

Зоологический журнал, 2022, T. 101, № 9, стр. 1061-1071
Новые находки редких видов рукокрылых (Chiroptera) на Восточном Кавказе (Дагестан)
Д. Г. Смирнов a, *, Г. С. Джамирзоев b, Ю. А. Быков c, В. П. Вехник d
a Пензенский государственный университет
440026 Пенза, Россия
b Институт экологии горных территорий имени А.К. Темботова РАН,
Государственный природный заповедник Дагестанский
367010 Махачкала, Россия
c Национальный парк “Мещèра”
601509 Гусь-Хрустальный, Россия
d Жигулевский государственный природный биосферный заповедник
имени И.И. Спрыгина
445362 Бахилова Поляна, Россия
* E-mail: eptesicus@mail.ru
Поступила в редакцию 08.10.2021
После доработки 08.12.2021
Принята к публикации 14.12.2021
- EDN: ZVNWQZ
- DOI: 10.31857/S0044513422090100
Аннотация
В ходе работ, проведенных в период с 2020 по 2021 гг. на территории Дагестана, получены новые сведения о распространении шести редких видов рукокрылых. Отловами подтверждено обитание Myotis alcathoe в северной части Восточного Кавказа. Новые находки M. emarginatus, так же как и предыдущие, сделаны исключительно в предгорье Центрального Дагестана и по большей части представлены крупными выводковыми колониями. У двух видов рода Barbastella установлены места зимовок и распространение в Дагестане, которое охватывает обширную территорию от южных границ республики до широты Кизилюрта на севере. У Rhinolophus mehelyi отмечено резкое сокращение численности в известной Карабудахкентской пещере. Новые находки Tadarida teniotis подтверждают широкое его распространение на территории республики. Данный вид впервые обнаружен на высоте 3763 м над уровнем моря, таким образом, установлен новый рекорд высотного предела его обитания. Для видов M. emarginatus, B. barbastellus и T. teniotis приведены характеристики эхолокационных сигналов.
До недавнего времени в фауне российского сектора Восточного Кавказа (Дагестан) насчитывали 16 видов рукокрылых (Амирханов, 1980). В последние два десятилетия благодаря интенсивным исследованиям были значительно расширены знания о структуре фауны рукокрылых, а количество видов увеличено до 26 (Смирнов и др., 2019). Среди впервые отмеченных видов к категории редких и требующих особого внимания относятся пять видов – Myotis emarginatus (Geoffroy 1806), M. alcathoe Von Helversen et Heller (2001), Nyctalus lasiopterus (Schreber 1780), Barbastella barbastellus Schreber (1774) и Tadarida teniotis Rafinesque (1814). С 2020 г. в ходе полевых исследований нам удалось получить новые данные о местах обитания и распространении этих и других редких видов в регионе.
В настоящей работе представлены сведения о новых находках шести редких видов рукокрылых на территории Республики Дагестан.
МАТЕРИАЛ И МЕТОДЫ
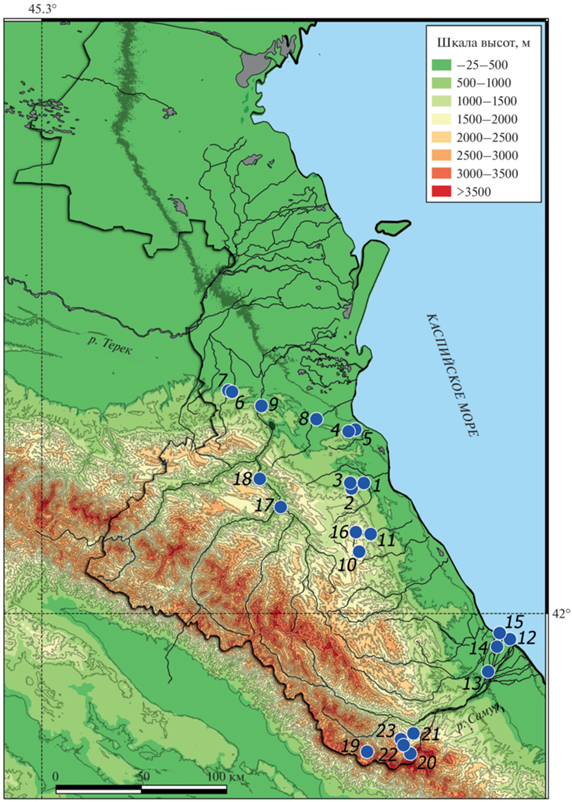
Работы проводили в июле–сентябре 2020–2021 гг. и с января по март 2021 г. Исследованиями были охвачены 12 административных районов, находящихся в пределах Кумской и Присулакской равнин, северо-западных, центральных и юго-восточных предгорий Внешнегорного Дагестана, центрального района Внутригорного и южных водораздельных хребтов Высокогорного Дагестана, а также дельты р. Самур. За указанный период зарегистрировано 24 вида. Из списка, представленного нами ранее (Смирнов и др., 2019), не удалось отметить M. bechsteinii Kuhl 1817 и Plecotus macrobullaris Kuzyakin 1965. Особое внимание было обращено на находки шести редких видов: M. emarginatus, M. alcathoe, B. barbastellus, T. teniotis, Rhinolophus mehelyi Matschie 1901 и B. leucomelas Cretzschmar 1826. Все они являются периферийными и представлены в России только (или в основном) на Кавказе, а два последних вида известны в стране исключительно с территории Дагестана. Обитание этих видов установлено в 23 локалитетах, которые указаны на рис. 1.
Рис. 1.
Места находок рукокрылых в Дагестане (координаты точек находок даны в тексте). Жирной линией оконтурены границы республики. 1 – Пещера “Живая” Карабудахкентский р-н, 2 и 3 – окр. с. Какамахи Карабудахкентский р-н, 4 и 5 – окр. с. Тарки Советский р-н, 6 и 7 – окр. с. Эндирей Хасавюртовский р-н, 8 – окр. ж/д станции Кумторкала Кумторкалинский р-н, 9 – окр. пос. Миатли Казбековский р-н, 10 – окр. с. Верх. Мулебки Левашинский р-н, 11 – окр. с. Ванашимахи Сергаколинский р-н, 12 – окр. пос. Приморский Магарамкентский р-н, 13 – окр. с. Бутказмаляр Магарамкентский р-н, 14 – окр. с. Самур Магарамкентский р-н, 15 – устье р. Карасу Магарамкентский р-н, 16 – окр. с. Зуригаудимахи Левашинский р-н, 17 – окр. с. Чалда Гергебильский р-н, 18 – окр. с. Унцукуль Унцукульский р-н, 19 – окр. с. Фий Ахтынский р-н, 20 – окр. с. Куруш Докузпаринский р-н, 21 – окр. с. Каладжух Докузпаринский р-н, 22 и 23 – гора Шалбуздаг Докузпаринский р-н.
Ночью рукокрылых отлавливали паутинными сетями по ранее описанной методике (Смирнов и др., 2017). В светлое время суток поиски рукокрылых осуществляли в потенциальных для них убежищах (пещеры, гроты, постройки человека и т.п.). Всех пойманных животных осматривали, проводили морфологические измерения (R – длина предплечья, D1 – длина первого пальца, Tib – длина голени), взвешивание (m – масса тела), кольцевание, фотографирование и после небольшой передержки отпускали в местах отлова.
Часть фаунистических данных получена путем анализа эхолокационных сигналов, записанных прибором BATLOGGER M (Elekon AG, Switzerland). Запись сигналов проходила в режиме time-expansion с фактором 10, частотой дискретизации 312.5 кГц и разрядностью 16 bit. Оцифрованные в формате “wav” записи анализировали в программе BatSound 3.31 (Pettersson Elektronik AB, Sweden). Автоматические классификаторы из-за низкой их разрешающей способности не применяли. Для анализа использовали только четкие импульсы из поисковых серий, не включающих фазы приближения и кормовые трели, а также с интенсивностью не менее –30 дБ. Из обработки были исключены записи, которые имели невысокую вероятность верного определения или по которым невозможно определить видовую принадлежность. Основные измерения в BatSound проводили на осциллограммах, спектрограммах (окно Хэннинга) и в окне спектра мощности с FFT размером 512 для Myotis и 1024 для представителей других родов. Разрешение по времени было улучшено перекрытием FFT 97%. Измеряли длину импульса (DUR), максимальную (Fmax), минимальную (Fmin), пиковую (Fpeak) частоты и расстояние между импульсами (IPI). Классификацию проводили по известным видовым характеристикам сигналов (Russo, Jones, 2002; Walters et al., 2012; Barataud, 2015; Russ, 2021), а также при их сравнении с ваучерными записями эхолокационных сигналов рукокрылых (личная колл. Д. Смирнова).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Rhinolophus mehelyi. Один из самых редких видов рукокрылых. Его находки известны только из двух точек Дагестана (Смирнов и др., 2019). Нами обитание подтверждено в пещере “Живая” (Карабудахкентская), расположенной недалеко от райцентра Карабудахкентского р-на (460 м над ур. м.) (рис. 1, точка 1). Ранее здесь существовала достаточно крупная зимующая колония, насчитывающая около 1000 особей (Амирханов, 1974, 1977), а также отмечено размножение (Кожурина, Стрелков, 1999; Смирнов и др., 2019). Однако в настоящее время из-за фактора беспокойства, вызванного частым посещением подземелья туристами, численность вида резко упала (Джамирзоев, Газарян, 2020). При осмотре пещеры 28.01.2021 нами было обнаружено всего 11 особей R. mehelyi, а при повторном посещении 24.03.2021 – 17 зверьков. В ходе осмотров, проведенных в летние месяцы 2020 и 2021 гг., особи этого вида здесь не отмечены. По всей видимости, эта пещера используется сейчас только как зимнее убежище. Однако существование здесь самой зимовки вызывает серьезные опасения. Для решения этой проблемы требуется организация ряда мер, связанных с законодательной охраной этого места в виде создания ООПТ, а также установление специальных защитных сооружений вокруг входа в пещеру для ограничения доступа людей.
Myotis emarginatus. До 2019 г. в России основное количество находок этого вида зарегистрировано на Западном Кавказе (Казаков, Ярмыш, 1974; Ярмыш и др., 1980; Газарян, 2002; колл. ЗИН РАН и ЗМ МГУ). Восточнее он был известен лишь по двум точкам из Кабардино-Балкарии (Темботов, Шхашамишев, 1984; Газарян, Темботова, 2007). На территории Дагестана M. emarginatus впервые отмечена в июле 2019 г. в искусственном подземелье в окрестностях с. Миатли (Смирнов и др., 2019). При повторном посещении этого убежища 03.07.2021 г. нами была обнаружена выводковая колония численностью около 1700 особей. Животные размещались несколькими, расположенными на небольшом расстоянии друг от друга, кластерами, три самых крупных из которых насчитывали примерно 300, 500 и 600 особей. На небольшом удалении от основных скоплений находились “ясли” приблизительно с 30 нелетными детенышами в возрасте от 4 до 11 дней. Часть из них были слепыми с небольшим пучком шерстки на холке, другая часть с почти открытыми глазами и короткой шерстью по всему телу. Кроме этого, нами был сделан ряд находок, подтверждающих обитание вида в российской части Восточного Кавказа. Одиночный зверек был обнаружен 21.07.2020 г. в одной из ракетных шахт Карабудахкентского р-на (рис. 1, точка 2: 42.64 N, 47.46 E; 704 м над ур. м.). 22.07.2020 г. в окрестностях с. Тарки Советского р-на (рис. 1, точка 4: 42.94 N, 47.45 E; 692 м над ур. м.) в заброшенном военном бункере была найдена крупная выводковая колония численностью около 300 особей. Взрослые и уже способные к полету молодые животные размещались здесь несколькими кластерами на потолке и стенах самой дальней части убежища. Ночью этого же дня, на расстоянии примерно 1 км от указанного бункера, с помощью паутинных сетей, установленных в дубо-грабовом лесном массиве, пойманы две молодые самки и один молодой самец. 23.07.2020 г. при осмотре заброшенных строений бывшего пионерского лагеря, расположенного в окр. с. Эндирей Хасавюртовского р-на (рис. 1, точка 6: 43.13 N, 46.62 E; 354 м над ур. м.), на потолке одной из комнат была обнаружена небольшая группа, состоящая из трех взрослых и трех молодых уже лётных особей. При повторном посещении этого места 14.07.2021 г. в том же помещении находилось шесть особей, все они были взрослыми. Еще одна молодая самка отловлена нами 23.07.2020 г. в окр. с. Эндирей в пойме р. Акташ (рис. 1, точка 7: 43.14 N, 46.61 E; 336 м над ур. м.).
Также в двух точках нам удалось записать эхолокационные сигналы, которые по основным параметрам импульсов соответствовали рассматриваемому виду. Их базовая частота составляла более 56 кГц, а начальная – превышала 94 кГц (табл. 1). Энергия импульсов имеет колоколообразное распределение по частоте без выраженного пика (рис. 2). Первая запись сделана 22.07.2020 г. у небольшого пожарного водоема на горе Таркитау в окр. Махачкалы (рис. 1, точка 5: 42.95 N, 47.47 E; 345 м над ур. м.), вторая – 27.07.2020 г. в долине р. Шура-Озень в 1 км выше ж/д станции Кумторкалы Кумторкалинского р-на (рис. 1, точка 8: 42.99 N, 47.21 E; 86 м над ур. м.).
Таблица 1.
Характеристики поисковых сигналов трех видов рукокрылых
| Вид | Название локалитета (число записей/ число импульсов) |
DUR, мс | Fmax, кГц | Fmin, кГц | Fpeak, кГц | IPI, мс |
|---|---|---|---|---|---|---|
| Myotis emarginatus |
Таркитау (2/18) |
4.3 ± 0.7 (3.0–5.1) |
107.1 ± 4.1 (98.9–113.1) |
37.4 ± 1.7 (34.2–39.8) |
62.8 ± 3.6 (56.2–71.0) |
71.9 ± 19.8 (43.8–119.0) |
| Шура-Озень (1/17) |
4.0 ± 0.4 (3.3–4.7) |
114.5 ± 12.3 (94.0–129.8) |
39.5 ± 1.2 (37.1–41.3) |
67.6 ± 8.4 (55.2–82.3) |
72.1 ± 19.2 (42.0–107.0) |
|
| Barbastella barbastellus | Таркитау Тип 1 (1/25) |
3.5 ± 0.4 (3.0–4.4) |
36.0 ± 0.7 (34.3–37.7) |
28.8 ± 0.4 (27.9–29.7) |
32.0 ± 0.5 (30.8–33.1) |
109.6 ± 20.3 (87.0–199.0) |
| Тип 2 (1/6) | 4.6 ± 0.8 (3.4–5.8) |
45.5 ± 2.6 (42.8–48.9) |
32.9 ± 2.4 (29.7–35.2) |
40.7 ± 0.9 (39.5–41.8) |
79.3 ± 22.2 (53.0–107.0) |
|
| Ванашимахи Тип 1 (1/17) |
4.8 ± 0.7 (3.5–5.9) |
37.6 ± 1.3 (35.5–40.0) |
28.4 ± 0.8 (27.3–29.7) |
32.6 ± 0.9 (31.5–35.4) |
133.2 ± 42.7 (82.0–232.0) |
|
| Тип 2 (1/8) | 5.3 ± 1.4 (3.8–8.4) |
43.2 ± 0.5 (42.6–44.1) |
37.0 ± 1.1 (35.5–38.3) |
41.3 ± 0.4 (40.4–41.9) |
– | |
| Tadarida teniotis |
Куруш (2/44) |
19.2 ± 2.7 (13.9–27.0) |
19.6 ± 1.6 (16.9–22.4) |
11.5 ± 0.5 (10.8–12.9) |
13.6 ± 0.7 (11.9–15.1) |
407.2 ± 73.8 (250.0–570.0) |
| Каладжух (3/7) |
19.7 ± 3.9 (15.6–24.9) |
20.5 ± 1.4 (18.9–22.6) |
12.7 ± 0.2 (12.4–12.8) |
14.8 ± 0.6 (14.3–15.9) |
713.0 ± 128.1 (549.0–862.0) |
|
| Шалбуздаг (15/167) |
15.1 ± 2.3 (9.0–22.4) |
21.2 ± 2.3 (16.9–28.5) |
11.4 ± 0.5 (10.5–12.6) |
13.5 ± 1.0 (11.7–16.4) |
461.3 ± 104.4 (230.0–770.0) |
|
| Ванашимахи (5/67) |
20.7 ± 2.3 (14.0–24.0) |
22.6 ± 4.3 (16.3–33.3) |
12.8 ± 0.7 (11.2–14.5) |
14.2 ± 1.1 (13.2–19.8) |
464.7 ± 113.4 (260.0–710.0) |
|
| Фий (5/51) |
18.6 ± 4.3 (11.9–26.0) |
19.3 ± 2.6 (14.6–26.1) |
10.8 ± 0.4 (10.2–11.4) |
12.9 ± 1.0 (11.1–14.8) |
355.2 ± 56.4 (270.0–490.0) |
Рис. 2.
Осциллограмма, спектрограмма и диаграмма спектральной плотности фрагмента записи сигналов Myotis emarginatus, сделанной 27.07.2020 г. в долине р. Шура-Озень.

Myotis alcathoe. Ранее вид был известен только с Западного Кавказа, откуда описана отдельная его подвидовая форма – M. a. circassicus (Benda et al., 2016). В российской части Восточного Кавказа он впервые зарегистрирован в 2019 г. из двух точек Дагестана. Здесь были сделаны записи эхолокационных сигналов, которые по основным характеристикам звуковых импульсов соответствовали M. alcathoe (Смирнов и др., 2019). 23 июля 2020 г. в долине р. Акташ, в окр. с. Эндирей Хасавюртовского р-на (рис. 1, точка 7: 43.14 N, 46.61 E; 336 м над ур. м.) нами были отловлены четыре особи: взрослая постлактирующая и молодая самки, взрослый и молодой самцы. Ключевые внешние промеры этих особей (табл. 2) полностью соответствовали размерам, известным для западно-кавказской и европейской популяций этой ночницы (Газярян, 2009; Dietz, Helversen, 2004; Dietz et al., 2009). Отловы в этом же локалитете других видов рукокрылых показали, что M. alcathoe встречается совместно с M. mystacinus (Kuhl 1817) и M. davidii (Peters 1869), от которых заметно отличается мелкими размерами. Таким образом, последняя наша находка подтверждает обитание вида в Дагестане. Эта территория может рассматривать как восточный форпост распространения M. alcathoe, если будет окончательно подтверждена видовая самостоятельность недавно описанной из Гирканских лесов провинции Голестан Северного Ирана морфологически сходной формы M. hyrcanicus Benda, Reiter et Vallo 2012. Предварительные результаты молекулярно-генетического анализа, проведенного с использованием одного митохондриального маркера, показали разобщение M. hyrcanicus и M. alcathoe на уровне 8.8% (Benda et al., 2012).
Таблица 2.
Внешние промеры (мм) трех видов ночниц, отловленных в окрестностях с. Эндирей Хасавюртовского р-на Дагестана
| Вид, пол, возраст | R | D1 | Tib | m | |
|---|---|---|---|---|---|
| Myotis alcathoe | Самка, ad | 32.4 | 3.6 | 14.2 | 3.8 |
| Самка, s/ad | 31.7 | 3.2 | 13.8 | 3.3 | |
| Самец, ad | 30.8 | 3.5 | 13.7 | 3.4 | |
| Самец, s/ad | 31.1 | 3.2 | 13.8 | 3.3 | |
| Myotis mystacinus | Самки, s/ad (n = 3) | 32.4–36.2 | 4.1–4.6 | 15.5–16.0 | 4.2–4.4 |
| Myotis davidii | Самец, ad | 36.2 | 5.1 | 17.0 | 5.0 |
| Самцы, s/ad (n = 3) | 33.5–34.5 | 4.4–4.9 | 15.9–16.5 | 4.2–4.8 | |
Barbastella barbastellus. Первые достоверные находки вида на территории Республики Дагестан сделаны сравнительно недавно (Смирнов и др., 2019). За период 2020–2021 нам удалось сделать еще ряд находок.
Одного взрослого самца мы нашли 21.07.2020 г. в заброшенном военном ангаре в окр. с. Какамахи Карабудахкентского р-на (рис. 1, точка 3: 42.65 N, 47.43 E; 672 м над ур. м.) и одна постлактирующая самка отловлена сетями 23.07.2020 в долине р. Акташ в окр. с. Эндирей (рис. 1, точка 7: 43.14 N, 46.61 N; 336 м над ур. м.). В зимнее время два самца обнаружены 27.01.2021 в заброшенной ракетной шахте в окр. с. Какамахи (рис. 1, точка 2: 42.64 N, 47.46 E; 704 м над ур. м.) и еще один самец – 03.02.2021 в старой штольне в окр. с. Верхние Мулебки Левашинского р-на (рис. 1, точка 10: 42.31 N, 47.51 E; 1601 м над ур. м.).
Кроме этого, эхолокационные сигналы, классифицированные нами как B. barbastellus, были записаны 22.07.2020 в лиственном лесу на горе Таркитау в окр. Махачкалы (рис. 1, точка 4: 42.95 N, 47.47 E; 345 м над ур. м.) и 13.07.2021 в долине р. Какаозень в окр. с. Ванашимахи Сергаколинского р-на (рис. 1, точка 11: 42.40 N, 47.59 E; 758 м над ур. м.) (табл. 1). Записи содержали два различных по структуре типа поисковых сигнала небольшой амплитуды: узкополосный громкий FM – “тип 1” и менее интенсивный выпуклый частотно-временнóй qCF-FM – “тип 2”. Животные издавали сигналы либо только первого типа, либо чередовали эти два типа (рис. 3).
Рис. 3.
Осциллограмма, спектрограмма и диаграмма спектральной плотности фрагмента записи сигналов Barbastella barbastellus, сделанной 13.07.2021 г. в окр. с. Ванашимахи.

Также записи сигналов с характерными параметрами для рассматриваемого вида сделаны в местах, где мы его уже регистрировали ранее: 26.07.2020 г. у рыборазводных прудов в Национальном парке “Самурский” (кластер “Дельта Самура”) в окр. пос. Приморский (рис. 1, точка 12: 41.86 N, 48.55 E; –22 м над ур. м.) и на Беркубинской лесной даче в окр. с. Бутказмаляр (рис. 1, точка 13: 41.69 N, 48.41 E; 256 м над ур. м.), 24.07.2020 г. в урочище Осел в окр. с. Самур (рис. 1, точка 14: 41.83 N, 48.47 E; 24 м над ур. м.) и 05.08.2020 г. на каспийском побережье Национального парка “Самурский” в устье р. Карасу (рис. 1, точка 15: 41.90 N, 48.50 E; 29 м над ур. м.).
Barbastella leucomelas. “Азиатских” широкоушек с Восточного Кавказа долгое время относили к B. leucomelas (Стрелков, 1981; Павлинов и др., 1995; Simmons, 2005), позже включали в состав B. darjelingensis (Hodgson 1855) (Benda et al., 2008). В последней ревизии рода Barbastella было показано, что экземпляры из Средней Азии генетически отличаются от синайских B. leucomelas и индийской B. darjelingensis, но морфологически сходны с кавказскими “азиатскими” широкоушками (Кruskop, 2015; Kruskop et al., 2019). Это обстоятельство послужило причиной объединения среднеазиатских и восточнокавказских “азиатских” широкоушек в состав одного вида – B. caspica (Satunin 1908), таксономическое описание которого было сделано по экземплярам из Азербайджана. Однако молекулярно-генетические исследования, проведенные нами на основе митохондриального маркера, показали, что экземпляры из Дагестана генетически идентичны синайской B. leucomelas, но хорошо разобщены со среднеазиатской линией (Смирнов и др., 2020). Поэтому в настоящее время для восточнокавказских “азиатских” широкоушек целесообразно использовать приоритетное в данном случае название – B. leucomelas. Окончательные выводы о видовой принадлежности данной формы можно будет сделать после изучения маркеров ядерной ДНК с привлечением материалов из Закавказья.
В России вид встречается только на Восточном Кавказе, где известен всего из 13 точек (Смирнов и др., 2019). Новые находки сделаны еще в пяти локалитетах. Так, 03.02.2021 один самец был обнаружен нами в состоянии глубокого оцепенения в небольшой штольне по добыче целестина, расположенной в окр. с. Зуригаудимахи Левашинского р-на (рис. 1, точка 16: 42.42 N, 47.50; 1203 м над ур. м.). Еще один самец найден 25.03.2021 в технической штольне недалеко от Миатлинского туннеля в окр. пос. Миатли Казбековского р-на (рис. 1, точка 9: 43.06 N, 46.82 E; 172 м над ур. м.). В самом Миатлинском туннеле в этот же день было насчитано 22 зимующих зверька.
Выявленные нами эхолокационные сигналы B. leucomelas имеют особенности и отличаются от уже описанных для этого вида сигналов (Benda et al., 2008), а также от сигналов B. barbastellus Schreber 1774. Однако в данной работе мы эти особенности не приводим, т.к. планируем посвятить этому вопросу отдельную публикацию.
Эхолокационные сигналы, классифицированные как B. leucomelas, были записаны 15.04.2021 в долине р. Аварское-Койсу в Гергебильском р-не в окрестностях сел Чалда (рис. 1, точка 17: 42.55 N, 46.96 E; 576 м над ур. м.) и Унцукуль (рис. 1, точка 18: 42.69 N, 46.82 E; 462 м над ур. м.), а также 23.07.2021 в Ахтынском р-не в окрестностях с. Фий (рис. 1, точка 19: 41.28 N, 47.56 E; 1944 м над ур. м.).
Tadarida teniotis. На территории России зверьков этого вида добывали только в двух точках Центрального Кавказа (Корнеев, Марисова, 1950; Темботов, Шабаев, 1962; Газарян, Темботова, 2007). Все остальные известные находки сделаны на основе анализа эхолокационных сигналов, в том числе и на Восточном Кавказе (Газарян, Джамирзоев, 2018; Смирнов и др., 2019). Новые находки в Республике Дагестан нами представлены также на основе анализа записей эхолокационных сигналов (табл., рис. 4).
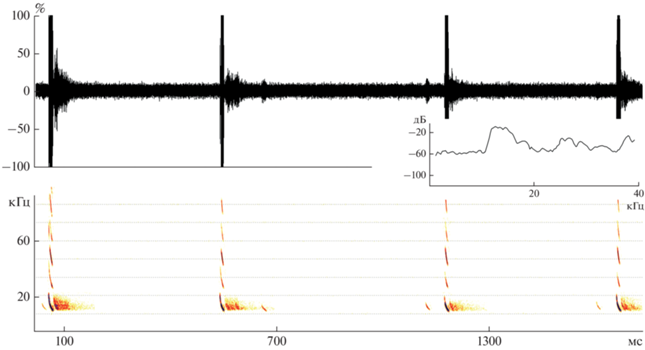
Рис. 4.
Осциллограмма, спектрограмма и диаграмма спектральной плотности фрагмента записи Tadarida teniotis, сделанной 16.07.2021 г. на горе Шалбуздаг.
По результатам записей ультразвуков летающие животные были зарегистрированы в нескольких точках. В Докузпаринском р-не в окр. с. Куруш (рис. 1, точка 20: 41.28 N, 47.83 E) на высоте 2600 м над ур. м. 18.07.2020 г. были записаны 10 последовательностей из 44 импульсов, из которых проанализировано 24. Первые сигналы были услышаны без детектора через 16 мин после захода солнца – в 19:40 по московскому времени. Далее животные непрерывно кормились в течение трех с половиной часов, летая над склонами альпийских лугов при температуре воздуха 13–15°С. В долине р. Усухчай недалеко от с. Каладжух (рис. 1, точка 21: 41.37 N, 47.89 E) на высоте 1257 м над ур. м. 19.07.2020 г. записаны четыре файла с 9 импульсами, из которых проанализировано 7. Сигналы были отмечены в период с 21:44 до 22:04, температура воздуха в это время составляла 22°С. Наибольшее количество записей было сделано вдоль скал и над склонами альпийских лугов горы Шалбуздаг. На южной ее экспозиции 16.07.2021 г. при движении по высоте от 2429 (41.27 N, 47.79 E) до 3763 м над ур. м. (41.33 N, 47.81 E) (рис. 1, точка 22) с 21.13 до 22.21, при температуре от 13 до 10°С, записан 61 файл с 282 импульсами, из которых проанализировано 119. На северной части этой же горы (рис. 1, точка 23: 41.36 N, 47.81 E) 21.07.2021 г. на высоте 3019 м над ур. м. с 23:00 до 02:04 при температуре воздуха 15°С, было записано 17 файлов, содержащих 126 импульсов. Из этого числа было проанализировано 48 импульсов.
В Сергоколинском р-не эхолокационные и коммуникативные сигналы охотящихся животных отмечены 13.07.2021 г. в долине р. Какаозень недалеко от с. Ванашимахи (рис. 1, точка 11: 42.40 N, 47.59 E; 758 м над ур. м.). Здесь за небольшой промежуток времени с 19:58 до 20:05, при температуре 23°С, записано 13 файлов с более 200 импульсами, из которых мы проанализировали 67.
В Ахтынском р-не записи сигналов одиночно летающих зверьков были сделаны 23 и 25.07.2021 г. в окр. с. Фий (рис. 1, точка 19: 41.28 N, 47.56 E; 1944 м над ур. м.). Первые сигналы были зарегистрированы через 40 мин после захода солнца, или в 19:58, а самые поздние – почти за 2 ч до восхода, или в 02:03, при температуре 15–18°С.
Сигналы, информация о которых приведена выше, относятся к T. teniotis. По основным характеристикам они полностью соответствуют сигналам этого вида. Возможность обитания в этих локалитетах N. lasiopterus, пиковая частота сигналов которого во время поискового полета может быть близкой T. teniotis (Haquart, Disca, 2007; Bec et al., 2008), очень маловероятна. Все записи были сделаны на достаточно большой высоте над уровнем моря, часто вблизи отвесных скал, и практически в отсутствие древесной растительности и, тем более, крупных лесных массивов, что крайне нетипично для мест обитания дендрофила N. lasiopterus. Тем не менее встречи последнего вида в таких местах, например на пролете во время осенних или весенних миграций, не исключены. Основанием для такого предположения могут служить отловы в середине июля 2011 г. трех самцов N. lasiopterus в верхнем течении р. Сулак грузинского района Тушети (J. Aihartza, личное сообщение) (Смирнов и др., 2019). Весь регион Тушети находится на высоте около 2000 м над ур. м. и в непосредственной близости от границ Дагестана. Бóльшая часть этой территории, в отличие от дагестанской стороны, занята лесными массивами, поэтому может быть перспективной для обитания N. lasiopterus.
Записи эхолокационных сигналов T. teniotis, сделанные нами во всех указанных локалитетах, характеризовались одной важной особенностью, которая ранее была впервые выявлена и описана для сигналов этого вида из Кабардино-Балкарии, но никак не отмечена в сигналах с территории Дагестана (Газарян, Джамирзоев, 2018). Во всех наших записях, где встречаются мощные импульсы, присутствует от пяти до восьми гармоник, общая полоса пропускания достигает 120 кГц, а энергия от самых низких гармонических составляющих к более высоким убывает не равномерно, а дискретно (рис. 5). Самой интенсивной после фундаментальной в большинстве случаев была третья гармоника. Почему наблюдается именно не равномерное распределение энергии по гармоникам – остается пока непонятным. При этом давно известно, что использование импульсов с множеством составляющих связано с ориентацией рукокрылых в пространстве в условиях ревербации и для улучшения “захвата” цели (Zbinden, 1988). Как показали исследования (Bates et al., 2011), гармоники позволяют сосредоточиться на релевантных объектах и тем самым изолировать цель от различных фоновых отражений. После “захвата” цели сонаром зверьки могут ее отслеживать, несмотря на множество эхосигналов от других объектов. Это происходит благодаря фокусировке центра луча-сонара на эхосигналах, где находится цель, а эхосигналы от помех на периферии не учитываются. Высокочастотные гармоники формируют более узкий луч, чем луч низкочастотных, и поэтому точнее концентрируются на целевых объектах, тогда как не целевые оказываются за пределами диапазона узкого луча. Кроме того, высокочастотные звуки быстрее затухают в воздухе, поэтому высокие гармоники, когда отражаются, например, от удаленных объектов, возвращаются к летучей мыши более слабыми. Исходя из сказанного, мы предполагаем, что перенос мощности со второй гармоники на третью гармонику связано с увеличением эффективности фокусировки на специфических объектах питания в момент охоты при больших скоростях полета.
Рис. 5.
Спектрограмма и диаграмма спектральной плотности мильтикомпонентного импульса Tadarida teniotis: h1 – фундаментальная (основная) гармоника, h2–h5 – высокочастотные гармоники.

T. teniotis был зарегистрирован нами в широком диапазоне высот, с максимальным значением более 3700 м над ур. м. Согласно литературным данным, в Средиземноморье (Dietz et al., 2009), на Кавказе (Газарян, Джамирзоев, 2018), в Закавказье (Кузякин, 1950; Явруян, Сафарян, 1975; Рахматулина, 2001), на Ближнем Востоке (Benda et al., 2006, 2008, 2010, 2012) и в Средней Азии (Богданов, 1953; Стрелков, Шаймарданов, 1983; Rybin et al., 1989) вид находили на высотах, не превышающих 1500 м над ур. м., в Непале – до 2500 м над ур. м. (Sharma et al., 2021), а известный предел высоты, отмеченный в Альпах, составляет 3100 м над ур. м. (Arlettaz et al., 2000; Benda, Piraccini, 2016). Таким образом, высота нашей находки, сделанной на юге Дагестана, превышает этот максимум на 660 м.
ЗАКЛЮЧЕНИЕ
Новые данные о находках рукокрылых значительно расширяют представления об их распространении и характере пребывания в регионе. Так, в Республике впервые отловлена M. alcathoe, которая в России до этого была известна только с территории Западного Кавказа, а на сопредельных территориях – в ряде районов Грузии и в Армении (Дундарован, 2018; Газарян, неопубликованные данные). Судя по данным отлова и регистрации, обитание этого вида приурочено к пойменным старовозрастным лесам, где он может встречаться совместно с M. mystacinus и M. davidii. Все находки M. emarginatus сделаны на предгорных территориях Центрального Дагестана. Местами его обитания здесь служат как крупные лесные массивы, так и открытые пространства с обязательным присутствием водоемов. Находки крупных выводковых колоний свидетельствуют о размножении и высокой концентрации особей вида в локальных местах обитания. Найденная нами колония в окр. с. Миатли – самое многочисленное из известных в настоящее время скоплений этого вида в ареале (Piraccini, 2016). Убежище, где находится колония, легко доступно для посещения, а животные крайне уязвимы к воздействию беспокойства и нуждаются в законодательной охране вместе с убежищем. У представителей рода Barbastella распространение в Дагестане охватывает обширную территорию, их находки известны от южных границ республики до широты Кизилюрта на севере и связаны с карстовыми районами в предгорьях и горными территориями до высот 2260 (B. barbastellus) и 2500 (B. leucomelas) м над ур. м. Основные летние места обитания у B. barbastellus приурочены к лесным стациям, а у B. leucomelas – к аридным территориям. R. mehelyi в России обнаружен только в Дагестане, где достоверно известна единственная колония в пещере Живая. За последние несколько десятилетий здесь отмечено резкое сокращение численности вида, где он еще пока встречается в небольшом количестве зимой и, возможно, в ранневесенний период. Пещера легкодоступна и нуждается как в физической защите входа, так и в законодательном установлении режима ее охраны. Новые находки T. teniotis подтверждают ранее высказанное мнение о широком его распространении на территории республики (Газарян, Джамирзоев, 2018; Смирнов и др., 2019). На настоящий момент в Дагестане, по данным записей эхолокационных сигналов, известно уже 13 точек обнаружения этого вида, а также установлен новый рекорд высотного предела его обитания.
Список литературы
Амирханов З.М., 1974. О зимовке подковоносов Мегели (Rhinolophus mehelyi Matschie) в Дагестане // Материалы 1 Всесоюз. совещ. по рукокрылым. Ленинград, Зоол. ин-т АН СССР. С. 91–92.
Амирханов З.М., 1977. Экологические наблюдения за зимней спячкой летучих мышей Дагестанской АССР // Материалы науч. конф. Дагестанского филиала географического общества СССР. Махачкала. № 7. С. 41–43.
Амирханов З.М., 1980. Размещение рукокрылых в Дагестане // Рукокрылые (Chiroptera). М.: Наука. С. 63–69.
Багданов О.П., 1953. Рукокрылые фауны Узбекской СССР. Т. 3. Млекопитающие. Вып. 2. Ташкент: АН УзбССР. 159 с.
Джамирзоев Г.С., Газарян С.В., 2020. Подковонос Мегели – Rhinolophus mehelyi (Blasius, 1853) // Красная книга Республики Дагестан. Махачкала: Типография ИП Джамалудинов М.А. С. 686–687.
Газарян С.В., 2002. Эколого-фаунистический анализ населения рукокрылых (Chiroptera) Западного Кавказа. Дис. … канд. биол. наук: 03.00.08. М. 225 с.
Газарян С.В., 2009. Новый вид ночниц на Кавказе: Myotis alcathoe или Myotis caucasicus? // Plecotus et al. № 11–12. С. 50–61.
Газарян С.В., Джамирзоев Г.С., 2018. Новые сведения о распространении широкоухого складчатогуба Tadarida teniotis (Chiroptera, Molossidae) в России // Plecotus et al. № 21. С. 3–18.
Газарян С.В., Темботова Ф.А., 2007. Новые находки рукокрылых (Chiroptera) на Центральном Кавказе // Зоологический журнал. Т. 86. № 6. С. 761–762.
Дундарова Х.И., 2018. Молекулярна филогения на морфо-групата: Myotis mystacinus (Chiroptera: Vespertilionidae) на Балканския полуостров. Дис. … за присъждане на образователна и научна степен “доктор”: научна специалност 01.06.02 “Зоология”. София. 81 с.
Казаков Б.А., Ярмыш Н.Н., 1974. О фауне рукокрылых Предкавказья // Материалы I Всесоюз. совещ. по рукокрылым (Сhiroptera). Л.: Зоол. ин-т АН СССР. С. 69–72.
Кожурина Е.И., Стрелков П.П., 1999. Редкие виды рукокрылых фауны бывшего СССР и России // Редкие виды млекопитающих России и сопредельных территорий. М.: Териологическое общество. С. 168–187.
Корнеев О.П., Марисова И.В., 1950. Новая находка в СССР широкоухого складчатогуба (Tadarida teniotis Rafin.) // Науковi записки Киiвського ун-та. Т. 9. Вып. 6. С. 159–160.
Кузякин А.П., 1950. Летучие мыши. М.: Советская наука. 443 с.
Павлинов И.Я., Борисенко А.В., Крускоп С.В., Яхонтов Е.Л., 1995. Млекопитающие Евразии. II. Non-Rodentia: систематико-географический справочник. Москва: МГУ. 338 с.
Рахматулина И.К., 2001. Вертикальное распространение рукокрылых в Восточном Закавказье // Plecotus et al. № 4. С. 41–50.
Смирнов Д.Г., Курмаева Н.М., Иваницкий А.Н., 2017. К изучению рукокрылых (Chiroptera) на востоке Крыма // Plecotus et al. № 20. С. 17–29.
Смирнов Д.Г., Джамирзоев Г.С., Газарян С.В., Вехник В.П., Быков Ю.А., 2019. Рукокрылые (Chiroptera) Дагестана: обзор фауны по итогам исследований в 2017–2019 // Plecotus et al. № 22. С. 3–48.
Смирнов Д.Г., Вехник В.П., Джамирзоев Г.С., 2020. О таксономическом статусе “азиатских” широкоушек (Chiroptera: Vespertilionidae: Barbastella) на Восточном Кавказе // Plecotus et al. № 23. С. 3–12.
Стрелков П.П., 1981. Отряд Chiroptera – рукокрылые // Каталог млекопитающих СССР. Плиоцен – современность. Ред. Громов И.М., Барановой Г.И. Ленинград: Наука. С. 29–53.
Стрелков П.П., Шаймарданов Р.Т., 1983. Новые данные по распространению летучих мышей (Chiroptera) в Казахстане // Труды Зоол. ин-та АН СССР. Т. 119. С. 3–37.
Темботов А.К., Шабаев М.И., 1962. Новый вид рукокрылых фауны Кабардино-Балкарии // Уч. зап. Каб.-Бал. ун-та. № 14. С. 124.
Темботов А.К., Шхашамишев Х.Х., 1984. Животный мир Кабардино-Балкарии. Нальчик: Эльбрус. 55 с.
Явруян Э.Г., Сафарян Л.А., 1975. Находки широкоухого складчатогуба (Tadarida teniotis Rafinesque) на территории Армянской ССР // Биологический журнал Армении. № 28 (7). С. 90–93.
Ярмыш Н.Н., Казаков Б.А., Сонина И.Ю., Усвайская А.Х., 1980. О колонии трехцветной ночницы и большого подковоноса в Хаджохской пещере // Рукокрылые. (Chiroptera). М.: Наука. С. 179–181.
Arlettaz R., Ruchet C., Aeschimann J., Brun E., Genoud M., Vogel P., 2000. Physiological traits affecting the distribution and wintering strategy of the bat Tadarida teniotis // Ecology. V. 81 (4). P. 1004–1014.
Bates M.E., Simmons J.F., Zorikov T.V., 2011. Bats Use Echo Harmonic Structure to Distinguish Their Targets from Background Clutter // Science. V. 333 (6042). P. 627–630. https://doi.org/10.1126/science.1202065
Barataud M., 2015. Acoustic ecology of European bats. Species Identification and Studies of Their Habitats and Foraging Behaviour. Biotope Editions, Mèze; National Museum of Natural History, Paris (collection Inventaires et biodiversité). 340 p.
Bec J., Haquart A., Julien J.-F., Disca T., 2008. New criteria for the acoustic identification of the Greater Noctule, Nyctalus lasiopterus, lead to a better knowledge of its distribution in France // In: Abstracts of XI European Bat Research Symposium, Cluj-Napoca, Romania. P. 21.
Benda P., Andreas M., Kock D., Lučan R. K., Munclinger P., Nová P., Obuch J., Ochman K., Reiter A., Uhrin M., Weinfurtová D., 2006. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean. Part 4. Bat fauna of Syria: distribution, systematics, ecology // Acta Societatis Zoologicae Bohemicae. V. 70. P. 1–329.
Benda P., Dietz C., Andreas M., Hotovy J., Lucan R.K. et al., 2008. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 6. Bats of Sinai (Egypt) with some taxonomic, ecological and echolocation data on that fauna // Acta Societatis Zoologicae Bohemicae. V. 72. P. 1–103.
Benda P., Lučan R.K., Obuch J., Reiter A., Andreas M., Bačkor P., Bohnenstengel T., Eid E.K., Ševčík M., Vallo P., Amr Z.S., 2010. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 8. Bats of Jordan: fauna, ecology, echolocation, ectoparasites // Acta Societatis Zoologicae Bohemicae. V. 74. P. 185–353.
Benda P., Faizolâhi K., Andreas M., Obuch J., Reiter A., Ševčík M., Uhrin M., Vallo P. Ashrafi S., 2012. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 10. Bat fauna of Iran // Acta Societatis Zoologicae Bohemicae. V. 76. P. 163–582.
Benda P., Gazaryan S., Vallo P., 2016. On the distribution and taxonomy of bats of the Myotis mystacinus morphogroup from the Caucasus region (Chiroptera: Vespertilionidae) // Turkish Journal of Zology. V. 40. P. 1–8.
Benda P., Piraccini R., 2016. Tadarida teniotis. The IUCN Red List of Threatened Species 2016: e.T21311A22114995. Downloaded on 21 September 2021.https://doi.org/10.2305/IUCN.UK.2016-2.RLTS. T21311A22114995.en
Dietz C., Helversen O. von, 2004. Illustrated identification key to the bats of Europe. Electronic publication. Version 1.0. Tuebingen & Erlangen. 72 p.
Ditz C., Von Helversen O., Nill D., 2009. Bats of Britain, Europe & Northwest Africa. London: Black Publisers Ltd. 400 p.
Haquart A., Disca T., 2007. Caractéristiques acoustiques et nouvelles données de Grande Noctule Nyctalus lasiopterus (Schreber, 1780) en France // Le Vespère. № 1. P. 15–20.
Kruskop S.V., 2015. Dark and pale: taxonomic status of the barbastelle (Barbastella: Vespertilionidae, Chiroptera) from Central Asia // Acta Chiropterologica. V. 17 (1). P. 49–57.
Kruskop S.V., Kawai K., Tiunov M.P., 2019. Taxonomic status of the barbastelles (Chiroptera: Vespertilionidae: Barbastella) from the Japanese archipelago and Kunashir Island // Zootaxa. V. 4567 (3). P. 461–476.
Piraccini R., 2016. Myotis emarginatus. The IUCN Red List of Threatened Species 2016: e.T14129A22051191. Downloaded on 21 September 2021.https://doi.org/10.2305/IUCN.UK.2016-2.RLTS. T14129A22051191.en
Rybin S.N., HoráČek I., Červený J., 1989. Bats of southern Kirghizia: distribution and faunal status // European Bat Research 1987. Hanák V., HoráČek I., Gaisler J. (eds). Praha: Charles University Press. P. 421–441.
Russo D., Jones G., 2002. Identification of twenty-two bat species (Mammalia: Chiroptera) from Italy by analysis of time-expanded recordings of echolocation calls // Journal of Zoology (Lond.). V. 258. P. 91–103.
Russ J., 2021. Bat Calls of Britain and Europe. A Guide to Species Identification. Milton Keynes: Pelagic Publishing Ltd. 432 p.
Sharma B., Chakravarty R., Acharya P.R., 2021. The first record of European free-tailed bat, Tadarida teniotis Rafinesque, 1814, and note on probable elevational movement from Nepal // Journal of Asia-Pacific Biodiversity. V. 14. P. 248–253.
Simmons N.B., 2005. Order Chiroptera // Mammal species of the World: a taxonomic and geographic reference, Third Edition. Vol. 1. Wilson D.E. & Reeder D.M. (Eds). Baltimore: Johns Hopkins University Press. P. 312–529.
Walters C.L., Freeman R., Collen A., Dietz C., Fenton M.B. et al., 2012. A continental-scale tool for acoustic identification of European bats Journal of Applied Ecology. V. 49. P. 1064–1074.
Zbinden K., 1988. Harmonic Structure of Bat Echolocation Signals // Animal Sonar. NATO ASI Science (Series A: Life Sciences). Nachtigall P.E., Moore P.W.B. (Eds). V. 156. Boston: Springer, MA. https://doi.org/10.1007/978-1-4684-7493-0_58
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал