БИОФИЗИКА, 2020, том 65, № 1, с. 125-137
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 519.876.5
НЕЙРОСЕТЕВАЯ МОДЕЛЬ ПРОГНОЗИРОВАНИЯ ФЕНОЛОГИИ
СКОРОСПЕЛЫХ СОРТОВ СОИ ПО КЛИМАТИЧЕСКИМ ФАКТОРАМ
© 2020 г. О.Д. Таратухин*, Л.Ю. Новикова*, **, И.В. Сеферова**, Герасимова Т.В.**,
С.В. Нуждин*, ***, М.Г. Самсонова*, К.Н. Козлов*
*Санкт-Петербургский политехнический университет Петра Великого,
195251, Санкт-Петербург, Политехническая ул., 29
**Федеральный исследовательский центр «Всероссийский институт генетических ресурсов растений
имени Н.И. Вавилова», 190000, Санкт-Петербург, ул. Большая Морская, 42-44
***Университет Южной Калифорнии, CA 90089, Лос-Анжелес, США
E-mail: kozlov_kn@spbstu.ru
Поступила в редакцию 30.10.2019 г.
После доработки 30.10.2019 г.
Принята к публикации 01.11.2019 г.
Фенология сои сильно зависит от температуры и продолжительности светового дня и четко
указывает на изменения климата. Авторами разработана модель, которая включает в себя три
искусственные нейронные сети для прогнозирования временных интервалов между посевом,
всходами, цветением и созреванием в зависимости климатических факторов, а также ансамбли
регрессионных моделей для прогнозирования урожайности, содержания белка и масла в семенах.
Проанализированы данные по созреванию скороспелых образцов сои, фенотипированных на двух
опытных станциях ВИР имени Н.И. Вавилова в Северо-Кавказском и Северо-Западном регионах
России. Модель реализована на языке Python с использованием библиотек Keras и TensorFlow.
Ключевые слова: климатические фактоpы, cоя, искусственные нейронные сети.
DOI: 10.31857/S0006302920010159
Благодаря комплексу ценных свойств расте-
Распространенным подходом является расчет
ний и зерна, соя (Glycine max (L.) Merr.) является
суммы накопленных «единиц тепла» в сутки.
одной из главнейших белково-масличных куль-
Продолжительность периода между фазами раз-
тур в мире. Высокое содержание белка (40-45%)
вития определяется нахождением суммы вкладов
и масла (20-25%) в семенах сои обеспечивает по-
всех дней до достижения заданной величины, не-
стоянный рост значения этой культуры в миро-
обходимой для завершения фазы. Вклад за день
вой экономике. Соя - растение короткого дня
часто принимается равным произведению функ-
теплообеспеченных регионов умеренных широт,
ций длины дня и средней суточной температуры
однако ее биологический потенциал позволяет
воздуха [6-9]. Динамические и имитационные
выращивать сорта в широком диапазоне клима-
модели [10] позволяют исследовать развитие рас-
тических условий. В условиях глобальных и реги-
тения.
ональных климатических трендов научной осно-
Искусственные нейронные сети являются од-
вой для изучения механизмов адаптации расте-
ним из широко применяемых методов машинно-
ний к изменению условий обитания является
го анализа, который используется в различных
математическое моделирование и прогнозирова-
областях, в том числе для описания хозяйствен-
ние [1-3]. Индикатором климатических измене-
но-ценных признаков растений [11-13]. Искус-
ний и степени пригодности сортов к складываю-
ственные нейронные сети основываются на кон-
щимся климатическим условиям является фено-
цепции слоев искусственных нейронов. Исход-
логия растения [4].
ные данные подаются в нейронную сеть на
Математические модели хозяйственно ценных
входной слой, с которого поступают на произ-
показателей, таких как, например, длина перио-
вольное число промежуточных слоев, а результат
дов «посев-всходы», «всходы-цветение», «цвете-
считывается с выходного слоя. Такие параметры,
ние-созревание», могут строиться различными
как размерность и количество скрытых слоев,
методами. Часто используются регрессионные
должны быть заданы с учетом объема имеющихся
модели с различными наборами факторов [5].
экспериментальных данных. Модели на основе
125
126
ТАРАТУХИН и др.
Таблица 1. Группировка сортов по областям селекции
Группа
Сорта
ЕС
Sito, Major
Канада
Maple Ridge, KG-20
Север
ПЭП 26, ПЭП 27, ПЭП 28, ПЭП 2, ПЭП 17, ПЭП 18
Сибирь
СибНИИСХ 6, Степная 90, СибНИИК 15/83, Алтом, Степная 85
Швеция
Fiskeby 840-2-7, Fiskeby 840-7-3, Fiskeby V, Fiskeby 1040-4-2, Bravalla
Центр
Соер 13-91, Мадева, Восход*1191/79, Белор, Магева, УСХИ 6, Соер 4, Окская, Светлая
нейронных сетей используются и для исследова-
изучаемых образцов за годы исследований и по
ния фенологических показателей [14].
вариантам опыта содержание белка и масла в се-
менах заметно варьировало. В условиях Северо-
Запада содержание белка колебалось от 27,4 до
ОБЪЕКТ ИССЛЕДОВАНИЯ
49,7%, а масла - от 13,8 до 20,9%. В Краснодар-
И ЭКСПЕРИМЕНТАЛЬНЫЕ ДАННЫЕ
ском крае содержание белка составляло от 32,8 до
46,1%, а масла - от 16,4 до 24,4%.
Сбор фенотипических данных, показателей
продуктивности и качества семян проводили в
ФИЦ «Всероссийский институт генетических ре-
ГРУППИРОВКА ДАННЫХ
сурсов растений им. Н.И. Вавилова» (ВИР) в двух
Исходные данные содержат информацию о
географических районах - в Санкт-Петербурге
сортах, которые были созданы в нескольких гео-
(г. Пушкин) и Краснодарском крае (на Кубан-
графических областях. Образцы были объедине-
ской опытной станции) в 2004-2006 гг. В изуче-
ны в группы по областям селекции (см. табл. 1)
ние были включены скороспелые сорта, способ-
для сравнения результатов моделирования.
ные вызревать в условиях Краснодарского края и
По происхождению образцы могут быть разде-
формировать выполненные семена в условиях
Северо-Западного региона. В Пушкине проведе-
лены на созданные в Европе (Sito, Major), в Кана-
но два посева - основной и ранний, 25 и 11 мая
де (Maple Ridge, KG-20), в сибирских селекцион-
ных центрах (СибНИИСХ 6, Степная 90, Сиб-
соответственно, на Кубани посев проводили с
НИИК 15/83, Алтом, Степная 85), в Швеции
7 по 13 мая. В 2004 г. в Пушкинском филиале сум-
(Fiskeby 840-2-7, Fiskeby 840-7-3, Fiskeby V, Fiske-
ма активных температур (выше +10°С) за вегета-
цию в зависимости от сроков посева составляла
by 1040-4-2, Bravalla), в селекционных центрах
от 1718,9 до 1721,5°С; сумма осадков за вегетацию
средней полосы РФ (Соер 13-91, Мадева, Вос-
составила от 181,1 до 182,3 мм. В 2005 г. сумма ак-
ход*1191/79, Белор, Магева, УСХИ 6, Соер 4, Ок-
ская, Светлая), а также экспериментальные об-
тивных температур составляла от
1768,3 до
разцы, созданные в ВИРе в Ленинградской обла-
1924,5°С, сумма осадков за вегетацию - от 82,8 до
90,6 мм. В 2006 г. сумма активных температур со-
сти (ПЭП 26, ПЭП 27, ПЭП 28, ПЭП 2, ПЭП 17,
ставляла от 1789,2 до 1930,8°С, сумма осадков за
ПЭП 18).
вег етац ию - от 58,2 до 84,7 мм. На Кубанско й
опытной станции (КОС ВИР) в 2004 г. сумма ак-
МОДЕЛЬ ДЛИНЫ ПЕРИОДОВ НА ОСНОВЕ
тивных температур за вегетацию была выше и со-
НЕЙРОННЫХ СЕТЕЙ
ставляла 2020°С при сумме осадков 339,7 мм; в
2005 г. сумма активных температур составляла
В работах [14, 17] была предложена модель на
2247°С, сумма осадков - 107 мм. Продолжитель-
основе нейронной сети, которая состоит из четы-
ность светового дня составляла от 16 ч (в мае) до
рех входных нейронов, двадцати нейронов в
18 ч (в июне) в Пушкине и от 14 ч (в мае) до 15 ч (в
единственном скрытом слое и единственного вы-
июне) на КОС ВИР.
ходного нейрона. В данной работе сеть была мо-
дернизирована для включения информации о
Полевые исследования проводили общепри-
принадлежности образца одной из групп Gk,
нятыми методами в соответствии с «Методиче-
k = 1, ... 6. Для каждого образца Gk = 1, если обра-
скими указаниями по изучению коллекции зер-
новых бобовых культур» [15] и «Международным
зец принадлежит к группе k, в иных случаях
классификатором СЭВ рода Glycine Willd» [16]. У
Gk = 0. Схема сети показана на рис. 1. Показаны
БИОФИЗИКА том 65
№ 1
2020
НЕЙРОСЕТЕВАЯ МОДЕЛЬ ПРОГНОЗИРОВАНИЯ ФЕНОЛОГИИ
127
Рис. 1. Нейронная сеть модели длины периодов «посев-всходы», «всходы-цветение» и «цветение-созревание».
На входы сети I1-I5 подаются результаты применения специально подобранных функций климатических факторов,
G1-G6 получают информацию о принадлежности к группе. Показаны связи только первого нейрона входного слоя с
нейронами скрытого слоя, чтобы не загромождать рисунок. Все нейроны входного слоя связаны с каждым нейроном
скрытого слоя.
связи только первого нейрона входного слоя с
обучения сети для достижения наилучших ре-
нейронами скрытого слоя, чтобы не загромож-
зультатов.
дать рисунок. Все нейроны входного слоя связа-
ны с каждым нейроном скрытого слоя.
МОДЕЛЬ УРОЖАЙНОСТИ, СОДЕРЖАНИЯ
На входы сети I1, I2, I3, I4, I5 подаются резуль-
БЕЛКА И МАСЛА В СЕМЕНАХ
таты применения специально подобранных
Модели прогнозирования урожайности, со-
функций, которые обобщают влияние ежеднев-
держания белка и масла в семенах строили с по-
ных показателей максимальной и минимальной
мощью программы autoML [18] на языке Python 3
температуры, осадков, солнечной радиации и
по признакам: «Группа», «Длина периода «по-
длины светового дня на состояние растения. Чис-
сев-всходы»», «Длина периода «всходы-цвете-
ло входных функций по сравнению с предыдущи-
ние»», «Длина периода «цветение-созревание»»,
ми моделями было увеличено до пяти.
«Температура
«посев-всходы»»,
«Осадки
«по-
Входные данные Ii вычисляются по формуле (1)
сев-всходы»»,
«Температура
«всходы-цвете-
для каждого дня D суммированием по всем
ние»», «Осадки «всходы-цветение»», «Темпера-
предыдущим дням j неотрицательных значений
тура «всходы-созревание»», «Осадки «всходы-
функций fi от gj - вектора ежедневных погодных
созревание»». AutoML использует автоматиче-
данных и параметров:
ское машинное обучение на основе байесовской
оптимизации для построения ансамбля регресси-
D
I
H f(
g f (g
),
(1)
онных моделей. Такой подход позволяет снизить
i
=∑
i
j
i
j
j=1
риск переобучения [18].
gj = {Tx,j, Tm,j, Pj, Rj, Lj, Tx,c, Tm,c, Pc, Rc, Lc},
(2)
где H - функция Хевисайда (равна единице для
ПРОГНОЗЫ ИЗМЕНЕНИЯ КЛИМАТА
неотрицательных аргументов, иначе
0); Tm,c,
Моделирование погоды на каждый день на Ку-
Tx,c - нижняя и верхняя температурные границы
бани с 2019 до 2030 года было проведено с помо-
роста; Lc - базовая длина светового дня; Tm,j, Tx,j,
щью генератора погоды - программы МаркСим
Lj - минимальная температура, максимальная
[19-23]. Были учтены социоэкономические сце-
нарии развития, которые описываются четырьмя
температура и длина светового дня за день j; Pc,
характерными профилями концентрации угле-
Pj - базовое и ежедневное количество осадков;
кислого газа (Representative Concentration Path-
Rc, Rj - базовая и ежедневная солнечная радиа-
ways, RCPs), принятые IPCC (Intergovernmental
ция. Конкретные значения параметров в векторе
Panel on Climate Change - Межправительствен-
gj и вид функций fj должны быть выяснены в ходе
ная группа экспертов по изменению климата) для
БИОФИЗИКА том 65
№ 1
2020
128
ТАРАТУХИН и др.
пятого оценочного отчета (Assessment Report,
AR5) в 2014 г. Профили соответствуют широкому
кругу возможных изменений будущих антропо-
генных выбросов парниковых газов и называются
rcp26, rcp45, rcp60 и rcp85 в соответствии с воз-
можными значениями нарушения радиационно-
го баланса Земли в 2100 г. относительно преинду-
стриальной эры (+2.6, +4.5, +6.0 и +8.5 Вт/м2 со-
ответственно) [24].
РЕЗУЛЬТАТЫ АДАПТАЦИИ МОДЕЛЕЙ
С помощью программы autoML [18] построе-
ны модели урожайности, содержания белка и
масла в семенах сои по признакам: «Группа»,
«Длина периода «посев-всходы»», «Длина перио-
да «всходы-цветение»», «Длина периода «цвете-
ние-созревание»», «Температура «посев-всхо-
ды»», «Осадки «посев-всходы»», «Температура
«всходы-цветение»»,
«Осадки
«всходы-цвете-
ние»»,
«Температура
«всходы-созревание»»,
«Осадки «всходы-созревание»». Набор данных
включал 194 записи. В программе autoML были
использованы параметры по умолчанию.
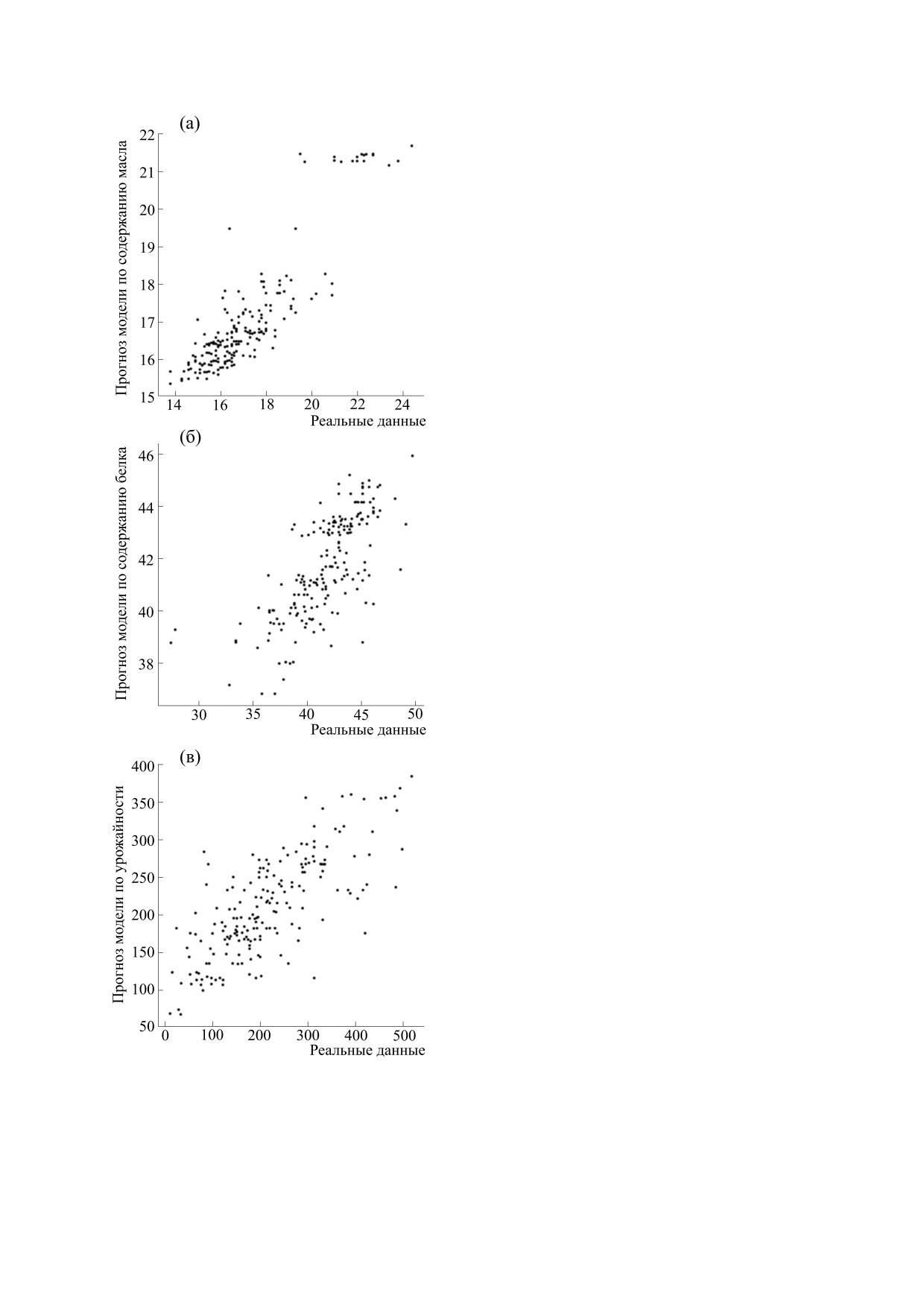
На рис. 2 показано сравнение данных о содер-
жании масла и белка в семенах и урожайности и
решений модели. Видно, что модель достаточно
точно описывает экспериментальные данные.
Коэффициент корреляции Пирсона на уровне
значимости <0.01 составил для масла 0.88, для
белка - 0.72, для урожайности - 0.75.
С использованием бутстрап-подхода в про-
грамме sklearn [25] были оценены относительные
доли влияния признаков на модели (см. табл. 2).
Урожайность и содержание белка в семенах в зна-
чительной степени зависели от генотипа, содер-
жание масла - от тепловлагообеспеченности ве-
гетации.
Модели для длин периодов «посев-всходы»,
«всходы-цветение» и
«цветение-созревание»
были запрограммированы на языке Python с ис-
пользованием Keras [26] и TensorFlow [26] в каче-
стве вычислителя. Данные о ежедневных макси-
мальных и минимальных температурах, осадках и
длине дня были взяты в открытом доступе в сети
функций fi, а также значение констант Tm,c, Tx,c,
Lc, Pc, Rc были получены путем минимизации от-
клонения решения модели от данных с использо-
ванием методов разностной эволюции [28, 29] и
грамматической эволюции [17, 30].
Точность моделей оценивали с помощью ко-
эффициента корреляции Пирсона между экспе-
Рис. 2. Сравнение данных и решения модели для
риментальными данными и решением модели.
содержания масла (а), белка (б) и урожайности (в).
Модели с наилучшей точностью были отобраны
БИОФИЗИКА том 65
№ 1
2020
НЕЙРОСЕТЕВАЯ МОДЕЛЬ ПРОГНОЗИРОВАНИЯ ФЕНОЛОГИИ
129
Таблица 2. Относительные доли влияния признаков в модели урожайности, содержания белка и масла в семенах
сои
Модель содержания
Модель содержания
Модель
Признак
масла
белка
урожайности
Группа
0.096
0.300
0.274
Длина периода «посев-всходы»
0.077
0.072
0.130
Длина периода «всходы-цветение»
0.044
0.072
0.096
Длина периода «цветение- созревание»
0.059
0.088
0.131
Температура периода «посев-всходы»
0.024
0.079
0.054
Осадки периода «посев-всходы»
0.088
0.091
0.037
Температура периода «всходы-
0.044
0.077
0.074
цветение»
Осадки периода «всходы-цветение»
0.044
0.043
0.054
Температура периода «всходы-
0.200
0.089
0.090
созревание»
Осадки периода «всходы-созревание»
0.324
0.089
0.059
Примечание. Значения округлены до трех знаков после запятой по правилам округления.
Таблица 3. Характеристика моделей периодов на основе нейронных сетей и
Период «цветение-
Имя
Период «посев-всходы»
Период «всходы-цветение»
созревание»
f1
Pj - Pc
Rj - Rc - Tx,j - Tx,c + 1/Tm,j
Rc
f2
(Tx,j - Tx,c) - (Tx,j - Tx,c)/Lj
1/(Tx,j- Tx,c)/(1/(Rj - Rc))
1/(Tx,j - Tx,c)
f3
1/(Lj - Lc) + (Tm,j- Tm,c) + 1/Pj
1/(Pj - Pc) - (Pj - Pc) + (Lj - Lc)
Tm,j
f4
(Tx,j - Tx,c)(Tm,j - Tm,c)
1/(Tm,j - Tm,c)
(Tx,j - Tx,c) + 1/(Tx,j- Tm,c)
f5
Tx,j - 1/(Tx,j - Tx,c)
1
Tm,j - Tx,c
Tm,c
6.683
10.174
10.975
Tx,c
26.136
29.868
23.805
Lc
-
15.493
15.544
Pc
7.288
-
0.376
Rc
-
6.479
0.090
r
0.79
0.78
0.067
Примечание. Tm,c, Tx,c - нижняя верхняя граница роста; Lc - базовая длина светового дня; Tm,j, Tx,j, Lj - минимальная,
максимальная температура и длина светового дня за день j; Pc, Pj - базовое и ежедневное количество осадков; Rc, Rj - базовая
и ежедневная солнечная радиация; r - коэффициент корреляции Пирсона между экспериментальными данными и
решением модели..
БИОФИЗИКА том 65
№ 1
2020
130
ТАРАТУХИН и др.
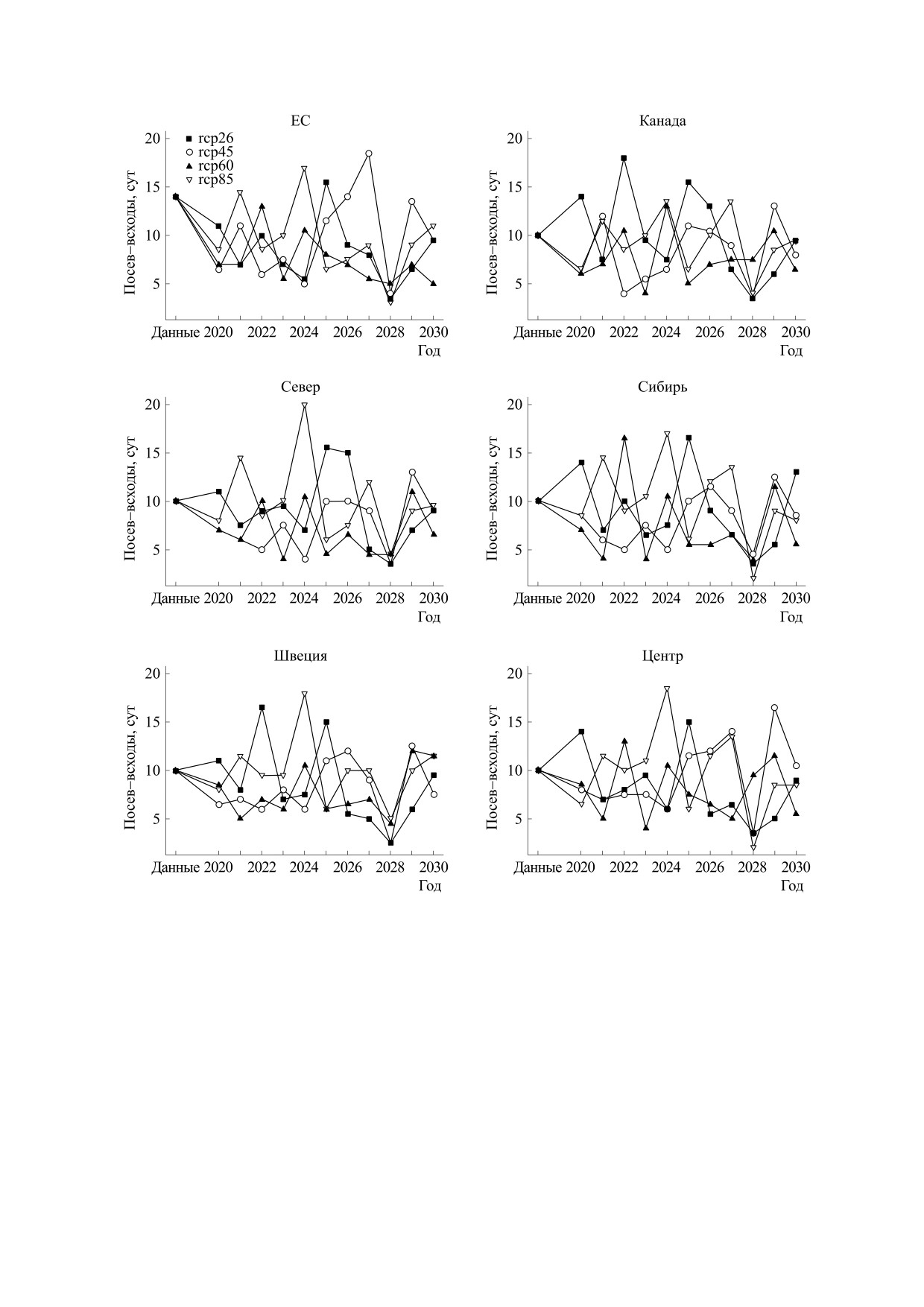
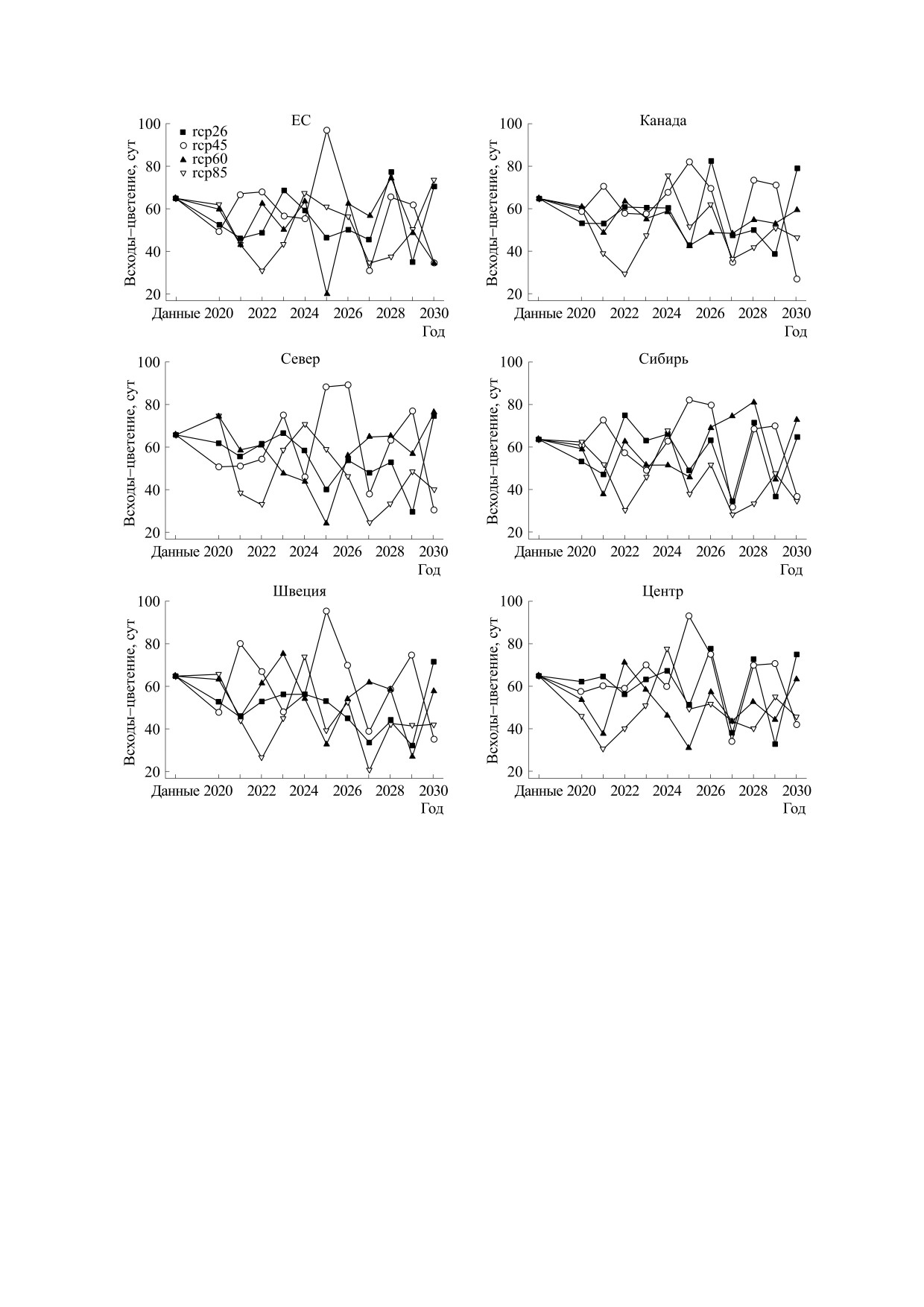
Рис. 3. Прогноз времени всходов сои по группам образцов разного происхождения. Различными символами —
квадрат, круг, треугольник и обратный треугольник - помечены значения, соответствующие четырем характерным
профилям концентрации углекислого газа: rcp26, rcp45, rcp60 и rcp85 соответственно.
для дальнейшего исследования (см. табл. 3). В
РЕЗУЛЬТАТЫ ИСПОЛЬЗОВАНИЯ МОДЕЛИ
каждой из моделей присутствуют линейные и не-
линейные функции. Модель для длины периода
Созданные погодные данные и построенные
«посев-всходы» оказалась независящей от сол-
модели были использованы для прогнозирования
нечной радиации, а для периода «всходы-цвете-
длин периодов
«посев-всходы»,
«всходы-
ние» - независящей от осадков. Модели показы-
цветение», «цветение-созревание», а затем для
вают удовлетворительную точность.
прогнозирования урожайности и содержания
БИОФИЗИКА том 65
№ 1
2020
НЕЙРОСЕТЕВАЯ МОДЕЛЬ ПРОГНОЗИРОВАНИЯ ФЕНОЛОГИИ
131
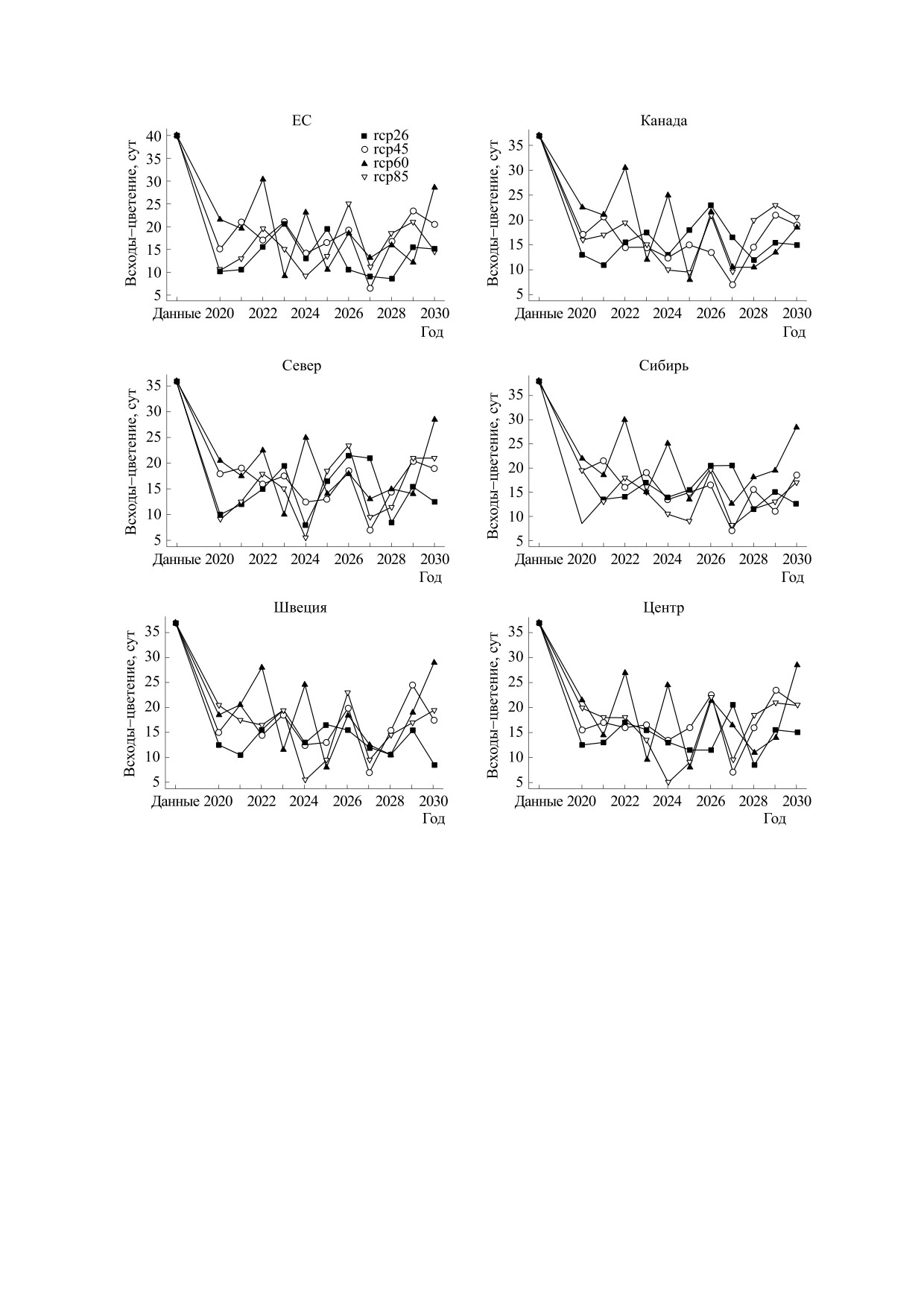
Рис. 4. Прогноз времени цветения сои, по группам образцов разного происхождения. Условные обозначения, как на
рис. 3.
белка и масла в семенах для модельного посева
тение» (см. рис. 4) имеют тенденцию к
10 мая шести групп сортов сои в Кубани в 2019-
уменьшению, однако также подвержены колеба-
2030 гг.
ниям. В отличие от прогноза времени до всходов,
верхняя граница колебаний существенно ниже
Результаты моделирования показывают, что
значений в реальных данных, в то время как ниж-
время до всходов (см. рис. 3) сильно колеблется
няя граница находится на уровне в три-четыре ра-
год от года в интервале примерно от -50% до
за ниже. Модельные значения длины периода
+50% к значениям в данных. При этом макси-
мальные и минимальные значения в большин-
«цветение-созревание» (см. рис. 5) изменяются
стве случаев относятся к профилю rcp85. Про-
колебательно, однако не имеют выраженной тен-
гнозные значения длины периода «всходы-цве-
денции к увеличению или уменьшению и лишь в
БИОФИЗИКА том 65
№ 1
2020
132
ТАРАТУХИН и др.
Рис. 5. Прогноз времени созревания сои, по группам образцов разного происхождения. Условные обозначения, как
на рис. 3.
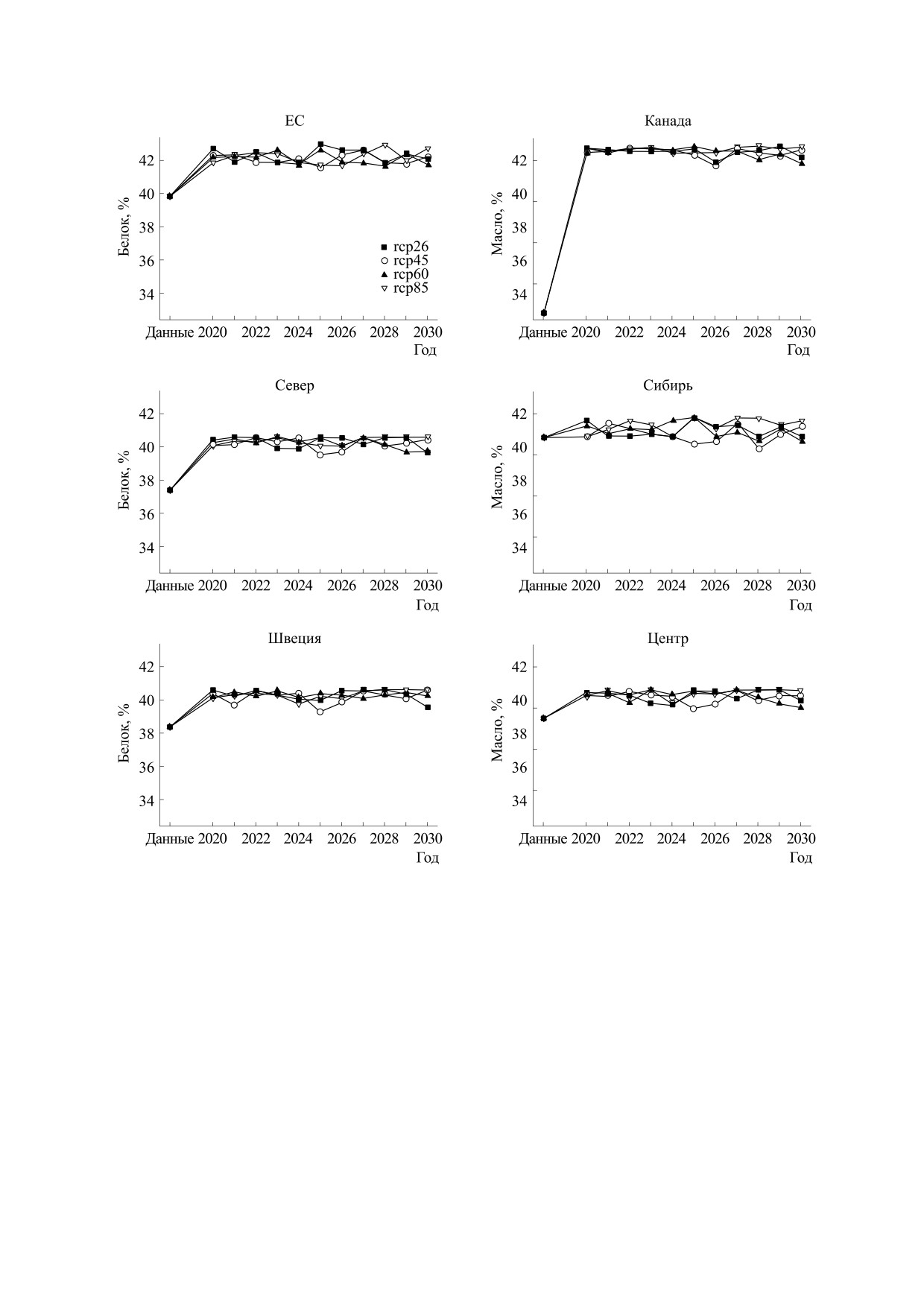
редких случаях отличаются от значений в экспе-
ри - на уровне 41%, для остальных сортов (ЕС и
риментальных данных больше чем на треть.
Канада) - на уровне 42%. При этом для Сибири и
Центра прогноз показывает лишь слабые колеба-
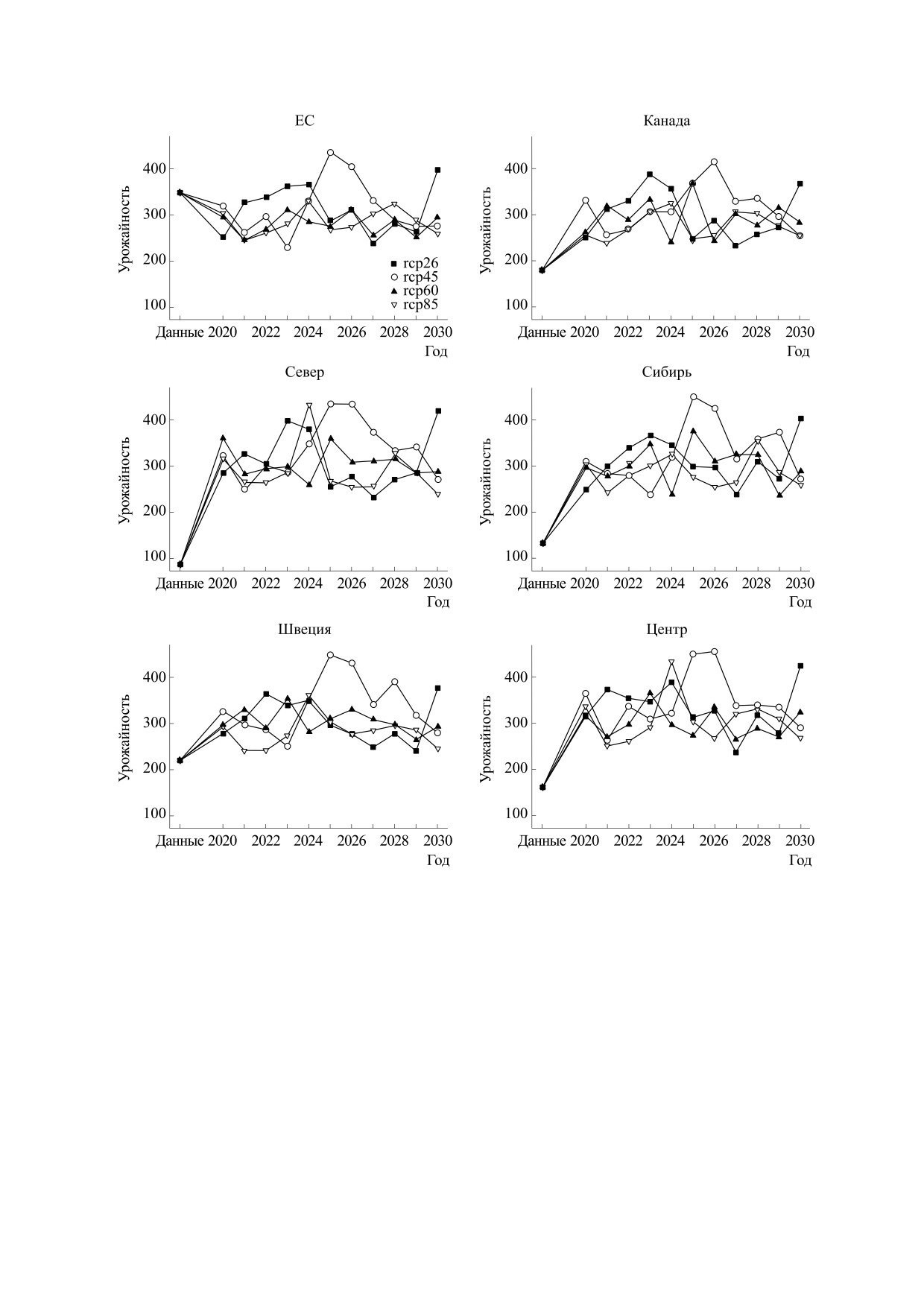
Результаты прогнозирования урожайности,
ния и незначительный прирост, в то время как
содержания белка и масла в семенах сои показа-
для остальных сортов прогнозируется прирост
ны соответственно на рис. 6, 7 и 8. Для всех сор-
содержания белка. Прогноз урожайности пока-
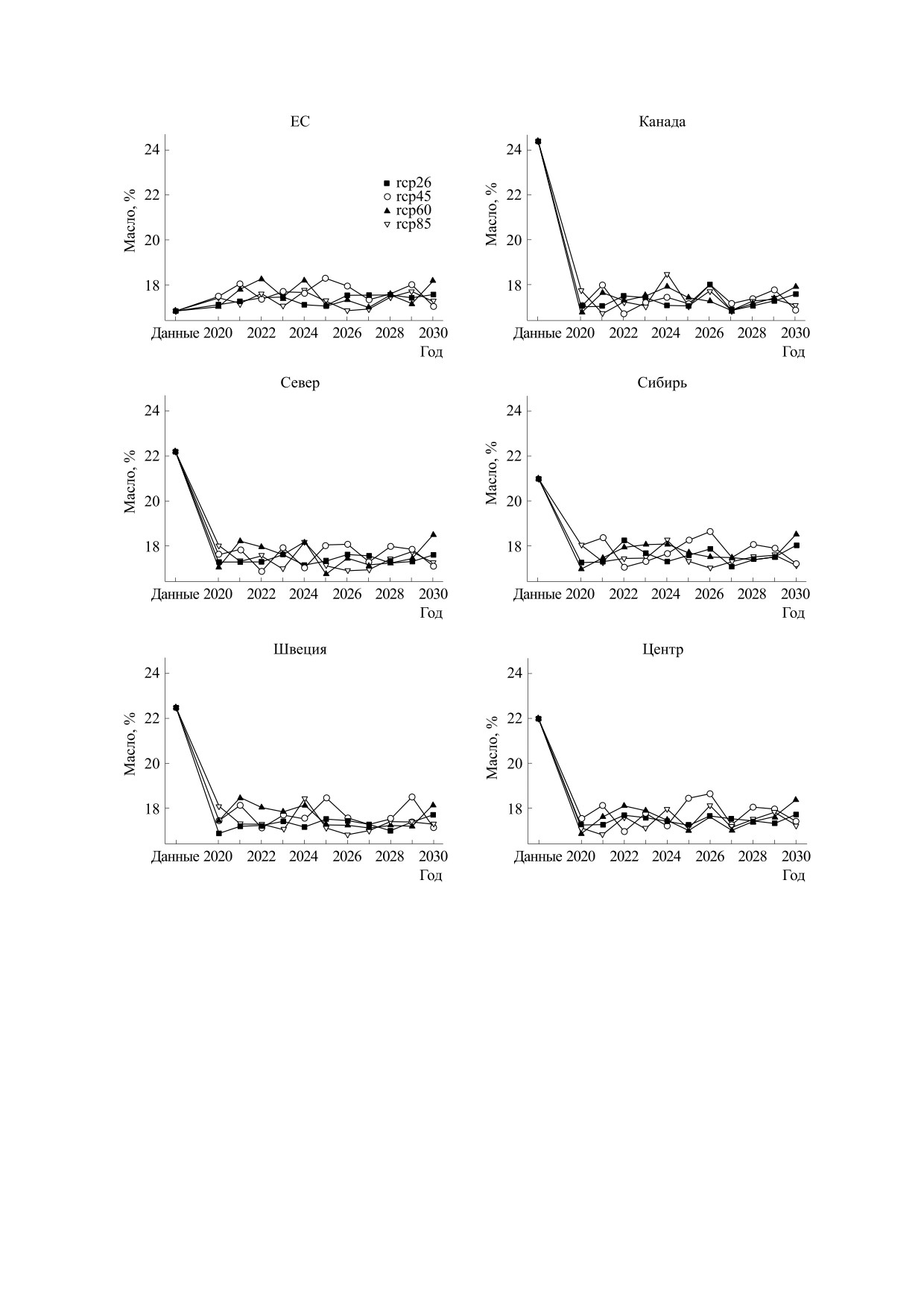
тов, кроме группы ЕС, в которой данные получе-
зывает ее рост для всех групп сортов, кроме груп-
ны для Ленинградской области, содержание мас-
пы ЕС, при наличии ярко выраженных коле-
ла прогнозируется значительно ниже значений в
баний.
реальных данных. Сорта делятся на три группы по
прогнозу содержания белка. Для сортов Севера,
Такие результаты можно объяснить тем, что
Швеции и Центра прогнозируется содержание
для моделей содержания белка в семенах и уро-
белка на уровне примерно 40%, для сортов Сиби-
жайности принадлежность образца к группе име-
БИОФИЗИКА том 65
№ 1
2020
НЕЙРОСЕТЕВАЯ МОДЕЛЬ ПРОГНОЗИРОВАНИЯ ФЕНОЛОГИИ
133
Рис. 6. Прогноз содержания масла в семенах сои, по группам образцов разного происхождения. Условные
обозначения, как на рис. 3.
ет сравнительно высокую относительную долю
в моделях и сложным балансом погодных факто-
влияния (см. табл. 2), поэтому прогнозы отлича-
ров. Одновременное снижение содержания масла
ются по группам (см. рис. 7 и 8). Колебания про-
в семенах сои и рост урожайности требует допол-
гнозируемых значений урожайности, белка и
нительного объяснения. Мы предполагаем, что в
масла порождаются колебаниями прогнозируе-
условиях прогноза может формироваться больше
мых значений длин периодов, которые можно
семян. Однако скороспелые сорта попадают при
объяснить стохастической природой моделей ге-
созревании в условия избыточно высоких темпе-
нерации погоды, а также наличием нелинейности
ратур, при которых накопление масла понижено.
БИОФИЗИКА том 65
№ 1
2020
134
ТАРАТУХИН и др.
Рис. 7. Прогноз содержания белка в семенах сои, по группам образцов разного происхождения. Условные
обозначения, как на рис. 3.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
мата. В данной работе были построены модели
для предсказания характеристик сортов сои, вы-
Использование математических моделей для
веденных в шести регионах и фенотипированных
прогнозирования хозяйственно-ценных характе-
в условиях Северо-Кавказского и Северо-Запад-
ристик растений, таких как длины периода «по-
ного регионов. Разработанные модели использу-
сев-всходы»,
«всходы-цветение»,
«цветение-
созревание», а также урожайность, содержание
ют новейшие подходы математического модели-
белка и масла в семенах сои, должно являться на-
рования, такие как автоматическое машинное
учной основой селекционного улучшения сортов
обучение, нейронные сети и грамматическую
культуры в условиях глобальных изменений кли-
эволюцию, с помощью которой были подобраны
БИОФИЗИКА том 65
№ 1
2020
НЕЙРОСЕТЕВАЯ МОДЕЛЬ ПРОГНОЗИРОВАНИЯ ФЕНОЛОГИИ
135
Рис. 8. Прогноз урожайности сои, по группам образцов разного происхождения. Условные обозначения, как на рис. 3.
функции погодных данных. Расчеты по имею-
Модельные прогнозы показали различную ре-
щимся экспериментальным данным показали
акцию групп сортов на изменение климата. Одна-
высокую точность моделирования. Прогноз ро-
ко для большинства сортов прогнозируется рост
ста содержания белка в семенах сои, не столь бла-
урожайности, что указывает на благоприятные
гоприятный прогноз для содержания масла, зави-
перспективы использования изменений климата.
симость содержания белка от сорта совпадают с
результатами, полученными по простым регрес-
Таким образом, показана пригодность модели
сионным уравнениям [31]. Однако новейшие
фенологии сои, созданной методом искусствен-
подходы показали более высокие прогностиче-
ных нейронных сетей, для прогнозирования в из-
ские способности моделей.
меняющемся климате.
БИОФИЗИКА том 65
№ 1
2020
136
ТАРАТУХИН и др.
ФИНАНСИРОВАНИЕ РАБОТЫ
12.
M. Kaul, R. L. Hill, and C. Walthall, Agricult. Systems
85 (1), 1 (2005).
Работа выполнена в рамках Федеральной це-
левой программы (проект
№14.575.21.0136 от
13.
A. Bagherzadeh, et al., Modeling Earth Systems and
26.09.2017, уникальный идентификатор проекта
Environment 2 (2), (2016).
RFMEFI57517X0136).
14.
D. A. Elizondo, R. W. McClendon, and G. Hoogen-
boom, Trans. ASAE 37 (3), 981 (1994).
БЛАГОДАРНОСТИ
15.
Н. И. Корсаков, О. П. Адамова и В. И. Будаова,
Вычисления были поведены в Суперкомпью-
Методические указания по изучению коллекции зер-
тером центре «Политехнический» CПбПУ и кла-
новых бобовых культур (ВНИИ растениеводства
стере Университета Южной Калифорнии.
им. Н. И. Вавилова, Ленинград, 1975).
Исходные данные получены на базе уникаль-
16.
Л. Щелчко, Международный классификатор СЭВ
ной научной установки Коллекция генетических
рода Glycine Willd (Науч.-техн. совет стран-членов
ресурсов растений ВИР.
СЭВ по коллекциям диких и культурных видов
растений, ВНИИ растениеводства им. Н. И. Вави-
лова, Ленинград, 1990).
КОНФЛИКТ ИНТЕРЕСОВ
17.
О. Д. Таратухин, Л. Ю. Новикова, И. В. Сеферова
Авторы заявляют об отсутствии конфликта
и К. Н. Козлов, Биофизика 64 (3) 563 (2019).
интересов.
18.
M. Feurer, et al. in Advances in Neural Information Pro-
cessing Systems, Ed. by C. Cortes et al. (Curran Associ-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
ates, Inc., 2015), pp. 2962-2970.
Настоящая работа не содержит описания ка-
19.
M. Srinivasa Rao, et al., PLoS One 10 (2), e0116762
ких-либо исследований с использованием людей
(2015).
и животных в качестве объектов.
20.
P. G. Jones and P. K. Thornton, Agricult. Forest Mete-
orol. 86 (1-2), 127 (1997).
СПИСОК ЛИТЕРАТУРЫ
21.
P. G. Jones and P. K. Thornton, Agricult. Forest Mete-
1. А. Г. Папцов и др., Адаптация сельского хозяйства
orol. 97 (3), 213 (1999).
России к глобальным изменениям климата. Проект
независимой международной организации Окс-
22.
P. G. Jones and P. K. Thornton, Agronomy J. 92, 445
фам
(2000).
iblock/f96/f9622b41f48549945438f2292f509d14.pdf
(Электронный ресурс, режим доступа: 06.01.2019).
23.
P. G. Jones and A. L. Jones, MarkSim: a computer tool
that generates simulated weather data for crop modeling
2. J. E. Olesen, et al., Eur. J. Agronomy 34 (2), 96 (2011).
and risk assessment (CIAT, 2002).
3. T. Carter and K. Mäkinen, Approaches to climate change
24.
D. P. van Vuuren, et al., Climatic Change 109 (1-2), 5
impact, adaptation and vulnerability assessment: towards
(2011).
a classification framework to serve decision-making: ME-
DIATION 2.1 (Finnish Environment Institute, Helsin-
25.
F. Pedregosa, et al., J. Machine Learning Res. 12, 2825
ki, Finland, 2011).
(2011).
4. A. D. Richardson, et al., Global Change Biology 18 (2)
26.
F. Chollet, et al., Keras (GitHub, 2015). URL: https://
566 (2012).
5. К. Н. Козлов, Л. Ю. Новикова, И. В. Сеферова и
27.
M. Abadi, et al., in Proc. 12th USENIX Conf. on Operat-
М. Г. Самсонова, Биофизика 63 (1), 175 (2018).
ing Systems Design and Implementation (Savannah, GA,
6. D. J. Major, et al., Crop Science 15, 174 (1975).
USA, 2016), pp. 265-283.
7. T. Hodges and V. French, Agronomy J. 77 (3), 500
28.
K. Kozlov, A. M. Samsonov, and M. Samsonova, Peer
(1985).
J. Comp. Sci. 2, e74 (2016).
8. P. Pedersen, et al., Agronomy J. 96, 556 (2004).
29.
R. Storn and K. Price, J. Global Optimization 11, 341
(1997).
9. T. D. Setiyono, et al., Field Crops Res. 100 (2-3), 257
(2007).
30.
M. O’Neill and C. Ryan, IEEE Trans. Evolutionary
10. Л. Ю. Новикова, И. В. Сеферова и К. Н. Козлов,
Computation 5 (4), 349 (2001).
Биофизика 63 (6) 1182 (2018).
31.
Л. Ю. Новикова, И. В. Сеферова, А. Ю. Некрасов
11. M. Abdipour, et al., J. Am. Oil Chemists’ Soc. 95 (3),
и др., Вавиловский журн. генетики и селекции 22
283 (2018).
(6) 708 (2018).
БИОФИЗИКА том 65
№ 1
2020
НЕЙРОСЕТЕВАЯ МОДЕЛЬ ПРОГНОЗИРОВАНИЯ ФЕНОЛОГИИ
137
An Artificial Neural Network Model for Prediction of Phenology of Early Maturing
Soybean Varieties in Relation to Climate Factors
O.D. Taratuhin*, L.Yu. Novikova*, **, I.V. Seferova**, T.V. Gerasimova**, S.V. Nuzhdin*, ***,
M.G. Samsonova*, and K.N. Kozlov*
*Peter the Great St. Petersburg Polytechnic University, ul. Polytekhnicheskaya 29, St. Petersburg, 195251 Russia
**Federal Research Center “N.I. Vavilov All-Russian Institute of Plant Genetic Resources”,
ul. Bolshaya Morskaya 42-44, St. Petersburg, 190000 Russia
***University of Southern California, Los Angeles, CA, 90089 USA
Soybean phenology is strongly influenced by temperature and day length and phenological records clearly re-
f lect changes in climatic conditions. We developed a model that includes four artificial neural networks for
forecasting time intervals between sowing, emergence, flowering and maturity in relation to climatic factors
and also formed assemblies of regression models to predict the yield, seed protein and oil content in soybean.
We analyzed data on maturation of early ripening soybean accessions phenotyped at two experimental sta-
tions of N.I. Vavilov Institute of Plant Genetic Resources in North-Caucasian and North-Western districts
of Russia. The model was implemented in Python using Keras and TensorFlow serving libraries.
Keywords: climatic factors, soybeans, artificial neural networks
БИОФИЗИКА том 65
№ 1
2020