БИОХИМИЯ, 2019, том 84, вып. 7, с. 995 - 1015
УДК 616
КАНЦЕРОГЕНЕЗ, АССОЦИИРОВАННЫЙ С ИНФЕКЦИЕЙ
ВИРУСАМИ ПАПИЛЛОМЫ ЧЕЛОВЕКА, ЕГО МЕХАНИЗМЫ
И ВОЗМОЖНОСТИ ИММУНОТЕРАПИИ
Обзор
© 2019
М.С. Вонский1,2, М.Г. Шабаева3, А.Л. Рунов1,2,4, Н.Н. Лебедева4,5,
С. Човдхари6, Д.М. Палефский6, М.Г. Исагулянц4,7,8,9*
1 Институт цитологии РАН, 194064 Санкт Петербург, Россия;
электронная почта: m.vonsky@gmail.com
2 Национальный медицинский исследовательский центр им. В. А. Алмазова
Минздрава России, 197341 Санкт Петербург, Россия
3 Первый Санкт Петербургский государственный медицинский университет им. акад. И.П. Павлова,
197022 Санкт Петербург, Россия; электронная почта: maria.dobro@mail.ru
4 Национальный исследовательский центр эпидемиологии и микробиологии
имени почетного академика Н.Ф. Гамалеи Минздрава России,
123098 Москва, Россия; электронная почта: maria.issagouliantis@rsu.lv
5 ГКУЗ МО «Центр по профилактике и борьбе со СПИДом и инфекционными заболеваниями»,
129110 Москва, Россия; электронная почта: lebedevanatalya@rocketmail.com
6 University of California, San Francisco School of Medicine,
CA 94143 San Francisco, USA; E mail: Joel.Palefsky@ucsf.edu
7 Федеральный научный центр исследований и разработки иммунобиологических
препаратов им. М.П. Чумакова РАН, 108819 Москва, Россия
8 Department of Microbiology, Tumor and Cell Biology, Karolinska Institutet,
SE 171 77, Stockholm, Sweden; E mail: maria.issagouliantis@ki.se
9 Department of Pathology, Riga Stradins University, LV 1007, Riga, Latvia
Поступила в редакцию 02.04.2019
После доработки 19.04.2019
Принята к публикации 21.04.2019
Папилломавирусная инфекция отвечает за 5% всех раковых заболеваний и 30% всех раков, вызванных ин
фекционными агентами. Ассоциированный с вирусами папилломы человека (ВПЧ) рак шейки матки явля
ется третьим по распространенности раком среди женщин в мире, при этом ~70% случаев обусловлены ин
фекцией ВПЧ высокого канцерогенного риска (ВПЧ ВКР) типа 16 и 18. Инфицирование ВПЧ происходит,
в основном, при половом контакте, однако существуют и другие способы передачи вируса, реализуемые в
рамках горизонтального и вертикального путей. После инфицирования кератиноцитов базального слоя
эпителия или клеток экто и эндоцервикальной зоны перехода ДНК ВПЧ персистирует в эписомальной
форме, и в большинстве случаев инфицированные клетки элиминируются иммунной системой. Однако в
ряде случаев элиминации не происходит, и инфекция переходит в хроническую фазу. Репликация вируса в
делящихся эпителиальных клетках сопровождается повышением уровня синтеза онкобелков Е6 и Е7, обла
дающих канцерогенной активностью. Они ответственны за индукцию геномной нестабильности, наруше
ние клеточного цикла, клеточную пролиферацию, иммортализацию и, в конечном счете, злокачественную
трансформацию ВПЧ инфицированных клеток. Эти белки вызывают также иммуносупрессию, препят
ствуя выявлению иммунной системой ВПЧ инфицированных и трансформированных клеток. Важной осо
бенностью ВПЧ ассоциированного канцерогенеза является интеграция ВПЧ в геномную ДНК клетки хо
зяина, приводящая к повышению экспрессии онкобелков Е6 и Е7 и стимулирующая онкогенез. Профилак
тические вакцины против ВПЧ позволяют предотвратить до 89% случаев развития ВПЧ ассоциированных
аногенитальных раков, однако успешность их применения связана с ВПЧ наивностью вакцинируемого ор
ганизма. Вызываемый ими иммунный ответ не способен элиминировать персистирующий вирус и предот
вратить развития ВПЧ ассоциированных неопластических изменений. Ограниченная эффективность про
филактической ВПЧ вакцинации при условии существования неконтролируемых вертикальных и горизон
тальных путей инфицирования новорожденных и детей до достижения ими возраста вакцинации требуют
создания терапевтических вакцин против ВПЧ и ВПЧ ассоциированных неоплазий.
КЛЮЧЕВЫЕ СЛОВА: вирусы папилломы человека (ВПЧ), канцерогенез, плоскоклеточный рак, рак шейки
матки, интраэпителиальные неоплазии, эпидемиология, онкобелки Е6 и Е7.
DOI: 10.1134/S0320972519070091
995
996
ВОНСКИЙ и др.
Вирусные инфекции обусловливают разви
ВПЧ рода β придерживаются cтратегии ми
тие 15-20% от общего числа онкологических за
нимизации экспрессии, их репликация в орга
болеваний, вирусы папилломы человека (ВПЧ),
низме подавляется иммунитетом хозяина. ВПЧ
считающиеся причиной возникновения 5% ра
рода α развили стратегию уклонения от имму
ковых заболеваний и 30% всех раков, вызван
нитета за счет его модуляции и/или подавления
ных инфекционными агентами, вносят один из
вирусными белками. Эти стратегии отражают
основных вкладов в эту статистику. ВПЧ пред
существенные различия в экспрессии вирусных
ставляют большое семейство малых безоболо
генов и в функциях одних и тех же вирусных
чечных двухцепочечных ДНК вирусов, являю
белков у ВПЧ различных родов, которые меня
щихся причиной доброкачественной пролифе
ются также и в зависимости от типа инфициру
рации эпителия или кондилом, впервые опи
емой ткани.
санных в 1933 г Shope и Hurst [1]. Развитие тех
нологий молекулярно генетического анализа
показало существование целого ряда типов
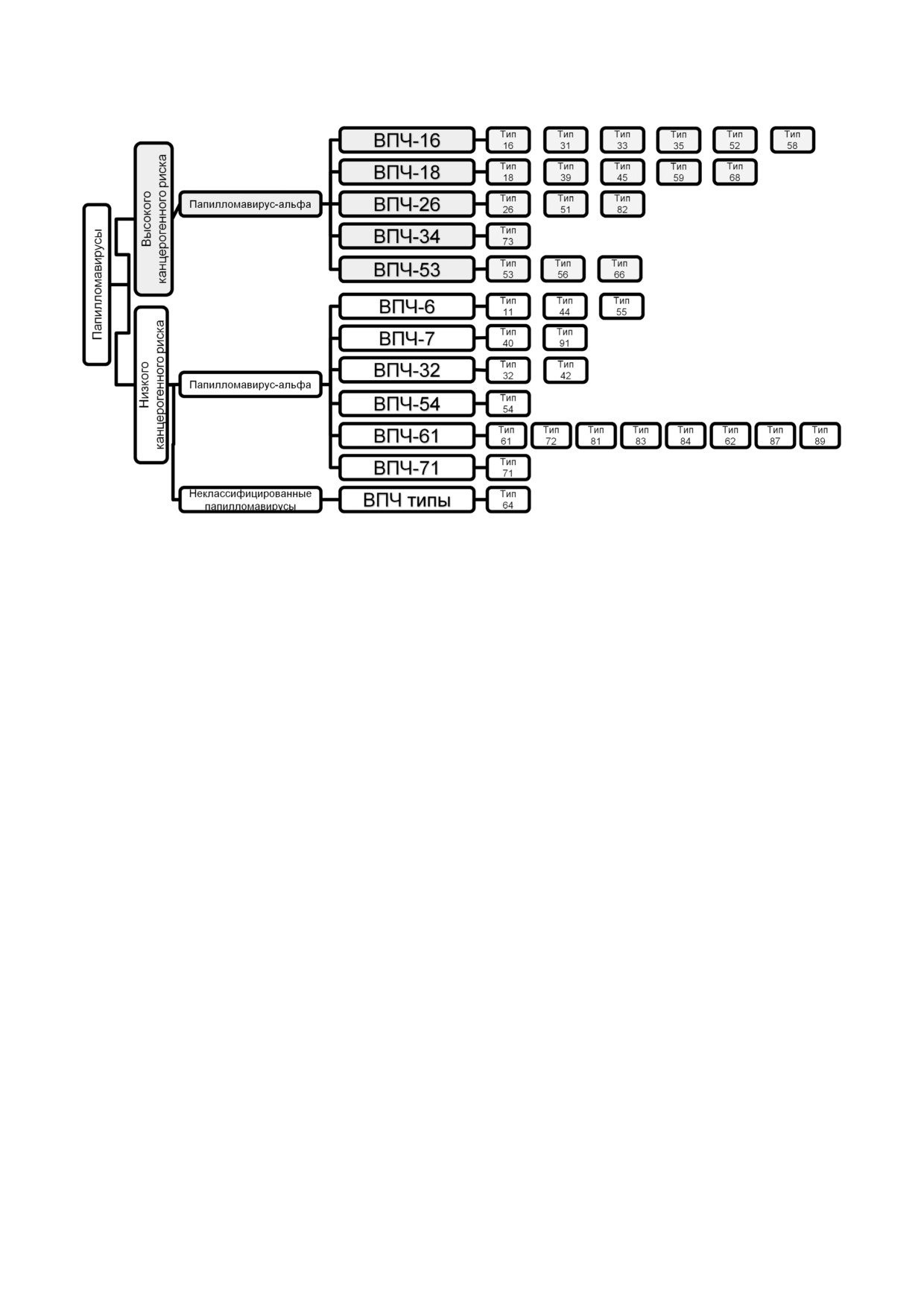
ТИПЫ ВПЧ, АССОЦИИРОВАННЫЕ
ВПЧ, обладающих тропизмом к поверхностям
С КАНЦЕРОГЕННЫМ РИСКОМ
слизистых оболочек или эпидермальному плос
кому эпителию и вызывающих как доброкачест
По клинической значимости ВПЧ разделили
венные, так и злокачественные новообразова
на две группы: генотипы (далее, типы) ВПЧ
ния, в частности, рак шейки матки (РШМ), вто
низкого канцерогенного риска, вызывающие в
рую по распространенности форму рака у жен
основном кондиломы, и типы ВПЧ высокого
щин [2, 3]. На лидирующее место по частоте вы
канцерогенного риска (ВКР), приводящие к ма
ходит ВПЧ ассоциированный рак носоглотки,
лигнизации инфицированных тканей. Между
его распространенность за последние три деся
народное агентство по исследованию рака
тилетия возросла на 225% и, предположительно,
(IARC) предложило более детальную классифи
к 2020 году превзойдет частоту РШМ [4].
цикацию типов ВПЧ [6]:
Вирусы папилломы как человека, так и жи
- неканцерогенные типы: ВПЧ 6, 11 (2);
вотных устойчиво размножаются в специфичес
- возможно канцерогенные типы: ВПЧ
ких типах многослойного эпителия хозяина,
26, 53, 66, 67, 70, 73, 82, 30, 34, 69, 85 и 97
вызывая хронические бессимптомные инфек
(12);
ции, сопровождающиеся длительной продукци
- вероятно канцерогенный тип: ВПЧ 68;
ей вирионов с ограниченной экспрессией ви
- высоко канцерогенные типы: ВПЧ 16, 18,
русных генов, что минимизирует риск иммун
31, 33, 35, 39, 45, 51, 52, 56, 58 и 59 (12).
ного клиренса. После начальной инфекции ре
К настоящему моменту спектр типов ВПЧ
пликация вирусного генома в делящихся клет
ВКР, передающихся половым путем и участвую
ках эпителия находится на низком уровне. Эти
щих в канцерогенезе, развитии аногенитального
клетки образуют резервуар инфекции, который
и орофарингеального плоскоклеточного рака,
может сохраняться десятилетиями. Когда инфи
был расширен до 20 типов [7].
цированные клетки дифференцируются, уро
ВПЧ ВКР в значительной степени ассоци
вень экспрессии многократно возрастает, при
ированы с аногенитальными раками (с раком
этом поверхностные эпителиальные клетки на
шейки матки в первую очередь), с рядом форм
чинают продуцировать вирионы [5]. К настоя
рака носоглотки, головы и шеи, а также с высо
щему времени идентифицировано уже >200 ге
ким уровнем развития интраэпителиальных нео
нотипов ВПЧ, выделенных из клинических об
плазий - предшественников аногенитальных
разцов. Их классификация основана на анализе
раков. Классификация неоплазий по системе
последовательности ДНК, номера генотипам
Bethesda [8] включает категории:
присваивают по мере выявления и идентифика
- атипичные клетки плоского эпителия не
ции (рис. 1).
ясного значения (ASC US);
Принятые сокращения: ВПЧ - вирусы папилломы человека; ВПЧ ВКР - ВПЧ высокого канцерогенного риска;
РШМ - рак шейки матки; СПИД - синдром приобретенного иммунодефицита; LSIL - плоскоклеточные интраэпители
альные повреждения низкой степени (Low grade squamous intraepithelial lesion); HSIL - плоскоклеточные интраэпители
альные повреждения высокой степени (High grade squamous intraepithelial lesion); CIN1 - легкая цервикальная интраэпи
телиальная неоплазия; CIN2 - умеренная интраэпителиальная неоплазия; CIN3 - тяжелая интраэпителиальная неопла
зия; АРТ - высокоактивная антиретровирусная терапия; ОРС - открытая рамка считывания; L1 - большой белок капси
да ВПЧ; L2 - малый белок капсида ВПЧ; LCR, NCR или URR - некодирующая регуляторную область ВПЧ; ММ - мо
лекулярная масса.
* Адресат для корреспонденции.
БИОХИМИЯ том 84 вып. 7 2019
ВПЧ АССОЦИИРОВАНЫЙ КАНЦЕРОГЕНЕЗ
997
Рис. 1. Таксономическая схема классификация вирусов папилломы человека, разработанная Международным Комитетом
- атипичные клетки плоского эпителия, не
РАСПРОСТРАНЕННОСТЬ ВПЧ
исключающие HSIL (ASC H);
И ВПЧ@АССОЦИИРОВАННЫХ
- плоскоклеточные интраэпителиальные
ФОРМ РАКА
повреждения низкой степени (Low grade squa
mous intraepithelial lesion; LGSIL or LSIL);
По оценке Центра по контролю и профилак
- плоскоклеточные интраэпителиальные
тике заболеваний США (CDC), >90% сексуаль
повреждения высокой степени (High grade squa
но активных мужчин и >80% женщин инфици
mous intraepithelial lesion; HGSIL or HSIL);
руются в течение жизни по крайней мере одним
- инвазивный рак шейки матки (плоскокле
типом ВПЧ [13], причем примерно в 50% проис
точная карцинома; Squamous cell carcinoma);
ходит инфицирование ВПЧ относящимися к
- атипичные железистые клетки, не опреде
группе высокого канцерогенного риска (https://
ленные иначе (AGC NOS);
- атипичные железистые клетки, с подозре
risk/infectious agents/hpv and cancer). В глобаль
нием на аденокарциному in situ (AIS) или нео
ном плане наиболее широко распространены
плазию (AGC neoplastic);
генотипы ВПЧ ВКР 16, 18, 31, 33, 35, 45, 52 и 58
- аденокарцинома in situ (Adenocarcinoma in
situ; AIS).
038414/en/). Частоты встречаемости различных
Плоскоклеточным интраэпителиальным
типов ВПЧ ВКР сильно разнятся географичес
повреждениям низкой степени LSIL соответ
ки, в том числе в зависимости от уровня эконо
ствует легкая цервикальная интраэпителиаль
мического развития и возможностей нацио
ная неоплазия (CIN1); высокой степени HSIL -
нальных систем здравоохранения. Частота
умеренная или тяжелая интраэпителиальная нео
встречаемости первого по распространенности
плазия (CIN2/3) [9]. До 75% всех плоскоклеточ
генотипа ВПЧ ВКР, ВПЧ 16, составляет в Герма
ных раков и 94% всех аденокарцином вызвано
нии 77%, в Южной Америке - 71%, в США -
инфекциями ВПЧ типов 16, 18 31, 33 и 45 [10].
59%, в Японии в 33-39%. Вторым по распрост
Наиболее распространенными типами ВПЧ вы
раненности является ВПЧ 18 с глобальной час
сокого риска, отвечающими за развитие >80%
тотой встречаемости 8%, в США он является пя
всех цервикальных раков, являются ВПЧ 16 и 18
тым по частоте после ВПЧ типов 16, 52, 51 и 31.
[11]. ВПЧ 16 также предоминирует и в случаях
В целом в Европе превалирует ВПЧ 33, в странах
анальных раков [12].
Азии - 52 и 58 (в Японии ВПЧ 52 обнаруживают
БИОХИМИЯ том 84 вып. 7 2019
998
ВОНСКИЙ и др.
в 20% случаев РШМ, тогда как в США его выяв
скрининга рака шейки матки, внедрению вак
ляют только в 2% случаев) [6, 14, 15]. В Южной
цинации против ВПЧ были получены в ходе ре
Африке наиболее часто детектируются ВПЧ 16
ализации 7 й рамочной программы Европей
(11,7%), 58 (10,3%), 51 (8,9%), 66 (8,6%) и 18
ской комиссии (проекты PREHDICT, HPV
(7,6%); в суб Сахаре, одном из самых поражен
AHEAD, HEALTH F3 2010 242061 и HEALTH
ных регионов мира, частота встречаемости ВПЧ
F2 2011 282562). Распространенность ВПЧ 16 и
ВКР 16, 18 и 45 повышена (3-4)× в случаях
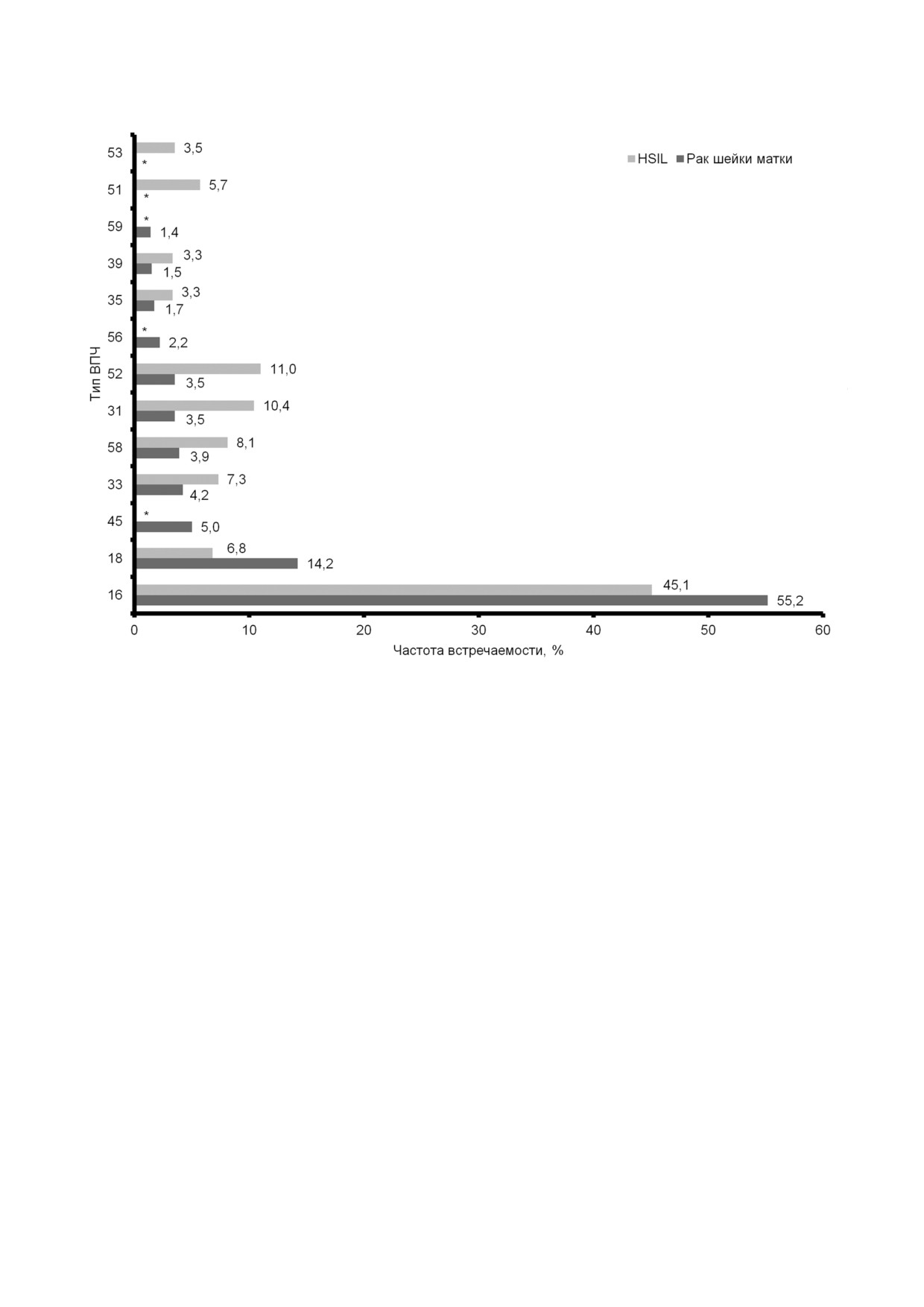
18 типов среди женщин с нормальной цитологи
РШМ у ВИЧ инфицированных [16-18].
ей составила 2,9 и 1,2%; с LSIL - 19,3 и 6,5%;
Среднемноголетний показатель общей забо
с HSIL - 45,1 и 6,8%; с инвазивным раком шей
леваемости ВПЧ ассоциированными новообра
ки матки - 55,2 и 14,2% соответственно [11]. По
зованиями за период 2007-2016 гг. среди муж
нашим данным, распространенность ВПЧ ВКР
ского населения Российской Федерации (РФ)
среди женщин детородного возраста в централь
составил 33,7, среди женского - 715,5 на 100 000
ной части России (n = 2783) составила в 2018 г.
человек [19]. В то время, как в странах Европы,
10%, из них 2% приходилось на ВПЧ 16, и 0,4% -
США, Австралии и Новой Зеландии распрост
на ВПЧ 18, остальную часть составили ВПЧ ВКР
раненность РШМ снижается, в РФ она растет.
других типов (неопубликованные данные). По
Среди женского населения РФ заболеваемость
лученные данные в целом совпадают с данными
РШМ матки растет в среднем на 2,26% в год:
по частоте встречаемости основных типов ВПЧ
в 2007 г она составляла 12,48, а к 2017 г. - уже
ВКР общемировой статистики (рис. 2).
15,76 на 100 тыс. населения. Число впервые ус
тановленных случаев в 2017 г. составило 17 587,
«грубый» и стандартизованный показатели за
ПУТИ ПЕРЕДАЧИ ВПЧ ИНФЕКЦИИ
болеваемости РШМ составили 22,3 и 15,76 слу
чаев на 100 000 населения соответственно [20].
Основной путь передачи ВПЧ инфекции -
При этом в отдельных регионах РФ стандарти
половой, во время традиционного, анального
зованный показатель заболеваемости РШМ
или орального секса с человеком, имеющим
превышает средний в 3 и более раза. Значитель
клинические или субклинические проявления
но меняется вклад РШМ в структуру смертнос
ВПЧ инфекции или вирусоносителем. ВПЧ -
ти от злокачественных новообразований в раз
одна из наиболее распространенных инфекций,
личных возрастных группах: он является причи
передаваемых половым путем, хотя имеется
ной смерти 7,1% женщин моложе 30 лет и ос
множество неоспоримых свидетельств верти
новной причиной (23,1%) - для возрастной
кальной и иных горизонтальных путей передачи
группы 30-39 лет. В последующих возрастных
вируса [22, 23]. На это прямо указывает высокая
группах его значимость снижается: в группе
частота обнаружения ВПЧ, включая ВПЧ ВКР, в
40-49 лет он занимает второе место (17,3%) пос
крайней плоти детей и подростков [24]. Распро
ле рака молочной железы (23,5%), в группе
странена также передача ВПЧ инфекции по вер
50-59 лет - 7,2%. Ежегодная смертность от
тикальному пути, от матери к плоду [25, 26].
РШМ в 2017 г. достигла 6480 случаев, прирост
Нуклеотидные последовательности - биомар
показателя смертности от РШМ в период
керы папиломавирусной инфекции обнаружи
2007-2017 г. составил 4,31% [20]. По другим дан
вают в полости рта новорожденных [26], груд
ным, РШМ в РФ занимает 1 ое место в структ
ном молоке [27], амниотической жидкости,
руре смертности от рака у женщин до 45 лет [19].
околоплодных водах, плаценте и пуповинной
В 2012 г. Роговской с соавт. был опубликован
крови [28]. ВПЧ, вызывающий папилломатоз
детальный анализ распространенности типов
гортани, может быть получен ребенком не толь
ВПЧ в разных регионах РФ в различных попу
ко при родах, но и in utero, так как его обнаружи
ляционных группах. Частота инфицирования
вают у детей, рожденных при помощи кесарева
ВПЧ ВКР в общей популяции варьировала от 13
сечения [22, 23]. При исследовании околоплод
до 40% и коррелировала с сексуальным поведе
ных вод у женщин, которые страдают гениталь
нием. В большинстве регионов РФ при РШМ и
ными кондиломами, и назофарингеальных ас
цервикальных неоплазиях тяжелой степени в
пиратов у рожденных ими детей определяются
81,8-100% случаев выявлялся ВПЧ ВКР, наибо
конкордатные типы вирусов папилломы, что
лее часто - 16 й и 18 й типы ВПЧ, при гени
указывает на трансплацентарную и интрана
тальных кондиломах - 6 й и 11 й типы [21]. Об
тальную передачу ВПЧ (в частности, ВПЧ 6 го и
ширные статистические данные по РШМ, дру
11 го типов). Интранатальное инфицирование
гим аногенитальным ракам, ракам головы и
ВПЧ может приводить к ювенильному рециди
шеи, частотам выявления ВПЧ, факторам, спо
вирующему респираторному папилломатозу,
собствующим раку шейки матки, практикам
частота которого составляет 1,7-2,6 на 100 000
БИОХИМИЯ том 84 вып. 7 2019
ВПЧ АССОЦИИРОВАНЫЙ КАНЦЕРОГЕНЕЗ
999
Рис 2. Встречаемость ВПЧ ВКР у женщин с плоскоклеточными интраэпителиальными повреждениями высокой степени
(HSIL) и раком шейки матки. * - Данные не представлены. Частоты встречаемости ранжированы по частоте выявления
при раке шейки матки (По данным Bruni et al., 2019 [11])
детей и 1 на 1500 родов среди женщин с гени
вали инфекцию ВПЧ, что позволило предполо
тальной ВПЧ инфекцией [29].
жить горизонтальный путь передачи инфекции
Метаанализ, охвативший 15 исследований в
внутри семьи [32].
медицинских учреждениях, общественных заве
Таким образом, возможны горизонтальный
дениях и 36 исследований пациентов, показал
и вертикальный пути передачи ВПЧ, в том чис
множественные горизонтальные пути передачи
ле от инфицированных родителей детям. Это
ВПЧ, как связанные, так и не связанные с поло
свидетельствует о важности профилактической
выми отношениями, в том числе передачу ВПЧ
вакцинации против ВПЧ не только для охраны
от рук к гениталиям или от гениталий к рукам,
здоровья женщин и мужчин, но и здоровья их
наличие инфекционных вирионов ВПЧ на по
детей, от зачатия и до достижения ими возраста
верхностях и оборудовании в медицинских уч
рекомендуемой профилактической ВПЧ вакци
реждениях и общественных местах [30]. Ряд ав
нации.
торов пишут о внутрисемейном пути передачи
инфекции [31]. В Испании было проведено рет
роспективное исследование, включавшее на
ВПЧ И ВПЧ@АССОЦИИРОВАННЫЕ
блюдение за ВПЧ положительными и ВПЧ от
ФОРМЫ РАКА НА ФОНЕ
рицательными беременными женщинами и их
ВИЧ ИНФЕКЦИИ
детьми, наблюдавшимися на протяжении 14 ме
сяцев. В 16,9% случаев среди детей, рожденных
Нарушение иммунного баланса у лиц, пере
от ВПЧ отрицательных женщин, диагностиро несших трансплантацию почек, печени или лег
БИОХИМИЯ том 84 вып. 7 2019
1000
ВОНСКИЙ и др.
ких, и заболевания, сопровождающиеся имму
слизистых оболочек у взрослых ВИЧ позитив
носупрессией, приводят к потере контроля над
ных мужчин и женщин. Выраженное проявле
ВПЧ инфекцией и повышению рисков развития
ние этой взаимосвязи среди пациентов с низки
ВПЧ ассоциированных форм рака. В частности,
ми уровнями CD4+ (<200 клеток/мм3) указывает
крайне высока частота выявляемости ВПЧ ВКР
на роль иммунной системы в контроле реплика
среди больных, инфицированных вирусом им
ции ВПЧ [35]. Наряду с этим показана также
мунодефицита (ВИЧ) [33, 34].
прямая корреляция между снижением уровня
Течение соматических и вторичных ВПЧ ас
CD4+ и повышением числа типов ВПЧ, обнару
социированных заболеваний напрямую связано
живаемых в аногенительных образцах у ВИЧ
с развитием ВИЧ обусловленной иммуносуп
инфицированных. Важную роль в увеличении
рессии и зависит от длительности заболевания,
числа типов ВПЧ и повышении вирусной на
уровня CD4+ лимфоцитов, активности реплика
грузки играет число половых контактов, но оп
ции ВИЧ, индивидуальных особенностей паци
ределяющей все же является потеря иммунного
ентов, а также влияния препаратов для высоко
контроля [35].
активной антиретровирусной терапии (АРТ)
Несмотря на наличие описанных выше кор
[34]. Интересно, что при ВПЧ ассоциирован
реляций, прямая зависимость между иммуно
ном канцерогенезе с иммуносупрессией связано
супрессией и частотой возникновения аногени
инфицирование ВПЧ и ранние стадии диспла
тального рака отсутствует. Сопоставление дан
зии, но не развитие рака. Результаты исследова
ных регистров больных с синдромом приобре
ний показывают корреляцию между иммуно
тенного иммунодефицита (СПИДом) и с раком
супрессией, регистрируемой по снижению
не позволяет выявить взаимосвязи между более
уровня CD4+ Т лимфоцитов (<500 клеток/мм3),
низкими уровнями CD4+, например, ниже 500
распространенностью цервикальной и анальной
клеток/мм3 или даже ниже 200 клеток/мм3, и
инфекции ВПЧ и уровнем ДНК ВПЧ в клетках
частотами развития цервикального или аналь
ного рака среди женщин и мужчин со СПИДом.
Не наблюдают также и явного увеличения забо
леваемости аногенитальным раком после диаг
ностирования СПИДа [36]. Ни регрессия нео
плазий, ни их прогрессия не оказываются свя
занными с иммунным статусом.
Неоднозначен и опыт использования АРТ
для контроля репликации ВПЧ у людей, ко ин
фицированных ВИЧ и ВПЧ. После начала сис
тематического применения АРТ заболеваемость
ВПЧ ассоциированными формами рака не сни
зилась (как в случае РШМ), а скорее увеличи
лась [37]. Некоторые исследования, посвящен
ные влиянию АРТ на естественную историю
ВПЧ ассоциированных интраэпителаильных
неоплазий, показали ее положительный эффект
[38], другие, напротив, отсутствие такого эф
фекта [39, 40]. При этом исследования, демон
стрирующие положительный эффект АРТ со
снижением уровня ВПЧ виремии, не обнару
жили регрессии неоплазий у большинства жен
щин с HSIL даже при нормализации уровня
CD4+ клеток [41, 42].
Этот феномен объясняют тем, что на позд
них стадиях заболевания восстановление им
мунного статуса уже «бесполезно». Вызываемая
ВПЧ хромосомная нестабильность (будет рас
мотрена ниже) приводит к накоплению в клет
ках необратимых генетических изменений, ко
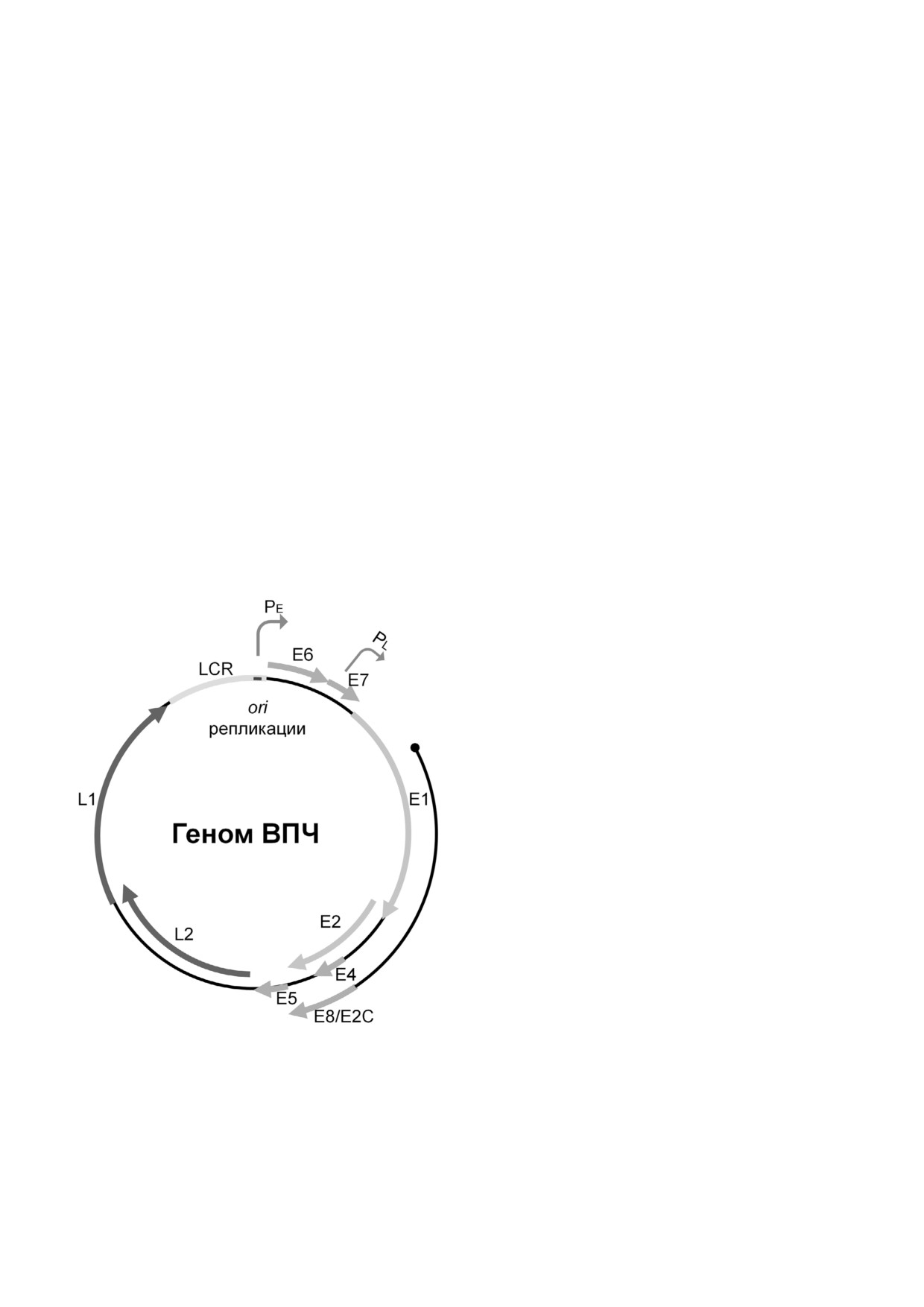
Рис. 3. Геном вируса папилломы человека (ВПЧ). Показа
на двуцепочечная кольцевая ДНК альфа семейства ВПЧ.
торые не поддаются исправлению иммунной
LCR - длинная регуляторная область; PE - ранний промо
системой, а сбой клеточного цикла - к тому,
тор; PL - поздний промотор; ori - участок инициации ре
пликации. Основные белки, участвующие в репликации
что такие «испорченные» клетки не уходят в
вируса - E1 и E2. Основные структурные белки - L1 и L2
апоптоз. Таким образом, причиной персисти
БИОХИМИЯ том 84 вып. 7 2019
ВПЧ АССОЦИИРОВАНЫЙ КАНЦЕРОГЕНЕЗ
1001
рующих интраэпителиальных поражений вы
нюю область (E), содержащую открытые рамки
сокой степени является не вирусная инфекция
считывания (ОРС), кодирующие не участвую
как таковая, а необратимые генетические изме
щие в репликации белки Е1, Е2, Е4, Е5, Е6 и
нения, в перспективе приводящие к инвазив
Е7, позднюю область (L), кодирующую боль
ному раку [39]. Понимание реальных механиз
шой (L1) и малый (L2) белки капсида и локали
мов развития ВПЧ ассоциированного рака,
зованную между ОРС L1 и Е6, некодирующую
особенно на фоне иммуносупрессии, имеет
регуляторную область, обозначаемую как LCR,
первостепенную важность для разработки ме
NCR или URR. Эта область содержит в своем
тодов его иммунотерапии, в том числе вакци
составе набор цис элементов, контролирую
нотерапии, давая основу для рационального
щих репликацию и транскрипцию вирусной
выбора стратегии вакцинации и вакцинных
ДНК, в частности, участок инициации репли
компонентов.
кации ori [5, 43].
Основные белки, кодируемые этими ОРС,
являются либо структурными, как капсидные
ГЕНОМ И ПРОТЕОМ ВПЧ
белки L1 и L2, либо функциональными, как E1
и E2, участвующие в репликации вирусной
Вирионы ВПЧ имеют консервативную ико
ДНК. Остальные белки (E4, E5, E6 и E7) коди
саэдрическую морфологию, вирусные частицы
руются не всеми папилломавирусами и могут
имеют диаметр 50-55 нм и молекулярную мас
рассматриваться как «эволюционные украше
су (ММ) 5 × 106 Да. Геномная организация всех
ния» [5]. Основные функции белков ВПЧ опи
папилломавирусов очень похожа: все вирусные
саны в таблице.
геномы состоят из двухцепочечной кольцевой
Белки L1 и L2. L1 - большой белок капсида
ДНК размером 6000-8000 п.о., ассоциирован
(ММ 55-57 кДа), обеспечивающий связывание
ной с гистон подобными белками [5] (рис. 3).
вирусной частицы с рецепторами гепарин суль
В составе генома выделяют три области: ран
фата, присутствующими на базальной мембране
Основные белки, кодируемые открытыми рамками считывания ВПЧ и их функции. Составлено на основе литературных
данных, рассмотренных ниже
Белок ВПЧ
ММ (кДа)
Основные функции
Е1
68-85 кДа
хеликаза; участвует в репликации вируса, в частности, в раскручивании вирусной ДНК
Е2
48 кДа
транскрипционный фактор; необходим для репликации вируса, сегрегации геномов и упа
ковки вирусной ДНК в капсид; регулятор транскрипции ОРС Е6 и Е7
Е3
нет данных
функция неизвестна, присутствует только в некоторых типах ВПЧ
Е4
10-44 кДа
связывается с белками цитоскелета, облегчает высвобождение и распространение вирионов
Е5
виропорин, обеспечивает проникновение вирусного генома в клетку; усиливает трансформи
14 кДа
рующую активность E6 и E7 путем регуляции экспрессии факторов роста и ингибирования
апоптоза, модулирует экспрессию генов, участвующих в адгезии клеток и клеточной подвиж
ности; снижает экспрессию молекул MHC классов I и II и подавляет противовирусный ин
терфероновый ответ
Е6
транскрипционный фактор; взаимодействует с рядом клеточных белков; индуцирует деграда
16-18 кДа
цию р53 и продукцию цитокинов
Е7
~10 кДа
подавляет активности pRb путем направления на протеасомную деградацию; нарушает тран
скрипционную регуляцию генов белков, участвующих в синтезе ДНК, нарушает эпигенетичес
кую регуляцию, стимулирует прогрессию фаз клеточного цикла и геномную нестабильность
Е8/Е2С
Е8 присутствует только в некоторых ВПЧ. В слитой форме с С терминальной частью Е2
20 кДа
функционирует как репрессор транскрипции и репликации ВПЧ ВКР
L1
57 кДа
большой белок капсида; организован в пентамерные капсомеры, формирующие икосаэдри
ческие вирионы
L2
малый белок капсида; участвует в инкапсуляции вирусной ДНК, обеспечивает внедрение и
43-53 кДа
транспорт вирионов
8 БИОХИМИЯ том 84 вып. 7 2019
1002
ВОНСКИЙ и др.
[44]. L1 кодируется ОРС L1, наиболее консерва
сомами, передавая, таким образом, эписомаль
тивной нуклеотидной последовательностью в
ные копии вируса после цитокинеза в дочерние
составе генома, вследствие чего она положена в
клетки [47]. E2 действует как регулятор тран
основу для построения филогенетической клас
скрипции ОРС Е6 и Е7 [9]. При повышении
сификации типов и субтипов ВПЧ. Малый бе
уровня экспрессии Е2 связывается с палиндром
лок капсида L2 (ММ 43-53 кДа) отвечает за упа
ной последовательностью 5' ACCG(N)4CGGT 3'
ковку вирусной ДНК. Белки L1 и L2 экспресси
в сайте Е2BS в области LCR в составе промото
руются на поздних стадиях, функционируют на
ра Р97 ВПЧ 16 [43], что предотвращает связыва
стадии сборки вирионов и инфекции и могут
ние этого участка с промоторным элементом
быть обнаружены преимущественно в диффе
РНК полимеразы II и подавляет экспрессию
ренцирующихся клетках эпителия [44].
белков Е6 и Е7. При низком уровне экспресии
Белок Е1. Высококонсервативный белок Е1,
Е2 способен рекрутировать транскрипционные
необходимый для репликации вирусов папилло
факторы, необходимые для вирусной реплика
мы, кодируется ОРС E1. Это вторая по консер
ции [49]. Таким образом, Е2 выступает в качест
вативности последовательность в составе гено
ве регулятора репродукции ВПЧ. Кроме того, Е2
ма ВПЧ [45, 46]. В составе белка Е1 выделяют
является эпигенетическим регулятором, спо
три функциональных домена - N концевой,
собным взаимодействовать с глобальным регу
центральный и наиболее значимый, С конце
лятором транскрипции - комплексом p300/
вой. В состав N концевого домена входят сиг
CBP p/CAF, что приводит к снижению тран
нальные последовательности ядерной локализа
скрипционной активности р53 вследствие гипо
ции и ядерного экспорта, мотив связывания с
ацетилирования хроматина [47]. Интересно, что
циклином Е/А и два сайта фосфорилирования
репликацию ВПЧ способны обеспечивать ком
циклин зависимой киназы Cdk2. Центральный
бинации Е1 и Е2 из вирусов разных типов, даже
домен отвечает за образование комплекса с бел
вирусов папилломы человека и животных, что
ком Е2 и его связывание с сайтом ori. С конце
указывают на высокую консервативность их
вой домен включает последовательность,
свойств.
действующую как хеликаза суперсемейства
Белок Е4. ОРС Е4 кодирует семейство бел
ААА+ АТФаз, связывающая и раскручивающая
ков, образуемое путем сплайсинга c последую
вирусную ДНК, что обеспечивает ее доступ
щей посттрансляционной модификацией. Е4
ность для репликативного комплекса клетки.
синтезируется в больших количествах в слоях
Все три домена играют важнейшую роль в ре
супрабазального и зернистого эпителия и обра
пликации вируса. Е1 вместе с Е2 образуют
зует кератин ассоциированные амилоидные
комплекс, связывающийся с участком инициа
филаменты в эпителиальных клетках среднего и
ции репликации вируса ori, причем в составе ori
верхнего слоев эпителиального пласта, повы
достаточно иметь один сайт связывания Е1 и
шая их хрупкость, что облегчает высвобождение
один сайт связывания Е2, при этом увеличение
и распространение вирионов ВПЧ [50].
числа сайтов связывания Е2 увеличивает эф
Белки Е5, Е6 и Е7 являются онкопротеинами
фективность репликации. После связывания,
ВПЧ, ответственными за иммортализацию и
Е2 вытесняется из комплекса, а Е1 формирует
трансформацию ВПЧ инфицированных кле
дигексамер, привлекающий топоизомеразу I,
ток. Основными онкогенными белками ВПЧ
ДНК полимеразу α и репликативный белок А
являются Е6 и Е7. Считается, что различия в он
(RPA), необходимые для репликации ВПЧ.
когенном потенциале ВПЧ связаны с различной
Кроме того, белок Е1 способствует формирова
выраженностью свойств Е6 и Е7, причем эти
нию разрывов ДНК в хроматине клетки хозяи
белки ВПЧ рода β менее эффективны в отноше
на, что способствует интеграции вируса [47, 48].
нии стимулирования онкогенеза с упором на
Белок Е2. Белок Е2 кодируется ОРС Е2. Бел
различные механизмы клеточной трансформа
ки Е1 и Е2 необходимы и достаточны для репли
ции, чем Е6 и Е7 ВПЧ рода α [51]. Основу онко
кации вирусов папилломы [45, 46]. Белок Е2
генной активности Е6 и Е7 составляет их спо
имеет N и С концевые трансактивирующие до
собность инактивировать онкосупрессорные
мены и действует как «загрузчик» Е1 хеликазы
белки р53 и pRb соответственно (рис. 4).
[47]. С концевой домен обеспечивает связыва
Белок Е5. Онкопротеин Е5 - гидрофобный
ние с белком BRD4, бромодомены которого взаи
трансмембранный белок с двумя цистеиновыми
модействуют с остатками лизина в составе гисто
остатками в С концевой части, стабилизирую
нов, что приводит к ремоделированию хромати
щими Е5 Е5 гомодимер. Молекулярная масса
на [49]. Кроме того, Е2 обеспечивает связь ви
Е5 у ВПЧ ВКР разных типов различается, сос
русной ДНК с хроматином клетки: комплекс
тавляя от 8,3 кДа для ВПЧ 18 до 9,4 кДа для
Е2-BRD4 связывается с митотическим хромо
ВПЧ 16. ВПЧ ВКР кодируют форму E5 (E5α) с
БИОХИМИЯ том 84 вып. 7 2019
ВПЧ АССОЦИИРОВАНЫЙ КАНЦЕРОГЕНЕЗ
1003
высокой про карциногенной активностью, а ва
работу иммунной системы. Он снижает
рианты ВПЧ низкого карциногенного риска -
экспрессию на поверхности инфицированных
E5 с пониженной активностью (E5β, γ, δ) или
клеток молекул главного комплекса гистосов
вообще не несут этой рамки считывания [52].
местимости (MHC) классов I и II и подавляет
Было предложено классифицировать Е5 как ви
противовирусный интерфероновый ответ и ак
ропорин, белок, образующий поры в клеточной
тивность естественных клеток киллеров (НК)
мембране, способный модулировать ионный го
[56]. В совокупности, это приводит к наруше
меостаз, перенос везикул, продукцию вирионов
нию презентации вирусных антигенов, подавле
и проникновение вирусного генома в клетку
нию воспалительной реакции и ослаблению
[53]. Онкопотенциал Е5 был выявлен сравни
противовирусного иммунного ответа, что спо
тельно недавно. Оказалось, что он играет клю
собствует выживанию инфицированных и
чевую роль в росте клеток, вмешиваясь в целый
трансформированных клеток [57].
ряд сигнальных путей. Как онкопротеин, Е5
Белок Е6. Онкопротеин Е6 состоит из
усиливает трансформирующую активность E6 и
151-158 а.о., не является структурным и не обла
E7 путем регуляции факторов роста, таких как
дает ферментативной активностью. Основной
рецептор эпителиального фактора роста EGF R,
клеточной мишенью белка E6 ВПЧ ВКР являет
Bcl 2, Bax, Fas и калнексина, участвующих в
ся туморсупрессорный белок p53. Уровень p53 в
контроле выживания, роста и дифференциров
клетке регулируется убиквитин лигазой Mdm2,
ки клеток [54]. Про канцерогенная активность
содержащей домен RING finger. Однако в усло
Е5 включает также ингибирование апоптоза,
виях стресса, например, при повреждении ДНК
индуцированного лигандом фактора некроза
или вирусной инфекции, этот механизм наруша
опухоли (TNFL) и лигандом CD95 (CD95L) [54].
ется, р53 стабилизируется и активируется путем
Помимо этого, Е5 модулирует экспрессию ге
фосфорилирования. В ВПЧ позитивных рако
нов, участвующих в адгезии клеток и клеточной
вых клетках Mdm2 зависимое регулирование
подвижности [55]. Е5 активно вмешивается в
уровня р53 полностью выключено, его уровень
Рис. 4. Основные механизмы ВПЧ Е6/Е7 индуцированного канцерогенеза
БИОХИМИЯ том 84 вып. 7 2019
8*
1004
ВОНСКИЙ и др.
регулируется белком Е6 [58]. Сам по себе E6 де
воздействуя на целый ряд контролирующих ее
градировать р53 не может, для этого он привле
клеточных факторов, что предотвращает укора
кает клеточную убиквитин лигазу E3 UBE3A/
чивание теломеров и старение клеток [62]. Со
E6AP (белок, ассоциированный с E6). UBE3A
вокупность свойств Е6 делает его необходимым
связывается с Е6 через мотив LXXLL, образуя
и достаточным фактором канцерогенеза, что
стабильный комплекс E6/E6AP, который и убик
подтверждается спонтанной индукцией карци
витинилирует р53, отправляя его на деградацию
ном у трансгенных по Е6 мышей [63].
протеасомой. Е6 обладает и другими анти апоп
Белок Е7. ОРС Е7 ВПЧ ВКР кодирует онко
тотическими свойствами. Ряд исследований по
белок размером до 127 а.о. Белок имеет три кон
казал взаимодействие Е6 с другими компонента
сервативных домена CD1, CD2 и CD3, послед
ми протеасомного пути деградации, также спо
ний гомологичен большому Т антигену SV40.
собствующими малигнизации [59]. В частности,
Наиболее важные функции Е7, определяющие
Е6 способствует протеасомной деградации Bak,
его онкогенность, связаны с доменами CD2 и
играющего важную роль в терминальной диффе
CD3. Домен CD2 содержит сайт фосфорилиро
ренциации клеток эпителия, и ряда других фак
вания протеинкиназы CKII и мотив LXCXE, оп
торов, задействованных в клеточном апоптозе -
ределяющий связывание с белком супрессором
рецептора фактора некроза опухолей 1 (TNF
ретинобластомы pRb. Белок pRb играет ключе
R1), адапторного белка Fas ассоциированного
вую роль в клеточном цикле, определяя переход
домена смерти (adaptor molecule Fas associated
клетки из фазы G1 в фазу S. В нормальных усло
death domain; FADD) и прокаспазы 8 (рис. 4).
виях, вначале G1 pRb не фосфорилирован и
Характерной особенностью Е6 является на
постепенно фосфорилируется при продвиже
личие двух цинковых пальцев, образованных
нии клетки к S фазе. Нефосфорилированный
двумя парами мотивов СХХС и домена PDZ
pRb взаимодействует с транскрипционными
класса I (PSD 95/Dlg/ZO 1) на С конце. Эти
факторами E2F и действует как репрессор тран
мотивы строго консервативны во всех белках
скрипции промоторов, содержащих сайты E2F.
Е6, их целостность важна для функциональной
Члены семейства E2F участвуют в зависимой от
активности онкопротеина [60]. Посредством
клеточного цикла транскрипционной регуля
цинковых пальцев E6, как транскрипционный
ции многих генов белков, обеспечивающих син
фактор, взаимодействует с широким спектром
тез ДНК [64]. Используя мотив LXCXE, E7 свя
клеточных субстратов, меняя профили экспрес
зывается с нефосфорилированным pRb, направ
сии генов, нарушая внутриклеточную регуля
ляя его на убиквитинилирование и последую
цию и сигналинг, что способствует утрате кле
щую деградацию протеасомой. Разрушение
точной полярности [61]. В частности, E6 участ
комплекса pRb E2F высвобождает E2F, что при
вует в регуляции трансактивации гена р53. Это
водит к E2F индуцированной транскрипции, в
уникальный дополнительный механизм регуля
результате которой повышаются уровни цик
ции активности р53, не зависящий от действия
лин зависимой киназы CDK2 и циклинов A и E
протеасомы. Мутанты Е6, неспособные вызвать
[59]. Стимулированная Е7 повышенная актив
деградацию р53, могут, тем не менее, блокиро
ность CDK2 определяет амплификацию цент
вать трансактивационную активность р53.
росом и их кластеризацию, являющиеся харак
Е6 опосредованная репрессия р53 зависимой
терным признаком раковых клеток [65]. Подав
активации коррелирует с ингибированием аце
ление активности pRb онкобелком E7 считается
тилирования р53 и коровых гистонов, что пре
критическим фактором стимулирования пере
пятствует их инкорпорации в хроматин и, соот
хода фаз клеточного цикла в дифференцирую
ветственно, осуществлению туморсупрессорных
щем эпителии, что обеспечивает оптимальные
функций. E6 также может контролировать
условия для репликации вирусной ДНК и од
p53 зависимую регуляцию генов путем взаимо
новременно способствует злокачественной
действия с ко активатором p300/CBP, это взаи
трансформации клеток.
модействие вносит свой вклад в ингибирование
Домен CD3 белка Е7 содержит четыре высо
р53 зависимой транскрипции [59].
коконсервативных остатка цистеина и участвует
Онкопротеин Е6 также может связываться с
во взаимодействиях с многочисленными кле
белками: XRCC1, интегральным регулятором
точными белками, включая ингибиторы р21 и
эксцизионной репарации оснований и О6 ме
р27 CDK. Е7 ингибирует их активность, позво
тилгуанозин ДНК трансферазой, затрудняя ре
ляя экспрессирующим его клеткам избежать
парацию одноцепочечных разрывов [47]. Па
ареста клеточного цикла, сопряженного с по
раллельно с этим, Е6 дерегулирует процесс ре
вреждением ДНК. Помимо этого, Е7 прямо свя
пликации ДНК путем активации теломеразы,
зывается и активирует ДНК метилтрансферазы,
обратной транскриптазы человека (hTERT),
что приводит к неконтролируемому повыше
БИОХИМИЯ том 84 вып. 7 2019
ВПЧ АССОЦИИРОВАНЫЙ КАНЦЕРОГЕНЕЗ
1005
нию уровня метилирования ДНК и нарушению
ИНФИЦИРОВАНИЕ
эпигенетической регуляции клеточных процес
И РЕПЛИКАЦИЯ ВИРУСА
сов [66].
Онкопротеины Е6 и Е7 действуют синергич
Фазы жизненного цикла ВПЧ представлены
но, индуцируя генетическую нестабильность,
на рис. 5. ВПЧ является исключительно интра
дестабилизируя транскрипционные комплексы
эпителиальным патогеном - как инфекция, так
и ремоделируя хроматин, что стимулирует не
и вегетативное размножение вируса полностью
контролируемую клеточную пролиферацию
зависят от реализации программы дифференци
(рис. 4). Гиперпролиферативная активность,
ровки кератиноцитов [70]. Показано, что вирус
стимулированная этими онкобелками, может
способен инфицировать «молодые» кератино
привести к репликативному стрессу и кластоге
циты базального слоя, возможно, активирован
незу, что также дестабилизирует геном [9].
ные кератиноциты в области повреждений эпи
30 летние исследования показали уникальную
телия [5, 9]. Именно повреждение эпителия
генетическую стабильность онкобелков ВПЧ
обеспечивает возможность сближения ВПЧ с
ВКР в неоплазиях и опухолях [67]. Исследовате
кератиноцитами базальной мембраны и связы
лям не удалось выявить взаимосвязи между раз
вание белка вирусного капсида L1 с внеклеточ
личающимися по нуклеотидным последователь
ным протеогликаном гепаран сульфатом, в ре
ностям вариантами ОРС Е6 и Е7 и различиями в
зультате которого происходит первичное при
степени интраэпителиальных поражений шейки
крепление вируса и его связывание с клетками
матки [68]. Уникальные канцерогенные свойства
базального слоя. После связывания капсида его
Е6 и Е7, их высокая консервативнось и консти
конформация изменяется, в результате протео
тутивная экспрессия в предраковых и злокачест
лиза, осуществляемого фурином, отщепляется
венных тканях делают их идеальными мишеня
N концевая часть белка L2. После этого проис
ми для вакцинной иммунотерапии ВПЧ ассоци
ходит второе изменение конформации капсида,
ированных предраковых состояний и рака [69].
в результате чего он получает возможность свя
Рис. 5. Схема развития папилломавирусной инфекции. Слева схематично представлен дифференцированный эпителий,
справа - экспрессируемые в инфицированных клетках белки ВПЧ. После инфицирования кератиноцитов базального
слоя вирус поддерживается в низкокопийном эписомальном состоянии. Белки Е6 и Е7 (красная стрелка) экспрессируют
ся с раннего промотора в нижних слоях эпителия (красные ядра). Синим цветом обозначены ядра неинфицированных
клеток. Экспрессия белка Е4, и, возможно, ряда других белков (зеленая стрелка), чьи ОРС лежат под поздним промото
ром, начинается раньше, чем прекращается экспрессия Е6 и Е7. Клетки, в которых экспрессируется Е4 (зеленые) и Е7 со
держат, скорее всего, все ранние белки, необходимые для репликации вирусной ДНК. Экспрессия белков капсида виру
са - L1 и L2 (оранжевая стрелка) следует за завершением амплификации геномной ДНК и происходит в клетках, экспрес
сирующих Е4 (на основе оригинального рисунка Zheng Z.M. и Baker C.C. [105], с модификациями)
БИОХИМИЯ том 84 вып. 7 2019
1006
ВОНСКИЙ и др.
зывания с рецепторами клетки, в частности с
ванных клетках наружного слоя эпителия [75].
интегринами α6 и α6β4, ламинин и коллаген
Высвобождение вирионов сопровождается ги
связывающими белками, задействованными в
белью дифференцированных клеток [44], что
адгезии кератиноцитов к дермису и диссемина
приводит к формированию койлоцитов [46].
ции эпителиальных клеток при заживлении ран
и канцерогенезе.
Последующая интернализация вируса в ва
ФОРМЫ ВПЧ ИНФЕКЦИИ
куолярные структуры осуществляется по меха
низму клатрин или кавеолин опосредованного
Инфекция ВПЧ может протекать в острой
эндоцитоза [71].
(не персистирующей, транзиторной или прехо
В течение длительного времени причиной
дящей), латентной или хронической формах.
ВПЧ индуцированных неоплазий считали ви
Острые, латентные и хронические инфекции
русную инфекцию клеток базального слоя сли
различаются по вирусной активности, экспрес
зистой оболочки. Однако, накопление данных,
сии вирусных и клеточных генов, влиянию на
подтверждающих критическую роль дискретной
динамику пролиферации клеток, способности
популяции клеток зоны перехода плоского эпи
вызывать локальную иммуносупрессию и онко
телия в цилиндрический в патогенезе прерако
генному потенциалу.
вых состояний и рака шейки матки, привело к
Генетический материал вируса может быть
пересмотру этой парадигмы [72]. В условиях
обнаружен после полового контакта с инфици
микроокружения экто эндоцервикальной зоны
рованным партнером в течение нескольких
перехода (ЭЭЗП) внутри или в непосредствен
дней, даже при отсутствии инфекции. После
ной близости от этой области присутствуют
инфицирования ДНК ВПЧ персистирует в эпи
уникальные эпителиальные клетки. Фенотипи
сомальной форме и в большинстве (до 90%) слу
чески близкие к клеткам плоскоклеточного
чаев элиминируется иммунной системой. Это
РШМ и его предшественника HSIL, они в высо
можно назвать «транзиторной инфекцией», хо
кой степени подверженны инфекции ВПЧ ВКР
тя термин «транзиторное обнаружение» может
[72].
быть более точным. Кроме того, в некоторых
Вследствие отсутствия собственных полиме
случаях ВПЧ может реплицироваться в инфи
раз, ВПЧ не способен к самостоятельной репли
цированных эпителиальных клетках и продуци
кации, что делает жизненный цикл вируса зави
ровать вирионы, однако, в конечном итоге, про
симым от клетки хозяина. После инфицирова
исходит спонтанная элиминация вируса
ния базальных клеток в области повреждения
(т.н. «острые инфекции») [76]. Однако в некото
происходит первый цикл репликации ДНК,
рых случаях происходит только кажущаяся эли
осуществляемый независимо от клеточного
минация вируса. Вирусные геномы сохраняются
цикла и амплифицирующий геном вируса до
в инфицированных клетках в неактивной форме
50-100 копий на клетку. Эта фаза инфекцион
(т.н. «латентные инфекции») [77]. В период ла
ного процесса характеризуется эписомальной
тентного носительства генетический материал
формой поддержания вирусного генома, при
ВПЧ может быть выявлен только случайно. Ре
которой дальнейшей репликации не происхо
активация латентной инфекции может про
дит - число геномов остается постоянным.
изойти через отдаленный период времени, на
Экспрессия белков вируса находится на мини
пример, вследствие иммунной супрессии, но за
мальном уровне под очень жестким контролем,
частую происходит и без каких либо видимых
продукты транскрипции вирусных генов, Е6 и
причин. Последовательное тип специфическое
Е7, в пролиферативном компартменте эпителия
определение вирусной нагрузки ВПЧ позволяет
практически не обнаруживаются.
дифференцировать регрессирующее поражение
Белки, кодируемые ранней областью генома
шейки матки и транзиторную вирион продуци
ВПЧ, индуцируют переход клетки в S фазу, что
рующую инфекцию [76].
обеспечивает вирус необходимыми ему полиме
Острые неэлиминированные инфекции, с
разами [73]. При попадании инфицированного
сохраняющейся с течением времени вирусной
кератиноцита в дифференцирующий компарт
активностью, классифицируют как «хроничес
мент, в шиповатый слой, выходящий из клеточ
кие инфекции» (хотя, со времением и они могут
ного цикла, происходит мощная активация
быть элиминированы естественным путем).
экспрессии как ранних, так и поздних вирусных
ВПЧ крайне успешен как инфекционный агент
генов, и репликации вирусной ДНК с увеличе
- вызванные им хронические инфекции прак
нием числа вирусных копий до многих тысяч
тически не имеют системных последствий и
копий на клетку [74]. Образование вирионов
редко убивают хозяина, который периодически
происходит только в полностью дифференциро
(в течение недель и месяцев) производит боль
БИОХИМИЯ том 84 вып. 7 2019
ВПЧ АССОЦИИРОВАНЫЙ КАНЦЕРОГЕНЕЗ
1007
шое количество инфекционных вирусных ча
свою активность. Они модулируют экспрессию,
стиц для передачи неинфицированным людям.
инактивируют гены супрессоры опухолей, ак
Возможны три варианта деления базальных
тивируют онкогены и индуцируют клеточную
клеток, инфицированных ВПЧ: (i) деление ин
пролиферацию, дополнительно повышая ге
фицированной клетки на две неделящиеся диф
номную нестабильность [83-85]. В совокупнос
ференцированные клетки; (ii) ассиметричное
ти, это приводит к развитию интраэпителиаль
деление на одну парабазальную и одну базаль
ных поражений: интраэпителиальным пораже
ную клетку; (iii) деление на две базальные клет
ниям шейки матки низкой или высокой степени
ки, при котором обе сохраняют способность к
(LSIL или HSIL) и РШМ, раку вульвы, влагали
делению (клональный путь ВПЧ трансформа
ща, полового члена, анальному раку, а также ра
ции). Цитологическое обнаружение LSIL в цер
ку головы, шеи и ротоглотки. Высокий уровень
викальном препарате является отличительным
гомологичности последовательностей ДНК
признаком продукции вирионов ВПЧ, а гало,
ВПЧ, обнаруженный при ВПЧ связанных рако
наблюдаемое при оптической микроскопии,
вых поражениях, а также в раковых тканях, ука
свидетельствует об аккумуляции нарабатывае
зывает на происхождение интегрированного
мых вирионов ВПЧ внутри дифференцирован
фрагмента от исходного инфицировавшего и
ных неделящихся клеток. Продукция в неделя
встроившегося в клетку вируса с дальнейшим
щихся клетках огромных количеств новых вири
размножением этой клетки, т.е. на клональность
онов гарантирует ВПЧ инфекцию новых кле
опухоли. В силу этого, высокий уровень гомоло
ток мишеней. Во всех этих случаях ДНК ВПЧ
гичности последовательностей ДНК ВПЧ явля
присутствует в эписомальной форме и может
ется одним из маркеров трансформации (хотя
упаковываться в вирионы с последующим их
формально и выглядит как консервативность
выходом из клетки и возможностью расшире
ВПЧ ВКР). В действительности же, увеличение
ния области инфекции [78].
вирусной нагрузки одного типа ВПЧ у женщин
В 10-12% случаев инфекция ВПЧ ВКР при
с инфекциями многими типами ВПЧ является
водит к интеграции вирусного генома в хромо
прогностическим признаком развития РШМ
сому клетки хозяина [79]. Интеграцию, наблю
[76].
даемую более чем в 70% ВПЧ ассоциированных
раков, считают ключевым событием канцероге
неза [80]. Она важна для трансформации клет
МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ
ки, но не является частью нормального цикла
ВПЧ@ИНДУЦИРОВАННОГО
жизни вируса [81]. Интересно, что интеграция
КАНЦЕРОГЕНЕЗА
во многом определяется присутствием в геноме
человека последовательностей, микрогомоло
Считают, что к необходимым условиям фор
гичных ВПЧ. При этом сайты интеграции ВПЧ
мирования необратимой неоплазии относятся:
в геноме челoвека расположены неравномерно:
1) активная первоначальная экспрессия ге
из 3667 сайтов, выявленных при исследовании
нов Е6 и Е7 ВПЧ ВКР, вызывающая в инфици
135 образцов клинического материала, более
рованной клетке генетическую нестабильность;
50% локализованы в «горячих точках» интегра
2) нарушение клеточного цикла, стимулиру
ции, в последовательностях генов POU5F1B
емое онкобелками Е6 и Е7, предотвращение
(9,7%), FHIT (8,7%), KLF12 (7,8%), HMGA2
апоптоза клеток с поврежденной ДНК, их им
(7,8%), KLF5 (6,8%), LRP1B (5,8%), LEPREL1
мортализация и трансформация;
(4,9%), DLG2 (4,9%), SEMA3D (4,9%) [82] (офи
3) стабильная экспрессия Е6 и Е7, защища
циальные названия генов приведены в соответ
ющая иммортализованные и трансформирован
ные клетки от апоптоза [86]. При этом времен
genecards.org/).
ное подавление экспрессии Е6 и Е7, например,
Вследствие этого, интеграция ВПЧ может
при гипоксии, несущественно. Гипоксия вызы
оказывать влияние на работу генов, в области
вает только временный арест пролиферации
которых произошло встраивание, как снижая,
клеток и роста опухоли, приводящий к утрате
так и повышая их экспрессию.
ответа на химиотерапию, ориентированную на
После интеграции, механизм которой вклю
подавление пролиферирующих клеток, и уход
чает потерю части вирусного генома, интегри
от иммунного контроля, специфичного к Е6/Е7,
рованная ДНК ВПЧ теряет свою инфекцион
с восстановлением пролиферации и роста после
ность и уже не может быть упакована в инфек
восстановления оксигенации [87];
ционные вирионы. При этом онкобелки ВПЧ
4) гормональная регуляция формирования и
Е6 и Е7, кодирующие последовательности кото
«поддержания» неопластических изменений
рых интегрируются в геном хозяина, сохраняют
тканей.
БИОХИМИЯ том 84 вып. 7 2019
1008
ВОНСКИЙ и др.
Индуцированные онкобелками ВПЧ нару
ством к эстрогеновым рецепторам, взаимодей
шения генетического аппарата клетки проявля
ствуя с ними, он во многом модулирует метабо
ются на ранних стадиях пре неопластических
литическую и пролиферативную активность
изменений, когда вирус еще находится в эписо
клеток. Исследование EPIC, в которое были
мальной форме [81]. Онкобелки Е6 и Е7 ВПЧ
вовлечены 308 036 женщин, включая 261 паци
ВКР индуцируют геномную нестабильность, яв
ентку с РШМ и 804 - с CIN3 (HSIL), показало
ляющуюся характерной чертой раковых клеток,
значительное повышение содержания эстрадио
характеризующихся геномными перестройками
ла в сыворотках крови пациенток с РШМ [91].
и генной амплификацией. Генетическая неста
Интересно, что сами клетки ВПЧ ассоцииро
бильность должна индуцировать апоптоз, одна
ванных опухолей не чувствительны к действию
ко он тормозится по нескольким параллельным
эстрогенов, т.к. не экспрессируют соответству
независимым механизмам, также опосредован
ющий рецептор, экспрессия эстрогенов значи
ным белками Е5, Е6 и Е7 (см. соответствующие
тельно повышена в перитуморозных тканях,
разделы).
поддерживающих рост опухоли [92].
Прямым следствием геномной нестабиль
ности, индуцированной онкобелками, является
интеграция вируса [9]. Интеграция имеет целый
ПРОФИЛАКТИЧЕСКИЕ ВПЧ ВАКЦИНЫ
ряд негативных последствий. Этот процесс час
И ВОЗМОЖНОСТЬ
то сопровождается делетированием ранних (E1,
ПРЕДОТВРАЩЕНИЯ РАКА
E2, E4 и E5) и поздних (L1 и L2) ОРС, но сохра
няет практически интактными ранние ОРС
ВПЧ инфекция часто встречается в половых
ВПЧ E6 и E7. После делеции L1 и L2 клетки, ин
путях у молодых сексуально активных людей.
фицированные ВПЧ, более не могут быть рас
Большинство из тех, у кого развиваются добро
познаны иммунной системой у лиц, получив
качественные ВПЧ ассоциированные измене
ших профилактическую вакцинацию. В силу
ния тканей, в конечном итоге вырабатывают
этого, интеграция вируса, сопровождаемая де
эффективный клеточный иммунный ответ,
лецией L1 и L2, делает бессмысленными попыт
приводящий к регрессии папиллом и кандилом
ки применения профилактических вакцин в те
и элиминации вируса. Однако в целом ряде слу
рапевтических целях. Происходящая при инте
чаев, особенно при иммуносупрессии, связан
грации делеция E2, негативного регулятора
ной, например, с ВИЧ инфекцией, элиминации
экспрессии E6/E7 [88], приводит к повышению
вируса не происходит. Персистенцию ВПЧ свя
экспрессии E6/E7, что способствует злокачест
зывают с его способностью к эффективному ук
венной трансформации клеток [89]. Кроме того,
лонению от врожденного иммунного ответа,
интеграция ВПЧ часто происходит в области
что задерживает развитие адаптивного иммуни
промотора гена TERT (5q15) или локуса 3q26,
тета и препятствует элиминации вируса на эта
что приводит к обильной экспрессии гена тело
пах, предшествующих неопластическим изме
меразы TERC [82] и иммортализации клеток,
нениям. Модификации иммунной системы,
несущих фрагменты генома ВПЧ, что способ
индуцируемые ВПЧ инфекцией, включают
ствует их неконтролируемой амплификации.
дифференцировку опухолеассоциированных
При этом надо отметить, что интегрированные
макрофагов (ТАМ), подавление активации и
нуклеотидные последовательности генома ВПЧ
созревания дендритных клеток и, как след
выявлены только в 62% всех образцов церви
ствие, нарушения клеточного иммунного отве
кальных карцином [90]. Хотя и была показана
та, включая аномальный дисбаланс между
связь между повышенной частотой интеграции
T хелперными клетками типа 1 (Th1) и 2 (Th2)
генома ВПЧ в случаях более выраженных пре
и индукцию регуляторных Т клеток [93]. Нес
неопластических изменений, вирусную инте
пособность выработать эффективный клеточ
грацию предлагают считать следствием хромо
ный иммунный ответ против ВПЧ приводит к
сомной нестабильности, вызванной действием
персистирующей инфекции. В случае «задерж
онкобелков, а не первопричиной злокачествен
ки» в организме ВПЧ ВКР высока вероятность
ной трансформации [9].
того, что за время персистенции, даже транзи
Основные молекулярные механизмы первых
торной, вирус успеет трансформировать эпите
трех процессов, регулируемых Е6 и Е7, были
лиальные клетки. На этом этапе элиминация
рассмотрены выше. Остановимся на последнем.
вируса иммунной системой уже не сможет пре
Шейка матки относится к эстрогеночувстви
дотвратить развития рака, что, с одной сторо
тельным тканям для неопластических процес
ны, указывает на необходимость своевременно
сов, в которых важную роль играют эстрогены, в
го проведения профилактической ВПЧ вакци
том числе - эстрадиол. Обладая высоким срод
нации, с другой, на необходимость разработки
БИОХИМИЯ том 84 вып. 7 2019
ВПЧ АССОЦИИРОВАНЫЙ КАНЦЕРОГЕНЕЗ
1009
терапевтических ВПЧ вакцин, способных ин
филогенетически родственны ВПЧ 16 го и 18 го
дуцировать клеточный иммунный ответ против
типа [96]. Продукция белка реализована в дрож
ВПЧ и элиминировать из организма ВПЧ ин
жах Saccharomyces cerevisiae, в качестве адьюван
фицированные клетки.
та использован гидроксифосфат сульфат алю
Исследования природной ВПЧ инфекции у
миния [97]. Уже во втором квартале 2007 г. эта
животных показали, что нейтрализующие анти
вакцина была разрешена к применению в 80
тела к белку вирусной оболочки L1 являются за
странах мира. При использовании, в соответ
щитными, их индукция может быть положена в
ствии с рекомендациями производителя,
основу эффективной стратегии профилактичес
Gardasil® обеспечивает эффективность 98% в за
кой вакцинации. Данные о том, как антитела
щите женщин от данных типов ВПЧ, элимини
против ВПЧ предотвращают инфекцию базаль
руя риск HSIL, плоскоклеточных цервикальных
ных эпителиальных клеток дают механистичес
карцином, аденокарцином и иных осложнений,
кое объяснение эффективности таких вак
обусловленных ВПЧ инфекцией [98].
цин [94]. Перед профилактическими ВПЧ вак
В 2008 г. была зарегистрирована также бива
цинами были поставлены задачи формирования
лентная вакцина Cervarix®, содержащая два типа
протективных антител, нарабатываемых в высо
капсидных белков L1 ВПЧ
16 и
18
ком титре и способных связать вирус до его по
(«GlaxoSmithKline», London, UK). Вакцина про
падания в эпителиальные клетки базального
дуцируется в инфицированных L1 рекомбина
слоя, а также генерации В клеточной памяти
нтным бакуловирусом клетках Spodoptera
для обеспечения длительного присутствия анти
frugiperda SF9 и Trichoplusia ni Hi 5 и использует в
тел, обеспечивающих локальную нейтрализа
качестве адьюванта AS04, включающий гидрок
цию вируса в течение периода половой актив
сид алюминия и 3 деацетилированный моно
ности иммунизированного человека. При этом
фосфорил липид А (детоксифицированную
иммунные корреляты защиты от ВПЧ - уровни
форму липосахарида - агониста TLR4). Эффек
антител, которые бы коррелировали со способ
тивность Cervarix® в предотвращении ВПЧ ас
ностью организма противостоять ВПЧ инфек
социированных поражений CIN3+ сохраняется
ции, как в случае инфекции вирусом гепатита В,
в течение >10 лет. Однако, если на момент вак
отсутствуют [94]. В отличие от профилактичес
цинации вакцинируемый уже был инфицирован
ких, терапевтические ВПЧ вакцины должны
ВПЧ 16, вакцина оказывается неэффективной
быть направлены против белков ВПЧ, экспрес
[99].
сирующихся в уже зараженных, особенно, в
В 2015 году Консультативным Комитетом по
трансформированных ВПЧ клетках, а именно,
Практикам Иммунизации (ACIP) была реко
белков Е6 и Е7, и вызывать клеточный иммун
мендована к применению нонавалентная вак
ный ответ, способный идентифицировать и эф
цина Gardasil® 9 («Merck & Co.», Darmstadt,
фективно элиминировать эти клетки, прежде
Germany). Gardasil® 9 обеспечивает протектив
всего в эпителии.
ный иммунитет к девяти типам ВПЧ: 6, 11, 16,
Существующие на данный момент коммер
18, 31 33, 45, 52 и 58. Как и в случае Gardasil®,
ческие профилактические вакцины против ВПЧ
производство рекомбинантных форм вариантов
основаны на использовании антигенов в составе
белка капсида L1, входящих в Gardasil® 9, реали
большого белка капсида L1, организуемых в
зуется в клетках дрожжей Saccharomyces cerevisi
процессе самосборки в вирус подобные части
ae с гидроксифосфат сульфатом алюминия в ка
цы [95]. В 2006 г. Управление по контролю за
честве адьюванта. Считается, что применение
продуктами питания и лекарственными сред
Gardasil® 9 позволит снизить риск генитальных
ствами США (Food and Drug Administration,
папиллом на 90%, а ВПЧ ассоциированных ра
FDA) зарегистрировало первую в мире квадри
ков шейки матки и анального канала - на 89%.
валентную вакцину Gardasil® («Merck & Co.»,
Важно отметить, что все профилактические вак
Darmstadt, Germany). Gardasil® продемонстри
цины против ВПЧ вводятся внутримышечно,
ровал эффективность в предупреждении инфи
чтобы принципиально обойти действие потен
цирования ведущими типами ВПЧ - ВПЧ ВКР
циально заложенных в вирусных антигенах мо
16 и 18, а также вызывающими генитальные па
тивов, обуславливающих уход от иммунного от
пилломы и рецидивирующий респираторный
вета в эпителии.
папилломатоз типами ВПЧ 6 и 11, и, кроме того,
Имеются все основания утверждать, что вак
частичную перекрестную защиту против пер
цины против ВПЧ станут вторыми по эффек
систирующей инфекции и цервикальных пора
тивности противовирусными противораковыми
жений, вызванных десятью невакцинными ти
вакцинами после вакцины против вирусного ге
пами ВПЧ высокого риска, включая ВПЧ 31 го,
патита В. ВПЧ вакцины уже показали себя, как
33 го, 35 го, 45 го, 52 го и 58 го типа, которые
мощный инструмент защиты от ВПЧ ассоци
БИОХИМИЯ том 84 вып. 7 2019
1010
ВОНСКИЙ и др.
ированных неоплазий. Мета анализ результатов
смотря на успех АРТ, распространенность ВПЧ
26 испытаний моновалентных (N = 1), двухвале
инфекции, интраэпителиальных поражений
нтных (N = 18) и четырехвалентных вакцин
низкого и высокого уровня, а также рака шейки
(N = 7) на, в общей сложности, 73 428 участни
матки остается стабильно высокой, а заболевае
ках, убедительно доказал, что вакцины против
мость соответствующими анальными форма
ВПЧ эффективно защищают от предраковых
ми - даже возрастает [37]. Больные ВИЧ, как и
поражений шейки матки девочек подростков и
люди с различными формами иммуносупресии,
молодых женщин в возрасте 15-26 лет, особен
нуждаются в обязательной вакцинации против
но при отборе участников, изначально неинфи
инфекции ВПЧ ВКР. Фармакоэкономическое
цированных ВПЧ ВКР. При оценке поражений
исследование показало, что профилактическая
защитный эффект оказался выше для пораже
вакцинация против ВПЧ в РФ при затратах в
ний, связанных с инфекцией ВПЧ 16/18, чем
3,4 млрд рублей могла бы сэкономить потери в
для поражений, вызыванных ВПЧ неопределен
19,4 млрд рублей, связанные с потерей трудос
ного типа. Были получены достоверные доказа
пособности и лечением больных с различными
тельства того, что вакцины против ВПЧ снижа
формами ВПЧ ассоциированных раков [104].
ют риск развития плоскоклеточных интраэпите
Одна только вакцинация против ВПЧ детей и
лиальных поражений высокой степени у пожи
подростков, живущих с ВИЧ, как одной из наи
лых женщин с изначально отрицательной реак
более уязвимых групп, позволила бы предотвра
цией на ВПЧ 16/18 [3].
тить до 50 000 случаев рака. Таким образом, про
Профилактическая ВПЧ вакцинация - гро
филактическая вакцинация против ВПЧ могла
мадный шаг вперед к предотвращению других
бы дать огромный социоэкономический эф
ВПЧ индуцированных неопластических изме
фект.
нений, в частности, пениса и анального канала у
мужчин, как за счет коллективного (популяци
Папилломавирусная инфекция считается
онного) иммунитета (heard effect), так и за счет
причиной возникновения 5% раковых заболева
расширяющейся практики прививок мальчиков
ний и 30% всех раков, вызванных инфекцион
и молодых людей [100, 101]. Однако, проведен
ными агентами. В настоящем обзоре рассмотре
ные к настоящему времени исследования недо
ны классификация ВПЧ, организация генома и
статочно объемны и недостаточно продолжи
белки, входящие в состав вируса, особенности
тельны, чтобы достоверно оценить уровень сни
репликации вируса и формы ВПЧ инфекции,
жения как РШМ, так и других форм ВПЧ ассо
обсуждены молекулярные механизмы ВПЧ ин
циированного рака [3]. Также трудно адекватно
дуцированного канцерогенеза. Отдельно рас
оценить косвенные эффекты вакцинации, в
смотрены распространенность ВПЧ и ВПЧ ас
частности, защиту от ВПЧ инфекции и ВПЧ
социированных форм рака, механизмы инфи
ассоциированных неоплазий мужчин, контак
цирования и пути передачи инфекции особен
тирующих с ВПЧ вакцинированными женщи
ности течения ВПЧ ассоциированных заболе
нами, и детей, рождающихся у ВПЧ вакцини
ваний на фоне иммуносупрессии.
рованных женщин.
Особое внимание в обзоре уделено профи
В странах же, не внедривших скрининг ра
лактической вакцинации и возможностям пре
ков шейки матки и анального канала и програм
дотвращения ВПЧ ассоциированного рака.
мы профилактической ВПЧ вакцинации под
Профилактическая вакцинация против ВПЧ,
ростков, частота выявления форм рака, ассоци
успехи которой отражены в обзоре, должна про
ированного с инфекцией ВПЧ ВКР, остается
водиться в преподростковый период, до того,
высокой. Особенно это касается лиц с различ
как произойдет инфицирование ВПЧ. Поздняя
ными формами иммунодефицитов. Группой по
вакцинация не может повлиять на персистен
Детской Онкологии выработаны рекомендации
цию ВПЧ у уже инфицированного индивида и
по ВПЧ вакцинации всех женщин с дефектами
не может предотвратить ВПЧ ассоциированные
иммунной системы, перенесших рак в детском
заболевания, а также горизонтальное и верти
возрасте [102]. У ВИЧ инфицированных лиц до
кальное ВПЧ инфицирование младенцев и де
70% раков вызваны инфекцией ВПЧ ВКР. Акти
тей до достижения ими возраста вакцинации.
вация/истощение Т клеток и миелоидных кле
Даже путем массовой вакцинации ВПЧ сложно
ток у ВИЧ инфицированных людей, получаю
добиться искоренения ВПЧ инфекции в попу
щих АРТ, при инфицировании ВКР типами
ляции как в ближайшем, так и в достаточно от
ВПЧ усиливается, что свидетельствует о вкладе
даленном будущем, что делает необходимым
ВПЧ в дисфункцию иммунных клеток [103].
проведение интенсивных работ по созданию те
Как было отмечено выше, АРТ не снижает рис
рапевтических вакцин против ВПЧ и ВПЧ ас
ков развития неоплазий и рака. [37, 103]. Не
социированных неоплазий.
БИОХИМИЯ том 84 вып. 7 2019
ВПЧ АССОЦИИРОВАНЫЙ КАНЦЕРОГЕНЕЗ
1011
Финансирование. Работа выполнена при фи
Конфликт интересов. Авторы заявляют об от
нансовой поддержке Российского фонда фунда
сутствии конфликта интересов.
ментальных исследований (гранты № 17_54_30002
Соблюдение этических норм. Настоящая
и 17_04_00583) и гранта #1R01CA217715 01
статья не содержит описания выполненных ав
Национальных Институтов Здоровья США
торами исследований с участием людей или ис
(NIH).
пользованием животных в качестве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Shope, R.E., and Hurst, E.W. (1933) Infectious papillo
14.
Heitmann, E., and Harper, D. (2012) Prophylactic HPV
matosis of rabbits, with a note on histopathology, J. Exp.
vaccines and the prevention of cervical intraepithelial neo
Med., 58, 607-624.
plasia, Cur. Obstet. Gynecol. Rep., 1, 95-105, doi: 10.1007/
2.
Bray, F., Ferlay, J., Soerjomataram, I., Siegel, R.L.,
s13669 012 0017 4.
Torre, L.A., and Jemal, A. (2018) Global cancer statistics
15.
Husain, R.S., and Ramakrishnan, V. (2015) Global varia
2018: GLOBOCAN estimates of incidence and mortality
tion of human papillomavirus genotypes and selected genes
worldwide for 36 cancers in 185 countries, CA Cancer
involved in cervical malignancies, An. Glob. Health, 81,
J. Clin., 68, 394-424, doi: 10.3322/caac.21492.
675-683, doi: 10.1016/j.aogh.2015.08.026.
3.
Arbyn, M., Xu, L., Simoens, C., and Martin Hirsch P.P.
16.
Mbulawa, Z.Z.A., van Schalkwyk, C., Hu, N.C., Meiring, T.L.,
(2018) Prophylactic vaccination against human papillo
Barnabas, S., Dabee, S., Jaspan, H., Kriek, J.M.,
maviruses to prevent cervical cancer and its precursors,
Jaumdally, S.Z., Muller, E., Bekker, L.G., Lewis, D.A.,
Cochrane Database Syst. Rev., 5, CD009069, doi: 10.1002/
Dietrich, J., Gray, G., Passmore, J.S., and Williamson, A.L.
14651858.CD009069.pub3.
(2018) High human papillomavirus (HPV) prevalence in
4.
Osazuwa Peters, N., Massa, S.T., Simpson, M.C., Adjei
South African adolescents and young women encourages
Boakye, E., and Varvares, M.A. (2018) Survival of human
expanded HPV vaccination campaigns, PLoS One, 13,
papillomavirus associated cancers: Filling in the gaps,
e0190166, doi: 10.1371/journal.pone.0190166.
Cancer, 124, 18-20, doi: 10.1002/cncr.30945.
17.
Nweke, М.С., Okolo, С.А., Daous, Y., and Esan, О.А.
5.
McBride, A.A. (2017) Mechanisms and strategies of papil
(2018) Challenges of human papillomavirus infection and
lomavirus replication, Biol. Chem.,
398,
919-927,
associated diseases in low resource countries, Arch. Pathol.
doi: 10.1515/hsz 2017 0113.
Lab. Med., 142, 696-699, doi: 10.5858/arpa.2017 0565
6.
IARC (2007). Human papillomaviruses. IARC Monogr.
RA.
Eval. Carcinog. Risks Hum., 90, 1-636.
18.
Clifford, G.M., Tully, S., and Franceschi, S.
(2017)
7.
Khallouf, H., Grabowska, A.K., and Riemer, A.B. (2014)
Carcinogenicity of human papillomavirus (HPV) types in
Therapeutic vaccine strategies against human papillomavirus,
HIV positive women: a meta analysis from HPV infection
Vaccines, 2, 422-462, doi: 10.3390/vaccines2020422.
to cervical cancer, Clin. Infect. Dis., 64, 1228-1235,
8.
Solomon, D., Davey, D., Kurman, R., Moriarty, A.,
doi: 10.1093/cid/cix135.
O’Connor, D., Prey, M., Raab, S., Sherman, M.,
19.
Лопухов П.Д. (2018) Научно методическое обоснование
Wilbur, D., Wright, T.Jr., and Young, N. (2018) The 2001
оптимизации эпидемиологического надзора и профилак
Bethesda System: terminology for reporting results of cer
тики папилломавирусной инфекции. Дис. канд. мед. на
vical cytology, JAMA, 287, 2114-2119, doi: 10.1001/
ук, Москва.
jama.287.16. 2114.
20.
Петрова Г.В., Грецова О.П., Шахзадова А.О., Прос
9.
Araldi, R., Sant’Ana, T.A., Modolo, D.G., de Melo, T.C.,
тов М.Ю., Простов Ю.И., Самсонов Ю.В. (2018) В кн.
Spadacci Morena, D.D., de Cassia Stocco, R.,
Злокачественные образования в России в 2017 г. Заболе
Cerutti, J.M., and de Souza, E.B. (2018) The human
ваемость и смертность (под ред. Каприна А. Д., Ста
papillomavirus (HPV) related cancer biology: an overview,
ринского В. В., Петровой Г.В.), ФГБУ «МНИОИ им.
Biomed. Pharmacother., 106, 1537-1556, doi: 10.1016/
П.А. Герцена», Москва, с. 4-130.
j.biopha.2018.06.149.
21.
Роговская С.И., Михеева И.В., Шипулина О.Ю.,
10.
De Vincenzo, R., Ricci, C., Conte, C., and Scambia, G.
Минкина Г.Н., Подзолкова Н.М., Радзинский В.Е.,
(2013) HPV vaccine cross protection: highlights on addi
Шипулин Г.А. (2012) Распространенность папиллома
tional clinical benefit, Gynecol. Oncol., 130, 642-651, doi:
вирусной инфекции в России, Эпидемиология и вакци
10.1016/j.ygyno.2013.05.033.
нопрофилактика, 1, 25-33.
11.
Bruni, L., Albero, G., Serrano, B., Mena, M., Gomez, D.,
22.
Касихина Е.И. (2011) Папилломавирусная инфекция
Munoz, J., Bosch, F.X., and de Sanjose, S. (2019) Human
сегодня: клиническое разнообразие, лечение и про
Papillomavirus and Related Diseases in the World. Summary
филактика, Лечащий врач, 10, 6-8.
Report 22 January 2019. ICO/IARC Information Centre
23.
Depuydt, C.E., Beert, J., Bosmans, E., and Salembier, G.
on HPV and Cancer (HPV Information Centre).
(2016) Human papillomavirus (HPV) virion induced can
12.
Wang, C.J., Sparano, J., and Palefsky, J.M. (2017) Human
cer and subfertility, two sides of the same coin, Facts Views
Immunodeficiency Virus/AIDS, Human Papillomavirus,
Vis. Obgyn., 8, 211-222.
and Anal Cancer, Surg. Oncol. Clin. N. Am., 26, 17-31,
24.
Lee, В., Lee, S.W., and Kim, D.I. (2017) HPV prevalence
doi: 10.1016/j.soc.2016.07.010.
in the foreskins of asymptomatic healthy infants and chil
13.
Chesson, H.W., Dunne, E.F., Hariri, S., and Markowitz, L.E.
dren: systematic review and meta analysis, Sci. Rep., 7,
(2014) The estimated lifetime probability of acquiring
7050, doi: 10.1038/s41598 017 07506 z.
human papillomavirus in the United States, Sex. Transm.
25.
Rintala, M.A., Grenman, S.E., Puranen, M.H., Isolauri, E.,
Dis., 41, 660-664, doi: 10.1097/OLQ.0000000000000193.
Ekblad, U., Kero, P.O., and Syrjanen, S.M.
(2005)
БИОХИМИЯ том 84 вып. 7 2019
1012
ВОНСКИЙ и др.
Transmission of high risk human papillomavirus (HPV)
39.
Palefsky, J.M., and Holly, E.A. (2003) Immunosuppression
between parents and infant: a prospective study of HPV in
and co infection with HIV 1, J. Natl. Cancer Inst. Monogr.,
families in Finland, J. Clin. Microbiol., 43, 376-381,
31, 41-46, doi: 10.1093/oxfordjournals.jncimonographs.
doi: 10.1128/JCM.43.1.376 381.2005.
a003481.
26.
Smith, E.M., Parker, M.A., Rubenstein, L.M., Haugen, T.H.,
40.
Ghebre, R.G., Grover, S., Xu, M.J., Chuang, L.T., and
Hamsikova, E., and Turek, L.P. (2010) Evidence for verti
Simonds, H. (2017) Cervical cancer control in HIV infect
cal transmission of HPV from mothers to infants, Infect.
ed women: past, present and future, Gynecol. Oncol. Rep.,
Dis. Obstet. Gynecol., 2010, 326389, doi: 10.1155/2010/
21, 101-108, doi: 10.1016/j.gore.2017.07.009.
326369.
41.
Paramsothy, P., Jamieson, D.J., and Heilig, C.M. (2009)
27.
Sarkola, M., Rintala, M., Grйnman, S., and Syrjanen, S.
The effect of highly active antiretroviral therapy on
(2008) Human papillomavirus DNA detected in breast
human papillomavirus clearance and cervical cytology,
milk, Pediatr. Infect. Dis. J.,
27,
557-558, doi:
Obstet. Gynecol.,
113,
26-31, doi:
10.1097/AOG.
10.1097/INF.0b013e318169ef47.
0b013e31819225cb.
28.
Teixeira, L.O., Amaral, S.C., Finger Jardim, F., da Hora, V.P.,
42.
Kelly, H., Weiss, H.A., Benavente, Y., de Sanjose, S., and
Gonçalves, C.V., Soares, M.A., and de Martinez, A.M.B.
Mayaud, P., ART and HPV Review Group
(2018)
(2015) Frequencia do papilomavirus humano na placenta,
Association of antiretroviral therapy with high risk human
no colostro e no sangue do cordao umbilical, Rev. Bras.
papillomavirus, cervical intraepithelial neoplasia, and
Ginecol. Obstet., 37, 203-207, doi: 10.1590/SO100
invasive cervical cancer in women living with HIV: a sys
720320150005293.
tematic review and meta analysis. Lancet HIV, 5, e45-e58.
29.
Green, G.E., Bauman, N.M., and Smith, R.J. (2000)
doi: 10.1016/S2352 3018(17)30149 2.
Pathogenesis and treatment of juvenile onset recurrent res
43.
Harden, M.E., and Munger, K. (2017) Human papillo
piratory papillomatosis, Otolaryngol. Clin. North. Am., 33,
mavirus molecular biology, Mutat. Res. Rev. Mutat. Res.,
187-207.
772, 3-12, doi: 10.1016/j.mrrev.2016.07.002.
30.
Liu, Z., Tasnuva, R., and Nyitray, A. (2015) Penises not
44.
Buck, C., Day, P., and Trus, B. (2013) The papillomavirus
required: a systematic review of the potential for human
major capsid protein L1, Virology,
445,
169-174,
papillomavirus horizontal transmission that is non sexual
doi: 10.1016/j.virol.2013.05.038.
or does not include penile penetration, Sex. Health, 13,
45.
Ustav, M., Ustav, E., Szymanski, P., and Stenlund, A.
10-21, doi: 10.1071/SH15089.
(1991) Identification of the origin of replication of bovine
31.
Зароченцева В., Белая Ю.М., Самсыгина Г.А., Щерба
papillomavirus and characterization of the viral origin
кова М.Ю., Выжлова Е.Н., Малиновская В.В. (2017)
recognition factor E1, EMBO J., 10, 4321-4329.
Папилломавирусная инфекция и ВПЧ ассоциирован
46.
Ferraro, C., Canedo, M., Oliveira, S., Carvalho, M., and
ные заболевания, Лечащий врач, 4, 56-63.
Dias, E. (2011) Infeccao oral pelo HPV e lesoes epiteliais
32.
Castellsague, X., Drudis, T., Canadas, M.P., Gonce, A.,
proliferativas associadas, J. Bras. Patol. Med. Lab., 47,
Ros, R., Perez, J.M., Quintana, M.J., Munoz, J., Albero, G.,
451-459.
de Sanjose, S., and Bosch, F.X. (2009) Human papillo
47.
Wallace, N.A., and Galloway, D.A. (2014) Manipulation of
mavirus (HPV) infection in pregnant women and mother -
cellular DNA damage repair machinery facilitates propa
to - child transmission of genital HPV genotypes: a
gation of human papillomaviruses, Semin. Cancer Biol., 26,
prospective study in Spain, BMC Infect. Dis., 9, 74, doi:
30-42, doi: 10.1016/j.semcancer.2013.12.003.
10.1186/1471 2334 9 74.
48.
Schuck, S., and Stenlund, A.A. (2015) Conserved regula
33.
Palefsky, J.M. (1998) Human papillomavirus infection and
tory module at the C terminus of the papillomavirus E1
anogenital neoplasia in human immunodeficiency virus
helicase domain controls E1 helicase assembly, J. Virol.,
positive men and women, J. Natl. Cancer Inst. Monogr., 23,
89, 1129-1142, doi: 10.1128/JVI.01903 14.
15-20.
49.
Helfer, C.M., Yan, J., and You, J. (2014) The cellular bro
34.
Рассохин В.В., Леонова О.Н., Пантелеева О.В., Смир
modomain protein Brd4 has multiple functions in E2
нова Н.Л., Фоменкова Н.В., Загдын З.М., Беляков
mediated papillomavirus transcription activation, Viruses,
Н.А. (2012) Частота и характер онкологических забо
6, 3228-3249, doi: 10.3390/v6083228.
леваний у больных с ВИЧ инфекцией до и на фоне
50.
Doorbar, J. (2017) Host control of human papillomavirus
применения высокоактивной антиретровирусной те
infection and disease, Best Pract. Res. Clin. Obstet.
рапии, ВИЧ инфекция и иммуносупрессии, 4, 34-43.
Gynaecol.,
47,
27-41, doi:
10.1016/j.bpobgyn.2017.
35.
Palefsky, J.M., Holly, E.A., Ralston, M.L., Da Costa, M.,
08.001.
and Greenblatt, R.M. (2001) Prevalence and risk factors
51.
Hoppe Seyler, K., Bossler, F., and Braun, J.A. (2018)
for anal HPV infection in HIV positive and high risk HIV
Down regulation of HPGD by miR 146b 3p promotes
negative women, J. Infect. Dis., 183, 383-391, doi:
cervical cancer cell proliferation, migration and anchor
10.1086/318071.
age independent growth through activation of STAT3 and
36.
Frisch, M., Biggar, R.J., and Goedert, J.J. (2000) Human
AKT pathways, Cell Death Dis., 9, 1055, doi: 10.1038/
papillomavirus associated cancers in patients with human
s41419 018 1059 y.
immunodeficiency virus infection and acquired immuno
52.
Muller, M., Prescott, E.L., Wasson, C.W., and Macdonald, A.
deficiency syndrome, J. Natl. Cancer Inst., 92, 1500-1510.
(2015) Human papillomavirus E5 oncoprotein: function
37.
Palefsky, J.M. (2017) Human papillomavirus associated
and potential target for antiviral therapeutics, Future
anal and cervical cancers in HIV infected individuals: inci
Virology, 10, 27-39, doi: 10.2217/fvl.14.99.
dence and prevention in the antiretroviral therapy era,
53.
Wetherill, L.F., Holmes, K.K., Verow, M., Muller, M.,
Curr. Opin. HIV AIDS,
12,
26-30, doi:
10.1097/
Howell, G., Harris, M., Fishwick, C., Stonehouse, N.,
COH.0000000000000336.
Foster, R., Blair, G.E., Griffin, S., and Macdonald A.
38.
Adler, D.H., Kakinami, L., Modisenyane, T., Tshabangu, N.,
(2012) High Risk Human Papillomavirus E5 Oncoprotein
Mohapi, L., De Bruyn, G., Martinson, N.A., and Omar, T.
Displays Channel Forming Activity Sensitive to Small
(2012) Increased regression and decreased incidence of
Molecule Inhibitors, J. Virol.,
86,
5341-5351,
human papillomavirus related cervical lesions among
doi: 10.1128/JVI.06243 11.
HIV infected women on HAART, AIDS, 26, 1645-1652,
54.
DiMaio, D., and Petti, L.M. (2013) The E5 proteins,
doi: 10.1097/QAD.0b013e32835536a3.
Virology, 445, 99-114, doi: 10.1016/j.virol.2013.05.006.
БИОХИМИЯ том 84 вып. 7 2019
ВПЧ АССОЦИИРОВАНЫЙ КАНЦЕРОГЕНЕЗ
1013
55.
Kivi, N., Greco, D., Auvinen, P., and Auvinen, E. (2008)
70.
Doorbar, J. (2005) The papillomavirus life cycle, J. Clin.
Genes involved in cell adhesion, cell motility and mito
Virol., 32, S7-S15, doi: 10.1016/j.jcv.2004.12.006.
genic signaling are altered due to HPV 16 E5 protein
71.
DiGiuseppe, S., Bienkowska Haba, M., Guion, L.G., and
expression, Oncogene, 27, 2532-2541, doi: 10.1038/sj.
Sapp, M. (2016) Cruising the cellular highways: how
onc.1210916.
human papillomavirus travels from the surface to the
56.
Venuti, A., Paolini, F., Nasir, L., Corteggio, A., Roperto,
nucleus, Virus Res., 231, 1-9, doi: 10.1016/j.virus
S., Campo, M.S., and Borzacchiello, G.
(2011)
res.2016.10.015.
Papillomavirus E5: the smallest oncoprotein with many
72.
Herfs, M., Soong, T.R., Delvenne, P., and Crum, C.P.
functions, Mol. Cancer, 10, 140, doi: 10.1186/1476 4598
(2017) Deciphering the multifactorial susceptibility of
10 140.
mucosal junction cells to HPV infection and related car
57.
Вонский М.С., Рунов А.Л., И.В. Гордейчук И.В., Иса
cinogenesis, Viruses, 9, E85, doi: 10.3390/v9040085.
гулянц М.Г. (2019) Терапевтические вакцины против
73.
Moody, C., and Laimins, L. (2010) Human papillomavirus
вирусов папилломы человека, достижения и перспек
oncoproteins: pathways to trans formation, Nat. Rev.
тивы, Биохимия, 84, 1016-1035.
Cancer, 10, 550-560, doi: 10.1038/nrc2886.
58.
Hengstermann, A., Linares, L.K., Ciechanover, A.,
74.
Stanley, M.A. (2012) Epithelial cell responses to infection
Whitaker, N.J., and Scheffner, M. (2001) Complete switch
with human papillomavirus, Clin. Microbiol. Rev., 25,
from Mdm2 to human papillomavirus E6 mediated degra
215-230, doi: 10.1128/CMR.05028 11.
dation of p53 in cervical cancer cells, Proc. Natl. Acad. Sci.
75.
Munday, J. (2014) Papillomaviruses in felids, Vet. J., 199,
USA, 98, 1218-1223, doi: 10.1073/pnas.98.3.1218.
340-347, doi: 10.1016/j.tvjl.2013.11.025.
59.
Tomaic, V. (2016) Functional roles of E6 and E7 oncopro
76.
Depuydt, C.E., Thys, S., Beert, J., Jonckheere, J.,
teins in HPV induced malignancies at diverse anatomical
Salembier, G., and Bogers, J.J. (2016) Linear viral load
sites, Cancers (Basel), 8, E95, doi: 10.3390/cancers8100095.
increase of a single HPV type in women with multiple
60.
Boon, S.S., Tomaic, V., Thomas, M., Roberts, S., and
HPV infections predicts progression to cervical cancer, Int.
Banks, L. (2015) Cancer causing human papillomavirus
J. Cancer, 139, 2021-2032, doi: 10.1002/ijc.30238.
E6 proteins display major differences in the phospho regu
77.
Brossfield, J.E., Chan, P.J., Patton, W.C., and King, A.
lation of their PDZ interactions, J. Virol., 89, 1579-1586,
(1999) Tenacity of exogenous human papillomavirus
doi: 10.1128/JVI.01961 14.
DNA in sperm washing, J. Assist. Reprod. Genet., 16,
61.
Songock, W.K., Kim, S.M., and Bodily, J.M. (2017) The
325-328.
human papillomavirus E7 oncoprotein as a regulator of
78.
Depuydt, C.E., Jonckheere, J., Berth, M., Salembier, G.M.,
transcription, Virus Res., 231, 56-75, doi: 10.1016/j.virus
Vereecken, A.J., and Bogers, J.J. (2015) Serial type specif
res.2016.10.017.
ic human papillomavirus (HPV) load measurement allows
62.
Katzenellenbogen, R. (2017) Telomerase induction in
differentiation between regressing cervical lesions and seri
HPV infection and oncogenesis, Viruses,
9, E180,
al virion productive transient infections, Cancer Med., 4,
doi: 10.3390/v9070180.
1294-1302, doi: 10.1002/cam4.473.
63.
Song, S., Pitot, H.C., and Lambert, P.F. (1999) The
79.
Gupta, S., Kumar, P., and Das, B.C. (2018) HPV:
human papillomavirus type 16 E6 gene alone is sufficient
Molecular pathways and targets, Curr. Probl. Cancer, 42,
to induce carcinomas in transgenic animals, J. Virol., 73,
161-174, doi: 10.1016/j.currproblcancer.2018.03.003.
5887-5893.
80.
Holmes, A., Lameiras, S., Jeannot, E., Marie, Y., Castera,
64.
Dyson, N.J. (2016) RB1: a prototype tumor suppressor and
L., Sastre Garau, X., and Nicolas, A. (2016) Mechanistic
an enigma, Genes Dev., 30, 1492-502, doi: 10.1101/
signatures of HPV insertions in cervical carcinomas, NPJ
gad.282145.116.
Genom. Med., 1, 16004, doi: 10.1038/npjgenmed.2016.4.
65.
Godinho, S.A., and Pellman, D. (2014) Causes and conse
81.
Akagi, K., Li, J., Broutian, T.R., Padilla Nash, H., Xiao, W.,
quences of centrosome abnormalities in cancer, Philos.
Jiang, B., Rocco, J.W., Teknos, T.N., Kumar, B., Wangsa, D.,
Trans. R. Soc. Lond. B Biol. Sci., 369, 20130467,
He, D., Ried, T., Symer, D.E., and Gillison, M.L. (2014)
doi: 10.1098/rstb.2013.0467.
Genome wide analysis of HPV integration in human can
66.
Sen, P., Ganguly, P., and Ganguly, N. (2017) Modulation
cers reveals recurrent, focal genomic instability, Genome
of DNA methylation by human papillomavirus E6 and E7
Res., 24, 185-199, doi: 10. 1101/gr.164806.113.
oncoproteins in cervical cancer, Oncol. Lett., 15, 11-22,
82.
Hu, Z., Zhu, D., Wang, W., Li, W., Jia, W., Zeng, X., Ding, W.,
doi: 10.3892/ol.2017.7292.
Yu, L., Wang, X., Wang, L., Shen, H., Zhang, C., Liu, H.,
67.
Mirabello, L., Yeager, M., Yu, K., Clifford, G.M., Xiao, Y.,
Liu, X., Zhao, Y., Fang, X., Li, S., Chen, W., Tang, T., Fu, A.,
Zhu, B., Cullen, M., Boland, J.F., Wentzensen, N.,
Wang, Z., Chen, G., Gao, Q., Li, S., Xi, L., Wang, C.,
Nelson, C.W., Raine Bennett, T., Chen, Z., Bass, S.,
Liao, S.., Ma, X., Wu, P., Li, K., Wang, S., Zhou, J., Wang, J.,
Song, L., Yang, Q., Steinberg, M., Burdett, L., Dean, M.,
Xu, X., Wang, H., and Ma, D. (2015) Genome wide pro
Roberson, D., Mitchell, J., Lorey, T., Franceschi, S.,
filing of HPV integration in cervical cancer identifies clus
Castle, P.E., Walker, J., Zuna, R., Kreimer, A.R., Beachler, D.C.,
tered genomic hot spots and a potential microhomology
Hildesheim, A., Gonzalez, P., Porras, C., Burk, R.D., and
mediated integration mechanism, Nat. Genet.,
47,
Schiffman, M. (2017) HPV16 E7 genetic conservation is
158-163, doi: 10.1038/ng.3178.
critical to carcinogenesis, Cell,
170,
1164-1174,
83.
Parfenov, M., Pedamallu, C.S., Gehlenborg, N., Freeman,
doi: 10.1016/j.cell.2017.08.001.
S.S., Danilova, L., Bristow, C.A., Lee, S., Hadjipanayis,
68.
Yu, J.H., Shi, W.W., Zhou, M.Y., Liu, J.M., Han, Q.Y.,
A.G., Ivanova, E.V., Wilkerson, M.D., Protopopov, A., Yang,
and Xu, H.H. (2019) Genetic variability and oncogenic
L., Seth, S., Song, X., Tang, J., Ren, X., Zhang, J., Pantazi,
risk association of human papillomavirus type 58 E6 and
A., Santoso, N., Xu, A.W., Mahadeshwar, H., Wheeler, D.A.,
E7 genes in Taizhou area, China, Gene, 686, 171-176,
Haddad, R.I., Jung, J., Ojesina, A,I., Issaeva, N., Yarbrough,
doi: 10.1016/j.gene.2018.11.066.
W.G., Hayes, D.N., Grandis, J.R., El Naggar, A.K.,
69.
Chabeda, A., Yanez, R.Jr., Lamprecht, R., Meyers, A.E.,
Meyerson, M., Park, P,J., Chin, L., Seidman, J.G.,
Rybicki, E.P., and Hitzeroth, I.I. (2017) Therapeutic vac
Hammerman, P.S., and Kucherlapati, R.
(2014)
cines
for
high risk
HPV associated diseases,
Characterization of HPV and host genome interactions in
Papillomavirus Res., 5, 46-58, doi: 10.1016/j.pvr.2017.
primary head and neck cancers, Proc. Natl. Acad. Sci. USA,
12.006.
111, 15544-15549, doi: 10.1073/pnas. 1416074111.
БИОХИМИЯ том 84 вып. 7 2019
1014
ВОНСКИЙ и др.
84.
Groves, I.J., and Coleman, N. (2018) Human papillo
or foes, Clinics (Sao Paulo), 73 (suppl. 1), e549s,
mavirus genome integration in squamous carcinogenesis:
doi: 10.6061/clinics/2018/e549s.
what have next generation sequencing studies taught us?
94. Stanley, M. (2010) HPV - immune response to infection
J. Pathol., 245, 9-18, doi: 10.1002/path.5058.
and vaccination, Infect. Agent. Cancer,
5,
19,
85.
Oyervides Munoz, M.A., Perez Maya, A.A., Rodriguez
doi: 10.1186/1750 9378 5 19.
Gutierrez, H.F., Gomez Macias, G.S., Fajardo Ramirez,
95. Ribeiro Muller, L., and Muller, M. (2014) Prophylactic
O.R., Trevino, V., Barrera Saldana, H.A., and Garza
papillomavirus vaccines, Clin. Dermatol., 32, 235-347,
Rodriguez, M.L. (2018) Understanding the HPV integra
doi: 10.1016/j.clindermatol.2013.08.008.
tion and its progression to cervical cancer, Infect. Genet.
96. Martinez Gomez, X., Curran, A., Campins, M., Alemany,
Evol., 61, 134-144, doi: 10.1016/j.meegid.2018.03.003.
L., Rodrigo Pendas, J., Borruel, N., Castellsague, X.,
86.
Magaldi, T.G., Almstead, L.L., Bellone, S., Prevatt, E.G.,
Diaz de Heredia, C., Moraga Llop, F., del Pino, M., and
Santin, A.D., and DiMaio, D. (2012) Primary human cer
Torne, A. (2019) Multidisciplinary, evidence based con
vical carcinoma cells require human papillomavirus E6 and
sensus guidelines for human papillomavirus (HPV) vacci
E7 expression for ongoing proliferation, Virology, 422,
nation in high risk populations, Spain, 2016, Euro
114-24, doi: 10.1016/j.virol.2011.10.012.
Surveill., 24, 1700857, doi: 10.2807/1560 7917.ES.2019.
87.
Bossler, F., Kuhn, B.J., Gunther, T., Kraemer, S.J.,
24.7.1700857.
Khalkar, P., Adrian, S., Lohrey, C., Holzer, A.,
97. Petrosky, E., Bocchini, J.A., Hariri S., Chesson, H.,
Shimobayashi, M., Durst, M., Mayer, A., Rosl, F.,
Curtis, R., Saraiya, M., Unger, E., Markowitz, L., and
Grundhoff, A., Krijgsveld, J., Hoppe Seyler, K., and
Centers for Disease Control and Prevention (CDC) (2015)
Hoppe Seyler, F. (2019) Repression of human papillo
Use of 9 valent human papillomavirus (HPV) vaccine:
mavirus oncogene expression under hypoxia is mediated by
updated HPV vaccination recommendations of the adviso
PI3K/mTORC2/AKT signaling, MBio, 10, e02323-18,
ry committee on immunization practices, MMWR Morb.
doi: 10.1128/mBio.02323 18.
Mortal. Wkly Rep., 64, 300-304.
88.
Johansson, C., Somberg, M., Li, X., Winquist, E., Fay, J.,
98. Markowitz, L.E., Dunne, E.F., Saraiya, M., Lawson,
Ryan, F., Pim, D., Banks, L., and Schwartz, S. (2012)
H.W., Chesson, H., Unger, E.R.; Centers for Disease
HPV 16 E2 contributes to induction of HPV 16 late gene
Control and Prevention (CDC); and Advisory Committee
expression by inhibiting early polyadenylation, EMBO J.,
on Immunization Practices (ACIP) (2007) Quadrivalent
31, 3212-3227, doi: 10.1038/emboj.2012.147.
human papillomavirus vaccine: recommendations of the
89.
Yang, A., Farmer, E., Wu, T.C., and Hung, C.F. (2016),
Advisory Committee on Immunization Practices (ACIP),
Perspectives for therapeutic HPV vaccine development,
MMWR Recomm. Rep., 56, 1-24.
J. Biomed. Sci., 23, 75, doi: 10.1186/s12929 016 0293 9.
99. Lehtinen, M., Lagheden, C., and Luostarinen, T. (2017)
90.
Vinokurova, S., Wentzensen, N., Kraus, I., Klaes, R.,
Ten year follow up of human papillomavirus vaccine effi
Driesch, C., Melsheimer, P., Kisseljov, F., Durst, M.,
cacy against the most stringent cervical neoplasia end
Schneider, A., and Doeberitz, M.V.K. (2008) Type depen
point registry based follow up of three cohorts from ran
dent integration frequency of human papillomavirus
domized trials, BMJ Open, 7, e015867, doi: 10.1136/
genomes in cervical lesions, Cancer Res., 68, 307-313,
bmjopen 2017 015867.
doi: 10.1158/0008 5472.CAN 07 2754.
100. Maver, P.J., and Poljak, M. (2018) Progress in prophylactic
91.
Roura, E., Travier, N., Waterboer, T., de Sanjose, S.,
human papillomavirus (HPV) vaccination in 2016: a liter
Bosch, F.X., Pawlita, M., Pala, V., Weiderpass, E., Margall, N.,
ature review, Vaccine, 36, 5416-5423, doi: 10.1016/j.vac
Dillner, J., Gram, I.T., Tjonneland, A., Munk, C., Palli, D.,
cine.2017.07.113.
Khaw, K.T., Overvad, K., Clavel Chapelon, F., Mesrine, S.,
101. Lehtinen, M., Baussano, I., Paavonen, J., Vanska, S., and
Fournier, A., Fortner, R.T., Ose, J., Steffen, A.,
Dillner, J. (2019) Eradication of human papillomavirus
Trichopoulou, A., Lagiou, P., Orfanos, P., Masala, G.,
and elimination of HPV related diseases - scientific basis
Tumino, R., Sacerdote, C., Polidoro, S., Mattiello, A.,
for global public health policies, Expert Rev. Vaccines, 18,
Lund, E., Peeters, P.H., Bueno de Mesquita, H.B.,
153-160, doi: 10.1080/14760584.2019.1568876.
Quiros, J.R., Sanchez, M.J., Navarro, C., Barricarte, A.,
102. Klosky, J.L., Gamble, H.L., Spunt, S.L., Randolph, M.E.,
Larraсaga, N., Ekstrom, J., Lindquist, D., Idahl, A.,
Green, D.M., and Hudson, M.M. (2009) Human papillo
Travis, R.C., Merritt, M.A., Gunter, M.J., Rinaldi, S.,
mavirus vaccination in survivors of childhood cancer,
Tommasino, M., Franceschi, S., Riboli, E., and
Cancer, 115, 5627-5636, doi: 10.1002/cncr.24669.
Castellsague, X. (2016) The influence of hormonal factors
103. Papasavvas, E., Surrey, L.F., Glencross, D.K., Azzoni, L.,
on the risk of developing cervical cancer and pre cancer:
Joseph, J., Omar, T., Feldman, M., Williamson, A.,
results from the EPIC cohort, PLoS One, 11, e0147029,
Siminya, M., Swarts, A., Yin, X., Liu, Q., Firnhaber, C.,
doi: 10.1371/journal.pone.0147029.
and Montanera, L.J. (2016) High risk oncogenic HPV
92.
den Boon, J.A., Pyeon, D., Wang, S.S., Horswill, M.,
genotype infection associates with increased immune acti
Schiffman, M., Sherman, M., Zuna, R.E., Wang, Z.,
vation and T cell exhaustion in ART suppressed HIV 1
Hewitt, S.M., Pearson, R., Schott, M., Chung, L., He, Q.,
infected women, Oncoimmunology,
5, e1128612,
Lambert, P., Walker, J., Newton, M.A., Wentzensen, N.,
doi: 10.1080/2162402X.2015.1128612.
and Ahlquist, P. (2015) Molecular transitions from papil
104. Костинов М.П., Зверев В.В. (2012) Экономическая
lomavirus infection to cervical precancer and cancer: Role
эффективность вакцинации против вируса папилло
of stromal estrogen receptor signaling, Proc. Natl. Acad.
мы человека в российской федерации, Журнал микро
Sci. USA,
112, E3255-E3264, doi:
10.1073/pnas.
биологии, эпидемиологии и иммунобиологии, 2, 43-50.
1509322112.
105. Zheng, Z.M., and Baker, C.C. (2006) Papillomavirus
93.
Nunes, R.A.L., Morale, M.G., Silva, G.Á.F., Villa, L.L.,
genome structure, expression, and post transcriptional
and Termini, L. (2018) Innate immunity and HPV: friends
regulation, Front. Biosci., 11, 2286-302.
БИОХИМИЯ том 84 вып. 7 2019
ВПЧ АССОЦИИРОВАНЫЙ КАНЦЕРОГЕНЕЗ
1015
CARCINOGENESIS ASSOCIATED WITH HUMAN PAPILLOMAVIRUS
INFECTION, MECHANISMS AND POTENTIAL FOR IMMUNOTHERAPY
M. S. Vonsky1,2, M. G. Shabaeva3, A. L. Runov1,2,4, N. N. Lebedeva4,5,
S. Chowdhury6, J. M. Palefsky6, and M. G. Isaguliants4,7,8,9*
1 Institute of Cytology, Russian Academy of Sciences, 194064 St. Petersburg, Russia; E mail: m.vonsky@gmail.com
2 Almazov National Medical Research Center, 19734, St. Petersburg, Russia; E mail: m.vonsky@gmail.com
3 Pavlov First St. Petersburg State Medical University, 197022 St. Petersburg, Russia; E mail: maria.dobro@mail.ru
4 Gamaleya Federal Research Center for Epidemiology and Microbiology,
123098 Moscow, Russia; E mail: maria.issagouliantis@rsu.lv
5 Moscow Regional Center of AIDS and Infectious Diseases Prevention and Treatment,
129110 Moscow, Russia; E mail: lebedevanatalya@rocketmail.com
6 University of California, San Francisco School of Medicine, San Francisco,
CA 94143, USA; E mail: Joel.Palefsky@ucsf.edu
7 Chumakov Federal Scientific Center for Research and Development of Immune and Biological Products,
Russian Academy of Sciences, 108819 Moscow, Russia
8 Department of Microbiology, Tumor and Cell Biology, Karolinska Institutet, SE 171 77, Stockholm, Sweden
9 Riga Stradins University, Department of Pathology, LV 1007, Riga, Latvia
Received April 2, 2019
Revised April 19, 2019
Accepted April 21, 2019
Human papillomavirus (HPV) infection is responsible for approximately 5% of all cancers and is associated with 30%
of all pathogen related cancers. Cervical cancer is the third most common cancer in women worldwide; about 70% of
cervical cancer cases are caused by the high risk HPVs (HR HPVs) of genotypes 16 and 18. HPV infection occurs
mainly through sexual contact; however, viral transmission via horizontal and vertical pathways is also possible. After
HPV infection of basal keratinocytes or ecto endocervical transition zone cells, viral DNA persists in the episomal
form. In most cases, infected cells are eliminated by the immune system. Occasionally, elimination fails, and HPV
infection becomes chronic. Replication of HPVs in dividing epithelial cells is accompanied by increased expression of
the E6 and E7 oncoproteins. These oncoproteins are responsible for genomic instability, disruption of the cell cycle,
cell proliferation, immortalization, and malignant transformation of HPV infected cells. Besides, E6 and E7 onco
proteins induce immunosuppression, preventing the detection of HPV infected and transformed cells by the immune
system. HPV integration into the genome of the host cell leads to the upregulation of E6 and E7 expression and con
tributes to HPV associated malignization. Prophylactic HPV vaccines can prevent over 80% of HPV associated
anogenital cancers. The vaccine elicits immune response that prevents initial infection with a given HPV type but does
not eliminate persistent virus once infection has occurred and does not prevent development of the HPV associated
neoplasias, which necessitates the development of therapeutic vaccines to treat chronic HPV infections and HPV
associated malignancies.
Keywords: human papillomavirus (HPV), squamous cell carcinoma, carcinogenesis, cervical cancer, intraepithelial
neoplasias, epidemiology, E6/E7 oncoproteins
БИОХИМИЯ том 84 вып. 7 2019