайге европейской части России апрель–май);
2) летний период – после вылета крылатых особей до выведения последнего поколения рабочих (Южная тайга – июнь–август);
3) осенний период – после выхода последнего поколения рабочих до ухода на зимовку (Южная тайга – сентябрь–ноябрь);
4) зимний период – зимовка (ноябрь–март).
Наиболее длинный из них период 4, зимовка длится около 5 мес. Муравьи проводят его в подземной части своего гнезда.
Каждый период имеет выраженную специфику (табл. 1) – в это время муравьи выполняют конкретные задачи. Эти задачи определяют направленность активности (на разные части гнезда, территории и источники пищи, на яйцекладущих самок и появившийся расплод) и ее уровень. И, прежде всего, происходит подбор исполнителей для решения этих задач.
Таблица 1.
Характеристики периодов годичного цикла жизни семьи рыжих лесных муравьев
| Период | Формы деятельности | |||||
|---|---|---|---|---|---|---|
| основной | переходный | репродуктивная | энергетическое обеспечение | гнездовая | демографическая | социоструктурная |
| Зимний–весенний | Начало репродуктивного периода, 1-я яйцекладка | За счет зимовочного пищевого запаса семьи | Разогрев гнезда и приведение его в функциональное состояние | Активизация рабочих, начало перехода от зимнего к летнему составу семьи | Активизация связей между колоннами одной семьи | |
| 1. Весенний | Выращивание весеннего поколения крылатых и 1–2 поколений рабочих | Интенсивный расход зимовочного запаса и постепенный переход на внешние источники питания | Ремонт, восстановление гнезда, установление устойчивого t°-режима во внутреннем конусе | Формирование летнего функционального состава рабочих особей | Формирование инфраструктуры кормового участка и системы межгнездовых связей | |
| Весенний–летний | Ротация самок и восстановление состава рабочих особей | Внешние источники белкового и углеводного питания | Завершение весенней реконструкции гнезда | Максимум численности функциональных групп | Полная развертка территориальных и внутрисемейных структур | |
| 2. Летний | Максимальный уровень продуктивности семьи: 2–3 поколения рабочих, факультативное июльское поколение крылатых | Максимум поступления белковой пищи в муравейник | Оптимизация гнезда, устойчивый t°-режим, пик строительной активности | Максимум общей активности при интенсивном росте численности фуражиров-охотников | Летний период реструктуризации и расселения | |
| Летний–осенний | Завершение репродуктивного периода: выход последнего поколения рабочих | Спад охотничьей активности, переключение на сбор пади (корневые тли) | Начало подготовки подземных зимовочных камер (зимовочного гнезда) | Формирования осеннего состава фуражиров с преобладанием сборщиков пади | Сворачивание территориальных систем | |
| 3. Осенний | Семья существует без расплода, самки перешли в зимовочные камеры | Сбор углеводов и формирование зимовочного запаса семьи | Подготовка гнезда к зимовке | Формирование зимнего функционального состава | Осенний период расселения и реструктуризации. Начало постепенной инкапсуляции муравейников | |
| Осенний–зимний | Репродуктивной активности нет | Переход на питание зимовочным запасом пищи | Зимовочное гнездо готово к зимовке | Зимний функциональный состав сформирован | Семьи инкапсулированы в зимовочных гнездах | |
| 4. Зимний | Репродуктивной активности нет | Питание за счет внутренних ресурсов | Гнездо закрыто | Зимний функциональный состав до весны | Отсутствует | |
Основная специфика каждого из периодов – устойчивость общего тренда активности муравьев в течение данного отрезка времени. И, соответственно этому, устойчивость функционального состава и занятий муравьев в гнезде и на участке.
Между устойчивыми периодами пролегают краткосрочные переходные периоды, в течение которых и происходит смена основных трендов деятельности муравьев от предыдущего состояния к последующему (табл. 1). Переход начинается по завершении предшествующего устойчивого периода, при этом сроки начала нового периода могут быть серьезно скорректированы погодными условиями и состоянием конкретного муравейника. Именно за счет переходных периодов происходит корректировка годичного цикла в различные годы, а также в зональном ряду. Так, средние сроки весенней активизации муравьев в Московской и Архангельской областях различаются всего на 10 дней – соответственно 05.IV и 14.IV. При этом задержка по основным фенологическим событиям в северной тайге (Пинежский заповедник) значительно больше (Федченко, 2008).
ФУНКЦИОНАЛЬНАЯ СТРУКТУРА ГОДИЧНОГО ЦИКЛА
Решаемые муравьями задачи можно представить, как набор специализированных форм деятельности: 1) репродуктивная, 2) обеспечения энергоресурсами, 3) гнездовая, 4) демографическая, 5) социоструктурная. Каждая из этих пяти форм деятельности имеет собственный годичный цикл, совокупность которых и образует годичный цикл жизни муравейника. Большинство таких форм деятельности (кроме первой) относится к компетенции сферы обеспечения (Захаров, 1991, 2015). Кратко охарактеризуем функциональные составляющие годичного цикла.
Репродуктивная деятельность
Данная деятельность обеспечивает само существование муравейника, создание его демографического ресурса и ротацию репродуктивных и рабочих особей. На уровне семьи формирование двух сфер функционирования – репродуктивной и обеспечения – является основным признаком эусоциальности у насекомых.
Сроки лёта крылатых особей регламентированы у всех видов. А процесс выведения рабочих многие привычно считают континуальным, хотя давно доказана серийность выведения рабочих и у видов, зимующих c расплодом: Myrmica rubra (Brian, 1957), Solenopsis invicta (O’Neil, Markin, 1975; Lofgren et al., 1986), и у видов, зимующих без него: Formica picea (Длусский, 2001), группа Formica rufa (Захаров, 1972). Семья Formica s. str., в зависимости от климатических, топических и погодных условий и силы муравейника, может вывести за сезон 3–5 поколений расплода (Захаров, 1991; Захаров, Захаров, 2013). Сериальное выведение расплода для муравьев исходно, о чем свидетельствует четкая периодичность яйцекладок в зарождающихся семьях всех видов с самками-основательницами (Stamps, Vinson, 1991; Красильников, 1998; Саблин-Яворский, 1998; Длусский, 2001; и др.).
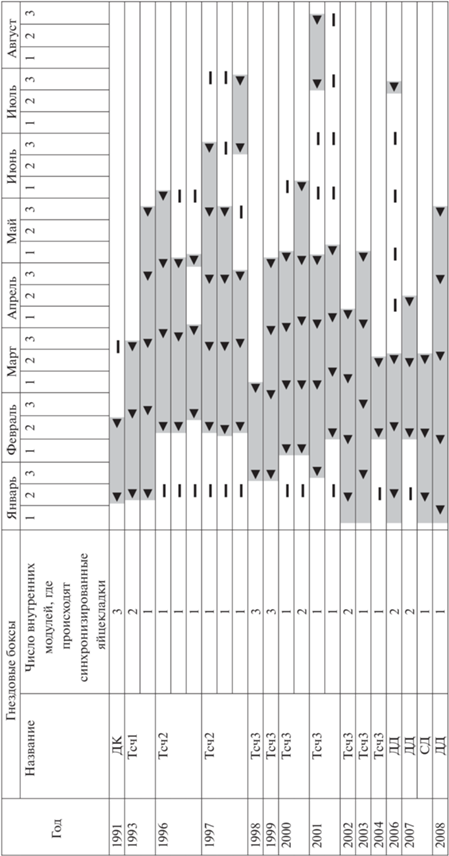
Эксперименты с лабораторными семьями выявили, что потенциальный диапазон сроков размножения у рыжих лесных муравьев значительно шире диапазона, реализуемого ими в природе – от 1-й декады января до 3-й декады августа (рис. 2), что соответствует девяти циклам яйцекладок. В различные годы 1-я яйцекладка начиналась в интервале 06.1–17.2, что соответствует самым ранним срокам весенней активизации муравейников в Европе (Gösswald, 1951; Дьяченко, 2017). Однако число кладок в непрерывной серии для семьи ни разу не превысило пяти. Максимальное число яиц в одной кладке в экспериментальной семье равнялось 2.0–2.5 nw, где n = 1–8 – число самок, w – число рабочих. Начиная с 3-й кладки, число отложенных яиц резко снижалось, а в малых группах падало до нуля. В новой серии эта тенденция повторялась. По-видимому, после нескольких последовательных яйцекладок самки нуждаются в длительном отдыхе (Захаров, Захаров, 2013). Муравьи в целом выдерживали примерно месячный интервал между яйцекладками. Развитие от яйца до имаго занимало около 40 дней.
Рис. 2.
Сроки яйцекладок в лабораторных семьях рыжих лесных муравьев. Черный треугольник – начало яйцекладки, прочерк – пропущенная яйцекладка; серая заливка – сроки одной серии яйцекладки. 1–3 – Декады месяца.
Брачный лёт крылатых особей происходит у рыжих лесных муравьев в конце мая–первой декаде июня (Длусский, 1967). Однако у двух видов этой группы, F. lugubris и F. polyctena, возможен и позднелетний лёт крылатых в конце июля – первой половине августа (Захаров, Захаров, 2013, 2014). В Сибири и Северной тайге F. lugubris реализует только второй вариант выведения крылатых (Дмитриенко, Петренко, 1976). Наличие у F. lugubris и F. polyctena двух сроков лёта крылатых делает их воспроизводство менее зависимым от зимних запасов пищи или длительных весенних холодов (Захаров, Захаров, 2013, 2014). У F. pratensis крылатые также могут вылетать два раза в год – весной и в конце лета (Otto, 1962; Schmidt, 1974).
Обеспечение семьи энергетическими ресурсами
Муравьи имеют значительное число “статей расхода” энергии: выкармливание расплода и уход за ним, обеспечение размножения, питание имаго, двигательная активность особей, социальные нужды (поддержание дорог и температурного режима гнезда, санитария, обмены), фуражировка (Бахем, Лампрехт, 1983; Coenen, 1988). Источником потребляемой энергии служит получаемая муравьями-фуражирами белковая и углеводная пища. Но лишь часть ее сразу же используется семьей для покрытия своих текущих расходов энергии. Как и все муравьи, Formica создают значительные социальные пищевые запасы.
Летом муравьи запасают пищу в зобиках и жировых телах молодых рабочих, что служит запасом семьи на случай плохой погоды и т.п. (Мартин и др., 1988). На зиму рабочие Formica накапливают в жировом теле запасы пищи, основным источником которых служит сахар, получаемый муравьями из пади и перерабатываемый биохимическим путем в гликоген (Chapman, 1998). Наращивание этих запасов продолжается до глубокой осени, пока сохраняется возможность собирать падь (Kirchner, 1964; Берман и др., 2007), а их размеры пропорциональны численности семьи. Так, у Formica s. str. жировое тело зимующего рабочего весит порядка 4 мг (Kneitz, 1967), т.е. муравейник с миллионным населением может заготовить на зиму около 4 кг пищевых запасов. Этого достаточно, чтобы семья перезимовала, произвела весенний разогрев купола и вырастила первое поколение потомства в следующем году.
Сезонный профиль активности фуражировки имеет у данной группы муравьев свою специфику: охотничья – один пик в летние месяцы, в период выкармливания расплода, сбор пади нарастает к середине лета и держится на высоком уровне до завершения процесса создания зимних запасов семьи (Плешанов, 1982; Кауль, 1984; Дьяченко, 2001).
Гнездовая деятельность. Сооружение и поддержание жилищ
Гнездо – наиболее значимый и заметный результат совместной деятельности рыжих лесных муравьев как “инженерных” видов. Наряду с пищевым потоком, оно выполняет важнейшую функцию консолидации семьи. Большинство событий в жизни семьи муравьев так или иначе проявляется в соответствующих подвижках в конструкции, материалах или состоянии гнезда, смысл которых заключается в обеспечении оптимальных условий обитания при цикличной смене ключевых задач. Терморегуляция сопряжена с постоянным контролем вентиляции и гнездовых выходов и, при необходимости, с трансформацией купола и подземной части гнезда (Захаров, Захаров, 2014; Горюнов, 2015; Дьяченко, 2017). Надо расширить внутренний конус – с кормового участка в гнездо идет поток крупных палочек, обновить покровный слой – возрастает поток хвои, укрепить купол и защитить его от плесени – увеличивается поступление смолы. Сезонные изменения в устройстве гнезд рыжих лесных муравьев изучались неоднократно (Wiśniewski, 1965; Малоземова, 1974; Малоземова, Овсяникова, 1987; Gösswald, 1989; и др.). В нормальных условиях строительная активность имеет два пика – поздней весной и осенью (Грауберг, 1988).
При этом всегда выражена тесная связь между организацией семьи и конструкцией гнезда. Ключевой приоритетной задачей муравьев на весь срок выращивания потомства (т.е. в течение весны и лета) является поддержание в гнезде температурного режима, оптимального для развития расплода.
Завершается сезон подготовкой муравейника к зиме. Купол частично обновляется, перед холодной зимой муравьи сооружают более мощным покровный слой гнезда. В подземной части гнезда муравьи сооружают разветвленную сеть зимовочных ходов и камер, что хорошо видно по свежим выбросам почвы, покрывающим осенью гнездовой вал и окаймляющим купол кольцом шириной до 0.5–1.0 м. Сооружение зимовочного гнезда начинается во второй половине августа (после выведения последних рабочих) и продолжается до осенних холодов.
Демографическая деятельность. Управление демографическим ресурсом семьи
Демографический ресурс семьи – это основной ресурс, обеспечивающий реализацию каждой из форм социальных структур, а также дееспособность муравейника во времени и при различных внешних воздействиях. Он характеризуется численностью и функциональной структурой населения, определяющими рабочий и репродуктивный потенциалы социума, – доступные уровни активности в гнезде и на участке, продукцию рабочих и репродуктивных особей. Поэтому именно акцент на демографический ресурс семьи позволяет связать в общую систему различные аспекты жизни социума муравьев и выработать его целостное восприятие (Захаров, 2015).
Ресурс, обеспечивающий рабочий потенциал социума, складывается из функционально активных, т.е. уже прошедших социализацию и входящих в состав функциональных групп особей (няньки, хранители запасов, строители, охотники, сборщики пади), и резерва рабочих, состоящего в основном из молодых особей, назовем его оперативным резервом социума. Наличие такого резерва позволяет муравейнику успешно решать задачи восполнения естественной смертности и текущих потерь фуражиров на кормовом участке, компенсировать крупные разовые потери и формировать дополнительные контингенты рабочих в экстренных ситуациях (появление массовой добычи, территориальные войны, тотальное переселение и т.п.). Размеры и специфика формирования и использования оперативного резерва относятся к наименее изученным разделам мирмекологии, хотя именно наличие этого ресурса позволяет социуму эффективно потреблять энергетические и географические ресурсы среды, развиваться и быть конкурентоспособным и устойчивым к негативным воздействиям.
Меняются выполняемые задачи, соответственно меняется состав или соотношение функциональных групп, – демографическая структура населения семьи. Сезонный характер распределения рабочих особей по функциональным группам позволяет муравьям более эффективно использовать имеющийся в гнезде демографический ресурс, что совокупно с серийным выведением расплода и формированием резерва функциональных особей создает надежную основу для устойчивого существования социума в ряду лет.
Социоструктурная деятельность. Сезонные изменения социальных структур
Здесь объединены регулярные сезонные трансформации структур, в т.ч. системы гнезд и внутрисемейных структур, кормовых участков, охраняемых территорий и многовидовых ассоциаций муравейников. Эти события также вписываются в годичный цикл жизни муравейников (Дьяченко, 1999, 2017; Сейма, 2008; Захаров, Захаров, 2013). Подробно проявления сезонной цикличности прослежены на разных представителях Formica – Formica s. str. (Захаров, 2015), Coptoformica (Pisarski, 1982; Chudzicka, 1982; Горюнов, 2003) и Serviformica (Rosengren, 1969, 1985; Бургов, 2015). Исследования охватывают не только системы жилых муравейников, но и вспомогательные гнезда на территории (например, Anderson, McShea, 2001).
Фенология жизни надсемейных структур. У видов, использующих наземные дороги в комплексе одиночных гнезд, группе колоний и в федерации, все межсемейные связи на зиму полностью прекращаются и каждой весной восстанавливаются заново. Это входит в число основных задач муравейника на этапе весеннего перестроения (период 1, переходный период весна–лето). При этом действует четкое правило: доминирующие (материнские) муравейники активизируются раньше вторичных гнезд и, как правило, становятся инициаторами активизации последних (Захаров, 1991, 2015). Туда приходят носильщики из гнезд-доминантов и налаживают обменные связи. Показательно, что в норме весной восстанавливается та же самая структура, которая функционировала минувшей осенью, что свидетельствует о хорошей структурной памяти у Formica. Система связей может быть утрачена при значительных нарушениях инфраструктуры поселения в результате гибели части крупных муравейников. И сам факт восстановления прежней схемы связей уже указывает на сохранение данной группой гнезд своей дееспособности после зимовки (Otto, 1962; Дьяченко, 2017). Успешность акции зависит от сроков восстановления связей между конкретными гнездами: связи, не установленные до конца мая, могут быть утрачены вообще.
Особое место в миграционных процессах в комплексах гнезд занимают временные объединения муравейников, возникающие после крупных поломок (Фоменко, 1991; Сейма, 2008; и др.). Основные факторы нарушения годичного цикла муравейника – зоогенные и климатические. Зоогенные факторы – это поломки гнезд позвоночными животными и браконьерами, связанные с разрушением гнезда и гибелью большей части его населения. Так, медведи обычно разоряют до 40–60% крупных муравейников, а при высокой численности до 90% гнезд (Гримальский, 1975; Mordosov, 2002; Захаров, 2008; Захаров, Рыков, 2018; Огурцов, Федосеева, 2018).
Фенологические аспекты жизни многовидовых ассоциаций муравейников. Взаимодействие муравейников разных видов, входящих в одну многовидовую ассоциацию муравейников, также имеет свою фенологическую специфику. Так, к числу общих характеристик многовидовых ассоциаций в умеренной зоне относятся сроки весенней активизации муравейников. Первыми активизируются семьи видов-доминантов (Горюнов, 2003; Захаров, 2015; Бургов, 2015), которые таким образом получают ситуационное преимущество перед субдоминантами. Подобная очередность, вероятно, более значима в неполных ассоциациях (без облигатного доминанта), где опережающая весенняя активизация одного из субдоминантов может обеспечить ему выход на позиции доминанта.
КОНВЕЙЕР ПРИОРИТЕТНЫХ ЗАДАЧ И ПРЕЕМСТВЕННОСТЬ ПЕРИОДОВ
Основные задачи социума муравьев – самосохранение в годичном цикле и расширенное воспроизводство населения, обеспечивающее развитие социума в многолетней перспективе. В отлаженной структуре годичного цикла муравейника согласованно решаются две важнейшие для существования социума задачи: адаптации к климатическим реалиям региона и погодным условиям года и рациональное использование имеющегося демографического ресурса. В каждом периоде у муравьев имеются свои задачи, являющиеся ключевыми для данного отрезка годичного цикла (например, разогрев гнезда и начало фуражировки весной или подготовка зимовочного гнезда и создание запасов питательных веществ на зиму осенью).
Цикл складывается из последовательных периодов, образующих “конвейер приоритетных дел”. В течение одного года в жизни муравейника сменяют друг друга качественно различающиеся периоды – пулы событий, имеющих определенную временну́ю привязку (табл. 1). Их содержание и значимость изменяются по периодам, но вместе они и образуют “конвейер приоритетных дел”: весенний разогрев гнезда и выведение первого расплода; организация вне гнезда системы снабжения семьи ресурсами; поддержание в гнезде оптимального терморежима для расплода; выращивание пополнения семьи; подготовка гнезда к зиме и создание зимних запасов; зимовка. Периоды годичного цикла облигатно сменяют друг друга, и на каждом предшествующем этапе формируются предпосылки и ресурсы для выполнения последующего.
Переходный период зима–весна. На этом временном отрезке происходит активизация семьи, ее выход из состояния зимовки. Муравьи активизируются и формируют в верхней трети купола “тепловое ядро”, в котором самки начинают первую в сезоне яйцекладку. Рабочие муравьи интенсивно расходуют резервные вещества на разогрев гнезда и выкармливание расплода, пополняя ряды активных функциональных групп (охотников, сборщиков пади, строителей, нянек), которые начинают ремонт гнезда, выращивание расплода и восстановление структуры участка.
Основной период 1 – весенний. Происходит переход семьи на внешние источники энергоресурсов, выведение репродуктивных особей (самцов и самок), а также мощное подкрепление семей выведшимися в те же сроки рабочими. Приоритетными задачами являются переход на внешние источники питания и формирование контингентов функциональных групп (нянек, фуражиров, строителей и охраны купола и т.д.), характерных для семьи рыжих лесных муравьев и действующих в течение всего вегетационного периода. Иначе все остальные задачи данного периода годичного цикла (выращивание первого поколения потомства, разворачивание структуры кормового участка и системы межсемейных связей, расселение) останутся невыполненными.
За периодом 1 следуют три действия, составляющие суть переходного периода весна–лето: а) ротация яйцекладущих самок и наращивание численности рабочих; б) полное развертывание социальных и территориальных структур, освоенных муравейником; в) выделение естественных отводков взрослыми активными семьями.
а) Ротация яйцекладущих самок и рабочих в действующих муравейниках способствует резкому увеличению продуктивности семей и обеспечивает максимум их летнего пополнения рабочими особями (Forelius et al., 1993). Важно, что схема наращивания плодовитости молодых самок связана со способом основания новых семей: у видов с независимым образованием семей плодовитость изначально низка и впоследствии постепенно увеличивается в течение длительного времени. А у полигинных видов с регулярной ротацией яйцекладущих самок плодовитость молодых самок достигает своего максимума уже в ближайшие недели после приема их в гнездо (Passera, Keller, 1990; Zakharov, 2011). С учетом отмеченной выше потребности яйцекладущих самок в отдыхе после каждой серии из 3–4 яйцекладок (рис. 2; Захаров, Захаров, 2013) примечательна “своевременность” ротации самок – именно к началу периода, когда плодовитость “старых” самок уже пошла на спад.
б) Полное развертывание социальных и территориальных структур. Завершение формирования контингента внегнездовых рабочих и мощное пополнение состава рабочих в конце мая позволяют семье занять максимально возможную охраняемую территорию и реализовать на ней полное развертывание структур (дорог, вспомогательных гнезд, убежищ).
в) Выделение отводков. Одновременные пополнение семьи молодыми самками и рабочими, а также высвобождение после вылета крылатых и выведения расплода большого числа ухаживавших за ними взрослых муравьев повышает гетерогенность и “пассионарность” системы и создает условия для выделения естественных отводков.
В целом за переходный период весна–лето муравейник создает предпосылки для достижения на следующем этапе годичного цикла максимальных продуктивности, мощности и структурной сложности системы.
Основной период 2 – летний. В норме складывается из трех серий выведения многочисленного расплода, обеспечивающих рост и развитие самого муравейника, а также формирование им поликалических и надсемейных структур. Мощный демографический ресурс, накапливаемый за этот период, позволяет муравьям успешно решать стратегические этапные задачи: а) пока есть расплод, в гнезде должно быть тепло, б) постоянно необходимо достаточно много рабочих каждой функциональной группы – для обогрева, ухода, кормления и защиты, в) муравейник должен утвердиться в системе межгнездовых и межвидовых отношений. И все это при регулярно или случайно меняющихся условиях. Так, потребность семьи в пище в течение всего периода закономерно меняется, – она возрастает при наличии и росте в гнезде личинок, спадая на фазе куколки и откладки яиц (Голосова, Захаров, 1974).
Выведение каждого поколения рабочих – это тоже определенная структуризация жизни семьи. Однако признать их периодами нельзя по ряду соображений. Во-первых, число таких поколений непостоянно (от 3 до 5 в одном гнезде) в зависимости от погодных условий и иных обстоятельств. Во-вторых, состав продукции также непостоянен, – большинство муравейников выращивают самок далеко не каждый год (Otto, 1962; Длусский, 1967; Gösswald, 1989). В обоих случаях сбои не препятствуют началу следующего периода годичного цикла, который начинается в установленные фенологические сроки. Поэтому мы можем рассматривать серийное выведение молоди и лабильность сроков выращивания репродуктивных особей у рыжих лесных муравьев как вторичные явления. Каждой яйцекладке предшествует подъем самок в верхние (прогреваемые) камеры гнезда, а после откладки яиц самки возвращаются в его нижнюю прохладную часть. Подъем самок всегда начинается за 2–4 дня до начала яйцекладки.
Переходный период лето–осень. После завершения выращивания расплода в задачах, стоящих перед семьей, происходят коренные изменения: а) падают до минимума потребность в белковой пище и связанная с ней общая активность на кормовом участке; б) исчезает потребность в разогреве гнезда, в) семья должна перестроиться для подготовки к зимовке: начать сооружение (обновление) зимовочного гнезда, переключиться на углеводное питание и формирование зимнего состава рабочих.
Основной период 3 – осенний. Приоритетные задачи периода: а) подготовка зимовочного гнезда и создание зимних запасов пищи; б) сворачивание внегнездовой активности и переход на питание из пищевых запасов семьи; в) формирование зимней функциональной структуры с абсолютным преобладанием муравьев – хранителей пищевых запасов, далее – просто хранители (supply keepers, keepers), и небольшим числом активных (мобильных) муравьев (mobile workers) и свиты самок (suite for queens). Доля участия хранителей запасов в составе рабочих увеличивается до 90–95%. Детали процесса осеннего перестроения функционального состава рабочих муравьев рассматриваются в следующем разделе статьи.
Переходный период осень–зима связан с распределением муравьев по зимовочным камерам и фактической “инкапсуляцией” семьи. Происходит консервация наземной части жилого гнезда. Консервация различных вспомогательных гнезд происходит раньше – по мере исключения их из режима активного использования.
Основной период 4 – зимний. За продолжительным, около 2 мес. (середина августа–середина октября), периодом накопления энергетических запасов семьи следует еще более длительный период зимовки (ноябрь–март, 5 мес.), в течение которого расход запасов невелик по причине низкой активности муравьев и соответственно минимальных трат ими пищевых ресурсов.
ИЗМЕНЕНИЕ ФУНКЦИОНАЛЬНОГО СОСТАВА РАБОЧИХ В ГОДИЧНОМ ЦИКЛЕ. СЕЗОННЫЙ ПОЛИЭТИЗМ
Смена задач в цикле, безусловно, должна обеспечиваться необходимыми для ее выполнения демографическими ресурсами. Как мы уже рассмотрели выше, в годичном цикле социума происходит закономерная смена общих приоритетных задач, комплексных по сути и требующих для своего решения участия больших масс муравьев (табл. 1). Для многих особей это связано со сменой выполняемых ими функций. Такое переключение функций рабочих муравьев в годичном цикле носит регулярный характер, хотя при быстротечной жизни большинства рабочих особей напрямую это касается лишь части из них. Тем не менее функции многих рабочих закономерно меняются не только с возрастом особи (возрастной полиэтизм) или в соответствии с размерными и психофизиологическими предпочтениями особей (постоянный, кастовый полиэтизм), но также в соответствии с сезонными приоритетными задачами, стоящими перед социумом (предварительно можно определить как сезонный полиэтизм). Меняются задачи, меняется и демографическая структура рабочего населения муравейника. Мобильность сезонного распределения рабочих по функциональным группам позволяет муравьям более эффективно использовать имеющийся в гнезде контингент рабочих особей.
На уровне семьи такая проблема должна быть решена системным путем: в социуме имеется постоянный контингент неспециализированных особей (полиэтическая группа “пассивные фуражиры”) либо формируется многочисленный резерв семьи из потенциально активных особей (Захаров, 1991, 2015). Первый вариант в полной мере реализуется у видов с системой массовых мобилизаций (r-стратегией фуражировки), второй – у К-стратегов с закрепленными функциями пассивных фуражиров.
Массовое переключение населения муравейника на выполнение очередной суперзадачи не означает даже временного исчезновения других внутригнездовых (свита самок), нагнездовых (муравьи-наблюдатели, строители) и внегнездовых (разведчики, охотники, сборщики пади) функциональных групп, являющихся системообразующими и обеспечивающими “самобытность” социума. Речь идет не о полном замещении одних функциональных групп другими, а о перераспределении больших масс рабочих между ними, чтобы в течение каждого периода обеспечить выполнение ключевых для него функций.
Серийность выведения молоди облегчает смену занятости для масс рабочих. Она позволяет начать формирование ресурсов для реализации стратегических задач семьи заблаговременно, задолго до собственно (самого) события. В гнезде одновременно присутствуют рабочие нескольких возрастов, которые обеспечивают плавную смену ведущих контингентов. Так, в конце мая–начале июня из раннелетнего поколения рабочих формируется контингент хранителей запасов, которым после зимовки суждено стать основой потоков фуражиров (Маавара, Мартин, 1983; Мартин, 1991). А в последнем, августовском поколении рабочих, составляющем около 0.3 численности населения муравейника (Калинин, 1998), почти все особи становятся хранителями запасов (Захаров, Захаров, 2014). Благодаря этому все взрослое население муравейника сохраняет дееспособность в течение длительного времени: пока заполняются жировые тела всех новых молодых рабочих, более старшие особи могут готовить в зимовке гнездо и собирать падь. Это крайне важно – при любых перестроениях семья должна постоянно сохранять свою функциональность.
На осеннем этапе абсолютное большинство населения (90–95%) гнезда должно стать хранителями (Kirchner, 1964; Kneitz, 1967). Наличие жировых тел лишь у 40–50% внутригнездовых рабочих не позволяет муравьям вырастить весной первое поколение расплода (Захаров, Захаров, 2014). Если бы не последнее поколение рабочих, то уже в конце августа хранителями вынуждены были бы стать взрослые активные рабочие, и семье не хватило бы сил для подготовки к зиме.
Деактивация семьи, сворачивание летней функциональной структуры происходит через формирование осеннего варианта – с преобладанием сборщиков пади и строителей. Далее идет формирование уже зимней функциональной структуры с абсолютным преобладанием хранителей пищевых запасов. Последовательность формирования зимнего состава и энергетического запаса семьи муравьев группы Formica rufa приведена на рис. 3. В первую очередь, к летним хранителям запаса семьи присоединяется последнее поколение рабочих, затем другие летние внутригнездовые рабочие. Охотники переключаются на сбор пади и строительного материала. Последними начинают накапливать запасы строители и сборщики пади, работающие на корневых тлях до предзимья.
Рис. 3.
Схема формирования зимнего состава и энергетического запаса семьи рыжих лесных муравьев. Горизонтальные сплошные стрелки означают переход из летних функциональных групп в зимние группы хранителей запасов и свиты самок; косые пунктирные линии – уход элитной части каждой из этих групп в функциональную группу мобильных рабочих; горизонтальная пунктирная линия – формирование из α-особей группы мобильных рабочих.
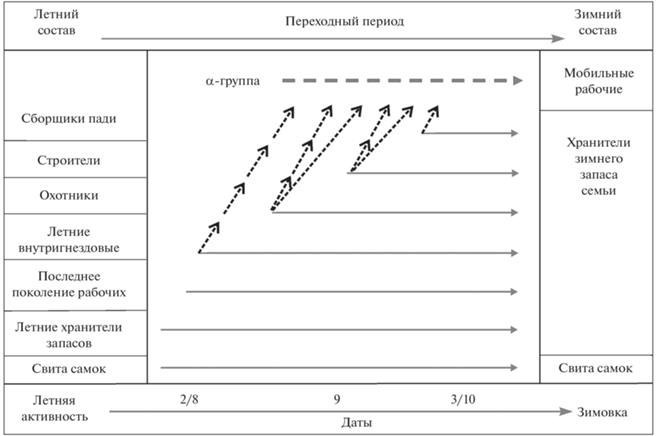
Возможна лишь ориентировочная оценка участия в формировании зимнего состава хранителей муравьев разных летних функциональных групп: 20–27% – это летние хранители и няньки, 23–30% – последнее поколение рабочих целиком, 15–25% – оперативный резерв семьи, который сначала усиливает группу сборщиков пади, а уже оттуда попадает в хранители, 15–22% – из строителей и охотников невысоких рангов. Абсолютное большинство рабочих, но не все, становятся при этом малоподвижными. Наиболее опытные особи (α-рабочие) из каждой функциональной группы жировым телом не обзаводятся и остаются активными, образуя на этот период специфическую функциональную группу мобильных рабочих. Мобильные рабочие рассредоточены по всем частям гнезда, активно перемещаются по тоннелям и камерам и в январе (Сейма, 1998), а весной обеспечивают общую скоординированную активизацию семьи.
Уход насекомых на зимовку происходит медленно. Это связано с постепенным падением осенних температур, а также с тем, что не все особи успевают быстро обеспечить себя резервными веществами, необходимыми для зимовки. Выход из зимовки, напротив, совершается очень быстро (Воронцов, 1963, 1982). Эти тенденции соблюдаются и у таких социальных насекомых, как муравьи. И здесь противоположно направленные процессы отражают общую стратегию, направленную на минимизацию сроков пребывания одиночного насекомого или семьи муравьев в беззащитном состоянии. Осенью растянутый, медленный уход муравьев на зимовку сохраняет хотя бы относительную мобильность семьи (т.е. ее возможность защиты и ремонта гнезда) поздней осенью и в предзимье. А весной решение этих же задач требует максимально быстрой активизации семьи. Иначе семья станет легкой добычей более мобильных соседей по сообществу. Эти задачи хорошо согласуются и с другими проблемами, которые приходится решать муравьям в периоды подготовки зимовки и выхода из нее.
Весенний расход энергетических запасов семьи должен происходить стремительно, поскольку меньше чем за месяц необходимо произвести полное перестроение сферы обеспечения с зимнего функционального состава на летний, т.е. реализовать приведенные на рис. 3 события в обратном порядке и сформировать все летние функциональные группы рабочих. Для этого необходима быстрая утилизация энергетических ресурсов семьи, чему способствует параллельное выполнение муравьями двух больших энергоемких задач: разогрева гнезда и выкармливания первого поколения расплода. Особенно жестко связано с освобождением рабочих особей от жировых тел сопровождение задач сферы обеспечения (Хансен, Вийк, 1981), ведь эффективное функционирование в качестве фуражиров и строителей просто недоступно физически особям с развитыми жировыми телами. Для этого они должны освободиться от своих жировых тел. Поэтому в апреле участие хранителей в составе семьи должно снизиться с зимних 95 до летних 25–30%.
ОБЩЕЕ ОБСУЖДЕНИЕ
Данное исследование во многом носит постановочный характер и имеет целью, прежде всего, привлечь внимание исследователей к необходимости соблюдения в мирмекологических и сопряженных с ними зоологических и синэкологических работах требований, связанных с явлениями цикличности и сезонности. Эти вопросы приобретают особое значение при многолетних исследованиях, будь то длительные эксперименты или стационарные мониторинговые наблюдения, поскольку несоблюдение фенологических рамок ведет к утрате ценной информации и делает выполняемые разными авторами исследования несопоставимыми.
При этом имеются серьезные трудности адекватного описания связей между годичным циклом и жизненным циклом семьи (colony life history) у эусоциальных насекомых (Stearns, 1992, 2000; Starr, 2006). Феномен сезонной цикличности у этих насекомых имеет свою специфику (Feller, 1989; Heinze, 1993; Heinze, Hölldobler, 1994; Starr, 2006; Захаров, Захаров, 2013, 2019). Так, можно полагать, что годичный цикл жизни является предельным временны́м интервалом корректного описания отдельного социума насекомых (например, отдельного муравейника) в природных условиях. При переходе к многолетним описаниям сообществ муравьев необходимо переходить на уровень поселения (комплекса гнезд конспецификов или многовидовых сообществ), поскольку миграционные потоки между составляющими поселение муравейниками, как правило, не позволяют, по крайней мере, на современном технологическом уровне исследований, прослеживать судьбы отдельных семей в потоке многолетних событий. Поэтому, хотя бы в приложении к Formica s. str., годичный цикл жизни муравейника является в определенном смысле идеализаций, как если бы модельный муравейник в течение своей жизни не менял своего статуса и не участвовал в общих структурных перестроениях, систематически происходивших в комплексе гнезд, частью которого он является. Данное обстоятельство не меняет общих принципов организации годичных циклов жизни поселения муравьев, но не позволяет выделять реальные границы родственных группировок различного масштаба (семей, колоний, федераций).
Следует также отметить принципиальное отличие применяемого нами понятия “годичный цикл жизни” или эквивалентного ему “годичный цикл активности” (Захаров, Захаров, 2013; Захаров, 2015) от используемого некоторыми авторами понятия “годичный цикл развития” (Кипятков, Лопатина, 2007). Если “годичный цикл жизни” охватывает все стороны жизни муравейника, связанные с гнездом, кормовым участком, выращиванием потомства, социотомией и т.д., то в рамках “годичного цикла развития” предмет рассмотрения ограничивается только схемами выращивания расплода, что делает всю процедуру уплощенной и односторонней. Эти различия особенно существенны применительно к модельным объектам нашего исследования, которые зимуют без расплода. Размножение у них носит сугубо сезонный характер, и уже поэтому реальная жизнь социума рыжих лесных муравьев оказывается существенно сложнее уплощенных схем “годичного цикла развития”.
Имеется качественная разница в использовании в годичном цикле демографического ресурса муравьями, зимующими с расплодом (личинками 2–3-го возраста) и без такового. У первых во время весенней реактивации личинки служат и энергетическим ресурсом, и объектами выращивания. У вторых весенний этап энергетически почти целиком обеспечивается созданными осенью запасами в жировых телах рабочих, что делает их менее зависимыми от обеспеченности внешними источниками пищи и от погодных условий в период весенней активизации. Однако при этом для них становится необходимой радикальная смена функций большинством рабочих особей в годичном цикле жизни семьи, что мы и обсуждали в разделе “Изменение функционального состава рабочих в годичном цикле. Сезонный полиэтизм”. Как и все Formicini, рыжие лесные муравьи выращивают расплод в течение одного сезона – важное отличие от видов, которые зимуют с расплодом и для которых прямая забота о расплоде является одной из важных составляющих всех периодов годичного цикла семьи. В средней полосе европейской части России в норме расплод находится в гнезде рыжих лесных муравьев 4.0–4.5 мес. (с апреля по август). Это означает, что около трети всего времени в годичном цикле и двух третей срока общей внегнездовой активности уход за молодью не входит в число задач, выполняемых семьей этих муравьев.
Выращивание расплода в течение одного сезона сопряжено у муравьев с повышенной скоростью развития расплода. Например, установлено, что у Formica и Cataglyphis скорость индивидуального развития значительно выше, чем у большинства видов с зимующими личинками. Среди северных муравьев расплод Formica отличается самым быстрым ростом и развивается почти вдвое быстрее расплода у зимующих с расплодом Myrmica и Leptothorax: развитие от яйца до куколки у Formica при 25–26°С происходит всего за 20–25 дней, а у M. rubra (Linnaeus 1758) при тех же температурах – за 34–35 дней, а при обычных для этого вида температурах около 22°С – за 40–45 дней, т.е. почти в 2 раза дольше (Кипятков, Лопатина, 2007). В итоге в средней полосе даже небольшая семья Serviformica может вырастить за лето 2–3 крупные партии расплода (Длусский, 2001; Кипятков, Лопатина, 2007), тогда как расплод Myrmica должен уходить на зимовку и завершает свое развитие только в следующем сезоне.
Одним из центральным вопросов, связанных с жизнеописанием социумов насекомых, является гипотеза о разделении этапов накопления ресурсов для выращивания репродуктивных особей и самого выращивания последних (Starr, 2006). Подобное разделение функций этапов саморазвития социума насекомых достаточно четко прослеживается у ресоциальных насекомых, таких как осы Dolichovespula (Hymenoptera, Vespidae) или шмели (Hymenoptera, Apidae) (Захаров, 1993; Радченко, Песенко, 1994), но применительно к муравьям с их поступательным развитием возникшего муравейника подобный подход вряд ли оправдан. Событийная насыщенность и вариантность развития в годичном цикле социума рыжих лесных муравьев – веский довод в пользу этого.
Наступление каждого из периодов достаточно надежно диагностируется по внешним признакам (табл. 2). Изменения в конструкции гнезда и поведении его обитателей являются не только следствием феноменологических особенностей любого из периодов, но и определяется качественными различиями в формах активности муравьев (табл. 3). В частности, поэтому незавершенность одного из периодов может стать критической для реализации всего годичного цикла и дальнейшей судьбы муравейника. В норме муравьи не приступят к выполнению следующего этапа годичного цикла, пока не завершен предыдущий. Так, не закончив процесса выращивания расплода рабочих, муравьи не могут начать подготовку гнезда к зимовке, поскольку им приходится все еще поддерживать в гнезде температурный режим. Хотя, следует отметить, если в гнезде остались уже только последние немногочисленные куколки рабочих, фуражиры могут начать сворачивать свою охотничью деятельность и переключатся на усиленный сбор пади и хвои.
Таблица 2.
Внешние признаки, индицирующие определенные этапы годичного цикла в гнезде рыжих лесных муравьев
| Период | Формы деятельности | |||||
|---|---|---|---|---|---|---|
| основной | переходный | репродуктивная | энергетическое обеспечение | гнездовая | демографическая | социоструктурная |
| Зимний–весенний | На куполе разогрев яйцекладущих самок, начало яйцекладки в “тепловом ядре” | Потоков фуражиров еще нет | Начало ремонта гнезда | На куполе множество муравьев-теплоносцев, появление первых “отрядов” фуражиров | На куполе переносы муравьев-имаго | |
| 1. Весенний | Переносы куколок рабочих по куполу, пустые коконы вокруг гнезда, крылатые на куполе | Кормовые дороги, фуражиры на кормовом участке – несут добычу, падь, сок растений | Ремонт повреждений и сушка гнезда, активный сбор смолы и других стройматериалов | На территории действуют массы фуражиров, в верхней части купола исчезают хранители запасов | Формирование кормовых и обменных дорог, вспомогательных гнезд, распределение колоний тлей | |
| Весенний–летний | Лёт крылатых, бескрылые самки около гнезд | Фуражиры во всех ярусах лесного сообщества | Ликвидированы все следы зимних и ранневесенних повреждений купола | В хорошую погоду муравейник “кипит” | Установление границ и нейтральных зон, иногда – муравьиные войны | |
| 2. Летний | Вынос пустых коконов рабочих, факультативно-июльское поколение крылатых | Обилие добычи, появление колоний корневых тлей | Интенсивное обновление покровного слоя и внутреннего конуса, устойчивый t°-режим | Нарастает агрессивность рабочих на куполе и на территории. Максимум охотничьей активности | Образование новых дорог, строительство новых (летних) отводков | |
| Летний–осенний | Пустые коконы у самого основания купола | Уменьшение доли белковой добычи на дорогах, усиление потоков сборщиков пади | Гнездо холодное. Кайма выбросов почвы вокруг купола, активный принос хвои и смолы | Формирование осеннего состава фуражиров с преобладанием сборщиков пади | Сокращение дорог 2-го и 3-го порядков, закрытие вспомогательных гнезд | |
| 3. Осенний | В гнездах нет расплода, самки спустились в подземную часть гнезда | Полное преобладание сборщиков пади среди фуражиров |
Кайма окружает купол гнезда, почвой и хвоей укрепляется покровный слой, почти все входы закрыты | В куполе преобладают рабочие с развитыми жировыми телами | Выделение (осенних) отводков и гнезд выселения на следующий год, угасание связей между гнездами | |
| Осенний–зимний | Расплода нет | Активны лишь колонии корневых тлей непосредственно у гнезда | Муравьи сконцентрированы в зимовочном гнезде | Полное преобладание рабочих с развитыми жировыми телами | Семьи инкапсулированы в зимовочных гнездах | |
| 4. Зимний | Расплода нет | Питание за счет внутренних ресурсов | Гнездо закрыто | Минимальная активность особей в зимовочных камерах | Колонны зимуют в соответствующих секторах зимовочного гнезда | |
Таблица 3.
Распределение состояний и активность по формам деятельности в годичном цикле жизни социума рыжих лесных муравьев
| Форма деятельности | ОР | Состояние по основным периодам годичного цикла | |||
|---|---|---|---|---|---|
| Зимний | Весенний | Летний | Осенний | ||
| а) Репродуктивная | 1 | Нет | Идет | Идет | Нет |
| б) Обеспечение энергетическими ресурсами | 1 | Нет | Начинается | Идет | Идет |
| в) Гнездовая | 1–2 | Нет | Идет | Идет | Идет |
| г) Демографическая (смена составов функциональных групп) | 2 | Зимний состав | Зимний состав → → Летний состав | Летний состав | Летний состав → → Зимний состав |
| д) Состояние социальных внегнездовых структур | 2 | Свернуты | Разворачиваются | Развернуты | Сворачиваются |
Сбой в годичном цикле по фазе активности означает отставание конкретного события в жизни социума от установленного для него времени, что чревато большими потерями для муравейника. Так, задержка лёта крылатых в одном из гнезд комплекса ухудшает его позиции при разделе территорий, а сдвиг сроков выведения из куколок последнего поколения рабочих рыжих лесных муравьев в осень означает острый дефицит рабочих особей в гнезде. Таким образом, успешное начало следующего сезона, включая выведение крылатых особей, во многом зависит от эффективности деятельности муравейника в предыдущем сезоне. Одновременно это означает потенциальную эквивалентность всех основных периодов годичного цикла.
Список литературы
Бахем И., Лампрехт И., 1983. Гнездо лесных муравьев Formica polyctena как модель экологической системы // Журнал общей биологии. Т. 44. № 1. С. 114–123.
Берман Д.И., Алфимов А.В., Жигульская З.А., Лейрих А.Н., 2007. Зимовка и холодоустойчивость муравьев на северо-востоке Азии. М.: Товарищество научных изданий КМК. 264 с.
Бургов Е.В., 2015. Многовидовая ассоциация муравейников: переход из неполной ассоциации в полную // Зоологический журнал. Т. 94. № 10. С. 1200–1210.
Воронцов А.И., 1963. Биологические основы защиты леса. М.: Высшая школа. 324 с.
Воронцов А.И., 1982. Лесная энтомология. М.: Высшая школа. 384 с.
Галахов Н.Н., 1959. Изучение структуры климатических сезонов года. М.: Изд. АН СССР. 182 с.
Голосова М.А., Захаров А.А., 1974. Эффективность действия муравьев Formica rufa на дубовую зеленую листовертку // Лесоведение. № 1. С. 37–42.
Горюнов Д.Н., 2003. Структура комплексов муравейников Formica exsecta и ее динамика в течение сезона // Успехи современной биологии. Т. 123. № 3. С. 249–256.
Горюнов Д.Н., 2015. Гнездостроение у муравьев Formica exsecta (Hymenoptera, Formicidae) // Зоологический журнал. Т. 94. № 10. С. 1132–1137.
Грауберг Р.Э., 1988. Состав и динамика сбора строительного материала северным лесным муравьем // Биологические основы использования полезных насекомых. М. С. 50–52.
Гримальский В.И., 1975. Влияние медведей на муравьев рода Formica в Березинском заповеднике // Муравьи и защита леса. Материалы V Всесоюзного мирмекологического симпозиума. М. С. 74–77.
Длусский Г.М., 1967. Муравьи рода Формика. М.: Наука. 236 с.
Длусский Г.М., 1980. Температурный режим в гнездах некоторых видов и пути эволюции терморегуляции у муравьев рода Formica // Физиология и популяционная экология животных. Саратов. Вып. 6/8. С. 13–36.
Длусский Г.М., 2001. Сезонная динамика развития семей Formica candida в изолированной болотной популяции // Муравьи и защита леса. Материалы XI Всероссийского мирмекологического симпозиума. Пермь. С. 69–71.
Дмитриенко В.К., Петренко Е.С., 1976. Муравьи таежных лесов Сибири. Новосибирск: Наука. 220 с.
Дьяченко Н.Г., 1999. Видовой состав муравьев рода Formica Беловежской Пущи и особенности их экологии // Parki Narodove I Rezerwaty Przyrody (Blałoweża). Т. 18. № 3. P. 81–90.
Дьяченко Н.Г., 2001. Суточные и сезонные биоритмы муравьев Formica s. str. в Беловежской пуще // Муравьи и защита леса. Материалы XI Всероссийского мирмекологического симпозиума. Пермь. С. 25–27.
Дьяченко Н.Г., 2017. Рыжие лесные муравьи Беловежской пущи. М.: Товарищество научных изданий КМК. 128 с.
Захаров А.А., 1972. Внутривидовые отношения у муравьев. М.: Наука. 216 с.
Захаров А.А.,1976. Использование метода исчерпывающих выборок при учете муравьев // Pedobiologia. Bd. 16. H. 8. S. 418–424.
Захаров А.А., 1991. Организация сообществ у муравьев. М.: Наука. 278 с.
Захаров А.А., 1993. Стадии развития эусоциальности у насекомых и критерии их выделения // Журнал общей биологии. Т. 54. № 5. С. 609–618.
Захаров А.А., 2015. Муравьи лесных сообществ. М.: Товарищество научных изданий КМК. 404 с.
Захаров А.А., Длусский Г.М., Горюнов Д.Н., Гилёв А.В., Зрянин В.А., Федосеева Е.Б., Гороховская Е.А., Радченко А.Г., 2013. Мониторинг муравьёв Формика. Информационно-методическое пособие. М.: Товарищество научных изданий КМК. 99 с.
Захаров А.А., Захаров Р.А., 2013. Влияние зоогенных и климатических факторов на годичный цикл жизни муравейника группы Formica rufa // Муравьи и защита леса. Материалы XIV Всероссийского мирмекологического симпозиума М.: Товарищество научных изданий КМК. С. 210–215.
Захаров А.А., Захаров Р.А., 2014. Муравьи в условиях экстремально жаркого лета // Зоологический журнал. Т. 93. № 1. С. 92–107.
Захаров А.А., Захаров Р.А., 2018. Фенологические аспекты мониторинга муравейников Formica s. str. // Проблемы экологического мониторинга и моделирования экосистем. Т. 29. № 4. С. 86–110.
Захаров А.А., Захаров Р.А., 2019. Муравьи и позвоночные животные. М.: Товарищество научных изданий КМК. 186 с.
Захаров А.А., Захаров Р.А., Федосеева Е.Б., 2015. Использование параметров гнезда рыжих лесных муравьев в мониторинге муравейников // Проблемы экологического мониторинга и моделирования экосистем. Т. 26. № 1. С. 68–90.
Захаров Р.А., 2008. Муравьи Formica (Formicidae) // Компоненты экосистем и биоразнообразие карстовых территорий европейского севера России (на примере заповедника “Пинежский”). Архангельск. С. 261–272.
Захаров Р.А., Захаров А.А., 2012. Мониторинг комплекса гнезд рыжих лесных муравьев “Красные Горы” // Многолетняя динамика экосистем природных комплексов Пинежского заповедника и сопредельных территорий. Архангельск. С. 78–85.
Захаров Р.А., Рыков А.М., 2018. Фотоловушки в мониторинге повреждений муравейников бурым медведем // Муравьи и защита леса. Материалы XV Всероссийского мирмекологического симпозиума. Екатеринбург. С. 157–160.
Калинин Д.А., 1998. Выплод рабочих у Formica aquilonia и метод его оценки // Муравьи и защита леса. Материалы X Всесоюз. мирмекологического симпозиума. М. С. 28–31.
Кауль Р.М., 1984. Суточная и сезонная динамика фуражировки у рыжих лесных муравьев // Насекомые в экосистемах лесной зоны Сибири. Томск. С. 41–44.
Кипятков В.Е., Лопатина Е.Б., 2007. Сезонные циклы и стратегии муравьев: структура, разнообразие и адаптивные особенности // Стратегия адаптаций наземных членистоногих к неблагоприятным условиям среды. СПб.: Изд-во СПб. ун-та. С. 107–192.
Красильников В.А., 1998. Экспериментальное изучение первичного плеометроза у дернового муравья Tetramorium caespitum (Formicidae) // Успехи современной биологии. Т. 118. Вып. 3. С. 323–330.
Лаврехин Ф.А., Паннова С.В., 1983. Биология медоносной пчелы. М.: Колос. 304 с.
Маавара В.Ю., Мартин А.-Й.А., 1983. Роль запасных особей в семье рыжих лесных муравьев // Механизмы поведения. Материалы III Всесоюз. конференция по поведению животных. М.: Наука. Т. 1. С. 102–104.
Малоземова Л.А., 1974. Закономерности динамики суточной и сезонной активности муравьев рода Formica // Рациональное использование биол. ресурсов Сибири. Красноярск. С. 185–189.
Малоземова Л.А., Овсянникова Н.П., 1987. Распределение и динамика “вентиляционных” отверстий на куполе муравейника // Муравьи и защита леса. Материалы VIII Всесоюз. мирмекологического симпозиума. Новосибирск. С. 122–124.
Мартин А.-Й.А., 1991. Поведение и значение запасных особей рыжих лесных муравьев // Муравьи и защита леса. Материалы IX Всесоюз. мирмекологического симпозиума. М. С. 98–101.
Мартин А.-Й.А., Мянд М.Ю., Маавара В.Ю., 1988. Влияние искусственного подогревания муравейника на жизнедеятельность семьи северного лесного муравья // Биологические основы использования полезных насекомых. М.: ИЭМЭЖ – ВООП. С. 84–86.
Огурцов С.С., Федосеева Е.Б., 2018. Муравьи в питании бурого медведя (Ursus arctos) в Центрально-Лесном заповеднике // Материалы XV Всероссийского мирмекологического симпозиума. Екатеринбург. С. 141–147.
Плешанов А.С., 1982. Насекомые – дефолианты лиственничных лесов Восточной Сибири. Новосибирск: Наука. 209 с.
Радченко А.Г. 2016. Муравьи (Hymenoptera, Formicidae) Украины. Киев: ИЗ НАНУ. 480 с.
Радченко В.Г., Песенко Ю.А., 1994. Биология пчел (Hymenoptera, Apoidae). СПб.: ЗИН РАН. 350 с.
Саблин-Яворский А.Д., 1998. Становление поселений Lasius niger в процессе роста и взаимодействия первичных микросемей // Муравьи и защита леса. Материалы X Всероссийского мирмекологического симпозиума. М. С. 20–22.
Cейма Ф.А., 1998. О внутригнездовых перемещениях рабочих Formica polyctena в зимний период // Муравьи и защита леса. Материалы X Всероссийского мирмекологического симпозиума. М. С. 117.
Cейма Ф.А., 2008. Структура населения муравьев тайги. Пермь: Пермский Государственный Университет. 166 с.
Соколов Л.В., 2010. Климат в жизни растений и животных. СПб.: ТЕССА. 344 с.
Соловьев А.Н., 2005. Биота и климат в ХХ столетии. Региональная фенология. М.: НПЦ “Пасьва”. 288 с.
Федченко И.А., 2008. Календарь природы // Компоненты экосистем и биоразнообразие карстовых территорий Европейского севера России. Гл. 5. Архангельск: ГПЗ “Пинежский”. С. 329–336.
Фоменко В.Ю., 1991. Влияние экзогенных факторов на процесс расселения в комплексе северного лесного муравья // Муравьи и защита леса. Материалы IX Всероссийского мирмекологического симпозиума. М. С. 69–71.
Хансен Т.Э., Вийк М.О., 1981. Сезонные изменения содержания резервных и холодозащитных веществ у Formica aquilonia (Hymenoptera, Formicidae) // Зоологический журнал. Т. 60. Вып. 3. С. 380–387.
Шульц Г.И., 1981. Общая фенология. Л.: Наука. 188 с.
Anderson C., McShea D.W., 2001. Intermediate-level parts in insect societies: adaptive structures that ants build away from the nest // Insectes Soc. V. 48. P. 291–301.
Brian M.V., 1957. Serial organization of brood in Myrmica // Insectes Soc. V. 4. № 3. P. 178–190.
Chapman R.F., 1998. The insects: structure and function. 4th ed. Cambridge un. press. 770 p.
Chudzicka E., 1982. Development de colonies de Formica (Coptoformica) exsecta Nyl. // Memorabilia Zoologica. V. 38. P. 205–237.
Coenen S.D., 1988. Zum Wȁrmehaushalt im Nest der Roten Wadlameise Formica polyctena (Hymenoptera, Formicidae) // Mitteilungen der Deutschen Gesellschaft für Allgemeine und Angewandte Entomologie. Bd. 6. № 1–3. S. 140–145.
Feller J.H., 1989. Daily and seasonal activity in woodland ants // Oecologia. V. 78. P. 69–76.
Fortelius W., Rosengren R., Cherix D., Chautems D., 1993. Queen recruitment in a highly polygynous supercolony of Formica lugubris (Hymenoptera, Formicidae) // Oikos. V. 67. P. 193–199.
Gösswald K., 1951. Die rote Waldameisen im Dünste der Waldhygiene. Lüneburg: Metta Kinau Verl. 160 S.
Gösswald K., 1989. Die Waldameise. V.1: Biologische Grudlagen, Ökologie und Verhalten. Wiesbaden. 660 S.
Heinze J., 1993. Life histories of subarctic ants // Arctic. V. 46. № 4. P. 354–358.
Heinze J., Hölldobler B., 1994. Ants in the cold // Memorabilia Zoologica. V. 48. P. 99–108.
Kirchner W., 1964. Jahreszyklische Untersuchungen zur Reservestoffspeicherung und Überlebensfähigkeit adulter Waldameisenarbeiterinnen (Gen. Formica, Hymenoptera, Formicidae) // Zoologische Jahrbücher. Abteilung für Systematik, Geographie und Biologie der Tiere. Bd. 71. № 1. S. 1–72.
Kneitz G., 1967. Stichproben zur jahreszeitlichen Dinamik des Waldameisenstaates (Formica polyctena Förster) // Progress in Soil Biology. Braunschweig, Amsterdam. P. 241–248.
Kneitz G.,1969. Jahreszeitliche Verȁnderungen der Ovariolenzustȁnde in der Arbeiterinnenkaste des Waldenameisenstaates von Formica polyctena Förster (Hymenoptera, Formicidae) // Zoologischer Anzeiger Rupplements. Bd. 33. Verh. Zool. S. 209–215.
Lofgren C., Banks W., Glancey R., 1986. Biology and control of imported fire ants // Annual Review of Entomology. V. 20. P. 1–30.
Mordosov I.I., 2002. Yakutia brown bear foods // International Bear News. V. 11. № 2. P. 6–7.
O’Neil J., Markin G.P., 1975. Brood development of the various castes of the imported fire ant Solenopsis invicta Buren // Journal of the Kansas Entomological Society. V. 48. № 2. P. 152–159.
Otto D., 1962. Die roten Waldameisen. Wittenberg. 152 p.
Passera L., Keller L., 1990. Relation entre la fecondite des reines de fourmis recemment accouplees el leur mode de fondation. Paris: Actes Coll., Insectes Sociaux. V. 6. P. 211–218.
Pisarski B. (ed.), 1982. Structure et organisation des sociétés de fourmis del l’espéce Formica exsecta Nyl. (Hymenoptera, Formicidae) // Memorabilia Zoologica. V. 38. P. 1–280.
Rissing S.W., Pollock G.B., 1988. Pleometrosis and polygyny in ants. Boulder: Westview press. Colorado. P. 172–222.
Roháček A., 1974. Medovice-vydatný zdroj našich lesů // Včelařstvi. P. 128–129.
Rosengren R., 1969. Notes fegarding the growth of the polycalic system in Formica uralensis Ruzsky // Notulae Entomol. V. 49. № 3. P. 211–230.
Rosengren R., 1985. Internest relations in polydomius Formica-colonies (Hymenoptera, Formicidae) // Mitteilungen der Deutschen Gesellschaft für Allgemeine und Angewandte Entomologie. Bd. 4. S. 288–291.
Schmidt H., 1974. Steuerung der Kastenbildung und Geschlechtsregulation in Waldameisenstaat. Stuttgart: Wiss. Verlag. S. 404–512.
Stamps W.T., Vinson S.B., 1991. Raiding in newly founder colonies of Solenopsis invicta Buren // Environmental Entomology. V. 20, Issue 4, 1 August 1991. P. 1037–1041.
Starr Ch.K., 2006. Steps toward a general theory of the colony cycle in social insects // Life Cycles in social insects: Behaviour, ecology and evolution. St. Petersburg Univ. Press. P. 1–20.
Stearns S.C., 1992. The evolution of life histories. Oxford: Oxford University press. 264 p.
Stearns S.C., 2000. Life history evolution: successes, limitations, and prospects // Naturwissenschaften. Bd. 87. S. 476–486.
Wiśniewski J., 1965. Pajeczaki towarzyszące mrowiskom Formica polyctena Först. (Hymenoptera, Formicidae) w nadleśnictwie dóswiadczalnym WSR Zielonka // Prace komisji Nauk Rolliczych i komisji Nauk Leśnych (Pożnan). V. 17. № 3. P. 537–584.
Zakharov A.A., 2011. Monogyny and polygyny in ant-hill life // Russian Entomological Journal. V. 20. № 3. P. 341–344.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал