БИОФИЗИКА, 2019, том 64, № 3, с. 457-467
МОЛЕКУЛЯРНАЯ БИОФИЗИКА
УДК 577.3
ИЗМЕНЕНИЕ УРОВНЕЙ ПОВРЕЖДЕНИЙ ДНК КЛЕТОК МЫШЕЙ
В ЗАВИСИМОСТИ ОТ АТМОСФЕРНЫХ ФАКТОРОВ
© 2019 г. Е.А. Кузнецова*, Н.П. Сирота*, Т.А. Зенченко* **
*Институт теоретической и экспериментальной биофизики РАН,
142290, Пущино Московской области, ул. Институтская, 3
**Инcтитут коcмичеcкиx иccледований PАН, 117997, Моcква, ул. Пpофcоюзная, 84/32
E-mail: kuzglu@rambler.ru
Поступила в редакцию 27.03.2019 г.
После доработки 27.03.2019 г.
Принята к публикации 28.03.2019 г.
Изучено изменение уровней повреждений ДНК по показателю %TDNA - процентной доли
ДНК в хвосте «кометы», определенной методом «комета-теста» в лейкоцитах крови, клетках
асцитной карциномы Эрлиха и лимфолейкоза Р388 мышей, в зависимости от атмосферных
факторов. Показано, что: повышенные значения %TDNA лейкоцитов регистрировались в узких
диапазонах атмосферной температуры (от -4°С до +3°С) и парциального содержания кислорода
воздуха (298-304 мг/л); по мере роста уровня геомагнитной активности увеличивался диапазон
вариаций
%TDNA лейкоцитов; изменения
%TDNA интактных или Х-облученных (4 Гр)
лейкоцитов и опухолевых клеток ассоциированы с изменениями температуры воздуха; выявлена
корреляция между температурой и %TDNA обоих видов интактных опухолевых клеток и
облученных клеток Р388. Можно полагать, что вариации %TDNA связаны с воздействием активных
форм кислорода, формирующихся в окружающей среде при температурах фазовых переходов воды.
Изменения базального уровня %TDNA клеток, по-видимому, могут приводить к изменению их
радиационного ответа.
Ключевые слова: comet assay, кровь мыши, клетки асцитной карциномы Эрлиха, клетки лимфолейкоза
Р388, рентгеновское излучение, температура воздуха
DOI: 10.1134/S0006302919030050
В исследованиях на уровне организма в рабо-
мого человека, так и условий окружающей среды,
не позволяет в настоящее время построить доста-
тах различных авторов многократно продемон-
точно полную непротиворечивую модель реак-
стрировано, что показатели сердечно-сосудистой
ции организма человека на комплекс факторов
системы человека изменялись под воздействием
земной и космической погоды [1]. В отличие от
факторов земной и космической погоды [1]. Так,
человека, лабораторные животные находятся в
на основании анализа динамики показателей ар-
стандартных стационарных условиях (местопо-
териального давления и микроциркуляции крови
ложение, освещенность, рацион питания). Они
у здоровых волонтеров сделано заключение, что
не подвергаются воздействиям, связанным с раз-
для климатического пояса Москвы и более север-
нообразием питания, передвижением, общением
ных широт наиболее биотропным атмосферным
с новыми индивидуумами, получением различ-
фактором являются вариации температуры воз-
ной информации и самовнушением. Несмотря на
духа, причем зависимость проявляется как син-
это, при проведении многочисленных экспери-
хронизация медленных волн физиологических и
ментов мы обнаружили, что в разные дни некото-
атмосферных показателей длительностью не-
рые показатели у одних и тех же клеточных попу-
сколько суток. [2-4]. К сожалению, значительная
ляций отличались друг от друга. Так, в экспери-
вариабельность амплитуд и даже знаков обнару-
ментах по определению уровней повреждений
женных эффектов, обусловленная, по-видимому,
ДНК лейкоцитов крови мышей методом «комета-
их зависимостью от текущего состояния, как са-
теста» было обнаружено, что изменялся базаль-
ный уровень повреждений ДНК в разные дни. В
Сокращения: АФК - активные формы кислорода, АКЭ -
асцитная карцинома Эрлиха, ГМА - геомагнитная актив-
этой связи было интересно выяснить, существует
ность.
ли зависимость наблюдающихся вариаций %TD-
457
458
КУЗНЕЦОВА и др.
NA (процентной доли ДНК в хвосте «кометы»,
Na2HPO4, 1,5 мМ KH2PO4, рН 7,2), содержащим
определенной методом «комета-теста») от атмо-
0,001 М ЭДТА.
сферных факторов, используя биологические мо-
Клетки асцитной карциномы Эрлиха (клетки
дели - мышиные клетки - при дополнительной
АКЭ), взятые на седьмые сутки после прививки
генотоксической нагрузке и без нее. В норме уро-
мышам-самцам неинбредной линии SHK, содер-
вень повреждений ДНК индивидуальных клеток
жащимся в стандартных условиях вивария ИТЭБ
отражает процессы как возникновения повре-
РАН, были получены, как указано в работах [8,9],
ждений, так и их репарации в ДНК, поскольку
и предоставлены для исследования С.П. Роман-
ДНК клеток постоянно подвергается спонтанной
ченко (лаборатория клеточной инженерии ИТЭБ
тепловой и гидролитической деградации, окисле-
РАН). Суспензию клеток трижды промывали
нию и неферментативному метилированию. Так,
раствором Хенкса, их количество подсчитывали в
только за счет апуринизации и окисления актив-
камере Горяева.
ными формами кислорода (АФК) в ДНК одной
Инокулят клеток лимфолейкоза Р388 (клетки
нормальной клетки человека в течение одного ча-
Р388) был получен от В.В. Шапошниковой из ла-
са возникает около 1000 поврежденных основа-
боратории окислительного стресса ИТЭБ РАН.
ний, что примерно равно количеству таких же по-
Клетки лимфолейкоза Р388 выращивали в брюш-
вреждений при гамма-облучении клеток в дозе
ной полости мышей-самцов линии DBA 2 (полу-
0,5 Гр. Как правило, эти повреждения ДНК по-
чены из НПП «Питомник лабораторных живот-
стоянно подвергаются репарации [5]. Повыше-
ных» ФИБХ РАН), прививали внутрибрюшинно
ние базального уровня повреждений ДНК лейко-
2 · 106 клеток на мышь. Через семь суток после
цитов может свидетельствовать об атаках гено-
прививки мышей умерщвляли методом церви-
токсических физико-химических факторов,
кальной дислокации, извлекали клетки из брюш-
например, АФК, что может сказаться на функци-
ной полости, промывали средой RPMI 1640 (Sig-
онировании ряда систем организма, в том числе
ma, США) и подсчитывали в камере Горяева.
сердечно-сосудистой системы. Поскольку воз-
Для приготовления агарозных слайдов ис-
действие редкоионизирующего излучения можно
пользовали цельную кровь мышей, разведенную
рассматривать как атаку АФК, где основным по-
содержащим ЭДТА фосфатно-солевым буфером
вреждающим фактором являются ОН-радикалы
в шесть раз; клетки АКЭ разводили раствором
[6], то в качестве такой генотоксической нагрузки
мы использовали воздействие рентгеновского из-
Хенкса, клетки Р388 - средой RPMI 1640 до кон-
лучения на клетки. Широкий спектр поврежде-
центрации 1 · 106 кл/мл.
ний ДНК - одно- и двунитевые разрывы, реали-
Работу с животными проводили в соответ-
зованные в разрывы щелочелабильные (апурино-
ствии с рекомендациями Комиссии по биологи-
вые/апиримидиновые) сайты
- позволяет
ческой безопасности и биоэтике ИТЭБ РАН.
выявить щелочная версия метода «комета-теста»
Облучение. Клетки крови, АКЭ или Р388 были
(метод ДНК-комет, comet assay) [7]. В этой работе
облучены в составе агарозных слайдов на льду.
методом «комета-теста» оценивали уровни по-
Облучение проводили на рентгеновской установ-
вреждений ДНК лейкоцитов крови нормальных
ке РУТ-250-15-1 в ЦКП «Источники излучений»
мышей и клеток асцитной карциномы Эрлиха
ИБК РАН (Пущино Московской области) при
или мышиного лимфолейкоза Р388 до и после
мощности дозы
1,12 Гр/мин, напряженности
воздействия рентгеновского излучения (Х-облу-
200 кВ, силе тока 20 мА, фильтрах 1 мм Al и 1 мм
чения) на эти клетки in vitro.
Cu, фокусном расстоянии 37 см. Дозиметрию
Цель работы: выявить, как соотносятся уровни
проводили по методу Фрике в присутствии бен-
повреждений ДНК интактных или Х-облученных
зойной кислоты и с помощью дозиметра VA-J-18
лейкоцитов периферической крови или опухоле-
(RFT Messelektronik, Германия). Слайды сразу
вых клеток мышей с изменениями показателей
после облучения помещали в охлажденный лизи-
атмосферного давления, температуры воздуха,
рующий раствор.
его относительной влажности, парциального со-
Метод
«комета-теста». Вначале предметные
держания кислорода и уровня геомагнитной ак-
стекла погружали в раствор 1%-й агарозы и высу-
тивности.
шивали. На эти стекла наносили слой 1%-й агаро-
зы и инкубировали в холодильнике до ее затверде-
вания (5-7 мин). Разведенную кровь или суспен-
МАТЕРИАЛЫ И МЕТОДЫ
зию клеток смешивали с равным объемом 1%-й
Клетки. Использовали мышей-самцов неин-
легкоплавкой агарозы (Sigma, США), расплавлен-
бредной линии SHK, содержащихся в стандарт-
ной при 70°С в фосфатно-солевом буфере, содер-
ных условиях вивария ИТЭБ РАН. Аликвоты
жащем ЭДТА, и инкубируемой при 37°С (конечная
крови (5 мкл), взятые при надрезании кончика
температура получаемой смеси 20-22°С). Смесь
хвоста, отбирали в пробирки с фосфатно-соле-
(15 мкл) наносили на приготовленный агарозный
вым буфером (136,7 мМ NaCl, 2,7 мМ KCl, 8,1 мМ
слой. После охлаждения и застывания содержащей
БИОФИЗИКА том 64
№ 3
2019
ИЗМЕНЕНИЕ УРОВНЕЙ ПОВРЕЖДЕНИЙ ДНК КЛЕТОК МЫШЕЙ
459
клетки агарозы на ее поверхность наносили новый
Весовое (парциальное) содержание кислорода
слой 0,5%-й легкоплавкой агарозы и инкубирова-
в атмосферном воздухе - Р(О2) было рассчитано
ли в холодильнике до затвердевания. Слайды под-
по формуле Клапейрона:
вергали воздействию рентгеновского излучения и
затем помещали в лизирующий раствор (2,5 М
P
атм
-
e
P
атм
−
e
3
P
=
⋅
0,232
=
80,83
(г/м
),
(1)
NаС1, 0,1 М ЭДТА, 0,01 М трис-НСl, рН 10, 1%
R⋅T
T
тритона Х-100) при 4°С на ночь. Затем следовала
где Pатм и e - соответственно атмосферное давле-
процедура денатурации ДНК - слайды перемеща-
ние и абсолютная влажность воздуха (в гПа), Т -
ли на 20 мин в щелочной раствор «А» (0,3 М
температура воздуха в К, R - газовая постоянная
NаОН, 0,001 М ЭДТА, рН > 13). Для всех экспери-
для сухого воздуха, равная 287 Дж/(кг·К), 0,232 -
ментов процедуру денатурации проводили при
весовая доля кислорода в сухом воздухе.
4°С, за исключением экспериментов с кровью, по-
Для оценки возможной зависимости значений
лученной из одной мыши, а именно: денатурацию
биологических показателей от атмосферных и
ДНК проводили как при 4°С, так и при 20°С. Далее
геомагнитных факторов использовали методы
слайды переносили в электрофоретическую каме-
корреляционного, регрессионного и дисперси-
ру SE-1/S-1N (ООО
«Компания Хеликон»,
онного анализов. Поскольку для большинства
Москва) и подвергали электрофорезу в свежей
проанализированных выборочных распределе-
порции раствора «А» в течение 20 мин при 4°С, при
ний биологических показателей не выполнялась
объеме буфера 250 мл, напряжении 27 В, силе тока
гипотеза о нормальности распределения (соглас-
260-270 мА (напряженность электрического поля
но критерию Жарка-Берра), были использованы
2 В/см). Все эксперименты проводили с использо-
непараметрические методы оценки, а именно:
ванием одной и той же электрофоретической ка-
расчет рангового коэффициента корреляции
меры. После электрофореза слайды промывали
Спирмена и критерий Вилкоксона для оценки
дистиллированной водой и окрашивали в течение
различия выборок. Поскольку большинство об-
1 ч в фосфатно-солевом буфере, содержащем
наруженных зависимостей имели нелинейный
2,0 мкг/мл этидиум бромида. Слайды анализиро-
характер, был использован коэффициент детер-
вали под флуоресцентным микроскопом
минации (R2), полученный при аппроксимации
ЛЮМАМ И-3 («ЛОМО», Санкт-Петербург). За-
данной зависимости полиномом второй степени.
хват изображений проводили цифровым фотоап-
паратом CoolPix 995 (Nikon, Япония) с последую-
щей передачей изображений в компьютер. Обра-
РЕЗУЛЬТАТЫ
ботку изображений выполняли с помощью
специализированного программного обеспечения,
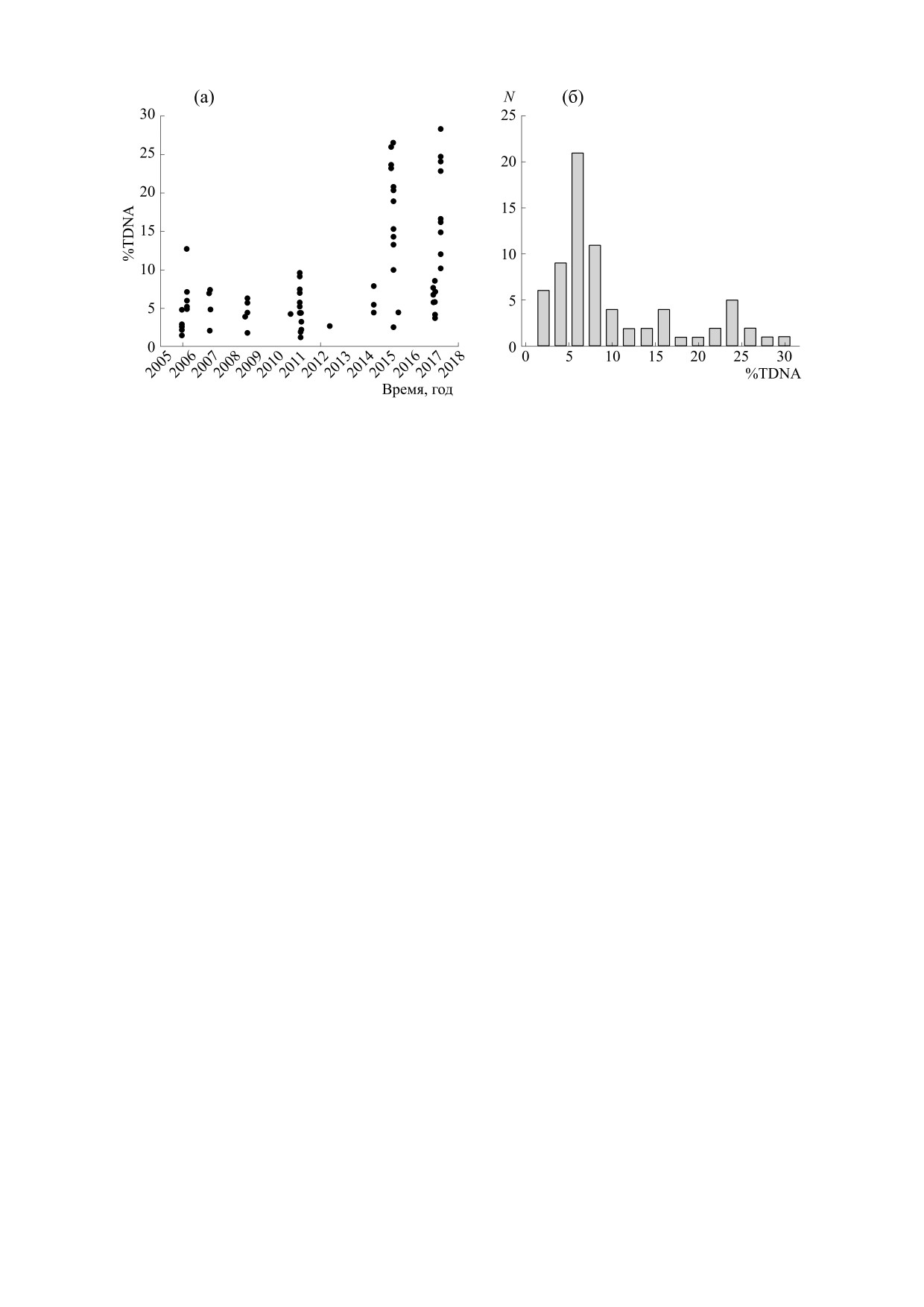
Вначале мы проанализировали изменения
где реализованы алгоритмы расчета стандартных
уровней повреждений ДНК лейкоцитов из крови
интактных мышей. Для анализа был использован
параметров «комет» [10]. Для оценки уровня по-
массив значений %TDNA лейкоцитов, получен-
вреждений ДНК использовался параметр, пред-
ный в экспериментах 2005-2017 гг., и каждой ве-
ставляющий процент ДНК в хвосте «кометы» (per
личине %TDNA соответствовали среднесуточные
cent of DNA in a comet tail - %TDNA). Для каждой
значения атмосферного давления, температуры
экспериментальной точки анализировали по три
воздуха, относительной влажности, содержания
слайда, вычисляя средние значения %TDNA и фо-
кислорода и уровня ГМА, оцененного по Кр-ин-
тографируя не менее чем по 50 клеток на слайд, со-
гласно рекомендациям, сделанным в работе [11].
дексу. На рис. 1 показаны распределения %TDNA
из использованного массива данных по датам
Метеорологические показатели и анализ результа-
экспериментов и по наблюдаемым значениям. Из
тов. Данные значений метеорологических парамет-
представленных 73 значений %TDNA 47 значе-
ров были получены с сайта https:// rp5.ru/ по г. Сер-
ний имели величину менее 10, и их распределение
пухов (30 км от места проведения экспериментов),
на гистограмме подчинялось нормальному зако-
по уровню геомагнитной активности - с сайта ftp://
ну (рис. 1б). В то же время 25 значений имели
ftp.ngdc.noaa.gov/STP/GEOMAGNETIC_DATA/
большую величину, и на графике выборочного
INDICES/KP_AP.
распределения их можно рассматривать либо как
«тяжелый хвост», либо как второе колоколооб-
В расчетах были использованы среднесуточ-
разное распределение с максимумом в районе 25.
ные значения, полученные вычислением средне-
Видно также, что значения %TDNA > 10 наблю-
го арифметического для атмосферного давления
дались неоднократно, а именно в четырех сериях
(Ратм), температуры (Татм), относительной влаж-
экспериментов (04.02.2015, 04.03.2015, 13.03.2015
ности (RH, Relative Humidity) и суммированием
и 30.03.2017). Если весь массив данных разбить по
для Кр-индекса, т.е. планетарного индекса гео-
месяцам проведения наблюдений, то достаточно
магнитной активности (ГМА), восьми трехчасо-
большие, сопоставимые между собой по величи-
вых показателей.
не выборки наблюдались для следующих перио-
БИОФИЗИКА том 64
№ 3
2019
460
КУЗНЕЦОВА и др.
Рис. 1. Распределение значений %TDNA лейкоцитов крови интактных мышей из использованного массива данных по
датам экспериментов (а) и по наблюдаемым величинам (б).
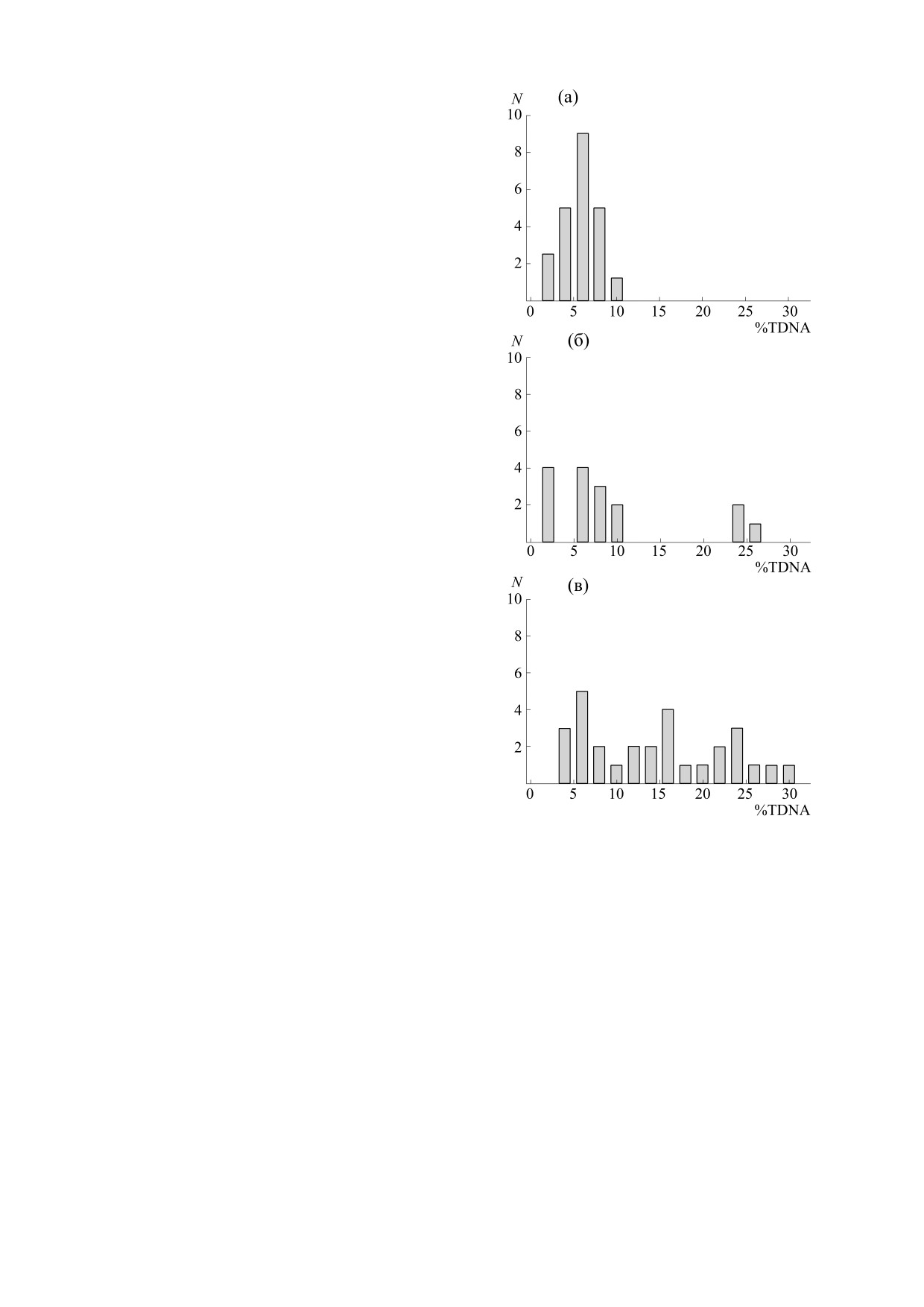
дов года: (а) - сентябрь-декабрь (22 измерения),
по величинам относительной влажности значе-
(б) - февраль (16 измерений), (в) - март (29 изме-
ния %TDNA > 10 наблюдались как при высоких,
рений). Распределения выборочных значений,
так и при низких значениях RH (рис. 3г). Необхо-
полученных для данных периодов, приведены на
димо отметить, что, как видно из формулы (1),
рис. 2. Видно, что в измерениях разных лет, про-
расчетное значение парциального содержания
веденных в сентябре-декабре, не встречались по-
кислорода в воздухе в наибольшей степени зави-
вышенные значений %TDNA, в феврале такие
село от соответствующей ему величины темпера-
значения составляли 19%, а в марте - 62%.
туры, в меньшей степени - от атмосферного дав-
ления и крайне слабо - от значения относитель-
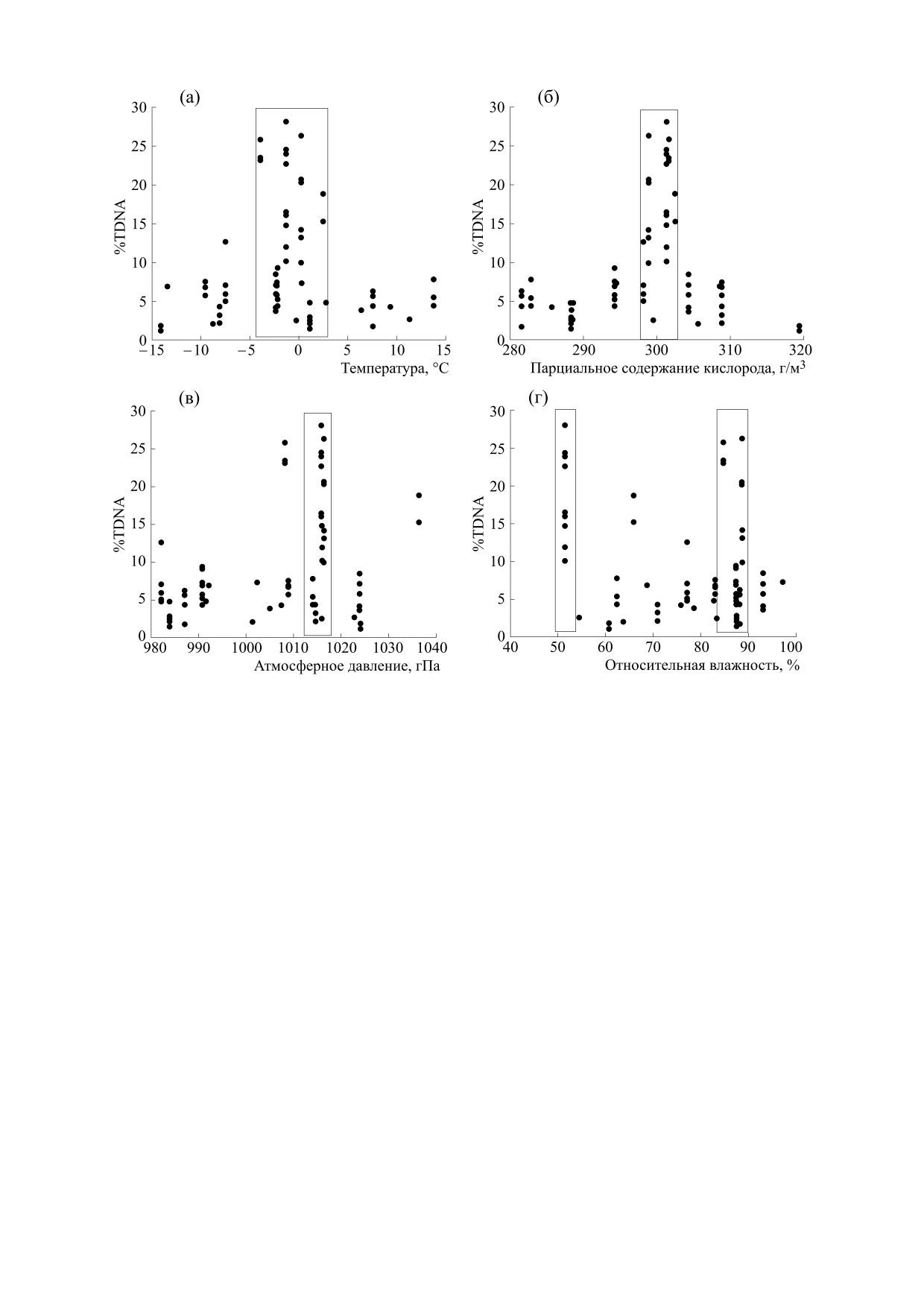
На рис. 3 показаны распределения %TDNA по
ной влажности. Можно полагать, что повы-
синхронным значениям температуры воздуха
шенные значения %TDNA зависят от показателя
(рис. 3а), парциального содержания кислорода в
Р(О2), реализуясь в очень узком диапазоне его
воздухе (рис.
3б), атмосферного давления
(рис. 3в) и относительной влажности (рис. 3г). Из
значений, а менее выраженные неоднородности
рис. 3а видно, что все (за исключением одного)
распределений %TDNA от Татм и Ратм являются
повышенные относительно фона значения %TD-
следствием функциональной связи этих атмо-
NA (%TDNA > 10) располагались в узком диапа-
сферных показателей со значениями Р(О2). То
зоне температур (-4°С - +3°С), составляющем
есть изменения %TDNA лейкоцитов крови мы-
менее четверти всего наблюдаемого диапазона.
шей были ассоциированы с вариациями темпера-
Аналогичная картина наблюдалась на рис. 3б: все
туры воздуха и парциального содержания кисло-
повышенные значения %TDNA располагались в
рода в окружающей среде.
диапазоне концентраций Р(О2) 298-304 мг/л, от-
Поскольку повышенные значения %TDNA
носительная ширина которого составляла всего
наблюдались в феврале-марте (рис. 2) и именно
15% от диапазона значений Р(О2), встретившихся
на этот период приходился максимум внутриго-
в эксперименте. С учетом достаточно равномер-
дового распределения уровня ГМА, чтобы прове-
ного распределения значений %TDNA < 10 веро-
рить возможную зависимость показателя
ятность случайного одновременного попадания
%TDNA от уровня ГМА было построено распре-
всех 25 повышенных значений, наблюдавшихся в
деление выборочных значений %TDNA от Кр-ин-
эксперименте, в интервал значений Р(О2) шири-
декса (рис. 4). Расчет рангового коэффициента
ной
0,15 составляла крайне малую величину
корреляции между %TDNA и соответствующими
(p ≈ 0,1524). В меньшей степени тенденция груп-
значениями Кр-индекса давал величину rs = 0,318
пирования повышенных значений %TDNA на-
с уровнем статистической значимости p = 0,006,
блюдалась на распределении по величинам Ратм.
однако, как и в случае атмосферных факторов,
Как видно из рис. 3в, пять показателей %TDNA >
вид зависимости был более сложным, чем прямая
10 не попадали в узкий интервал значений атмо-
пропорциональность, выявляемая методом кор-
сферного давления. В случае же распределения
реляционного анализа. Из рис. 4 видно, что при
БИОФИЗИКА том 64
№ 3
2019
ИЗМЕНЕНИЕ УРОВНЕЙ ПОВРЕЖДЕНИЙ ДНК КЛЕТОК МЫШЕЙ
461
низком уровне ГМА высокие значения %TDNA
не наблюдались, а по мере роста возмущенности
геомагнитной обстановки увеличивался и диапа-
зон значений %TDNA: чем выше было текущее
значение Кр-индекса, тем выше располагалась
верхняя грань наблюдаемых значений %TDNA.
Чтобы выяснить, как изменяются уровни по-
вреждений ДНК клеток при генотоксической на-
грузке, в качестве источника АФК использовали
воздействие рентгеновского излучения на клетки
in vitro. Для того чтобы избежать индивидуальных
различий, обычно наблюдаемых в группе живот-
ных, все эксперименты были проведены на кро-
ви, полученной от одной мыши в разные дни. На
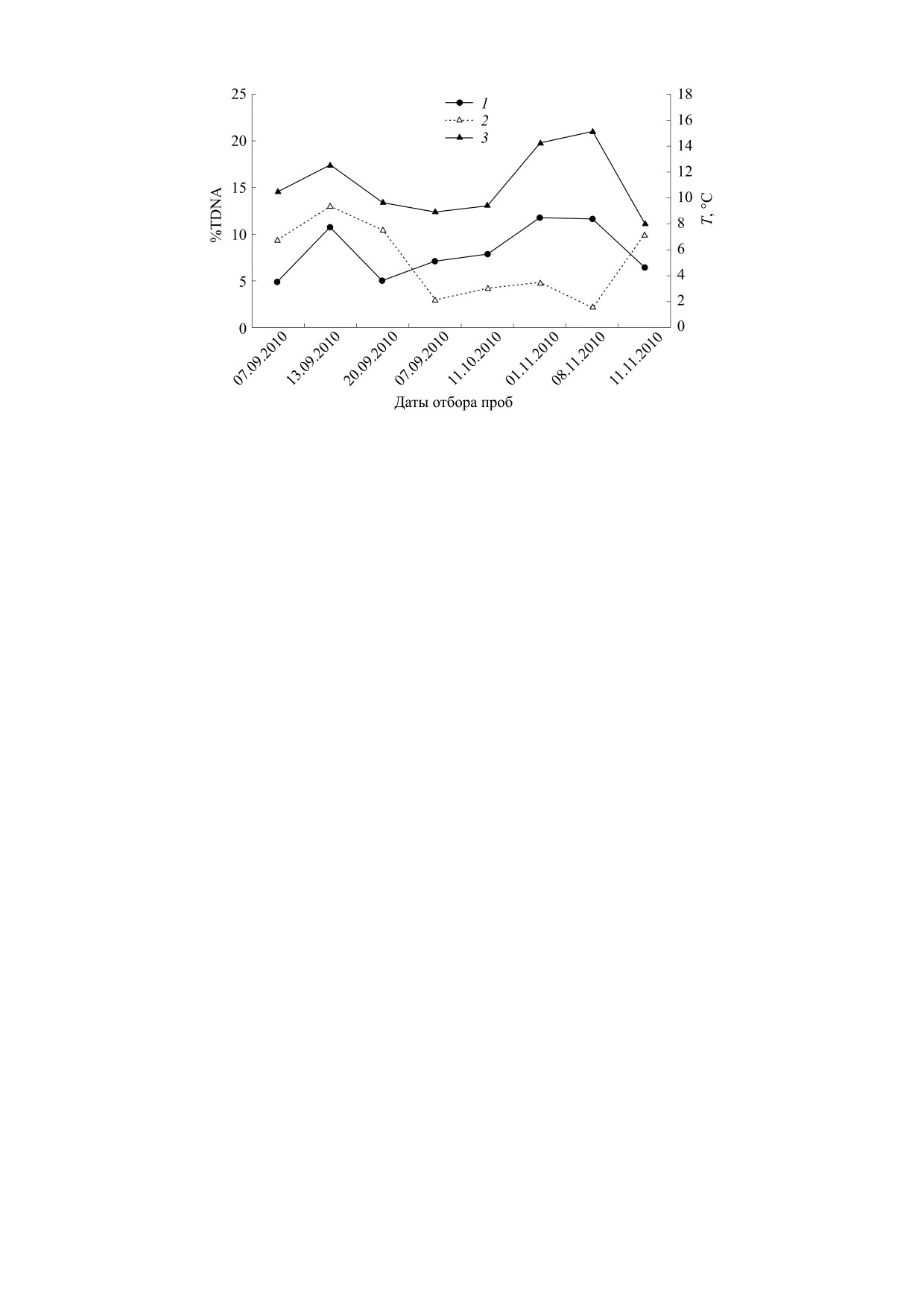
рис. 5 показаны изменения %TDNA облученных
в дозе 4 Гр лейкоцитов периферической крови
мыши и соответствующих среднесуточных тем-
ператур воздуха в дни отбора проб крови. Уровень
повреждений ДНК представлен как разность
средних значений %TDNA облученных и необлу-
ченных лейкоцитов. Показаны данные двух вари-
антов экспериментов, в которых проводилась де-
натурация ДНК (один из этапов метода «комета-
теста»): при стандартной температуре 4°С и при
20°С. Различия значений %TDNA для необлучен-
ных (контрольных) клеток при разных темпера-
турах денатурации были статистически недосто-
верны. Видно, что при одной и той же дозе облу-
чения в случае денатурации при 20°С наблюдался
более высокий уровень повреждений ДНК, чем
при 4°С, и что в дни, когда температура воздуха не
превышала +4°С, показатель %TDNA увеличи-
вался. Таким образом, изменение значений
%TDNA облученных лейкоцитов было ассоции-
ровано с изменением температуры воздуха. Не-
смотря на то, что среди проанализированных ве-
личин %TDNA у необлученных лейкоцитов не
встречалось значений выше 10, эти данные вхо-
дили в массив, для которого в приведенном выше
исследовании была обнаружена зависимость
%TDNA от температуры воздуха и парциального
содержания кислорода.
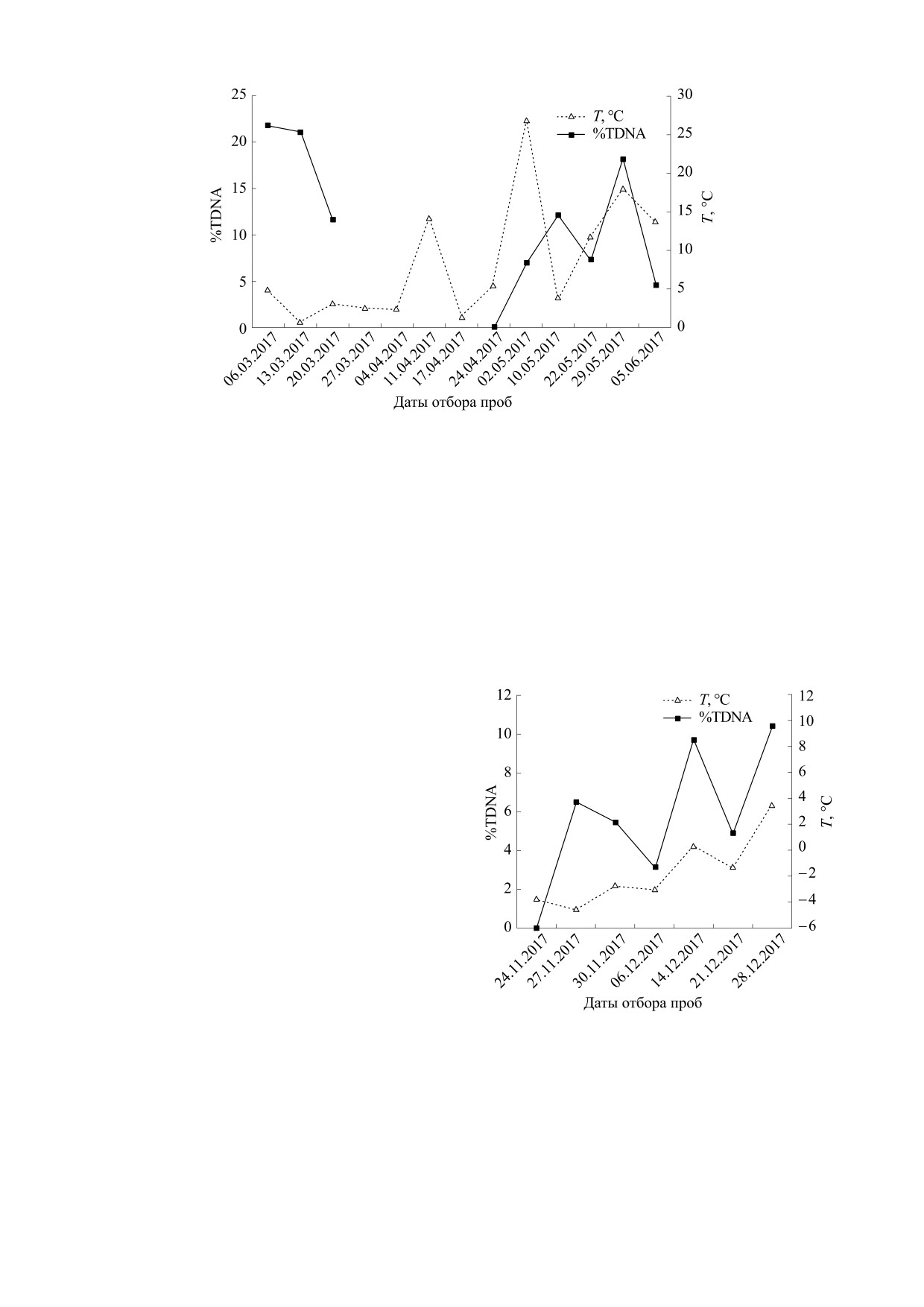
На рис. 6 показаны изменения %TDNA клеток
Рис.
2.
Распределения выборочных значений
АКЭ и соответствующих среднесуточных темпе-
%TDNA лейкоцитов крови интактных мышей в раз-
ратур воздуха в дни отбора асцита. Уровень по-
личные сезоны года: (а) - сентябрь-декабрь, (б) -
вреждений ДНК представлен как разность сред-
февраль, (в) - март.
них значений %TDNA облученных в дозе 4 Гр и
необлученных клеток. Видно, что эти изменения
%TDNA ассоциированы с изменениями темпера-
вреждений ДНК представлен как разность сред-
туры воздуха. При этом изменения температуры
них значений %TDNA облученных в дозе 4 Гр и
несколько опережали изменения %TDNA. Для
необлученных клеток. Видно, что изменения
%TDNA необлученных клеток АКЭ была обнару-
%TDNA ассоциированы с изменениями темпера-
жена статистически значимая корреляция с син-
туры воздуха. Выявлена статистически значимая
хронными значениями атмосферной температу-
ры воздуха (rs = 0,577, p < 0,05).
корреляция между температурой воздуха и уров-
нями повреждений ДНК интактных (rs = -0,883,
На рис. 7 показаны изменения %TDNA клеток
p < 0,05) и облученных (rs = -0,847, p < 0,05) кле-
Р388 и соответствующих среднесуточных темпе-
ратур воздуха в дни отбора асцита. Уровень по-
ток, а также температурой воздуха и разницей
БИОФИЗИКА том 64
№ 3
2019
462
КУЗНЕЦОВА и др.
Рис. 3. Распределение значений %TDNA лейкоцитов крови интактных мышей по синхронным значениям температу-
ры воздуха (а), парциального содержания кислорода (б), атмосферного давления (в) и относительной влажности воз-
духа (г).
%TDNA между облученными и необлученными
спектрально-временную картину вариаций маг-
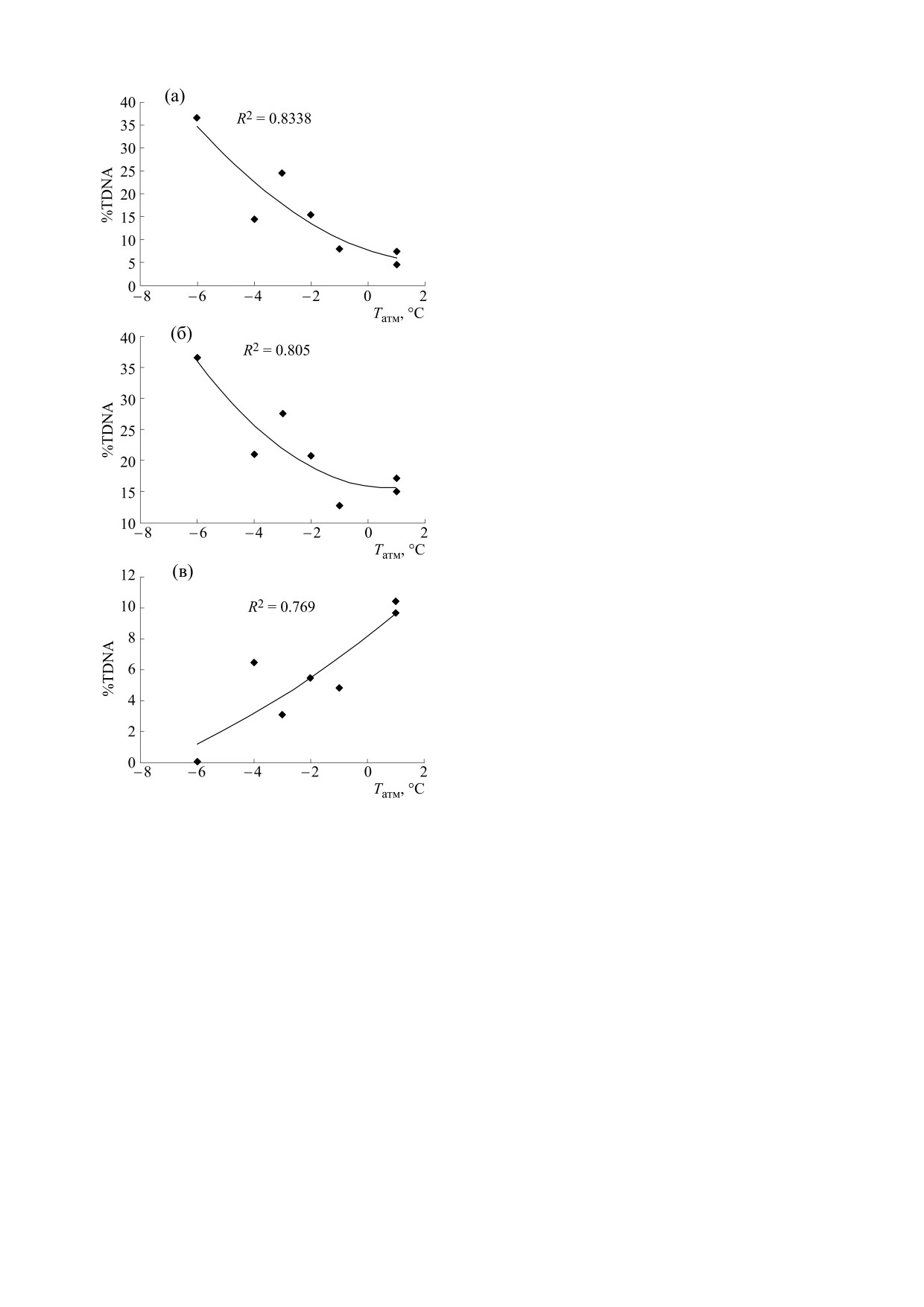
клетками (rs = 0,739, p < 0,1). На рис. 8 указаны
нитного поля и атмосферного давления [12].
значения объясненной дисперсии R2 при аппрок-
Вначале мы сопоставляли изменения %TDNA
симации зависимостей полиномом второй степе-
лейкоцитов крови с изменениями атмосферных
ни, что свидетельствует о зависимости %TDNA
факторов у интактных мышей. В предыдущих и
от температуры воздуха как интактных, так и об-
настоящих исследованиях в наших условиях экс-
лученных клеток.
перимента в большинстве случаев значения
%TDNA лейкоцитов не превышали 10% как у мы-
шей, так и у человека [13], поэтому все значения
ОБСУЖДЕНИЕ
%TDNA, равные или ниже 10%, считали фоновы-
Изучая влияние природных факторов - атмо-
ми/базальными (рис. 1). Было обнаружено, что
сферных и геомагнитных - на мышей, содержа-
не наблюдается какой-либо зависимости
щихся в виварии, следует отметить, что из-за по-
%TDNA лейкоцитов мышей от влажности возду-
стоянной вентиляции в помещениях вивария
ха и атмосферного давления (рис. 3). Все повы-
идет активный обмен воздуха с внешней средой.
шенные относительно фона значения %TDNA
То есть на животных оказывали влияние такие
располагались в окрестности нуля температуры
факторы как состав и влажность воздуха улицы,
(-4°С - +3°С), то есть в интервале температур,
магнитное поле, атмосферное давление. Как бы-
при которых происходят фазовые переходы воды.
ло показано ранее, наличие здания не изменяло
Ранее высказывалась гипотеза о существовании
БИОФИЗИКА том 64
№ 3
2019
ИЗМЕНЕНИЕ УРОВНЕЙ ПОВРЕЖДЕНИЙ ДНК КЛЕТОК МЫШЕЙ
463
механохимических реакций радикальной диссо-
циации воды: в отличие от фотолиза или радио-
лиза воды диссоциация воды предполагалась воз-
можной и в случаях, когда плотность энергии ак-
тивации заведомо меньше необходимой для
диссоциации отдельных молекул воды [14]. Как
считают авторы, появление H2O2 при фазовых
превращениях воды («вода-лед-вода» и «вода-
пар-вода») можно объяснить именно механохи-
мическими реакциями радикальной диссоциа-
ции воды, что также было подтверждено ими экс-
периментально. Было показано, что подобно то-
му, как это происходит при расплавлении и
отвердевании полимеров, после фазовых перехо-
дов и в талой, и в конденсированной воде образу-
ются ОН-радикалы и Н-атомы и накапливается
Рис. 4. Распределение значений параметра %TDNA
H2O2. Это осуществляется и в отсутствие воздей-
лейкоцитов крови интактных мышей по синхронным
ствия света. В экспериментах содержание H2O2 в
значениям уровня геомагнитной активности.
талой воде еще некоторое время после таяния мо-
нотонно увеличивалось, достигая насыщения че-
рез несколько часов, что свидетельствовало о зна-
ми/апиримидиновыми или содержащими моди-
чительной продолжительности жизни радикалов
фицированные основания сайтами ДНК. Следует
воды в клетках структур воды - кластерах. Суще-
отметить, что все повышенные относительно фо-
ствование кластеров водяного пара в атмосфере
на %TDNA располагались также в узком диапазо-
не вызывает сомнений, имеются предположения,
не значений парциального содержания кислоро-
что именно атмосферные кластеры воды способ-
да (298-304 мг/л, рис. 3). По-видимому, уровень
ствуют перемещению веществ, переносимых па-
H2O2, сформировавшийся в окружающей среде
рами воды из водных растворов [14]. Возможно,
при атмосферных температурах, при которых
обнаруженное нами повышение значений
происходят фазовые переходы воды, соотносился
%TDNA лейкоцитов связано с воздействием на
с определенными концентрациями кислорода в
них АФК, формирующихся при фазовых перехо-
воздухе, тем более что расчетное значение парци-
дах воды в окружающей среде: АФК при актив-
ального содержания кислорода в наибольшей
ной вентиляции проникали в помещения вива-
степени зависело от соответствующей ему темпе-
рия и посредством газообмена в легких могли
ратуры воздуха, как видно из формулы (1).
воздействовать на клетки крови мышей. При
Вариации %TDNA наблюдались и при изме-
среднесуточных температурах воздуха, находя-
нении ГМА: при низком уровне ГМА высоких
щихся в узком диапазоне -4°С-+3°С, у одной ча-
значений %TDNA не наблюдалось, а по мере ро-
сти животных %TDNA лейкоцитов находился в
ста уровня ГМА увеличивался и диапазон разбро-
пределах нормы, тогда как у другой части наблю-
са значений %TDNA (рис. 4). Однако высокие
дались существенно более высокие %TDNA. Об-
значения %TDNA наблюдались и в дни с низким
наружено, что дням, в которые регистрировали
уровнем ГМА (Кр -индекс < 20). Если весь массив
максимальные значения %TDNA, предшествова-
данных разбить по месяцам проведения наблюде-
ло от 5 до 10 дней, когда среднесуточные темпера-
ний, то наибольший разброс значений %TDNA
туры воздуха находились в интервале -4°С-
приходился на март (рис. 2). В Московской обла-
+3°С, то есть высокие уровни повреждений ДНК
сти в этот период температура воздуха часто нахо-
лейкоцитов у мышей наблюдались в тех случаях,
дилась в границах, соответствующих фазовым пе-
когда формирующиеся в окружающей среде по-
реходам воды. Исходя из представленного, мож-
вышенные уровни АФК воздействовали на жи-
но полагать, что повышение уровня повреждений
вотных в течение некоторого времени. Выявляе-
ДНК в основном связано с увеличением уровня
мые щелочной версией метода «комета-теста»
АФК в воздухе, опосредованного изменениями
(рН > 13) повреждения ДНК обусловлены либо
температуры окружающей среды.
уже существующими разрывами ДНК, либо реа-
лизованными в разрывы щелочелабильными
Чтобы выяснить, как изменяются уровни по-
(апуриновыми/апиримидиновыми) сайтами: из-
вреждений ДНК лейкоцитов при дополнитель-
вестно, что в щелочных условиях эти сайты пре-
ном воздействии на них АФК, следующие экспе-
вращаются в разрывы ДНК [7,15,16]. Поэтому и
рименты были проведены в течение октября-но-
регистрируемые нами повышенные уровни по-
ября на пробах крови, полученных от одной
вреждений ДНК лейкоцитов мышей могут быть
мыши в разные дни, поскольку исследование на
обусловлены как разрывами, так и апуриновы-
клетках одного и того же животного позволяет из-
БИОФИЗИКА том 64
№ 3
2019
464
КУЗНЕЦОВА и др.
Рис. 5. Изменения %TDNA лейкоцитов периферической крови мыши и среднесуточной температуры воздуха в дни
отбора проб крови. Уровень повреждений ДНК представлен как разность средних значений %TDNA облученных и
необлученных лейкоцитов крови: 1 - денатурация при 4°С; 2 - денатурация при 20°С; 3 - температура воздуха, °С.
бежать индивидуальных различий, обычно на-
Поскольку лейкоциты крови представлены в
блюдаемых в группе животных. Обнаружили, что
подавляющем большинстве зрелыми неделящи-
при одной и той же дозе облучения (4 Гр) в усло-
мися клетками, то следующие эксперименты бы-
виях денатурации при 20°С наблюдался более вы-
ли проведены на делящихся мышиных клетках
сокий уровень повреждений ДНК, чем при 4°С
лимфолейкоза Р388 и клетках АКЭ. Так, напри-
(рис. 5), что согласуется с литературными данны-
мер, на седьмые сутки после перевивки клетки
ми [17]. Поскольку процесс реализации щелоче-
АКЭ различались по стадиям клеточного цикла
лабильных сайтов в разрывы ДНК зависел от тем-
следующим образом: количество клеток в стадии
пературы [15], то очевидно, что при 20°С допол-
SubG0 в контроле колебалось в пределах 7% от
нительно выявлялись щелочелабильные сайты,
общего числа клеток; в фазе G0/G1 регистриро-
не реализовавшиеся в разрывы при 4°С. Уровни
валось примерно 62% клеток, тогда как в фазах S
повреждений ДНК для необлученных клеток ста-
и G2/M - по 15% и 16% соответственно [18]. В на-
тистически не различались при разных темпера-
ших экспериментах была обнаружена статистиче-
турах денатурации ДНК, что позволяет считать,
ски значимая корреляция %TDNA необлученных
что регистрируемые при 20°С щелочелабильные
клеток АКЭ с синхронными значениями атмо-
сайты возникли в ДНК в результате воздействия
сферной температуры воздуха (rs
=
0,577,
ионизирующей радиации, и эти сайты выявляют-
p < 0,05). Индуцированные облучением измене-
ся при более высокой температуре обработки.
ния %TDNA у этих клеток, так же как и у лейко-
Индуцированные
облучением
изменения
цитов мышей, были ассоциированы с изменени-
%TDNA были ассоциированы с изменениями
ями среднесуточной температуры, хотя корреля-
температуры воздуха; вариации %TDNA по-раз-
ция между значениями %TDNA и температурой
ному соотносились с температурой: в интервале
воздуха выявлена не было (рис. 6). При пониже-
выше (данные, полученные в период 07.09.2010 -
нии температуры воздуха у облученных клеток
05.10.2010) и примерно равными или ниже +4°С
наблюдалось увеличение %TDNA. При этом из-
(данные, полученные в период
05.10.2010-
менения температуры воздуха несколько опере-
11.11.2010) (рис. 5). С понижением температуры
жали изменения %TDNA. Похожие изменения
%TDNA наблюдались и для клеток Р388 (рис. 7,
воздуха до
+4оС наблюдалось увеличение
8). Однако, в отличие от клеток АКЭ, у клеток
%TDNA при обеих температурах денатурации,
Р388 была выявлена статистически значимая кор-
хотя корреляции между значениями %TDNA и
реляция между температурой воздуха и %TDNA
среднесуточными температурами выявлено не
не только у необлученных (rs = -0,883, p < 0,05),
было. Из представленного материала видно, что
изменения индуцированных облучением уровней
но и у облученных (rs = -0,847, p < 0,05) клеток, а
повреждений ДНК лейкоцитов мышей соотноси-
также между температурой и разностью значений
лись с изменениями температуры воздуха.
%TDNA между облученными и необлученными
БИОФИЗИКА том 64
№ 3
2019
ИЗМЕНЕНИЕ УРОВНЕЙ ПОВРЕЖДЕНИЙ ДНК КЛЕТОК МЫШЕЙ
465
Рис. 6. Изменения %TDNA клеток АКЭ и среднесуточной температуры воздуха в дни отбора асцита. Уровень повре-
ждений ДНК представлен как разность средних значений %TDNA облученных и необлученных клеток АКЭ.
клетками (rs = 0,739, p < 0,1). Здесь необходимо
мость становится более выраженной, когда на
подчеркнуть, что эксперименты с клетками Р388
клетки оказывают дополнительное воздействие
были проведены в дни, когда температура воздуха
АФК. Несмотря на то что повышенные уровни
находилась в интервале -4°С- +4°С, по сравне-
повреждений ДНК клеток ассоциированы также
нию с клетками АКЭ, когда наблюдались и более
с определенным количеством кислорода в возду-
высокие атмосферные температуры. Кривая из-
хе и высоким уровнем ГМА, температура воздуха,
менений %TDNA у облученных клеток Р388 бо-
являющаяся результирующим показателем цело-
лее точно соответствовала изменениям темпера-
го ряда атмосферных явлений, по всей видимо-
туры воздуха по сравнению с клетками АКЭ, что,
возможно, обусловлено также и различиями кле-
точных популяций: клетки Р388 представлены
двумя типами клеток [19], в то время как клетки
АКЭ представляют собой в высшей степени де-
дифференцированные элементы довольно разно-
образной величины [20]. Некоторые авторы вы-
деляют не менее трех субпопуляций АКЭ [18].
Как соотносятся по радиочувствительности раз-
ные субклеточные популяции у изучаемых опухо-
левых клеток - неизвестно. По сравнению с эти-
ми опухолевыми клетками лейкоциты мыши, ко-
торые в большинстве своем являются лимфо-
цитами, в свою очередь представлены в виде трех
различных по радиочувствительности субпопуля-
ций с D0(1) = 0,36 Гр, D0(2) = 0,74 Гр и D0(3) =
= 4,37 Гр; на радиорезистентную фракцию
(D0 = 4,37 Гр) приходится примерно 30% клеток
[21]. Таким образом, во всех случаях изучаемые
клетки были представлены в той или иной степе-
ни геторогенными популяциями. Тем не менее
видно, что вариации %TDNA интактных или об-
лученных опухолевых клеток АКЭ и Р388, так же
Рис. 7. Изменения %TDNA клеток лимфолейкоза
как и у лейкоцитов, ассоциированы с изменения-
Р388 и среднесуточной температуры воздуха в дни от-
ми температуры воздуха. Сравнивая зависимости
бора асцита. Уровень повреждений ДНК представлен
%TDNA интактных и облученных клеток от вари-
как разность средних значений %TDNA облученных
аций температуры, можно видеть, что эта зависи-
и необлученных клеток АКЭ.
БИОФИЗИКА том 64
№ 3
2019
466
КУЗНЕЦОВА и др.
клеток мышей ассоциированы с изменениями
температуры воздуха, что, по-видимому, обу-
словлено воздействием активных форм кислоро-
да, формирующихся в диапазоне температур -
4°С-+3°С в окружающей среде. Дополнительное
воздействие АФК увеличивало %TDNA этих кле-
ток, и индуцированные облучением уровни по-
вреждений ДНК также были ассоциированы с из-
менениями температуры воздуха. Изменения ба-
зального уровня повреждений ДНК в этих
клетках, по-видимому, могут приводить к изме-
нению их радиационного ответа.
Авторы выражают искреннюю благодарность
А.И. Газиеву, И.Ю. Митрошиной и М.Г. Ломае-
вой за помощь в проведении некоторых экспери-
ментов, а также С.П. Романченко (лаборатория
клеточной инженерии ИТЭБ РАН, руководитель
группы цитогенетических исследований
-
С.И. Заичкина) за предоставление клеток асцит-
ной карциномы Эрлиха и В.В. Шапошниковой
(лаборатория окислительного стресса ИТЭБ
РАН, руководитель - Ю.Н. Корыстов) за предо-
ставление инокулята клеток мышиного лимфо-
лейкоза Р388.
Работа выполнена при частичной финансо-
вой поддержке Российского фонда фундамен-
тальных исследований (грант
№ 17-29-01007
«офи_м»).
СПИСОК ЛИТЕРАТУРЫ
1. Т. А. Зенченко, Дис. … д-ра биол. наук (ИТЭБ
РАН, Пущино, 2016).
2. Т. А. Зенченко, А. Н. Скавуляк, Н. И. Хорсева
и др., Геофизические процессы и биосфера 12 (1),
22 (2013).
3. Т. А. Зенченко и Н. Г. Варламова, Геофизические
процессы и биосфера 14 (2), 50 (2015).
4. А. Л. Марков, Т. А. Зенченко, Ю. Г. Солонин и др.,
Авиакосмич. и экологич. медицина 47 (2),
29
(2013).
5. А. И. Газиев, Радиац. биология. Радиоэкология 39
(6), 630 (1999).
Рис. 8. Зависимость между атмосферной температу-
6. E. I. Azzam, J. P. Jay-Gerin, and D. Pain, Cancer Lett.
рой воздуха и уровнями повреждений ДНК интакт-
327, 48 (2012).
ных (а) и облученных (б) клеток лимфолейкоза мыши
7. A. R. Collins, A. A. Oscoz, G. Brunborg, et al., Muta-
Р388, а также их разности (в). Указаны значения объ-
genesis 23 (3), 143 (2008).
ясненной дисперсии R2 при аппроксимации зависи-
8. Д. А. Мошков, С. П. Романченко, Е. Ю. Парныш-
мостей полиномом второй степени.
кова и др., Бюл. эксперим. биологии и медицины
154 (11), 646 (2012).
9. Е. В. Инжеваткин, Практикум по эксперименталь-
сти, может служить удобным параметром. Реги-
ной онкологии на примере асцитной карциномы Эр-
стрируя изменения температуры, по-видимому,
лиха (Красноярский гос. ун-т, Красноярск, 2004).
можно прогнозировать реакцию живых клеток на
10. N. K. Chemeris, A. B. Gapeyev, N. P. Sirota, et al.,
воздействие генотоксических факторов по крите-
Mutat. Res. 558, 27 (2004).
рию уровня повреждений ДНК этих клеток, од-
11. D. P. Lovell and T. Omori, Mutagenesis 23 (3), 171
нако здесь необходимы дополнительные экспе-
(2008).
рименты.
12. П. М. Нагорский, Т. А. Зенченко, К. Н. Пустова-
Таким образом, можно полагать, что вариации
лов и др., Вестн. КРАУНЦ. Физ.-мат. науки 4 (20),
%TDNA у интактных лейкоцитов или опухолевых
64 (2017).
БИОФИЗИКА том 64
№ 3
2019
ИЗМЕНЕНИЕ УРОВНЕЙ ПОВРЕЖДЕНИЙ ДНК КЛЕТОК МЫШЕЙ
467
13.
N. Sirota, E. Kuznetsova, and I. Mitroshina, Radiat.
17. G. Speit, K. Trenz, et al., Toxicol. Lett. 110, 73 (1999).
Environ. Biophys. 57 (2) 115 (2018).
18. Е. С. Менчинская, Дис. … канд. биол. наук (Тихо-
14.
Г. А. Домрачев, Д. А. Селивановский, П. А. Стун-
океанский институт биоорганической химии им.
жас и др. Эффективность образования пероксида
Г.Б. Елякова ДВО РАН, Владивосток, 2014).
водорода и радикалов воды в природе, Препринт
ИПФ РАН № 537 (Нижний Новгород, 2000)
strains/char_strains/P388.
peroksida-vodoroda-i-radikalov.html,
dia.ru/text/80/052/50489.php).
20. Модели и методы экспериментальной онкологии,
под ред. А. Д. Тимофеевского (Медгиз, М., 1960)
15.
M. V. M. Lafleur, J. Woldhuis, and H. Loman, Int. J.
Radiat. Biol. 39 (2), 113 (1981).
16.
M. V. M. Lafleur, J. Woldhuis, and H. Loman, Nucl.
21. Л. А. Суворова и Л. Ю. Нугис, Мед. радиология и
Acids Res. 9 (23), 6591 (1981).
радиац. безопасность 57 (1), 30 (2012).
Changes in the Level of DNA Damage in Mouse Cells
as Induced by Atmospheric Factors
E.A. Kuznetsova*, N.P. Sirota*, and T.A. Zenchenko* **
*Institute of Theoretical and Experimental Biophysics, Russian Academy of Sciences,
Institutskaya 3, Pushchino, Moscow region, 142290 Russia
**Institute of Space Research, Russian Academy of Sciences,
Profsoyuznaya ul. 84/32, Moscow, 117997 Russia
Changes in the level of DNA damage as induced by atmospheric factors were studied according to the per-
centage of DNA in the tail (%TDNA) using the comet assay in blood leukocytes, Ehrlich ascites carcinoma
and lympholeukosis P388murine cells. It was shown that: (1) high values of DNA damage of blood leukocytes
(%TDNA) are recorded within narrow ranges of atmospheric temperature (from -4°С to +3°С) and partial
ambient oxygen level of 298-304 mg/l; (2) the range of variations in %TDNA of leukocytes increases with
increasing geomagnetic activity; (3) the changes in %TDNA of intact and irradiated (4 Gy) leukocytes and of
tumor cells are associated with changes in air temperature; and (4) there is a correlation between temperature
and %TDNA in both types of intact tumor cells and irradiated P388 cells. Thus, variations in %TDNA are
apparently due to the effect of environmental reactive oxygen species generated at temperatures of phase tran-
sitions of water. Presumably, changes in the basal %TDNA of cells may lead to changes in their radiation re-
sponse.
Keywords: comet assay, murine blood, Ehrlich ascites carcinoma cells, lympholeukosis P388 cells, x-ray, air tem-
perature
БИОФИЗИКА том 64
№ 3
2019