

Зоологический журнал, 2023, T. 102, № 7, стр. 826-840
Одонтологическая изменчивость инвазивного вида на стадии расселения: Microtus rossiaemeridionalis (Arvicolinae, Rodentia) в Иркутской области, Южное Предбайкалье
Е. А. Маркова a, *, С. А. Борисов b, **, С. В. Зыков a, ***, П. А. Сибиряков a, ****, Л. Э. Ялковская a, *****, С. В. Булычева a, ******
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
b Иркутский научно-исследовательский противочумный институт Роспотребнадзора
664047 Иркутск, ул. Трилиссера, 78, Россия
* E-mail: emrk@yandex.ru
** E-mail: borisov.irk@mail.ru
*** E-mail: svzykov@yandex.ru
**** E-mail: p_sibiryakov@rambler.ru
***** E-mail: lida@ipae.uran.ru
****** E-mail: bulycheva.2201@mail.ru
Поступила в редакцию 01.04.2023
После доработки 27.04.2023
Принята к публикации 10.05.2023
- EDN: AQAWSA
- DOI: 10.31857/S0044513423070073
Аннотация
Восточноевропейская полевка – факультативно синантропный вид, активно расселяющийся в восточной части северной Евразии. Территория распространения вида в Иркутской области известна как дизъюнктивный участок ареала с 1980-х. Изучено 98 особей, отловленных в 2016–2017 и 2021–2023 гг. на восточном участке расселения, где вид впервые был обнаружен в начале 2010-х гг. Животных отлавливали на антропогенно трансформированных территориях от окрестностей г. Иркутcк до 23-го километра Голоустненского тракта; в естественных биотопах вид в отловах не встречен. Для видовой идентификации использованы молекулярно-генетические маркеры. Морфологические и микроструктурные характеристики щечных зубов изучены с применением методов световой и электронной микроскопии. В ходе анализа одонтологической изменчивости оценивали сложность жевательной поверхности, регулярность чередования ее функциональных элементов – режущих граней, а также отмечали наличие или отсутствие атипичных призматических складок и призм (редких невидоспецифичных признаков, проявляющихся и накапливающихся у разных видов Arvicolinae при близкородственном скрещивании, но ранее не известных у восточноевропейской полевки). Частоты признаков, характеризующих сложность жевательной поверхности и регулярность ее функциональных элементов, у изученных животных находятся в пределах значений, известных для основного ареала вида. В двух точках отлова впервые для восточноевропейской полевки обнаружены особи с атипичными призматическими складками. При максимальном проявлении этих складок от задней непарной петли нижних зубов отделяется сверхкомплектная призма, топологически соответствующая гипокониду. Распределение ламеллярной и радиальной эмали на сверхкомплектной призме не согласуется с функционально обусловленным и эволюционно закрепленным распределением типов эмали на зубах рода Microtus. Сделан вывод о том, что в изученной локальной микропопуляции проявляются фенотипические последствия восстановления численности из небольшого числа основателей. По-видимому, в районе исследований вид сталкивается с множественными последовательными случаями закрепления (натурализации) на новой территории. Полученные результаты подтверждают перспективность использования дентальных признаков для мониторинга видов Arvicolinae, границы распространения которых подвергаются значительным пространственным изменениям в относительно краткие сроки, доступные для наблюдения.
Биологическая инвазия – это процесс вторжения вида и формирования его устойчивой популяции в регионах за пределами естественного ареала, включающий три основные стадии: перемещение (транспортировка, интродукция), закрепление (натурализация) и расселение (например, Blackburn et al., 2011; Richardson, Riccardi, 2013). Деятельность человека – важнейший фактор, способствующий биологическим инвазиям, и с ростом антропогенного влияния на биосферу все чаще инвазии чужеродных видов рассматривают как угрозу для биоразнообразия естественных экосистем (Galiana et al., 2014; Mačić et al., 2018). Для инвазивных видов процесс расселения за пределы основной части ареала на первый взгляд положителен, поскольку демонстрирует черты биологического прогресса. Вместе с тем оценка эволюционного потенциала инвазивных популяций – это комплексная междисциплинарная задача, требующая не только длительных наблюдений, но и синтеза данных разных дисциплин (например, Sherpa, Després, 2021). Фенотипические последствия прохождения через стадии инвазии при этом рассматривают в связи со способностью вида приспосабливаться к новым условиям. Однако стадия перемещения в большинстве случаев связана с критическим сокращением численности, что, теоретически, может иметь и негативные последствия (Allendorf, Lundquist, 2003). На практике увеличение частоты отдельных рецессивных признаков и проявление фенотипических аберраций отмечают в популяциях транслоцированных видов млекопитающих, которые, тем не менее, успешно осваивают новые территории (например, Кораблев и др., 2018). Каким образом инвазивные виды преодолевают негативные последствия восстановления численности из небольшого числа основателей – это один из фундаментальных вопросов при изучении инвазионного процесса.
Среди млекопитающих одним из перспективных объектов изучения фенотипических последствий инвазии является восточноевропейская полевка (Microtus rossiaemeridionalis Ognev 1924) – факультативно синантропный вид, активно расселяющийся в восточной части северной Евразии (Мейер и др., 1996; Малыгин и др., 2019). Дизъюнктивные участки ареала, появляющиеся в результате непреднамеренной интродукции, служат естественными лабораториями, в которых могут быть прослежены фенотипические последствия формирования популяции из ограниченного числа основателей (Markova et al., 2019). В трех из пяти изученных ранее дизъюнктивных участков ареала обнаружено проявление скрытой фенотипической изменчивости: резкое повышение доли резервных морфотипов зубов, появление редукций и накопление редких одонтологических вариаций (Markova et al., 2019). Материалы из Иркутской обл. также были включены в процитированную работу (небольшая серия черепов из окрестностей пос. Новонукутский, коллекция М.Н. Мейер, ЗИН РАН), однако фенотипических последствий формирования популяции из ограниченного числа основателей выявлено не было, что могло быть связано как с небольшим объемом выборки, так и с отсутствием фенотипически выраженных эффектов критического сокращения численности.
Территория распространения восточноевропейской полевки в Иркутской обл. известна как дизъюнктивный участок ареала с 1980-х годов, когда впервые по кариологическим данным была установлена видовая принадлежность полевок из окрестностей пос. Новонукутский (Липин и др., 1987; Мейер и др., 1996). Источник проникновения вида в регион не известен, предположительно восточноевропейская полевка была завезена по железной дороге (Обыкновенная полевка…, 1994). Расселение в Иркутской обл. шло во всех направлениях: к 2008–2013 гг. получены многочисленные подтвержденные находки восточноевропейской полевки, относящиеся к северному, восточному и юго-восточному направлениям расселения (Малышев, 2013). Наибольшего распространения и численности вид достиг в агроценозах левого берега Ангары в Верхнем Приангарье; известны находки на правобережье Ангары (Малышев, 2013), на о-ве Ольхон (Pavlova, Tchabovsky, 2011), а также в соседних регионах, например в Бурятии, в г. Улан-Уде и его окрестностях (Моролдоев, Картавцева, 2017; Моролдоев и др., 2017). Распространение вида в Иркутской обл. достаточно подробно документировано (Липин и др., 1987; Демидович, 2006, 2016; Попов, 2011; Малышев, 2013), что делает этот участок ареала перспективным для изучения фенотипической изменчивости инвазивного вида на стадии расселения.
Цель работы – ответить на вопрос, прослеживаются ли у восточноевропейской полевки в Иркутской обл. фенотипические последствия восстановления численности из небольшого числа основателей.
Для анализа фенотипической изменчивости выбраны одонтологические признаки. Морфология зубов восточноевропейской полевки сформировалась в ходе эволюции в направлении повышения приспособленности к питанию растительными кормами с высоким содержанием клетчатки (более подробно – см. Markova et al., 2019). Несмотря на строгую связь морфологии зубов с особенностями их функции, при близкородственном скрещивании даже у наиболее высоко специализированных представителей подсемейства Arvicolinae проявляются редкие невидоспецифичные дентальные признаки, снижающие приспособленность (Markova, Smirnov, 2018; Markova et al., 2020). Эта особенность зубной системы делает ее перспективным объектом изучения для выявления фенотипических последствий критического сокращения численности и, вместе с тем, позволяет сопоставлять выявляемые тенденции внутривидовой изменчивости с масштабами эволюционных изменений от предковых форм к современным таксонам.
МАТЕРИАЛ И МЕТОДЫ
Сбор материала проводили на восточном участке расселения M. rossiaemeridionalis в Иркутской обл. Точки отловов расположены в Иркутском р-не в направлении от окрестностей Иркутска до 23-го километра Голоустненского тракта, который связывает областной центр с пос. Большое Голоустное на берегу оз. Байкал. Отловы проведены С.А. Борисовым в четырех точках. Ловушки устанавливали в местах обнаружения нор со следами жизнедеятельности полевок, при наличии снежного покрова – вблизи отдушин на поверхности снега. Оценка численности грызунов не входила в задачи отловов.
Точка a – 23-й километр Голоустненского тракта, садовое некоммерческое товарищество (СНТ) для выращивания столовых корне- и клубнеплодных культур и многолетних культурных растений. Отловы на территории СНТ и в зоне естественной растительности за пределами товарищества проводили круглогодично с 1999 г., восточноевропейская полевка отмечена с 2011 г. только на территории СНТ. В естественных ландшафтах за пределами СНТ на протяжении всего периода отловов восточноевропейская полевка не обнаружена. Серые полевки в естественных ландшафтах представлены полевкой- экономкой и темной полевкой; периодически оба вида проникали на территорию СНТ, не формируя постоянных поселений. Материал для анализа одонтологической изменчивости собран в 2016 (5 особей), 2017 (14 особей), 2021 (25 особей), 2022 (30 особей) и 2023 (2 особи) годах.
Точка b – 15-й километр Голоустненского тракта. Отловы проводили в апреле 2022 г. на окраине д. Худяково в зарослях рудеральной растительности вдоль линии электропередач (14 особей).
Точка c – 9-й километр Голоустненского тракта. Отловы проводили в апреле 2022 г. на узкой полосе земли между автодорогой и нерегулярно возделываемым сельскохозяйственным полем (4 особи).
Точка d – р. Кая, окраина рабочего пос. Маркова близ г. Иркутск, левобережье р. Ангара. Отловы проводили на урбанизированной территории в октябре 2022 г. (4 особи).
Точки отлова a–c (рис. 1) расположены на правобережье Ангары на Иркутско-Черемховской равнине в направлении к предгорьям Онотской возвышенности и Приморского хребта, в зоне подтаежных подгорных лесов (Растительный покров…, 2013), преимущественно сосновых и лиственнично-сосновых, с участками осиново-березовых лесов и присутствием осоковых и осоково-гипновых болот с березой, кедром, елью и лиственницей с небольшой долей заболоченных лугов и лесов. Коренная растительность в значительной мере трансформирована хозяйственной деятельностью человека и подвержена частым лесным пожарам (например, Баянова, 2018). Точка d расположена на левобережье Ангары, в окрестностях г. Иркутск (рис. 1).
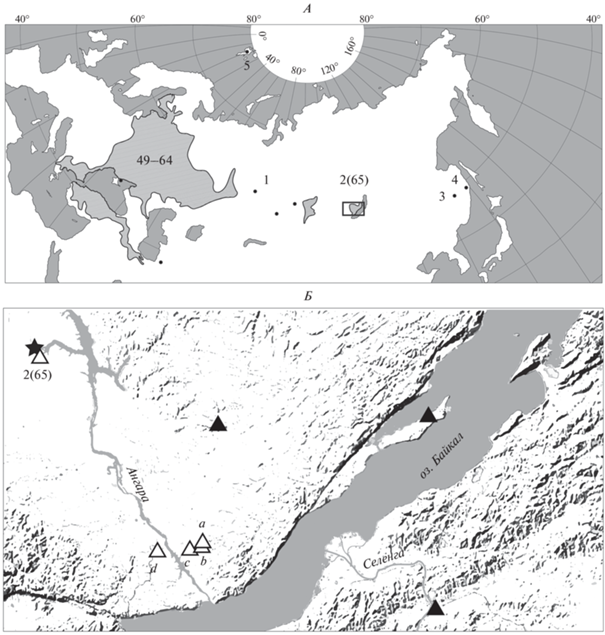
Рис. 1.
Географическое положение региона исследований и точки сбора материала. А – ареал восточноевропейской полевки с указанием положения региона исследований (прямоугольник) и номеров выборок, использованных в качестве сравнительного материала (49–65 – по: Markova et al., 2010; 1–5 – по: Markova et al., 2019). Б – точки сбора материала в Иркутской области: a – 23-й километр Голоустненского тракта, b – 9-й километр Голоустненского тракта, c – 15-й километр Голоустненского тракта, d – р. Кая. Звездочка – первая подтвержденная находка вида в регионе (Липин и др., 1987), черные треугольники – опубликованные данные о подтвержденных находках вида (Pavlova, Tchabovsky, 2011; Малышев, 2013; Моролдоев и др., 2017), белые треугольники – точки, включенные в анализ морфологических признаков.
Видовая идентификация всех особей восточноевропейской полевки из точек отлова a–d проведена по молекулярно-генетическим маркерам в лаборатории филогенетики и биохронологии ИЭРиЖ УрО РАН. Тотальную ДНК выделяли из образцов мышечной (фиксация в 96% этаноле) или костной (нижняя челюсть) ткани с использованием наборов “ДНК-Экстран-2” (“СИНТОЛ”) согласно протоколу производителя. Для ПЦР-типирования использовали пары праймеров, рекомендованные для идентификации видов-двойников обыкновенной полевки (Nekrutenko et al., 1999):
Mar14F 5'_CCCCTAAAGATTGTTACAGAAACCATC_3'
Mar14R 5'_AAGCCCAACTTGTAACCAGATAAAGCAC_3'
Mro16F 5'_GCTCGCTGGTAGAGACAGTCACC_3'
Mro16R 5'_GAAGGCAGATTGACCAAGATTTCC_3'
В работе использована терминология зубной системы Arvicolinae (Van Der Meulen, 1973) и Cricetidae (Reig, 1977; Maridet, Ni, 2013). Анализ одонтологической изменчивости проводили с использованием разработанных ранее подходов (Маркова, 2013; Markova et al., 2010, 2019, 2020). Рассматривали три группы признаков. Группа I – cложность жевательной поверхности; оценивали в тех отделах зубов, которые усложнялись в ходе эволюции Arvicolinae от предковых форм к современным за счет добавления новых призм – типичных элементов сложности для подсемейства Arvicolinae (рис. 2). Для оценки использовали ранги сложности (Маркова, 2013) и традиционные морфотипы (Rörig, Börner, 1905; Markova et al., 2010). Группа II – регулярность чередования эмалевых граней как функциональных элементов жевательной поверхности; оценивали на примере m1 и m2 в наиболее узкой части на контакте призм коронки (рис. 3). Группа III – редкие атипичные призматические складки или сверхкомплектные призмы на месте элементов сложности бугорчатого зуба предковых форм (рис. 2, признаки 1–7) и складки эмали во входящих углах передней непарной петли m1 и задней непарной петли М3 (рис. 2, признак 8). Обозначение признаков – по: Markova et al., 2020.
Рис. 2.
Номенклатура жевательной поверхности щечных зубов полевок (по: Van Der Meulen, 1973), места оценки рангов сложности и традиционных морфотипов в отделах, которые усложнялись в ходе эволюции полевок от предковых форм к современным за счет добавления новых призм – типичных элементов сложности для подсемейства Arvicolinae (серый цвет) и места проверки на наличие атипичных элементов жевательной поверхности (стрелки). Черные кружки – расположение сверхкомплектных призм или складок на месте элементов сложности бугорчатого зуба ранних хомякообразных, которые у полевок в норме не проявляются, но могут быть обнаружены при близкородственном скрещивании; белые кружки – эмалевые складки в основании головки передней непарной петли m1 и пятки M3; 1–8 – номера признаков по: Markova et al., 2020.
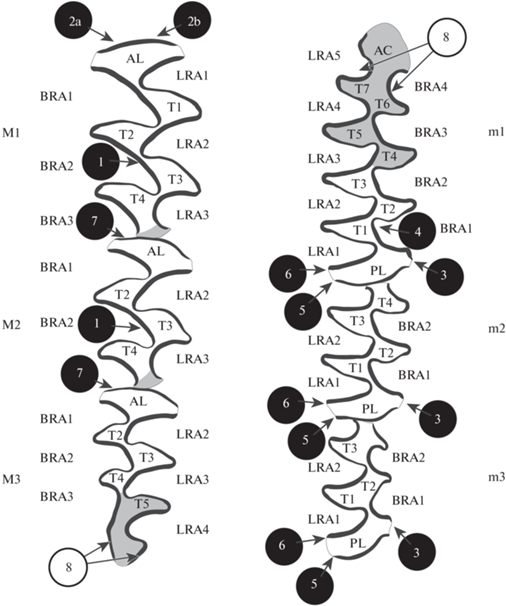
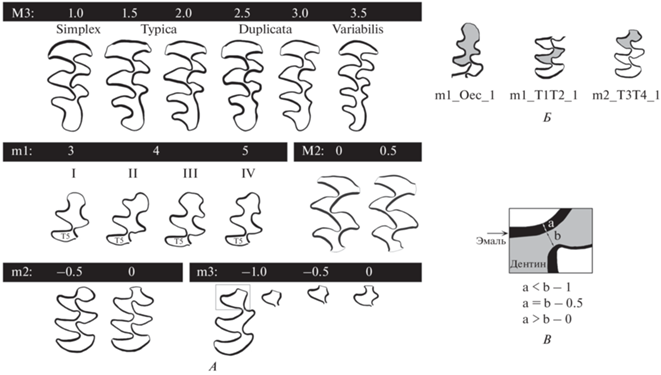
Рис. 3.
Признаки группы I, характеризующие сложность жевательной поверхности (A), и группы II, характеризующие регулярность режущих граней эмали (Б–В). A – На черном фоне показаны ранги сложности, ниже – традиционные морфотипы. Б – Примеры отклонений от регулярного чередования режущих граней эмали на m1 и m2. В – Принцип оценки регулярности режущих граней по ширине дентинового слияния между соседними призмами (а – ширина наиболее толстой эмалевой грани в месте контакта двух призм, b – расстояние между эмалевыми гранями противолежащих входящих углов, 0, 0.5, 1 – степень слияния полей жевательной поверхности от полного разделения (0) до неполного разделения (0.5) и полного слияния (1)).
В анализ включены правые и левые щечные зубы животных на пост-ювенильных стадиях (жевательная поверхность всех зубов, включая М3, полностью сформирована, отсутствует ювенильная складчатость эмали).
Всех особей проверяли на наличие функциональных патологий (значительных нарушений окклюзии, стирания или роста зубов), болезней ротовой полости (кариес, актиномикоз), следов травм или структурных нарушений тканей зубов, видимых в световой микроскоп.
Анализ микроструктуры эмали проводили с использованием сканирующего электронного микроскопа TESCAN VEGA3. Для получения микрофотографий зубы извлекали из челюсти, заливали эпоксидной смолой, после чего жевательную поверхность зуба выравнивали и обрабатывали 10% HCl в течение 3 с. Определение типов эмали проводили по: von Koenigswald, 1980.
Материал, использованный в данной работе, находится на хранении в Музее Института экологии растений и животных УрО РАН. Сравнительный материал для анализа одонтологической изменчивости (рис. 1A) включает опубликованные данные о выборках восточноевропейской полевки из основной части ареала (Markova et al., 2010) и из дизъюнктивных участков (Markova et al., 2019). В т.ч. изучена коллекция М.Н. Мейер из окрестностей пос. Новонукутский, Иркутская обл. (в предыдущих работах под номерами 65 (Markova et al., 2010) или 2 (Markova et al., 2019)), хранящаяся в ЗИН РАН и включающая материалы 1984 г. (ZIN70593-70602) и 5 черепов животных без музейных номеров, датированных 1985 г.
Обозначения зубов: m1, m2, m3 – нижние, M1, M2, M3 – верхние зубы. Элементы жевательной поверхности зубов подсемейства Arvicolinae: AC – головка передней непарной петли m1, AL – передняя непарная лопасть верхних зубов, BRA – буккальные входящие углы, LRA – лингвальные входящие углы, PL – задние непарные лопасти нижних зубов, T – треугольные призмы жевательной поверхности, ПНП – передняя непарная петля m1 (ПНП = Т6 + Т7 + AC). Элементы жевательной поверхности зубов Cricetidae (и соответствующее обозначение по терминологии, принятой для Arvicolinae, где применимо): ed – энтоконид (= Т1), hd – гипоконид (= Т0), hl – гиполофулид, lpsd – лабиальный постеросинусид (= BRA0), md – метаконид (= Т3), pld – постеролофид, prd – протоконид (= T2). При обозначении признаков на отдельных зубах сначала указывается зуб, после этого – признак, например, m1_lpsd – лабиальный постеросинусид на m1 и т.д. Признаки, характеризующие степень слияния или разделения дентиновых полей (например, отклонения от регулярного чередования эмалевых граней), обозначали, например, следующим образом: m1_Т1Т2_0.5 (т.е. между призмами T1 и Т2 на m1 имеется дентиновое слияние, равное 0.5) (по: рис. 3В).
РЕЗУЛЬТАТЫ
Изученные особи из точек отлова а–d не имели признаков функциональных патологий (нарушений окклюзии, стирания или роста зубов), болезней ротовой полости (кариес, актиномикоз), следов травм или структурных нарушений тканей зубов, видимых в световой микроскоп.
Частоты признаков, характеризующих сложность жевательной поверхности и регулярность ее функциональных элементов – режущих граней эмали (табл. 1, I , II), находятся в пределах значений, известных для основного ареала (Markova et al., 2010, выборки 49–64), и близки к значениям, опубликованным для выборки 65(2) из окрестностей пос. Новонукутский.
Таблица 1.
Частоты одонтологических признаков (%), обнаруженных у восточноевропейской полевки в точках отлова a–d и общее число особей на пост-ювенильных онтогенетических стадиях, включенных в анализ одонтологической изменчивости
| Признаки | Точки и годы отлова | ||||||
|---|---|---|---|---|---|---|---|
| а | b | c | d | ||||
| 2016 | 2017 | 2021 | 2022 | 2022 | 2022 | 2022 | |
| I – Ранги сложности зубов (в скобках – традиционные морфотипы) | |||||||
| M3_1 (simplex) | 0.0 | 0.0 | 22.0 | 2.0 | 0.0 | 0.0 | 0.0 |
| M3_1.5 (typica) | 50.0 | 23.1 | 22.2 | 28.6 | 18.5 | 0.0 | 0.0 |
| M3_2 (typica) | 0.0 | 57.7 | 44.4 | 40.8 | 37.0 | 28.6 | 100.0 |
| M3_2.5 (duplicata) | 20.0 | 0.0 | 8.9 | 6.1 | 11.1 | 0.0 | 0.0 |
| M3_3 (duplicata) | 10.0 | 11.5 | 22.2 | 20.4 | 33.3 | 14.3 | 0.0 |
| M3_3.5 (variabilis) | 20.0 | 7.7 | 0.0 | 2.0 | 0.0 | 57.1 | 0.0 |
| Всего M3 | 10 | 26 | 45 | 49 | 27 | 7 | 8 |
| m1_rank3 (I) | 80.0 | 100.0 | 91.7 | 87.5 | 96.4 | 50.0 | 100.0 |
| m1_rank4 (II) | 20.0 | 0.0 | 2.1 | 1.8 | 0.0 | 25.0 | 0.0 |
| m1_rank4 (III) | 0.0 | 0.0 | 4.2 | 5.4 | 3.6 | 12.5 | 0.0 |
| m1_rank5 (IV) | 0.0 | 0.0 | 2.1 | 5.4 | 0.0 | 12.5 | 0.0 |
| Всего m1 | 10 | 26 | 48 | 56 | 28 | 8 | 8 |
| M2_rank0 | 60.0 | 84.6 | 84.4 | 88.9 | 92.9 | 71.4 | 100.0 |
| M2_rank0.5 | 40.0 | 15.4 | 15.6 | 11.1 | 7.1 | 28.6 | 0.0 |
| Всего M2 | 10 | 26 | 45 | 54 | 28 | 7 | 8 |
| m2_rank0 | 90.0 | 100.0 | 95.7 | 92.9 | 96.4 | 100.0 | 100.0 |
| m2_rank-0.5 | 10.0 | 0.0 | 4.3* | 7.1* | 3.6 | 0.0 | 0.0 |
| Всего m2 | 10 | 26 | 46 | 56 | 28 | 8 | 8 |
| m3_rank0 | 30.0 | 28.6 | 33.3 | 37.7 | 64.3 | 100.0 | 75.0 |
| m3_rank-0.5 | 70.0 | 39.3 | 37.5 | 41.5 | 25.0 | 0.0 | 25.0 |
| m3_rank-1 | 0.0 | 32.1 | 29.2 | 20.8 | 10.7 | 0.0 | 0.0 |
| Всего m3 | 10 | 28 | 48 | 53 | 28 | 8 | 8 |
| II – Отклонения от регулярного чередования режущих элементов жевательной поверхности | |||||||
| m1_T6T7AC_0 (= maskii) | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 0.0 |
| m1_T5ПНП (= Oec)_0.5 | 0.0 | 0.0 | 2.1 | 0.0 | 0.0 | 0.0 | 0.0 |
| m1_T1T2_0.5 | 10.0 | 3.8 | 6.3 | 1.8 | 7.1 | 0.0 | 0.0 |
| m1_T1T2_1 | 0.0 | 3.8 | 8.3 | 0.0 | 3.6 | 0.0 | 0.0 |
| m2_T3T4_0.5 | 0.0 | 3.8 | 4.3 | 0.0 | 3.6 | 12.5 | 0.0 |
| m2_T3T4_1 | 20.0 | 7.7 | 26.1 | 8.9 | 0.0 | 0.0 | 0.0 |
| m2_T1T2_0.5 | 0.0 | 0.0 | 2.2 | 0.0 | 0.0 | 0.0 | 0.0 |
| m2_T1T2_1 | 0.0 | 11.5 | 8.7 | 0.0 | 0.0 | 0.0 | 0.0 |
| III – Атипичные элементы жевательной поверхности | |||||||
| m1_lpsd (= признак 3) | 0.0 | 0.0 | 4.2 | 8.9 | 0.0 | 0.0 | 0.0 |
| m2_lpsd (= признак 3) | 0.0 | 0.0 | 2.1 | 5.4 | 0.0 | 0.0 | 0.0 |
| m3_lpsd (= признак 3) | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 12.5 | 0.0 |
| m3_hl (= признак 6) | 0.0 | 0.0 | 0.0 | 1.8 | 0.0 | 0.0 | 0.0 |
| M3_признак 8 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 0.0 |
| Число особей | 5 | 14 | 25 | 29 | 14 | 4 | 4 |
В двух локалитетах (точки отловов а и с) обнаружены редкие атипичные признаки – призматические складки на месте элементов сложности бугорчатых зубов хомякообразных предков (рис. 4–5, табл. 1 , III). Из семи редких дентальных признаков, связанных у Arvicolinae с близкородственным скрещиванием (рис. 2, 1–7 ), встречено два: признак 3 – дополнительный входящий угол BRA0 на месте lpsd бугорчатого зуба хомякообразных и признак 6 – дополнительный входящий угол LRA1a на месте hl хомякообразных. Это первый случай обнаружения признаков данной группы у M. rossiaemeridionalis.
Рис. 4.
Редкие одонтологические признаки, обнаруженные у восточноевропейской полевки в Иркутской обл.: атипичные входящие углы на задней непарной петле нижних зубов (белые стрелки) и неполная редукция призмы Т4 на m2 (черные стрелки). A, Б – левые и правые m1 и m2 с признаком 3 (самец IPAE835372/И107, июль 2022, точка отловов a), В – m2 и m3 особи IPAE835473/И208 (пол неизвестен, февраль 2022, точка отловов a), Г – m3 с признаком 3 (самец IPAE835356.2/И89, апрель 2022, точка отловов c), Д – m3 cо слабо выраженным признаком 6 (самец IPAE1700481/И2, декабрь 2021, точка отловов a). Масштаб 3 мм.
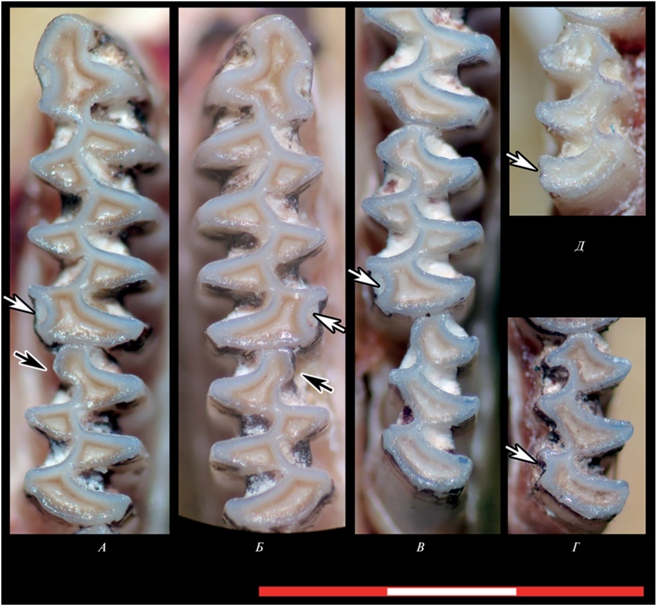
Рис. 5.
Левые нижние m1 и m2 взрослой самки IPAE835373/И108 (точка отловов a, сентябрь 2022 г.) и левый нижний m3 полувзрослого самца IPAE835356.2/И89 (точка отловов с, апрель 2022 г.) с атипичным входящим углом BRA0. А, В, Д – контуры жевательной поверхности; Б, Г, Е – микроструктура эмали. А, Б – m1; B, Г – m2; Д, Е – m3. Стрелками показано расположение типов эмали на призме hd (= T0), не соответствующее “микротусному” варианту дифференциации эмали при максимальной выраженности признака 3 (особь IPAE835373/И108): тройные черные стрелки – толстая стенка, сформированная радиальной и ламеллярной эмалью, одиночная белая стрелка – тонкая стенка, сформированная радиальной эмалью. Масштаб 200 мкм.
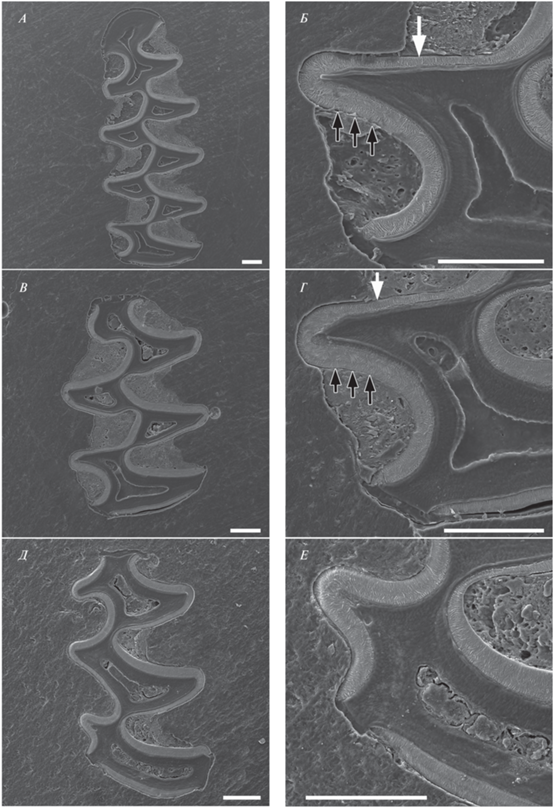
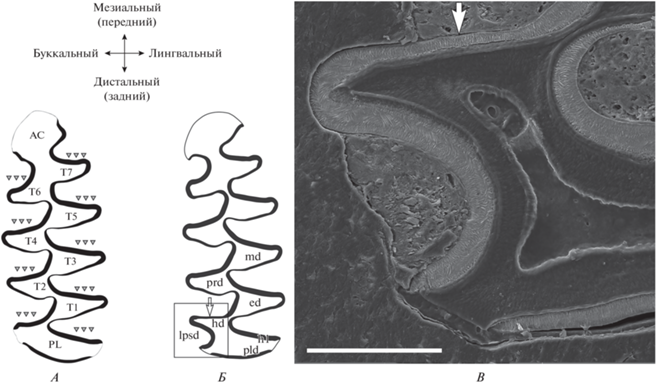
Рис. 6.
Обратное расположение ламеллярной и радиальной эмали на передней и задней стенках сверхкомплектной призмы hd (= Т0) при формировании атипичного входящего угла lpsd (= BRA0) у восточноевропейской полевки. А – Схема эволюционно закрепленного и функционально значимого “микротусного” типа дифференциации эмали, при котором передние (ведущие) стенки призм Т1–Т5 сформированы ламеллярной и радиальной эмалью и утолщены, а задние (ведомые) стенки тонкие, сформированы радиальной эмалью; тройные треугольные стрелки – ведущие эмалевые грани призм; обозначение элементов зуба – по традиционной схеме для Arvicolinae (Van Der Meulen, 1973). Б – Расположение типов эмали на сверхкомплектной призме hd (= T0), не согласующееся с “микротусным” типом дифференциации эмали: передняя стенка тонкая, задняя толстая; обозначение элементов зуба – по схеме, принятой для Cricetidae (Reig, 1977; Maridet, Ni, 2013). B – Микроструктура зубной эмали при формировании lpsd; белая одиночная стрелка – тонкая передняя стенка сверхкомплектной призмы hd (= T0), сформированная радиальной эмалью; масштаб 200 мкм.
Признак 6 (m1_hl) встречен единично (точка отловов а) и выражен слабо (рис. 4 Д). Признак 3 (lpsd на m1, m2 или m3) встречен в двух локалитетах, удаленных друг от друга на 8 км. В точке отловов c одна особь из четырех отловленных имела признак 3 на левом m3, остальные зубы этой особи были обычного для вида строения, без атипичных элементов (рис. 4Г). В точке а в июле 2021 г. отловлен один взрослый самец с хорошо выраженным признаком 3 на правом и левом m1 и левом m2, в феврале 2022 г. – одна особь с признаком 3 на левом m1 и правом m2 (рис. 4В), в июле 2022 – один полувзрослый самец с признаком 3 на правом и левом m1 и незначительным изгибом дистальных стенок m2 (рис. 4А–4Б), в сентябре 2022 – взрослая размножавшаяся самка с признаком 3 на правых и левых m1 и m2 (рис. 5А–5Г). Оценки частоты рассматриваемых признаков и объем выборок приведены в табл. 1 .
ОБСУЖДЕНИЕ И ЗАКЛЮЧЕНИЕ
В ходе расселения восточноевропейской полевки в антропогенно трансформированных ландшафтах зоны подтаежных подгорных лесов южного Предбайкалья формируются поселения вида, в которых на локальном уровне происходят накопление (до 9%) и усиление фенотипической выраженности редких одонтологических признаков. Обнаруженные признаки проявляются у представителей разных родов Arvicolinae вне зависимости от филогенетического родства, и их выраженность усиливается в условиях инбридинга (Markova, Smirnov, 2018; Markova et al., 2020). Учитывая приуроченность M. rossiaemeridionalis к территории садоводства и отсутствие находок в близлежащих естественных биотопах, точка отловов а представляет собой частично изолированное поселение, из которого вид на протяжении периода отловов не расселялся активно. Наиболее вероятно, в настоящий момент восточноевропейская полевка находится на стадии закрепления (т.е. натурализации) на данной территории. Неоднократное появление и усиление выраженности редких фенотипических признаков является косвенным свидетельством того, что этой локальной группировке пока не удается преодолевать последствия близкородственного скрещивания и редкие одонтологические признаки имеют тенденцию к усилению своего проявления. Произойдет ли их элиминация, будут ли они персистировать как редкие периодически проявляющиеся вариации или их частота будет увеличиваться – это вопрос для дальнейших наблюдений. Однако обнаруженный признак 3 (lpsd) не появился de novo в локалитете a. Присутствие в точке отлова c особи с атипичным входящим углом BRA0 на одном из m3 (начальная стадия фенотипического проявления признака 3 при близкородственном скрещивании у Arvicolinae (Markova et al., 2020)) указывает на то, что восточноевропейская полевка на правобережье Ангары предрасположена к проявлению этого признака. Остается неясным, проявляется ли этот признак лишь на правобережье Ангары, либо его фенотипическое проявление связано с эффектом основателя для всей иркутской популяции вида? Для ответа на этот вопрос требуются дополнительные исследования. Отсутствие редких одонтологических признаков в выборке 2 (65) из окрестностей Новонукутского (предположительно зона исходного расселения восточноевропейской полевки в регионе) и их проявление в пределах области современного расселения вида согласуются с моделью множественных последовательных случаев закрепления (натурализации) расселяющейся инвазивной популяции (Blackburn et al., 2011). При такой интерпретации можно ожидать, что редкие фенотипические признаки будет легче обнаружить в небольших изолированных поселениях на стадии закрепления на новой территории, чем на стадии расселения, когда высока численность особей с типичной для вида морфологией, а фон отклонений относительно низок. Имеющихся данных недостаточно, чтобы определить стадию инвазионного процесса, на которой впервые проявился обнаруженный редкий признак 3 (lpsd) в Иркутской обл. Для ответа на этот вопрос целесообразно сравнить локальные частично изолированные микропопуляции вида, формирующиеся в нескольких направлениях расселения в Верхнем Приангарье и Южном Прибайкалье.
Накопление редких одонтологических признаков отмечено и в других дизъюнктивных участках ареала M. rossiaemeridionalis (Markova et al., 2019), однако признаки 3 и 6 у этого вида до сих пор не были выявлены. Как и в других изолированных участках ареала восточноевропейской полевки, обнаруженные в локалитетах a и c редкие одонтологичесские признаки инадаптивны для зеленоядного вида и не согласуются с исходным для вида направлением эволюционных преобразований зубной системы. Ранее было высказано предположение, что такие проявления деспециализации зубной системы могут потенциально быть перспективны при переходе на более генерализованную диету (Markova et al., 2019), однако для проверки этого предположения требуются целенаправленные исследования.
У близкого вида – обыкновенной полевки (Microtus arvalis Pallas 1778) кариоформы arvalis – признак 3 (lpsd) на m1 был обнаружен в северо-западной части Иберийского п-ова, на территории Испании (Markova et al., 2010). Палеонтологические находки указывают на присутствие этого признака у обыкновенной полевки в северо-восточной части полуострова в позднем плейстоцене (Luzi, Lopez-Garcia, 2019, 2019a). Несмотря на то, что одонтологическая изменчивость M. arvalis в пределах области распространения формы arvalis традиционно привлекает внимание палеонтологов (например, Chaline, 1972; Nadachowski, 1982), а также описано множество редких признаков и фенотипических аберраций, встречающихся в Европе в настоящее время (Janossy, Schmidt, 1960; 1975; Kraft, 2000; Kapischke, 2014; Kapischke et al., 2015; Jentzsch et al., 2020), признак 3 (lpsd) пока ни разу не был обнаружен у этого таксона за пределами Испании.
У обыкновенной полевки формы obscurus признак 3 был обнаружен лишь при лабораторном разведении (Markova et al., 2020), признак проявился сначала на m3 (в третьем поколении) и лишь затем на m1 (c 6 поколения). Такая же последовательность проявления признака 3 характерна и для копытных леммингов рода Dicrostonyx в лабораторных колониях, однако на m3 признак был обнаружен и в природных популяциях, из которых получены основатели колоний (Markova, Smirnov, 2018). На m2 признак 3 ранее у Arvicolinae никогда не был обнаружен. Примечательно, что у восточноевропейской полевки признак 3 на m2 во всех случаях ассоциирован с частичной редукцией T4. В случаях, когда призма Т0 (гипоконид) отделена от PL достаточно сильно, на ней прослеживается реверсия типов эмали. Причем эта реверсия не согласуется с функционально значимым и эволюционно закрепленным типом дифференциации эмали, специфичным для рода Microtus (рис. 6). Наиболее вероятным объяснением этому может служить модификация морфогенеза зуба с изменением экспрессии генов в мезиально-дистальном градиенте на стадии закладки бугорков таким образом, что в норме стабильный дистальный отдел нижнего зуба начинает дифференцироваться, а развитие мезиального отдела замедляется. В результате на нижних зубах образуется сверхкомплектная призма, топологически соответствующая гипокониду бугорчатозубых грызунов, и дифференциация эмали на ней оказывается отображена зеркально в передне-заднем направлении.
Изучение природы редких атипичных призматических складок щечных зубов Arvicolinae, рекапитулирующих элементы сложности низкокоронковых зубов их хомякообразных предков, имеет фундаментальное значение в связи с развитием представлений о норме строения и фоне отклонений (Коваленко, 2003). В локалитетах a и c в Иркутской обл. мы обнаружили два из семи атипичных признаков, известных для Arvicolinae (признаки 1–7 по: Markova et al., 2020). В случае своей максимальной фенотипической выраженности эти признаки могут демонстрировать сходство с элементами зубов мутантных мышей с экспериментально модифицированной экспрессией генов сигнального пути EDA (Kangas et al., 2004; Rodrigues et al., 2013). При этом аномалий зубной системы, которые проявляются при экспериментальной модификации других сигнальных путей, вовлеченных в развитие зубов у млекопитающих (Seppala et al., 2017; Cho et al., 2011), у полевочьих пока не обнаружено. Сигнальный путь гена EDA, кодирующего белок эктодисплазин-А, является эволюционно древним (Pantalacci et al., 2008), вовлечен в формирование производных эктодермы, а его нарушения связаны с развитием синдромных аномалий. Вероятно, первым шагом для выяснения факторов проявления атипичных складок зубов у полевок, рекапитулирующих признаки ранних хомякообразных, может быть проверка гипотезы о связи проявления этих признаков с изменениями в сигнальном пути гена EDA. Примечательно, что, согласно имеющимся сборкам геномов, ген EDA у полевок расположен на Х-хромосоме (например, MicOch1.0 для Microtus ochrogaster Wagner 1842 (Broad Institute…, 2012) и mArvAmp1.2 для Arvicola amphibius Linnaeus 1758 (Wellcome Sanger Institute…, 2021), так же как и у других млекопитающих, включая домовую мышь и человека (Genome Reference Consortium…, 2020, 2022). Проявление необычного варианта Х-хромосомы у M. rossiaemeridionalis в регионе исследований (Pavlova, Tchabovsky, 2011), а также мутации Х-хромосомы в лабораторных колониях копытных леммингов (Gileva, Chebotar, 1979; Gileva, 2004), где впоследствии описано наибольшее разнообразие дентальных аномалий, соответствующих признакам 1–7 (Markova, Smirnov, 2018; Markova et al., 2020; Чепраков, 2022), косвенно подтверждает целесообразность проверки выдвинутой гипотезы.
Повышение частоты встречаемости редких дентальных признаков в условиях частичной изоляции не является специфической особенностью инвазивных видов и проявляется у Arvicoinae в местах флуктуации границ распространения и при периодических миграциях из оптимальных местообитаний в субоптимальные с образованием эфемерных локальных поселений (Markova et al., 2020). В случае, когда граница распространения остается стабильной, накопление редких признаков не прослеживается, как, например, было показано для обыкновенной полевки формы obscurus близ северной границы ареала на Урале (Маркова и др., 2013). Можно предполагать, что фенотипические последствия близкородственного скрещивания у полевочьих проявляются в результате снижения генетического разнообразия в локальных микропопуляциях, формирующих так называемый фронт экспансии (Swaegers et al., 2013). Полученные нами результаты подтверждают перспективность использования дентальных признаков для мониторинга видов Arvicolinae, границы распространения которых подвергаются значительным пространственным изменениям в относительно краткие сроки, доступные для наблюдения.
Однако то, какие механизмы стоят за проявлением скрытой (латентной) одонтологической изменчивости в небольших изолированных группировках, – еще предстоит установить. Это может быть связано с фиксацией редких аллелей, всплеском эпигенетической изменчивости, например за счет метилирования ДНК (Chapelle, Silvestre, 2022), или сочетанием генетических и эпигенетических механизмов. В этой связи совместный анализ морфологической и генетической изменчивости является перспективным направлением исследований. Обнаружение редкого варианта X-хромосомы у восточноевропейской полевки на о-ве Ольхон (Pavlova, Tchabovsky, 2011) позволяет предполагать, что специфика изолированных популяций восточноевропейской полевки может быть прослежена на разных уровнях организации.
Список литературы
Баянова А.А., 2018. Анализ горимости лесных ресурсов Иркутской области // Мониторинг. Наука и технологии. № 2. С. 35–38.
Демидович А.П., 2006. Антропогенная трансформация сообществ грызунов как компонента паразитарных систем // Бюллетень ВСНЦ СО РАМН. № 2 (48). С. 28–33.
Демидович А.П., 2016. Сообщества грызунов сельскохозяйственных угодий Иркутской области // Вестник Иркутской государственной сельскохозяйственной академии. № 76. С. 97–102.
Коваленко Е.Е., 2003. Эффект нормы признака и его теоретическое значение // Эволюционная биология: история и теория. Вып. 2. / Отв. ред. Колчинский Э.И. Ред.-сост. Попов И.Ю. СПб.: Политехника-сервис. С. 66–87.
Кораблев Н.П., Кораблев П.Н., Кораблев М.П., 2018. Микроэволюционные процессы в популяциях транслоцированных видов: евроазиатский бобр, енотовидная собака, американская норка. М.: Товарищество научных изданий КМК. 402 с.
Липин С.И., Хромичек С.И., Похряева А.Н., Суханов Н.А., Мирончук Ю.В., Якубенок М.И., 1987. Восточноевропейская полевка – носитель туляремии в южных районах Восточной Сибири // Вопросы региональной гигиены, санитарии и эпидемиологии. Тез. докл. науч.-практ. конференции, 18–19 дек., 1987. Вып. 2. Якутск, 1987. С. 167–169.
Малыгин В.М., Баскевич М.И., Хляп Л.А., 2019. Инвазии видов-двойников обыкновенной полевки // Российский журнал биологических инвазий. № 4. С. 71–93.
Малышев Ю.С., 2013. К вопросу о формировании нового участка ареала восточноевропейской полевки Microtus rossiaemeridionalis Ognev в Прибайкалье // Байкальский зоологический журнал. № 1 (12). С. 105– 108.
Маркова Е.А., 2013. Оценка сложности щечных зубов полевок (Arvicolinae, Rodentia): ранжированный морфотипический подход // Зоологический журнал. Т. 92. № 8. С. 968–980.
Маркова Е.А., Ялковская Л.Э., Зыков С.В., 2013. Морфологическая и хромосомная изменчивость обыкновенной полевки Microtus arvalis Pall. на северной границе распространения // ДАН. Т. 448. № 1. С. 109–112.
Мейер М.Н., Голенищев Ф.Н., Раджабли С.И., Саблина О.В., 1996. Серые полевки (подрод Microtus) фауны России и сопредельных территорий. СПб.: Зоол. ин-т РАН. 320 с.
Моролдоев И.В., Картавцева И.В., 2017. Новые данные об инвазии восточноевропейской полевки (Microtus rossiaemeridionalis) на восток от г. Улан-Удэ // Вестник Бурятского государственного университета. Биология. География. Вып. 3. С. 130–134.
Моролдоев И.В., Шереметьева И.Н., Картавцева И.В., 2017. Первая находка восточноевропейской полевки (Microtus rossiaemeridionalis) в Бурятии // Российский журнал биологических инвазий. № 2. С. 88–94.
Обыкновенная полевка: виды-двойники, 1994. / В.Е. Соколов [и др.]. М.: Наука. 432 с.
Попов В. В., 2011. Млекопитающие Иркутской области (аннотированный список) // Байкальский зоологический журнал. № 1 (6). С. 69–78.
Растительный покров Иркутской области, 2013. [Электронный ресурс] ИРКИПЕДИЯ. Эл. дан. URL: http://irkipedia.ru/content/rastitelnyy_pokrov_atlas (дата обращения 20.03.2023)
Чепраков М.И., 2022. Фенотипическая изменчивость и наследование нетипичной формы anterior lobe M1 копытных леммингов (Dicrostonyx, Rodentia, Arvicolinae) // Известия РАН. Серия биологическая. № 5. С. 482–488.
Allendorf F.W., Lundquist L.L., 2003. Introduction: population biology, evolution, and control of invasive species // Conservation Biology. V. 17. P. 24–30.
Blackburn T.M., Pyšek P., Bacher S., Carlton J.T., Duncan R.P., Jarošık V., Wilson J.R.U., Richardson D.M., 2011. A proposed unified framework for biological invasions // Trends in Ecology & Evolution. V. 26. P. 333–339.
Broad Institute, Microtus ochrogaster genome assembly MicOch1.0, 2012. [Электронный ресурс]. Режим доступа: https://www.ncbi.nlm.nih.gov/assembly/GCF_ 000317375.1. Дата обновления: 07.12.2012. Дата последнего доступа: 25.04.2023.
Chaline J., 1972. Les rongeurs du Pleistocene moyen et superieur de France (Systematique – Biostratigraphie – Paleoclimatologie), Cahiers de Paléontologie. Paris: C.N.R.S. 410 p.
Chapelle V., Silvestre F., 2022. Population Epigenetics: The Extent of DNA Methylation Variation in Wild Animal Populations // Epigenomes. V. 6. № 4. P. 31.
Cho S.-W., Kwak S., Woolley T.E., Lee M.J., Kim E.J., Baker R.E., Kim H.J., Shin J.S., Tickle C., Maini P.K., Jung H.S., 2011. Interactions between Shh, Sostdc1 and Wnt signalling and a new feedback loop for spatial patterning of the teeth // Development. V. 138. P. 1807– 1816.
Galiana N., Lurgi M., Montoya J.M., López B.C., 2014. Invasions cause biodiversity loss and community simplification in vertebrate food webs // Oikos. V. 123 (6). P. 721–728.
Genome Reference Consortium, Mouse Build 39 (GRCm39), 2020. [Электронный ресурс]. Режим доступа: https:// www.ncbi.nlm.nih.gov/assembly/GCF_000001635.27. Дата обновления: 24.06.2020. Дата последнего доступа: 25.04.2023.
Genome Reference Consortium, Human Build 38 patch release 14 (GRCh38.p14), 2022. [Электронный ресурс]. Режим доступа: https://www.ncbi.nlm.nih.gov/assembly/ GCF_000001405.40. Дата обновления: 03.02.2022. Дата последнего доступа: 25.04.2023.
Gileva E.A., 2004. The B chromosome system in the varying lemming Dicrostonyx torquatus Pall., 1779 from natural and laboratory populations // Russian Journal of Genetics. V. 40. P. 1399–1406.
Gileva E.A, Chebotar N.A., 1979. Fertile XO males and females in the varying lemming, Dicrostonyx torquatus Pall. (1779) // Heredity. V. 42. P. 67–77.
Janossy D., Schmidt E., 1960. Extreme Varianten des M1 der Feldmaus (Microtus arvalis Pallas) in Ungarn // Vertebrata Hungarica. V. 2. P. 137–142.
Janossy D., Schmidt E., 1975. Extreme Varianten des M1 der Feldmaus (Microtus arvalis Pallas) in Ungarn. II // Zeitschrift für Säugetierkunde. V. 40. P. 34–36.
Jentzsch M., Kraft R., Lemkul A., Kapischke H.-J., Maternowski H.J., Wolf R., 2020. Anomalies and pathological changes of skulls and dentition of wild small mammal species from Germany // Journal of Vertebrate Biology. V. 69 (4). 20072.
Kangas A.T., Evans A.R., Thesleff I., Jernvall J., 2004. Nonindependence of mammalian dental characters // Nature. V. 432. P. 211–214.
Kapischke H., 2014. Zur Variabilität der Zähne von Feldmäusen Microtus arvalis (Pallas, 1779) aus Sachsen (Übersicht zu einer Sammlung im Rahmen des Projektes: Atlas der Säugetiere Sachsens) // Veröffentlichungen des Museums der Westlausitz Kamenz. H. 32. S. 85–110.
Kapischke H., Wilhelm M., Fabian K., 2015. Zahnbesonderheit einer Feldmaus Microtus arvalis aus Eulengewöllen // Ornithologische Mitteilungen. № 67. S. 128.
Koenigswald W von., 1980. Schmeltzstruktur und Morphologie in den Molaren der Arvicolidae (Rodentia) // Abhandlungen der Senckenbergische Naturforschende Gesellschaft. H. 239. S. 1–139.
Kraft R., 2000. Ungewöhnliche Molarenbildungen bei Feldmaus (Microtus arvalis) und Erdmaus (Microtus agrestis) // Säugetierkundliche Informationen. 4. H. 23–24. S. 587–589.
Luzi E., Lopez-García H.-M., 2019. Relative size variations in two vole species: A climatic proxy for the identification of humid-arid pulses during Late Pleistocene in Southwestern Europe? // Quaternary Science Reviews. V. 223. 105920.
Luzi E., Lopez-García H.-M., 2019a. Patterns of variation in Microtus arvalis and Microtus agrestis populations from Middle to Late Pleistocene in southwestern Europe // Historical Biology. V. 31. № 5. P. 535–543.
Mačić V., Albano P.G., Almpanidou V., Claudet J., Corrales X., Essl F., Evagelopoulos A., Giovos I., Jimenez C., Kark S., Marković O., Mazaris A.D., Ólafsdóttir G.Á., Panayotova M., Petović S., Rabitsch W., Ramdani M., Rilov G., Tricarico E., Vega Fernández T., Sini M., Trygonis V., Katsanevakis S., 2018. Biological invasions in conservation planning: a global systematic review // Frontiers in Marine Science. V. 5. P. 178.
Maridet O., Ni X., 2013. A new cricetid rodent from the early Oligocene of Yunnan, China, and its evolutionary implications for early Eurasian cricetids // Journal of Vertebrate Paleontology. V. 33. № 1. P. 185–194.
Markova E., Bobretsov A., Borodin A., Rakitin S., Sibiryakov P., Smirnov N., Yalkovskaya L., Zykov S., 2020. The effects of population bottlenecks on dental phenotype in extant arvicoline rodents: implications for studies of the Quaternary fossil record // Quaternary Science Reviews. V. 228. 106045.
Markova E.A., Malygin V.M., Montuire S., Nadachowski A., Quéré J.-P., Ochman K., 2010. Dental variation in sibling species Microtus arvalis and M. rossiaemeridionalis (Arvicolinae, Rodentia): between-species comparisons and geography of morphotype dental patterns // Journal of Mammalian Evolution. V. 17. P. 121–139.
Markova E., Smirnov N., 2018. Phenotypic diversity arising from a limited number of founders: A study of dental variation in laboratory colonies of collared lemmings, Dicrostonyx (Rodentia: Arvicolinae) // Biological Journal of the Linnean Society. V. 125. P. 777–793.
Markova E.A., Sibiryakov P.A., Kartavtseva I.V., Lapin A.S., Morozkina A.V., Petukhov V.A., Tiunov M.P., Starikov V.P., 2019. What can an invasive species tell us about evolution? A study of dental variation in disjunctive populations of Microtus rossiaemeridionalis (Arvicolinae, Rodentia) // Journal of Mammalian Evolution. V. 26 (2). P. 267–282.
Nadachowski A., 1982. Late quaternary rodents of Poland with special reference to morphotype dentition analysis of voles. Warszawa; Krakow: Panstwowe wydawnictwo naukowe. 110 p.
Nekrutenko A., Makova K.D., Chesser R.K., Baker R.J., 1999. Representational difference analysis to distinguish cryptic species // Mol. Ecol. V. 8. P. 1235–1237.
Pantalacci S., Chaumot A., Benoît G., Sadier A., Delsuc F., Douzery E.J.P., Laudet V., 2008. Conserved Features and Evolutionary Shifts of the EDA Signaling Pathway Involved in Vertebrate Skin Appendage Development // Molecular Biology and Evolution. V. 25. № 5. P. 912–928.
Pavlova S.V., Tchabovsky A.V., 2011. Presence of the 54-chromosome common vole (Mammalia) on Olkhon Island (Lake Baikal, East Siberia, Russia), and the occurrence of an unusual X-chromosome variant // Comparative Cytogenetics. V. 5. № 5. P. 433–440.
Reig O.A., 1977. A proposed unified nomenclature for the enameled components of the molar teeth of the Cricetidae (Rodentia) // Journal of Zoology. V. 181. P. 227–241.
Rörig G., Börner C., 1905. Studien über das Gebiss mitteleuropäischer recenter Mäuse // Arbeit aus der Kaiserlichen Biologischen Anstalt für Land- und Forstwirtschaft. Berlin: Paul Parey-Springer. V. 5. № 2. P. 35–96.
Richardson D.M., Riccardi A., 2013. Misleading criticisms of invasion science: a field guide // Diversity and Distributions. V. 19. P. 1461–1467.
Rodrigues H.G., Renaud S., Charles C., Poul Y., Solé F., Aguilar J.-P., Michaux J., Tafforeau P., Headon D., Jernvall J., Viriot L., 2013. Roles of dental development and adaptation in rodent evolution // Nature Communications. V. 4. P. 2504.
Seppala M., Fraser G.J., Birjandi A.A., Xavier G.M., Cobourne M.T., 2017. Sonic hedgehog signaling and development of the dentition // Journal of Developmental Biology. V. 5. № 2. P. 6.
Sherpa S., Després L., 2021. The evolutionary dynamics of biological invasions: A multi-approach perspective // Evolutionary applications. V. 14 (6). P. 1463–1484.
Swaegers J., Mergeay J., Therry L., Larmuseau M.H.D., Bonte D., Stoks R., 2013. Rapid range expansion increases genetic differentiation while causing limited reduction in genetic diversity in a damselfly // Heredity. V. 111. P. 422–429.
Van Der Meulen A.J., 1973. Middle Pleistocene smaller mammals from the Monte Pegalia (Orvieto, Italy), with special reference to the phylogeny of Microtus (Arvicolidae, Rodentia) // Quaternaria. V. 17. P. 1–144.
Wellcome Sanger Institute, Arvicola amphibius genome assembly mArvAmp1.2, 2021. [Электронный ресурс]. Режим доступа: https://www.ncbi.nlm.nih.gov/assembly/GCF_903992535.2. Дата обновления: 15.05.2021. Дата последнего доступа: 25.04.2023.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал