БИОХИМИЯ, 2021, том 86, вып. 2, с. 259 - 273
УДК 576.52, 576.54
УРОВНИ БЕЛКОВ CRABP1 И CRABP2 НЕ КОРРЕЛИРУЮТ
С ЧУВСТВИТЕЛЬНОСТЬЮ КЛЕТОК РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ
К РЕТИНОЕВОЙ КИСЛОТЕ, НО КОРРЕЛИРУЮТ МЕЖДУ СОБОЙ,
ПРИЧЕМ CRABP2 ЯВЛЯЕТСЯ РЕГУЛЯТОРОМ ПРОДУКЦИИ CRABP1*
© 2021
А.Д. Еникеев, А.В. Комельков**, М.Е. Аксельрод, С.А. Галецкий,
С.А. Кузьмичев, Е.М. Чевкина
ФГБУ «НМИЦ онкологии им. Н.Н. Блохина» Минздрава России,
115478 Москва, Россия; электронная почта: komelkov@gmail.com
Поступила в редакцию 17.08.2020
После доработки 30.09.2020
Принята к публикации 30.09.2020
Белки, связывающие ретиноевую кислоту (РК), CRABP1 и CRABP2 являются молекулярными шаперона
ми, обеспечивающими реализацию внутриклеточной активности РК, важнейшего стимулятора клеточной
дифференцировки, обладающего опухоль супрессорной активностью. Одной из основных функций
CRABP2 является доставка и передача ретиноевой кислоты ядерным рецепторам RAR/RXR, что приводит
к активации транскрипции широкого спектра ретиноид респонсивных генов. Функции CRABP1 менее изу
чены, но связаны, по видимому, с удержанием ретиноевой кислоты в цитоплазме и ограничением ее актив
ности в отношении регуляции транскрипции генов, что предполагает участие данного белка в формирова
нии РК резистентности. Также малопонятны и механизмы, регулирующие активность CRABP1. Впервые
проведенное в данной работе сравнение уровней CRABP1 в линиях опухолевых клеток различного проис
хождения показало отсутствие этого белка в клетках опухолей, считающихся РК резистентными, и его вы
раженную продукцию в РК чувствительных клетках. Вместе с тем анализ, проведенный на панели клеточ
ных линий рака молочной железы с различным уровнем РК чувствительности, показал отсутствие корре
ляции продукции белка CRABP1 с чувствительностью клеток к ретиноевой кислоте. При этом впервые об
наружена корреляция продукции белков CRABP1 и CRABP2 во всех исследованных типах клеток, вне за
висимости от их происхождения и РК чувствительности/резистентности. Более того, впервые показано,
что нокдаун гена CRABP2 приводит к подавлению продукции белка CRABP1 как в РК чувствительных, так
и в РК резистентных клетках. Обнаруженная нами CRABP2 зависимая регуляция уровня CRABP1 являет
ся новым механизмом внутриклеточной ретиноевой сигнальной системы.
КЛЮЧЕВЫЕ СЛОВА: ретиноевая кислота, ATRA, CRABP1, CRABP2, пролиферация, регуляция
экспрессии.
DOI: 10.31857/S032097252102010X
ВВЕДЕНИЕ
тов ретинола (витамина А). Функциональная
активность РК связана с регуляцией транскрип
Ретиноевая кислота (РК) является одним из
ции более пяти сотен генов, в промоторной об
наиболее активных внутриклеточных метаболи ласти которых имеются ретиноид респонсив
ные элементы (RARE). Она реализуется с по
Принятые сокращения: НМРЛ - немелкоклеточ
ный рак легкого; РК - ретиноевая кислота; РМЖ - рак
мощью ядерных рецепторов РК, прежде всего
молочной железы; ATRA - полностью транс ретиноевая
белков RAR (RARα, β и γ) и RXR, которые
кислота (all trans retinoic acid); CRABP1 и CRABP2 - бел
функционируют в качестве транскрипционных
ки, связывающие ретиноевую кислоту, 1 и 2 (cellular факторов в составе гетеродимеров, а также ре
retinoic acid binding proteins 1 and 2); FABP5 - белок, свя
цептора PPARβ/δ, также образующего гетероди
зывающий жирные кислоты (fatty acid binding protein);
GFP - зеленый флуоресцентный белок (green fluorescent
мер с RXR [1]. За счет способности контролиро
protein); PPAR - рецепторы, активируемые пролиферато
вать транскрипционную активность ряда генов
рами пероксисом (peroxisome proliferator activated recep
РК регулирует важнейшие биологические про
tors); RAR - рецептор ретиноевой кислоты (retinoic acid цессы, связанные с дифференцировкой клеток,
receptor).
ремоделированием тканей и иммунным отве
* Первоначально английский вариант рукописи опубли
том. В большинстве клеток РК стимулирует
msu.ru/biokhimiya в рубрике «Papers in Press», BM20 267,
дифференцировку или апоптоз и даже является
25.01.2021.
негативным регулятором пролиферации, в свя
** Адресат для корреспонденции.
зи с чем ее принято считать опухолевым супрес
259
8*
260
ЕНИКЕЕВ и др.
сором. В этой связи предпринимаются актив
(РМЖ), которые показали, что FABP5 преиму
ные попытки разработки препаратов для тера
щественно экспрессируется в ER негативных
пии опухолей на основе РК и ее синтетических
(абб. ER от англ. estrogen negative) и трижды не
аналогов, однако до сих пор применение рети
гативных опухолях, которые обычно демон
ноидов в клинической практике ограничено (за
стрируют РК резистентность [8]. Наибольшей
исключением острого промиелоцитарного лей
аффинностью к РК среди РК связывающих
коза) прежде всего быстрым приобретением
белков обладает белок CRABP1 [13], однако о
злокачественными клетками устойчивости к те
его функциональном значении известно очень
рапии на основе производных или синтетичес
мало, а данные о его роли в трансформации кле
ких ретиноидов [2].
ток и опухолевой прогрессии еще более проти
Реализация функциональной активности РК
воречивы.
во многом определяется ее доставкой к ядерным
Предположительно, связывание CRABP1 с
рецепторам. За внутриклеточный транспорт
РК приводит к удержанию ее в цитоплазме [14],
гидрофобных молекул РК к рецепторам отвеча
ограничению активности (включая доступ РК к
ют представители семейства липид связываю
белку CRABP2 по конкурентному механизму
щих белков, прежде всего два белка, связываю
[15]), и даже может способствовать метаболизму
щих РК, CRABP1 и CRABP2 (cellular retinoic
РК [16-18]. В соответствии с этим можно сде
acid binding proteins 1 and 2), а также представи
лать предположение о роли CRABP1 в приобре
тель семейства белков, связывающих жирные
тении клетками устойчивости к РК. Однако за
кислоты, FABP5 (fatty acid binding protein 5) [3].
исключением единственной работы, где показа
Согласно некоторым данным, белки CRABP в
но увеличение экспрессии CRABP1 в
целом (или как минимум CRABP2) обеспечива
ER/PR негативных (абб. PR от англ. proges
ют взаимодействие РК с рецепторами
terone negative) опухолях молочной железы [15],
RAR/RXR, в то время как FABP5 доставляет РК
исследований о связи данного белка с чувстви
к рецептору PPARβ/δ. Предполагается, что взаи
тельностью к РК нам не известно. По другим
модействие РК с разными ядерными рецептора
данным, CRABP1 обладает схожими с CRABP2
ми приводит к противоположному эффекту на
функциями - он также направляет РК к рецеп
клетки:
CRABP2 зависимая
активация
торам RAR, однако не взаимодействует с ними
RAR/RXR способствует стимуляции дифферен
напрямую, передаче РК рецептору предшеству
цировки и реализации проапоптотической и ан
ет диссоциация комплекса РК-CRABP1 [13]. В
типролиферативной активности РК, в то время
этой связи остается непонятной функция этого
как опосредуемое FABP5 взаимодействие РК с
белка в ядре, хотя он, как и CRABP2, имеет сиг
PPARβ/δ способствует активации транскрип
нал ядерной локализации, а присутствие
ции генов, ответственных за выживание, проли
CRABP1 в ядре подтверждается как нашими
ферацию и ангиогенез [4]. Аффинность РК к
предшествующими исследованиями [19], так и
белкам CRABP выше, чем к FABP5 [5], кроме
данными литературы [20].
того, в большинстве клеток соотношение уров
О возможной взаиморегуляции белков
ня экспрессии белков CRABP2/FABP5 и
CRABP в настоящее время ничего не известно,
RAR/PPARβ/δ сдвинуто в сторону CRABP-RAR
хотя ее наличие кажется достаточно вероятным,
пути, что определяет опухоль супрессорную
учитывая то, что оба белка обеспечивают реали
функцию РК в большинстве типов тканей [6].
зацию сходных РК регулируемых процессов и
Существуют данные, указывающие на то, что
сами, в свою очередь, являются продуктами
увеличение соотношения FABP5/CRABP2 кор
экспрессии ретиноид зависимых генов. На та
релирует с устойчивостью клеток к РК [7, 8]. В
кую возможность указывают и полученные на
связи с описанной функцией CRABP2 его при
ми ранее данные [21].
нято считать опухолевым супрессором. Более
Тем не менее среди достаточно многочис
того, для него показана активность в цитоплаз
ленных исследований РК связывающих белков,
ме в отсутствие РК, не связанная с активацией
за исключением цитируемой выше работы по
ее рецепторов, которая также способствует оста
изучению функций CRABP1/2 при раке молоч
новке клеточного цикла и стабилизации тран
ной железы [15] и нашей работы, выполненной
скриптов проапоптотических генов [9, 10]. При
на образцах немелкоклеточного рака легкого
этом в некоторых типах опухолей CRABP2 игра
[21], практически отсутствуют исследования, в
ет, по видимому, опухоль промоторную роль
которых анализировались бы одновременно
[11, 12], однако механизмы такой активности
белки CRABP1 и CRABP2.
остаются малопонятными. Косвенно на участие
В данной работе мы впервые исследовали
FABP5 в РК резистентности указывают и ре
экспрессию обоих гомологов CRABP на широ
зультаты исследования рака молочной железы
кой панели клеточных линий и проанализирова
БИОХИМИЯ том 86 вып. 2 2021
КОРРЕЛЯЦИЯ CRABP2, CRABP1 И РК ЧУВСТВИТЕЛЬНОСТИ
261
ли их возможную связь с чувствительностью/ре
живали в количестве 50-300 тысяч на 60 мм
зистентностью клеток к РК. Мы не выявили
чашки Петри («SPL Life Sciences», США) в зави
строгой ассоциации между продукцией исследу
симости от клеточной линии и культивировали
емых белков и РК чувствительностью, хотя
в течение 5 сут в присутствии ATRA в диапазоне
большинство РК чувствительных линий харак
концентраций 0,1-100 мкM. Снимали клетки
теризовалось высоким уровнем CRABP1/2, а на
0,25% ным раствором трипсин ЭДТА, смеши
иболее РК резистентные линии демонстрирова
вали с красителем трипановый синий в соотно
ли отсутствие указанных белков. При этом была
шении 1 : 1 и подсчитывали в камере Горяева. В
обнаружена корреляция между продукцией
качестве контроля для каждой линии использо
CRABP1 и CRABP2 - все линии, в которых
вали клетки, культивируемые параллельно в те
экспрессируется CRABP1, характеризуются вы
чение того же времени в среде без ATRA, содер
соким уровнем продукции CRABP2, в то время
жащей ДМСО в соответствующей концентра
как во всех линиях клеток с низким уровнем или
ции. Количество живых клеток в контроле для
отсутствием CRABP2 продукция CRABP1 не ре
каждой линии брали за 100%.
гистрируется. Более того, впервые было показа
Получение производных линий клеток с гипер;
но, что экспрессия белков CRABP имеет функ
экспрессией экзогенного CRABP1. Молекуляр
циональную связь, в которой CRABP2 является
ное клонирование и ретровирусную инфекцию
регулятором продукции CRABP1.
проводили по описанной ранее методике [22].
Кодирующая последовательность CRABP1 была
амплифицирована с помощью ПЦР с использо
МАТЕРИАЛЫ И МЕТОДЫ
ванием праймеров к последовательностям,
фланкирующим кодирующую область мРНК
Клеточные линии. Были использованы следу
CRABP1:
ющие линии клеток из коллекции ATCC: немел
коклеточного рака легкого (НМРЛ) - A549,
CRABP1 cloning F: 5′ ATTCTCGAGCCACCATGC
H1299, H460; глиобластомы - LN229 и U87; ра
CCAACTTC 3′ и
ка яичника - OVCAR 8, SK OV 3, EFO 21; нейро
CRABP1 cloning R: 5′ ACAGGATCCCTGCCTT
бластомы - SK N AS, SH SY 5Y, IMR 32; рака
CACTCTCGG 3′
молочной железы (РМЖ) - MCF7, T47D,
SKBR3, HCC1954, MDA MB 453, HCC1937,
(сайты рестрикции выделены курсивом, компле
MDA MB 468, MDA MB 231, HBL100. Для по
ментарные мРНК последовательности подчерк
лучения псевдоретровирусных частиц использо
нуты). Матрицей для ПЦР служила кДНК, син
вались линии эпителиальных клеток 293FT
тезированная с мРНК, выделенной из ткани ра
(«Invitrogen», США) и GP293
(«Clontech»,
ка легкого человека. Амплификацию проводили
США). Клетки культивировали в среде DMEM
в присутствии высокоточной полимеразы PFX
(«ПанЭко», Россия), содержащей 0,294 мг/мл
(«Invitrogen», США) согласно протоколу произ
L глутамина, 10% эмбриональной телячьей сы
водителя. Кодирующая последовательность бы
воротки («PAA Laboratories», Канада), 0,1 мг/мл
ла клонирована в ретровирусный вектор pLXSN
стрептомицина, 100 ед/мл пенициллина при
(«Clontech», США) по сайтам рестрикции XhoI
37 °С и 5% CO2. Клетки MCF10A эпителия мо
и BamHI. Проверку осуществляли с помощью
лочной железы культивировали в среде
рестрикции полученных конструктов, а также
DMEM/F 12 («Gibco», США), содержащей 5%
методом ПЦР с помощью следующих прайме
лошадиной сыворотки
(«Gibco», США),
ров:
20 нг/мл ростового фактора EGF («Thermo
Fisher Scientific», США), 0,5 мг/мл гидрокорти
pLXSN F: 5′ CCCTTGAACCTCCTCGTTCG 3′ и
зона («Sigma Aldrich», США), 100 нг/мл холер
pLXSN R: 5′ TTTCCACACCTGGTTGCTGA 3′,
ного токсина
(«Sigma Aldrich», США) и
10 мкг/мл инсулина («Sigma Aldrich», США).
подобранных к участкам вектора, фланкирую
Отсутствие контаминации микоплазмой прове
щим вставку. Соответствие последовательности
ряли стандартными методами, ПЦР и окраши
CRABP1 человека подтверждали с помощью сек
ванием Hoechst
33342
(«Thermo Fisher
венирования. Псевдоретровирусные частицы
Scientific», США).
получали с помощью трансфекции клеток
Анализ чувствительности клеток к ретиноевой
GP293. Для трансфекции брали 2 мкг ДНК сме
кислоте. Полностью транс ретиноевую кислоту
си pLXSN CRABP1 и pVSV G («Clontech»,
(абб. ATRA от англ. all trans retinoic acid) раство
США) в эквимолярном соотношении. Транс
ряли в ДМСО для приготовления стокового
фекцию проводили с использованием липофек
раствора с концентрацией 10 мМ. Клетки выса
тамина 2000 («Invitrogen», США) согласно про
БИОХИМИЯ том 86 вып. 2 2021
262
ЕНИКЕЕВ и др.
токолу производителя. По достижении 20-30%
лизировали в буфере RIPA (50 мМ Tris HCl
конфлюэнтности клеток добавляли 24, 48 и
(pH 7,5), 150 мМ NaCl, 0,5% DoX, 1% NP 40,
72 часовые инокуляты вируса, смешанные со
0,1% SDS, 2 мМ ЭДТА) с добавлением смеси ин
средой в соотношении 1 : 1, с добавлением
гибиторов протеаз Complete Protease Inhibitor
8 мкг/мл полибрена («Sigma Aldrich», США).
Cocktail («Roche», Швейцария). Концентрацию
Селекцию проводили на G418 («Sigma Aldrich»,
белка определяли методом Бредфорда с по
США) в концентрации 1000 мкг/мл в течение
мощью набора Bradford Assay Kit («Bio Rad»,
8-9 дней. Для контрольных экспериментов по
США). Белок в количестве 15 мкг разделяли в
лучали производные тех же линий, в которые
15% ном полиакриламидном геле, переносили
трансдуцировали не содержащий вставки век
на PVDF мембрану («Millipore», США). Мемб
тор pLXSN.
рану инкубировали в блокирующем растворе
Получение производных линий клеток с нок;
5% ного BSA
(«PAA Laboratories GmbH»,
дауном CRABP2. Предшественники малых шпи
Австрия) в
буфере ТBS, содержащем
лечных РНК (shRNA) к кодирующей последова
0,1% TWEEN 20 («Sigma Aldrich», США). Затем
тельности CRABP2 были клонированы в ленти
мембрану инкубировали первичными антитела
вирусный вектор pLKO.1 puro
(«Addgene»,
ми к CRABP1
(«Sigma Aldrich», США,
США) по сайтам рестрикции AgeI и EcoRI. По
HPA17203; 1 : 1000) или CRABP2 («Sigma
следовательности:
Aldrich», США, HPA004135; 1 : 500) 12 ч при
+4 °С. В качестве контроля нанесения белка ис
sh1CRABP2: 5′ CCGGGAAATGGGAGAGTGA
пользовали антитела к β актину («Abcam», Ве
GAATAACTCGAGTTATTCTCACTCTCCCA
ликобритания, ab8227; 1 : 5000). После стандарт
TTTCTTTTTG 3′ (TRCN0000021371),
ной отмывки мембрану инкубировали со вто
ричными антителами, конъюгированными c пе
sh2CRABP2: 5′ CCGGCGAGGAATTGCTCAAA
роксидазой хрена («Cell Signaling Technology»,
GTGCTCTCGAGAGCACTTTGAGCAATTCCT
США, 29902; 1 : 65 000) 1 ч при комнатной тем
CGTTTTTG 3′ (TRCN0000021370)
пературе. После отмывки мембрану проявляли с
помощью реагента ECL (Enhanced Chemilumi
(смысловая и антисмысловая последователь
nescence, «Millipore», США). Хемилюминесцент
ности подчеркнуты). Результаты проверяли с
ную реакцию регистрировали на приборе
помощью ПЦР с использованием праймеров
Kodak Gel Logic 2200 Imaging System с последу
pLKO.1 seq standart и с помощью секвенирова
ющей обработкой с помощью программы
ния (ID последовательности AH002814.2, Homo
Carestream Molecular Imaging Software SE ver.
sapiens retinoic acid binding protein II (CRABP II)
5.0.1.27.
gene, complete cds, доступна по адресу https://
Статистический анализ. Все данные были по
лучены в результате трех независимых экспери
Для получения псевдоретровирусных частиц
ментов. Данные представлены в виде среднего ±
использовали клетки 293FT при 70% конфлю
стандартного отклонения (SD). Для расчетов и
энтности. Для трансфекции брали 2 мкг ДНК
построения графиков была использована прог
смеси векторов pLKO.1 puro, pVSVG
рамма GraphPad Prism 8.3 (GraphPad Software,
(«Clontech», США) и pCMV delta R8.2
США).
(«Addgene», США) в эквимолярном соотноше
нии. Трансфекцию проводили с использовани
ем липофектамина 2000 («Invitrogen», США),
РЕЗУЛЬТАТЫ
согласно протоколу производителя. По дости
жении 20-30% конфлюэнтности клеток добав
Продукция CRABP1 и CRABP2 коррелирует в
ляли 24, 48 и 72 часовые инокуляты вируса, сме
клеточных линиях различного происхождения.
шанные со средой в соотношении 1 : 1, с добав
Ранее в процессе изучения РК связывающих
лением 8 мкг/мл полибрена («Sigma Aldrich»,
белков мы обнаружили высокую вариабель
США). Селекцию проводили на пуромицине
ность уровней CRABP (в особенности CRABP1)
(«Sigma Aldrich», США) в концентрации
в линиях опухолевых клеток различного проис
1-2 мкг/мл в зависимости от клеточной линии в
хождения. Учитывая данные литературы о воз
течение 4-5 дней. Для контрольных экспери
можной роли СRABP1 в качестве внутриклеточ
ментов получали производные тех же линий,
ного регулятора биодоступности РК, мы пред
экспрессирующие shRNA к зеленому флуорес
положили, что этот белок может быть вовлечен
центному белку (shGFP).
в формирование РК резистентности. В этой
Иммуноблоттинг. Иммуноблоттинг проводи
связи мы сравнили продукцию белков CRABP в
ли по описанной ранее методике [22]. Клетки
различных типах опухолевых клеток. По резуль
БИОХИМИЯ том 86 вып. 2 2021
КОРРЕЛЯЦИЯ CRABP2, CRABP1 И РК ЧУВСТВИТЕЛЬНОСТИ
263
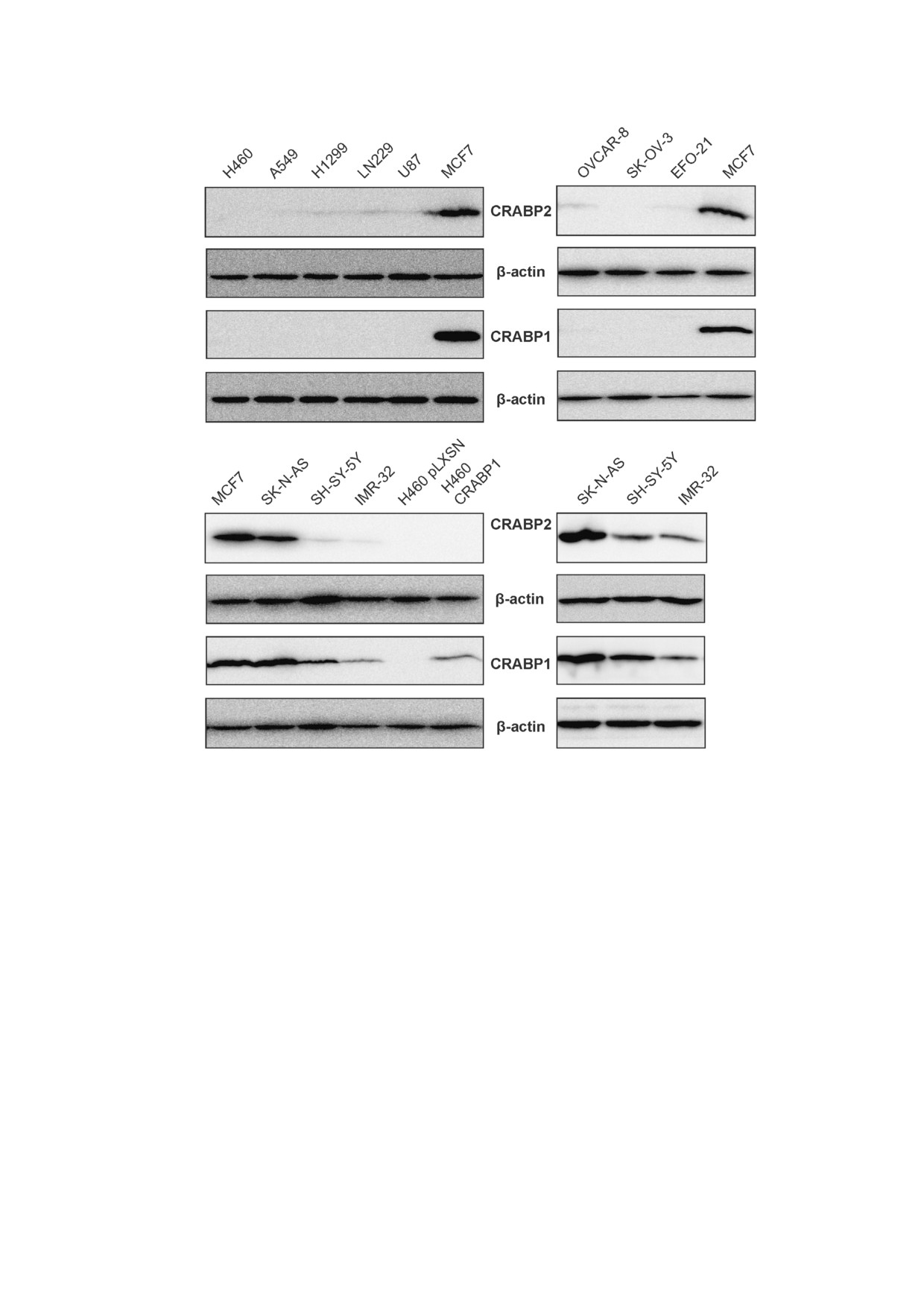
Рис. 1. Сравнение продукции белков CRABP1 и CRABP2 методом иммуноблоттинга в линиях опухолевых клеток различ
ного происхождения: НМРЛ (H1299, A549, H460), рака яичника (OVCAR 8, SC OV 3, EFO 21), глиобластомы (LN229,
U87), нейробластомы (IMR 32, SK N AS, SH SY 5Y). В качестве положительного контроля экспрессии CRABP1 исполь
зована линия РМЖ MCF7. В качестве контроля специфичности антител в отношении белков CRABP1 и CRABP2 исполь
зованы производные сублинии Н460 с гиперэкспрессией CRABP1 (Н460 CRABP1) и экспрессией вектора без вставки
(H460 pLXSN). Результаты по линиям нейробластомы приведены в нижней части рисунка дважды - совместно с клетка
ми MCF7 (слева) и в отсутствие MCF7 (справа) во избежание нивелирования («маскировки») различий в экспрессии бел
ков CRABP1 и CRABP2 между линиями при той же экспозиции мембраны в присутствии сильного сигнала от MCF7
татам анализа во всех линиях клеток НМРЛ
клетках с экспрессией CRABP1 вне зависимости
(А549, Н1299, Н460), рака яичников (SK OV 3,
от их происхождения наблюдался высокий уро
OVCAR 8, EFO 21) и глиобластомы (LN229,
вень CRABP2, в то время как в клетках с отсут
U87) мы наблюдали отсутствие белка CRABP1,
ствием экспрессии CRABP1 уровень CRABP2
в то время как во всех исследованных линиях
был заметно снижен или белок не регистриро
нейробластомы (SK N AS, SH SY 5Y, IMR 32)
вался. Для подтверждения специфичности ра
этот белок регистрировался на достаточно высо
боты используемых антител в отношении этих
ком уровне (рис. 1). В связи с тем, что в ряде
двух белков на рис. 1 представлены результаты
клеточных линий белок CRABP1 не регистриро
анализа CRABP1 и CRABP2 в клетках исходной
вался, мы использовали линию MCF7 РМЖ в
линии Н460 в отсутствие эндогенной экспрес
качестве положительного контроля наличия
сии CRABP1 и в ее производной сублинии с
сигнала. Мы также обнаружили, что во всех
экспрессией экзогенного CRABP1 (H460
БИОХИМИЯ том 86 вып. 2 2021
264
ЕНИКЕЕВ и др.
CRABP1), а также в контрольной линии, экс
течение 5 сут в стандартной среде (контроль) и в
прессирующей вектор без вставки (Н460
среде с ATRA в различных концентрациях, пос
pLXSN). Как видно на рисунке, во всех произ
ле чего проводили подсчет живых клеток. Диа
водных сублиниях экспрессия белка CRABP2 не
пазон концентраций ATRA варьировал от 0,1 до
регистрировалась вне зависимости от экспрес
100 мкМ. По результатам титрования ATRA и
сии CRABP1, что подтверждает специфичность
анализа количества живых клеток были выбра
антител.
ны пороговые концентрации ATRA и критерии,
По данным литературы, НМРЛ, рак яичника
позволяющие разделить линии клеток на
и глиобластомы считаются в целом РК резистент
РК чувствительные и РК резистентные. РК чув
ными, а нейробластомы - РК чувствительными
ствительными считали линии, показавшие сни
типами опухолей. Таким образом, наше предпо
жение пролиферации в два и более раза при ин
ложение о связи РК чувствительности клеток с
кубации с ATRA в концентрации до 10 мкМ. К
экспрессией CRABP1 подтверждалось. На осно
этой группе относились линии MCF7, T47D,
вании полученных данных мы также предполо
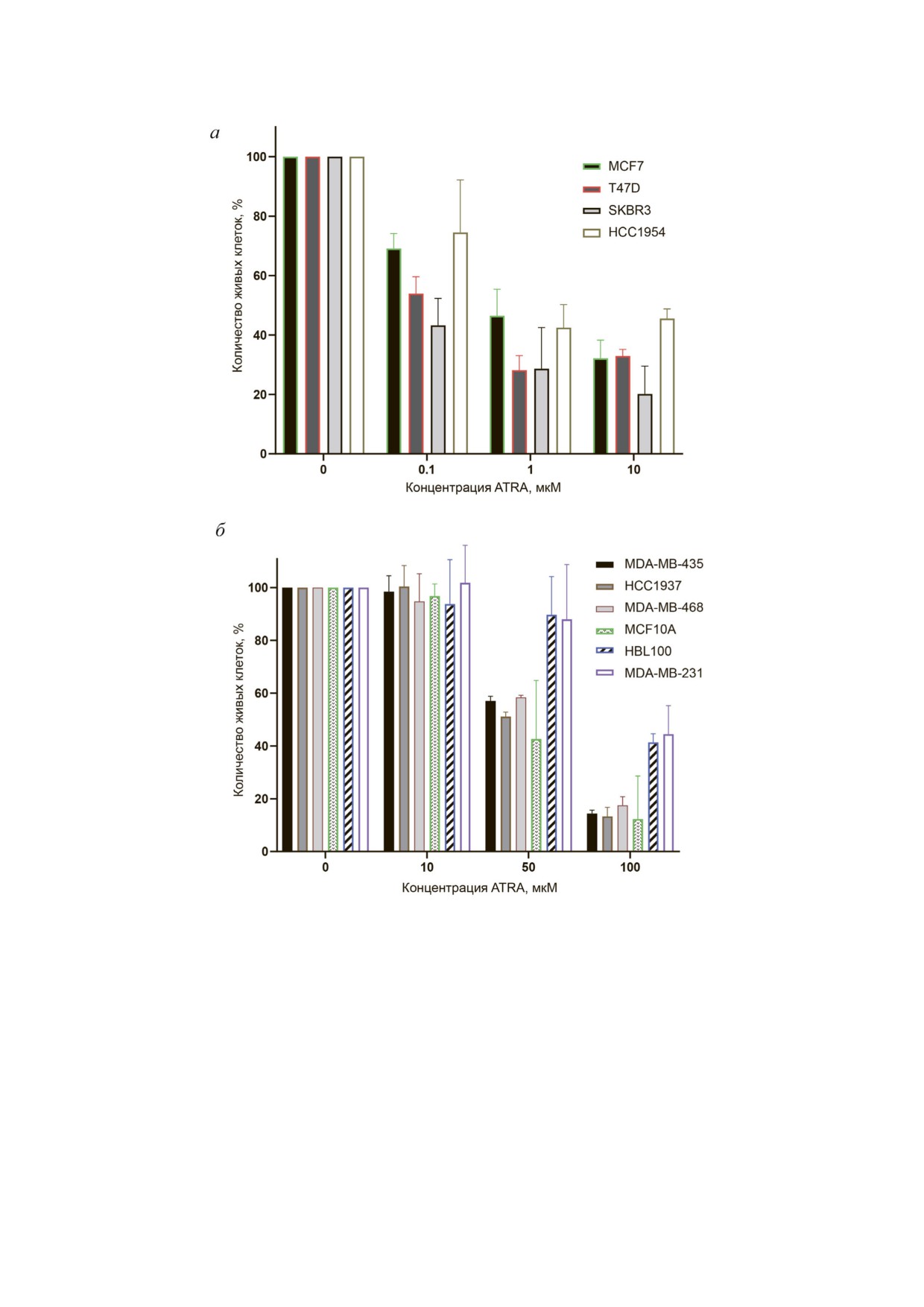
SKBR3 и HCC1954 (рис. 2, а). При этом
жили наличие регуляторной связи между белка
чувствительность отдельных линий внутри
ми CRABP1 и CRABP2. Однако обе гипотезы
группы варьировала - так, наиболее чувстви
необходимо было проверить, желательно с ис
тельными оказались линии SKBR3 и T47D, ко
пользованием клеточных линий сходного про
торые продемонстрировали двукратное сниже
исхождения, но различающихся по чувствитель
ние пролиферации по сравнению с контролем
ности к РК.
даже при концентрации ATRA 0,1 мкМ. В то же
Сравнение чувствительности к ретиноевой
время динамика пролиферации клеток в группе
кислоте различных линий клеток рака молочной
РК резистентных линий (MDA MB 453,
железы. Для проверки указанных гипотез была
MDA MB 468, MDA MB 231, HCC1937,
выбрана экспериментальная модель РМЖ.
HBL100, а также MCF10A) при добавлении
Во первых, выбор обусловливался большим пу
10 мкМ ATRA не менялась (рис. 2, б). Более то
лом преклинических данных, свидетельствую
го, количество живых клеток в этой группе ли
щих о перспективах использования РК в качест
ний не снижалось и при инкубации с ATRA в
ве агента для профилактики и лечения данного
концентрации 20 мкМ. Тем не менее в группе
типа опухолей [23-25]. Во вторых, РМЖ явля
РК резистентных клеток можно было выделить
ется гетерогенным типом опухолей, как по про
линии, динамика пролиферации которых не ме
исхождению, так и по молекулярным характе
нялась по сравнению с контролем даже при ин
ристикам, к которым относятся прежде всего
кубации с
50
мкМ ATRA (HBL100, MDA
экспрессия рецепторов эстрогена и прогестеро
MB 231), и линии, для которых было характер
на, а также статус HER2/neu. По данным лите
но снижение пролиферации при данных усло
ратуры, чувствительность этих опухолей к РК
виях (MDA MB 453, MDA MB 468, HCC1937,
также сильно варьирует [26, 27]. Различаются и
MCF10A).
данные в отношении уровня чувствительности к
Это означает, что чувствительность к РК
РК разных линий клеток РМЖ. Важно отме
варьирует в достаточно широком диапазоне, од
тить, что в разных работах для оценки
нако порогом можно считать интервал концент
РК чувствительности применяются различные
раций 1-10 мкМ, при котором пролиферация
подходы и методы оценки, а также значения
РК чувствительных клеток снижается в два и
концентрации РК. Поэтому на первом этапе не
более раза по сравнению с контролем, в то вре
обходимо было сравнить чувствительность к РК
мя как пролиферация РК резистентных клеток
широкой панели клеточных линий РМЖ в еди
либо не меняется, либо начинает снижаться
ной серии экспериментов, выбрать условия и
лишь при 50 мкМ ATRA - то есть, при концент
критерий (пороговое значение РК) для разделе
рациях РК, значительно превышающих физио
ния линий на РК чувствительные и РК резис
логические значения.
тентные. В исследование включили 9 линий
Продукция белков CRABP1 и CRABP2 в клет;
РМЖ, а также линию MCF10A, полученную из
ках РМЖ связана между собой, причем CRABP2
эпителия молочной железы, которая стандартно
является регулятором CRABP1. Далее мы прове
используется в качестве условно нормальной
ли анализ продукции РК связывающих белков
(нетуморогенной) линии клеток. Чувствитель
CRABP1 и CRABP2 в РК чувствительных и
ность к полностью транс ретиноевой кислоте
РК резистентных линиях РМЖ. По результа
(наиболее распространенная в клетках и биоло
там анализа мы не обнаружили достоверной ас
гически активная изоформа РК) анализировали
социации между экспрессией CRABP1 и
методом оценки динамики пролиферации. Для
РК чувствительностью. Так, в РК чувствитель
этого клетки каждой линии культивировали в
ной линии SKBR3 продукция CRABP1 отсут
БИОХИМИЯ том 86 вып. 2 2021
КОРРЕЛЯЦИЯ CRABP2, CRABP1 И РК ЧУВСТВИТЕЛЬНОСТИ
265
Рис. 2. Сравнение динамики пролиферации клеток РМЖ при инкубации с ATRA. Указан процент живых клеток после
5 сут культивирования в присутствии ATRA в различных концентрациях по сравнению с контролем. Контроль: за 100%
взято количество живых клеток каждой линии после культивирования в течение того же времени в стандартной среде
DMEM с добавлением ДМСО в концентрациях, соответствующих таковым при разведении ATRA. а - РК чувствитель
ные линии демонстрировали снижение пролиферации (падение ≥ 50% по сравнению с контролем) при концентрации
ATRA 0,1-10 мкМ; б - РК резистентные линии демонстрировали снижение пролиферации при концентрации ATRA
от 50 мкМ и выше. Диаграммы построены на основании средних значений для трех независимых повторов экспери
мента
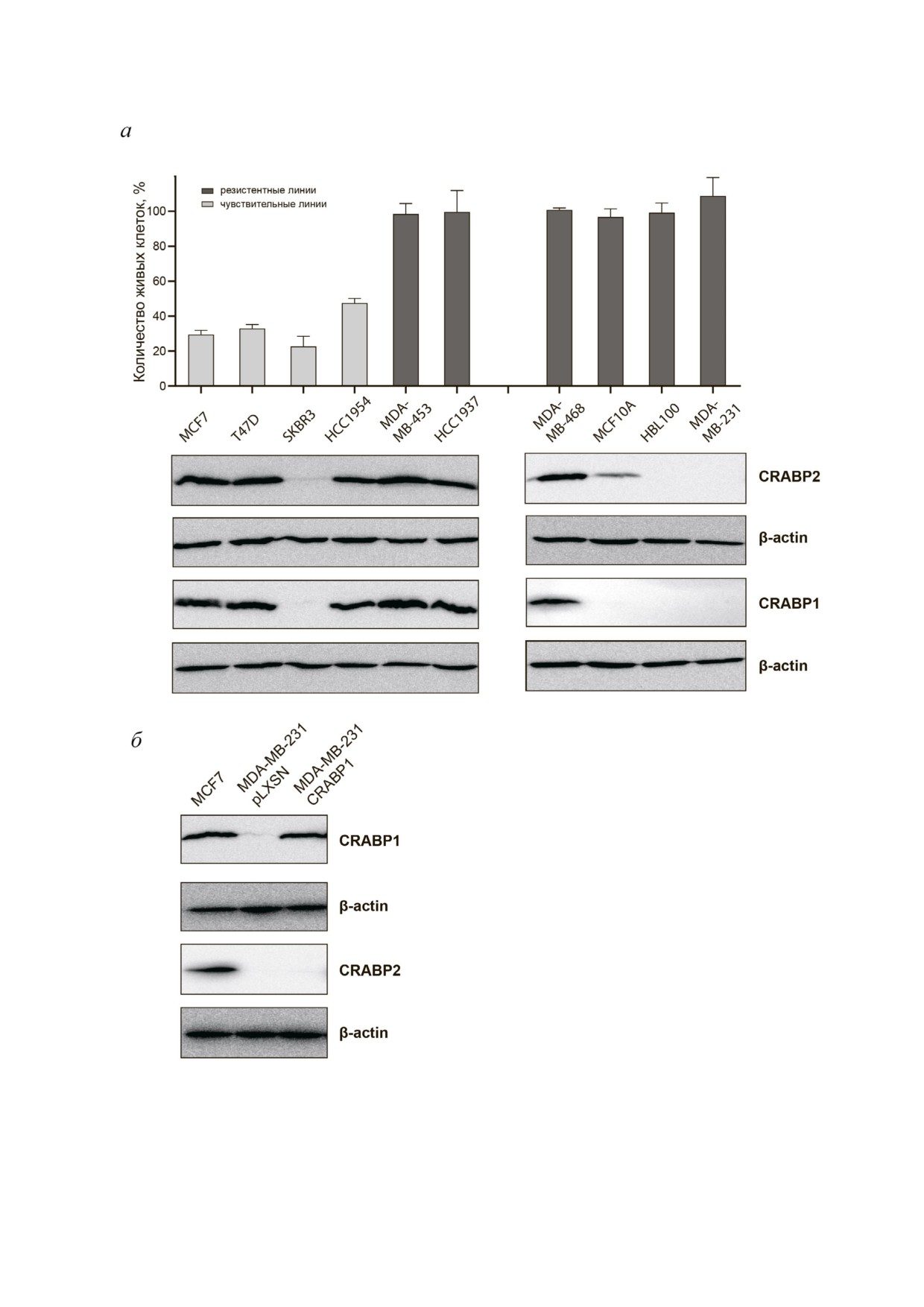
ствовала, а в РК резистентных линиях
Тем не менее стоит отметить, что в наиболее
MDA MB 453, MDA MB 468 и HCC1937 реги
устойчивых линиях MDA MB 231 и HBL100,
стрировался высокий уровень данного белка
пролиферация которых значимо не снижалась
(рис. 3, а).
даже при концентрации ATRA 50 мкМ, продук
БИОХИМИЯ том 86 вып. 2 2021
266
ЕНИКЕЕВ и др.
Рис. 3. Сравнение продукции белков CRABP1 и CRABP2 в линиях РМЖ с различной РК чувствительностью. а - Про
цент живых клеток каждой линии по сравнению с соответствующим контролем (взят за 100%) при инкубации с ATRA в
концентрации 10 мкМ и результаты иммуноблоттинга тех же линий; б - для подтверждения специфичности антител в от
ношении белков CRABP1 и CRABP2 приведены данные полученных производных сублиний клеток MDA MB 231 с ги
перэкспрессией CRABP1 (MDA MB 231 CRABP1) и экспрессией вектора без вставки (MDA MB 231 pLXSN)
БИОХИМИЯ том 86 вып. 2 2021
КОРРЕЛЯЦИЯ CRABP2, CRABP1 И РК ЧУВСТВИТЕЛЬНОСТИ
267
ция CRABP1 отсутствовала. Таким образом, для
водных сублиний на пуромицине. Эффектив
клеток РМЖ РК чувствительность либо не свя
ность обеих шпилечных конструкций оказалась
зана с экспрессией CRABP1, либо определяется
высокой. По данным денситометрического ана
совокупностью факторов, в числе которых мо
лиза результатов иммуноблоттинга, снижение
жет быть статус рецепторов ER, PR, HER2/neu и
продукции CRABP2 по сравнению с контроль
другие характеристики. В то же время получен
ными линиями, экспрессирующими последова
ные данные на линиях клеток РМЖ подтверди
тельность shRNA к зеленому флуоресцентному
ли нашу гипотезу о корреляции уровня белков
белку (shGFP), в разных линиях варьировала от
CRABP1 и CRABP2. Так, во всех без исключе
1,6 до 10 раз для sh1CRABP2 и от 2,5 до 10 раз для
ния линиях с выраженной экспрессией CRABP2
sh2CRABP2 (рис. 4). Анализ продукции белка
продукция CRABP1 также была представлена на
CRABP1 в производных сублиниях показал зна
высоком уровне; в линиях, где продукция
чительное снижение во всех клетках с нокдау
CRABP2 значительно снижена, CRABP1 почти
ном гена CRABP2 (рис. 4) по сравнению с соот
не детектировался; в линиях с отсутствием
ветствующими контролями. Это означает, что
CRABP2 также не детектировался и CRABP1.
уровень CRABP1 регулируется белком CRABP2.
Мы провели дополнительную проверку специ
Таким образом, мы впервые выявили нали
фичности работы антител, для чего трансдуци
чие строгой корреляции между экспрессией
ровали кодирующую последовательность
белков CRABP1 и CRABP2 и показали меха
CRABP1 в линию MDA MB 231, где отсутство
низм, при котором белок CRABP2 является вы
вала эндогенная экспрессия данного белка. Как
шестоящим регулятором (upstream regulator)
видно на рис. 3, б, гиперэкспрессия CRABP1 в
экспрессии CRABP1. Универсальность обнару
производной сублинии MDA MB 231 CRABP1
женного феномена свидетельствует о его биоло
хорошо детектируется антителами к данному
гической значимости. CRABP2 зависимая регу
белку, но не сопровождается увеличением
ляция продукции CRABP1 является дополни
экспрессии CRABP2, что свидетельствует, в
тельным механизмом реализации внутрикле
частности, об избирательности антител в отно
точной активности ретиноевой кислоты.
шении данных белков.
С учетом результатов анализа белков CRABP
в клетках отличного от РМЖ происхождения,
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
полученные результаты свидетельствуют об
универсальности обнаруженной нами корреля
Формирование РК резистентности является
ции между экспрессией белков CRABP1 и
одним из основных ограничений применения
CRABP2, что предполагает наличие регулятор
РК в клинической практике, несмотря на то, что
ной зависимости между данными белками. Важ
активная разработка ее и других натуральных
но отметить, что отсутствие CRABP1 наблюда
или синтетических ретиноидов в качестве про
лось при снижении CRABP2, но не наоборот -
тивоопухолевых препаратов ведется уже в тече
не было примеров сниженной продукции
ние долгого времени [28]. Механизмы приобре
CRABP1 и отсутствия CRABP2. Также важно,
тения устойчивости к РК до сих пор остаются не
что гиперэкспрессия CRABP1 в обоих случаях (в
до конца понятными. Одним из кандидатов на
линиях Н460 и MDA MB 231) не приводила к
роль «посредника» в формировании РК резис
изменению экспрессии CRABP2. Поэтому мы
тентности может быть белок CRABP1. Такое
предположили, что в данной паре белков
предположение основано как на его возможном
CRABP2, вероятно, является регулятором
участии в ограничении биодоступности РК в
CRABP1.
клетке за счет удержания ее в цитоплазме или
Для проверки этого предположения мы по
даже стимуляции ее катаболизма цитохромом
давили эндогенную экспрессию CRABP2 в 4 х
Cyp26A1 [18], так и на данных о метилировании
линиях РМЖ с различным уровнем РК чув
гена CRABP1 в ряде опухолей [29-33], которые
ствительности: РК чувствительных линиях
считаются преимущественно РК резистентны
MCF7 и T47D и РК резистентных линиях
ми. В то же время по нашим данным, в клетках
MDA MB 453 и MDA MB 468. Нокдаун прово
нейробластомы (классический пример РК чув
дили методом РНК интерференции с помощью
ствительных опухолей) CRABP1 экспрессирует
экспрессии предшественников малых шпилеч
ся на высоком уровне, причем его продукция
ных РНК (shRNA) к гену CRABP2. Две последо
коррелирует с уровнем дифференцировки тка
вательности shRNA (sh1CRABP2 и sh2CRABP2)
ней нейробластомы [34]. Предположительно,
были клонированы в вектор pLKO.1 puro и
CRABP1 может предохранять такие РК чув
трансдуцированы в клетки методом лентивирус
ствительные клетки от избытка РК. В то же вре
ной инфекции с последующим отбором произ
мя для ряда опухолей нами [22] и другими авто
БИОХИМИЯ том 86 вып. 2 2021
268
ЕНИКЕЕВ и др.
Рис. 4. Влияние нокдауна CRABP2 на продукцию CRABP1 в линиях РМЖ. Для анализа использованы производные
РК чувствительных (MCF7, T47D) и РК резистентных (MDA MB 453, MDA MB 468) линий РМЖ, экспрессирующих
последовательности малых шпилечных РНК к мРНК CRABP2 (sh1CRABP2 и sh2CRABP2). В качестве контролей использо
ваны те же линии, экспрессирующие малые шпилечные РНК к зеленому флуоресцентному белку (shGFP). Диаграммы
построены на основании данных денситометрического подсчета результатов трех независимых повторов эксперимента
рами [11, 12, 35, 36] показана опухоль промо
опухоль супрессорной активности РК, которая
торная роль CRABP1, которая может быть свя
осуществляется за счет ее доставки к ядерным
зана с конкурентным ограничением передачи
рецепторам RAR/RXR и дальнейшей активации
РК белку CRABP2, основному «проводнику»
транскрипции проапоптотических, продиффе
БИОХИМИЯ том 86 вып. 2 2021
КОРРЕЛЯЦИЯ CRABP2, CRABP1 И РК ЧУВСТВИТЕЛЬНОСТИ
269
ренцировочных и антипролиферативных генов
РК чувствительные клетки РМЖ были
(рис. 5, а). Иными словами, как опухоль промо
CRABP1 позитивными за исключением линии
торная, так и опухоль супрессорная (как в слу
SKBR3. Такие неоднозначные результаты не
чае нейробластомы) роль CRABP1 может быть
позволяют сделать четкий вывод об участии
связана с его функцией ограничения активнос
CRABP1 в формировании устойчивости клеток
ти РК. Возможно, именно разной чувствитель
к РК прежде всего потому, что механизмов
ностью конкретных опухолевых клеток к РК
РК резистентности может быть много, о чем
объясняются существенные противоречия в ли
свидетельствуют и данные литературы [40]. Так
тературных данных относительно функцио
же важно учитывать упомянутую гетерогенность
нального значения CRABP1 (опухоль промо
рака молочной железы, развитие резистентнос
торная или опухоль супрессорная роль) в кан
ти которого к РК может проходить по разным
церогенезе. Косвенным подтверждением учас
молекулярным сценариям [41].
тия CRABP1 в формировании РК резистент
Наиболее интересным результатом данного
ности могут служить и данные исследования, в
исследования можно считать обнаруженную
котором была показана связь между экспресси
100% ную корреляцию между экспрессией
ей CRABP1 и рецептор негативным статусом
CRABP2 и CRABP1. Следует подчеркнуть, что
РМЖ [15], который, в свою очередь, часто свя
этот феномен был подтвержден на всех типах
зан с РК резистентностью данного типа опухо
опухолевых клеток вне зависимости от их про
лей [37-39]. Мы предположили, что CRABP1
исхождения и РК чувствительности, что свиде
может способствовать приобретению РК резис
тельствует о его биологической значимости. На
тентности изначально РК чувствительных кле
основании полученных данных мы предполо
ток, ограничивая активность РК в отношении
жили, что продукция белков CRABP1 и
регуляции транскрипции РК зависимых генов.
CRABP2 является функционально связанной,
В РК резистентных клетках такие ограничения
причем вышестоящим регулятором является,
становятся «ненужными», что может приводить
по видимому, CRABP2. Ранее исследований
к метилированию гена CRABP1. В нашей работе
этого вопроса не проводилось. Данные един
мы впервые провели сравнение экспрессии
ственной работы, в которой анализировались
CRABP1 в клеточных линиях опухолей различ
уровни экспрессии и функциональное значение
ного происхождения и обнаружили, что в лини
обоих белков CRABP, свидетельствуют скорее о
ях клеток опухолей, считающихся РК резистент
«конкурентных взаимоотношениях» между бел
ными (три линии НМРЛ, две линии глиобласто
ками CRABP1 и CRABP2. Авторы статьи пред
мы, три линии рака яичников), продукция
полагают, что CRABP1 ограничивает функцию
CRABP1 отсутствует, в то время как в
CRABP2 в отношении реализации ядерной ак
РК чувствительных клетках (все три линии ней
тивности РК за счет удержания ее в цитоплазме
робластомы) данный белок детектируется на
[15]. Однако речь идет о функциональной кон
достаточно высоком уровне. РМЖ является
куренции, а не о регуляции количества белка. В
крайне гетерогенным типом опухолей, как по
этом контексте представляют интерес наши ра
морфологии, так и по молекулярным характе
нее опубликованные данные о корреляции меж
ристикам, включая чувствительность к РК. Для
ду экспрессией CRABP1 и CRABP2 в образцах
того, чтобы понять, действительно ли существу
тканей аденокарциномы легкого. Корреляция
ет прямая корреляция между экспрессией
была обнаружена нами как на уровне мРНК, так
CRABP1 и РК чувствительностью, мы провели
и на уровне белковой продукции, причем мак
анализ влияния РК на динамику пролиферации
симальное значение коэффициента корреляции
9 ти линий РМЖ и обнаружили широкий
Спирмена было характерно для высоко или
спектр реакции на ATRA, при этом линии хоро
умеренно дифференцированных опухолей, в то
шо разделялись на те, у которых значительно (в
время как в низкодифференцированных опухо
2 и более раз) снижалась пролиферация уже при
лях корреляция отсутствовала [21]. Эти данные
концентрации ATRA 1-10 мкМ (РК чувстви
косвенно указывают на регуляторную связь
тельные), и те, пролиферация которых не меня
между белками CRABP1 и CRABP2 и предпола
лась при концентрации ATRA 10-20 мкМ
гают наличие некоего регуляторного механизма,
(РК резистентные). Сравнение продукции
который, по видимому, утрачивается в ходе опу
CRABP1 в этих линиях не выявило корреляции
холевой прогрессии и дедифференцировки кле
с чувствительностью к РК, хотя следует отме
ток.
тить, что в максимально РК резистентных ли
Настоящее исследование подтвердило нали
ниях, пролиферация которых не менялась при
чие такой регуляции, а также показало, что
инкубации с ATRA даже в концентрации
CRABP2 является регулятором продукции
50 мкМ, продукция CRABP1 отсутствовала.
CRABP1. Как может осуществляться CRABP2
БИОХИМИЯ том 86 вып. 2 2021
270
ЕНИКЕЕВ и др.
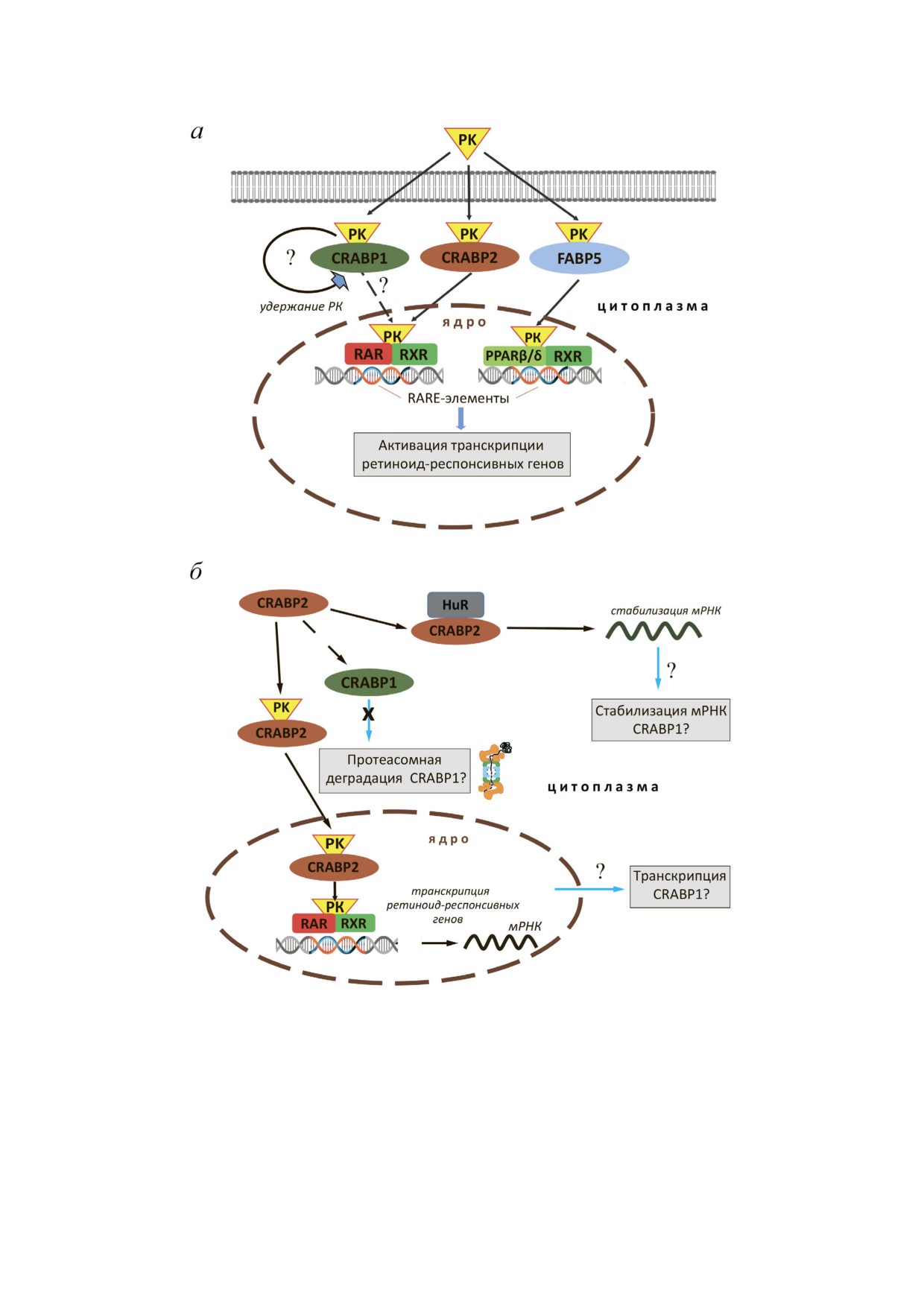
Рис. 5. Возможные механизмы влияния CRABP2 на внутриклеточный уровень белка CRABP1. а - Схема реализации
функциональной активности РК. Белок CRABP2 обеспечивает транспортировку РК в ядро и передачу ее ядерным рецеп
торам RAR. Белок FABP5 осуществляет доставку РК к ядерному рецептору PPARβ/δ. Формирование комплекса РК с ре
цепторами в составе гетеродимеров с RXR стимулирует взаимодействие с RARE в промоторах генов, привлечение ко
активаторов и активацию транскрипции. Сходная активность, предположительно, имеется и у белка CRABP1, однако, в
отличие от CRABP2, он не образует комплекса с рецепторами РК - предполагается, что передаче РК ядерным рецепто
рам в этом случае предшествует диссоциация комплекса РК-CRABP1; б - гипотетическая схема CRABP2 зависимой ре
гуляции продукции CRABP1. Такая регуляция может реализовываться с помощью CRABP2 зависимой активации тран
скрипции CRABP1, CRABP2 зависимой стабилизации мРНК CRABP1 при участии представителей группы Hu белков
(например, HuR), а также возможной негативной регуляции протеолитической деградации CRABP1
БИОХИМИЯ том 86 вып. 2 2021
КОРРЕЛЯЦИЯ CRABP2, CRABP1 И РК ЧУВСТВИТЕЛЬНОСТИ
271
зависимая регуляция белка CRABP1? Можно
тавки РК к ядерным рецепторам. Можно также
предположить несколько сценариев (рис. 5, б).
предположить, что CRABP2 зависимая регуля
Первый вариант предполагает прямую регу
ция происходит не на уровне транскрипции или
ляцию экспрессии, когда CRABP2 зависимая
посттранскрипционной регуляции мРНК, а ре
доставка РК к ядерным рецепторам приводит к
ализуется за счет протеолитической деградации
активации транскрипции CRABP1, в промоторе
CRABP1, активируемой в отсутствие CRABP2.
которого имеется РК респонсивный элемент.
Механизмы регуляции, реализуемые по этому
Помимо трансактивации транскрипции можно
сценарию, показаны для функционально свя
предположить наличие механизма CRABP2 за
занных или родственных белков. Характерным
висимой стабилизации транскриптов CRABP1.
примером могут служить белки флотиллин 1 и
Эта гипотеза основана на показанной в не
флотиллин 2, для которых неоднократно пока
скольких работах активности CRABP2, не свя
зано, что подавление одного из гомологов при
занной с РК и реализуемой с помощью взаимо
водит к снижению уровня другого [44-46]. Все
действия с представителями группы Hu белков,
вышеперечисленные возможные механизмы бу
относящихся к семейству ELAV (embryonic lethal
дут исследоваться нами в дальнейшем и способ
abnormal vision) РНК связывающих белков. Ос
ствовать пониманию фундаментальных процес
новная их функция связана с посттранскрипци
сов реализации внутриклеточной активности
онной регуляцией экспрессии генов и стабили
ретиноевой кислоты.
зацией транскриптов мРНК и некоторых других
типов РНК (например, микроРНК)
[42].
Финансирование. Работа выполнена при фи
CRABP2, по видимому, имеет несколько мише
нансовой поддержке Российского фонда фунда
ней из этой группы белков, в частности, белки
ментальных исследований (грант № 19 015
HuD и HuB [9, 10, 43]. Взаимодействие CRABP2
00027А).
с Hu белками приводит к их активации и как
Конфликт интересов. Авторы заявляют об от
результат - к повышению продукции целого ря
сутствии конфликта интересов.
да регуляторных и сигнальных белков. Интерес
Соблюдение этических норм. Настоящая
но, что HuR, в свою очередь, способствует
статья не содержит описания каких либо иссле
транспортировке CRABP2 в ядро, стимулируя
дований с участием людей или животных в каче
таким образом его активность в отношении дос
стве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Connolly, R. M., Nguyen, N. K., and Sukumar, S. (2013)
breast cancer: implication for retinoic acid therapy,
Molecular pathways: current role and future directions of
Am. J. Pathol., 178, 9971008, doi: 10.1016/j.ajpath.
the retinoic acid pathway in cancer prevention and treat
2010.11.075.
ment, Clin. Cancer Res., 19, 1651 1959, doi: 10.1158/
9. Vreeland, A. C., Levi, L., Zhang, W., Berry, D. C., and
1078 0432.CCR 12 3175.
Noy, N. (2014) Cellular retinoic acid binding protein 2
2.
Schenk, T., Stengel, S., and Zelent, A. (2014) Unlocking
inhibits tumor growth by two distinct mechanisms, J. Biol.
the potential of retinoic acid in anticancer therapy,
Chem., 289, 34065 34073, doi: 10.1074/jbc.M114.604041.
Br. J. Cancer, 111, 2039 2045, doi: 10.1038/bjc.2014.412.
10. Vreeland, A. C., Yu, S., Levi, L., de Barros Rossetto, D.,
3.
Chevkina, E. M., and Favorskaya, I. A. (2015) CRABP pro
and Noy, N. (2014) Transcript stabilization by the RNA
teins - relatives or namesakers? [in Russian], Uspekhi Mol.
binding protein HuR is regulated by cellular retinoic acid
Onkol., 2, 6 16, doi: 10.17650/2313 805X.2015.2.2.6 16.
binding protein 2, Mol. Cell. Biol., 34, 21352146,
4.
Tchevkina, E. M. (2017) Retinoic acid binding proteins
doi: 10.1128/mcb. 00281 14.
and cancer: similarity or polarity? Cancer Ther. Oncol.
11. Mallikarjuna, K., Sundaram, C. S., Sharma, Y., Deepa,
Int. J., 8, 555733, doi: 10.19080/ctoij.2017.08.555733.
P. R., Khetan, V., et al. (2010) Comparative proteomic
5.
Sussman, F., and De Lera, A. R. (2005) Ligand recognition
analysis of differentially expressed proteins in primary
by RAR and RXR receptors: binding and selectivity,
retinoblastoma tumors, Proteom. Clin. Appl., 4, 449 463,
J. Med. Chem., 48, 6212 6219, doi: 10.1021/jm050285w.
doi: 10.1002/prca.200900069.
6.
Schug, T. T., Berry, D. C., Shaw, N. S., Travis, S. N., and
12. Liu, R. Z., Li, S., Garcia, E., Glubrecht, D. D., Yin
Noy, N. (2007) Opposing effects of retinoic acid on cell
Poon, H., et al. (2016) Association between cytoplasmic
growth result from alternate activation of two different
CRABP2, altered retinoic acid signaling, and poor progno
nuclear receptors, Cell, 129, 723733, doi: 10.1016/
sis in glioblastoma, Glia, 64, 963976, doi: 10.1002/
j.cell.2007.02.050.
glia.22976.
7.
Schug, T. T., Berry, D. C., Toshkov, I. A., Cheng, L.,
13. Dong, D., Ruuska, S. E., Levinthal, D. J., and Noy, N.
Nikitin, A. Y., and Noy, N. (2008) Overcoming retinoic
(1999) Distinct roles for cellular retinoic acid binding pro
acid resistance of mammary carcinomas by diverting
teins I and II in regulating signaling by retinoic acid,
retinoic acid from PPARβ/δ to RAR, Proc. Natl. Acad. Sci.
J. Biol. Chem., 274, 23695 23698, doi: 10.1074/jbc.274.34.
USA, 105, 7546 7551, doi: 10.1073/pnas.0709981105.
23695.
8.
Liu, R. Z., Graham, K., Glubrecht, D. D., Germain,
14. Blaese, M. A., Santo Hoeltje, L., and Rodemann, H. P.
D. R., Mackey, J. R., and Godbout, R. (2011) Association
(2003) CRABP I expression and the mediation of the sen
of FABP5 expression with poor survival in triple negative
sitivity of human tumour cells to retinoic acid and irradia
БИОХИМИЯ том 86 вып. 2 2021
272
ЕНИКЕЕВ и др.
tion, Int. J. Radiat. Biol., 79, 981 991, doi:
10.1080/
cancer and its promise for therapy, J. Carcinogen. Mutagen.,
09553000310001632949.
doi: 10.4172/2157 2518.s7 006.
15.
Liu, R. Z., Garcia, E., Glubrecht, D. D., Poon, H. Y.,
29.
Miyake, T., Ueda, Y., Matsuzaki, S., Miyatake, T.,
Mackey, J. R., and Godbout, R. (2015) CRABP1 is associ
Yoshino, K., et al. (2011) CRABP1 reduced expression is
ated with a poor prognosis in breast cancer: adding to the
associated with poorer prognosis in serous and clear cell
complexity of breast cancer cell response to retinoic acid,
ovarian adenocarcinoma, J. Cancer Res. Clin. Oncol., 137,
Mol. Cancer, 14, 129, doi: 10.1186/s12943 015 0380 7.
715 722, doi: 10.1007/s00432 010 0930 8.
16.
Fiorella, P. D., and Napoli, J. L. (1991) Expression of cel
30.
Tanaka, K., Imoto, I., Inoue, J., Kozaki, K., Tsuda, H.,
lular retinoic acid binding protein (CRABP) in Escherichia
et al. (2007) Frequent methylation associated silencing of
coli. Characterization and evidence that holo CRABP is a
a candidate tumor suppressor, CRABP1, in esophageal
substrate in retinoic acid metabolism, J. Biol. Chem., 266,
squamous cell carcinoma, Oncogene, 26, 64566468,
16572 16579.
doi: 10.1038/sj.onc.1210459.
17.
Boylan, J. F., and Gudas, L. J. (1992) The level of CRABP
31.
Lind, G. E., Kleivi, K., Meling, G. I., Teixeira, M. R.,
I expression influences the amounts and types of all trans
Thiis Evensen, E., et al. (2006) ADAMTS1, CRABP1, and
retinoic acid metabolites in F9 teratocarcinoma stem cells,
NR3C1 identified as epigenetically deregulated genes in
J. Biol. Chem., 267, 21486 21491.
colorectal tumorigenesis, Cell. Oncol., 28, 259272,
18.
Won, J. Y., Nam, E. C., Yoo, S. J., Kwon, H. J., Um, S. J.,
doi: 10.1155/2006/949506.
et al. (2004) The effect of cellular retinoic acid binding pro
32.
Wu, Q., Lothe, R. A., Ahlquist, T., Silins, I., Tropé, C. G.,
tein I expression on the CYP26 mediated catabolism of
et al. (2007) DNA methylation profiling of ovarian carci
all trans retinoic acid and cell proliferation in head and
nomas and their in vitro models identifies HOXA9,
neck squamous cell carcinoma, Metab. Clin. Exp., 53,
HOXB5, SCGB3A1, and CRABP1 as novel targets, Mol.
1007 1012, doi: 10.1016/j.metabol.2003.12.015.
Cancer, 6, 42, doi: 10.1186/1476 4598 6 45.
19.
Delektorskaya, V. V., Komel’kov, A. V., Zborovskaya, I. B.,
33.
Wang, F., Yang, Y., Fu, Z., Xu, N., Chen, F., et al. (2014)
Enikeev, A. D., Safronova, V. M., and Chevkina, E. M.
Differential DNA methylation status between breast carci
(2017) Nuclear localization of cellular retinoic acid bind
nomatous and normal tissues, Biomed. Pharmacother., 68,
ing protein 1 (Crabp1) is associated with malignancy level
699 707, doi: 10.1016/j.biopha.2014.07.014.
in lung neuroendocrine tumors [in Russian], Voprosy
34.
Stroganova, A. M., Chemeris, G. Yu., Chevkina, E. M.,
Onkologii, 63, 886 893.
Senderovich, A., Karseladze, A. I. (2016) CRABP protein
20.
Gaub, M. P., Lutz, Y., Ghyselinck, N. B., Scheuer, I.,
1 and its role in the process of differentiation neuroblas
Pfister, V., et al. (1998) Nuclear detection of cellular
toma, Vestnik RONTs im. N. N. Blokhina, 27, 157 163.
retinoic acid binding proteins I and II with new antibodies,
35.
Bertucci, F., Houlgatte, R., Benziane, A., Granjeaud, S.,
J. Histochem. Cytochem., 46, 1103 1111, doi: 10.1177/
Adéla de, J., et al. (2000) Gene expression profiling of pri
002215549804601002.
mary breast carcinomas using arrays of candidate genes,
21.
Favorskaya, I., Kainov, Y., Chemeris, G., Komelkov, A.,
Hum. Mol. Genet., 9, 2981 2991, doi: 10.1093/hmg/9.20.
Zborovskaya, I., and Tchevkina, E. (2014) Expression and
2981.
clinical significance of CRABP1 and CRABP2 in non
36.
Tsibris, J. C. M., Segars, J., Coppola, D., Mane, S.,
small cell lung cancer, Tumor Biol., 35, 10295 10300,
Wilbanks, G. D., et al. (2002) Insights from gene arrays on
doi: 10.1007/s13277 014 2348 4.
the development and growth regulation of uterine leiomy
22.
Kainov, Y., Favorskaya, I., Delektorskaya, V.,
omata, Fertil. Steril., 78, 114 121, doi: 10.1016/S0015
Chemeris, G., Komelkov, A., et al. (2014) CRABP1 pro
0282(02)03191 6.
vides high malignancy of transformed mesenchymal cells
37.
Fontana, J. A. (1992) Responses to retinoic acid of tamox
and contributes to the pathogenesis of mesenchymal and
ifen sensitive and resistant sublines of human breast can
neuroendocrine tumors, Cell Cycle,
13,
15301539,
cer cell line MCF 7, Cancer Res., 52, 6164 6167.
doi: 10.4161/cc.28475.
38.
Fontana, J. A. (1987) Interaction of retinoids and tamox
23.
Rossetti, S., and Sacchi, N. (2019) 3D mammary epithelial
ifen on the inhibition of human mammary carcinoma cell
cell models: a goldmine of dcis biomarkers and morpho
proliferation, Pathobiology, 55, 136 144, doi: 10.1159/
genetic mechanisms, Cancers, 11, 130, doi: 10.3390/can
000163409.
cers11020130.
39.
Van der Leede, B. J. M., Folkers, G. E., van den Brink, C.
24.
Garattini, E., Bolis, M., Garattini, S. K., Fratelli, M.,
E., van der Saag, P. T., and van der Burg, B. (1995)
Centritto, F., et al. (2014) Retinoids and breast cancer:
Retinoic acid receptor α1 isoform is induced by estradiol
from basic studies to the clinic and back again, Cancer
and confers retinoic acid sensitivity in human breast cancer
Treat. Rev., 40, 739 749, doi: 10.1016/j.ctrv.2014.01.001.
cells, Mol. Cell. Endocrinol., 109, 77 86, doi: 10.1016/
25.
Coyle, K. M., Dean, C. A., Thomas, M. L., Vidovic, D.,
0303 7207(95)03487 R.
Giacomantonio, C. A., et al. (2018) DNA methylation
40.
Chlapek, P., Slavikova, V., Mazanek, P., Sterba, J., and
predicts the response of triple negative breast cancers to
Veselska, R. (2018) Why differentiation therapy sometimes
all trans retinoic acid, Cancers, 10, 397, doi: 10.3390/can
fails: Molecular mechanisms of resistance to retinoids,
cers10110397.
Int. J. Mol. Sci., 19, 132, doi: 10.3390/ijms19010132.
26.
Centritto, F., Paroni, G., Bolis, M., Garattini, S. K.,
41.
Tari, A. M., Lim, S. J., Hung, M. C., Esteva, F. J., and
Kurosaki, M., et al. (2015) Cellular and molecular deter
Lopez Berestein, G. (2002) Her2/neu induces all trans
minants of all trans retinoic acid sensitivity in breast can
retinoic acid (ATRA) resistance in breast cancer cells,
cer: luminal phenotype and RARα expression, EMBO
Oncogene, 21, 5224 5232, doi: 10.1038/sj.onc.1205660.
Mol. Med., 7, 950 972, doi: 10.15252/emmm.201404670.
42.
Wang, J., Guo, Y., Chu, H., Guan, Y., Bi, J., and Wang, B.
27.
Bolis, M., Garattini, E., Paroni, G., Zanetti, A.,
(2013) Multiple functions of the RNA binding protein
Kurosaki, M., et al. (2017) Network guided modeling
HuR in cancer progression, treatment responses and prog
allows tumor type independent prediction of sensitivity to
nosis, Int. J. Mol. Sci., 14, 10015 10041, doi: 10.3390/
all trans retinoic acid, Ann. Oncology, 28, 611621,
ijms140510015.
doi: 10.1093/annonc/mdw660.
43.
Gupta, A., Williams, B. R. G., Hanash, S. M., and
28.
Coyle, K. M., Sultan, M., Thomas, M. L., Vaghar
Rawwas, J. (2006) Cellular retinoic acid binding protein II
Kashani, A., and Marcato, P. (2013) Retinoid signaling in
is a direct transcriptional target of MycN in neuroblas
БИОХИМИЯ том 86 вып. 2 2021
КОРРЕЛЯЦИЯ CRABP2, CRABP1 И РК ЧУВСТВИТЕЛЬНОСТИ
273
toma, Cancer Res., 66, 8100 8108, doi: 10.1158/0008
flotillins induces formation of membrane microdomains,
5472.CAN 05 4519.
membrane curvature, and vesicle budding, Curr. Biol., 17,
44. Babuke, T., Ruonala, M., Meister, M., Amaddii, M.,
1151 1156, doi: 10.1016/j.cub.2007.05.078.
Genzler, C., Esposito, A., and Tikkanen, R.
(2009)
46. Solis, G. P., Hoegg, M., Munderloh, C., Schrock, Y.,
Hetero oligomerization of reggie 1/flotillin 2 and reggie
Malaga Trillo, E., Rivera Milla, E., and Stuermer,
2/flotillin 1 is required for their endocytosis, Cell. Signal.,
C. A. O. (2007) Reggie/flotillin proteins are organized into
21, 1287 1297, doi: 10.1016/j.cellsig.2009.03.012.
stable tetramers in membrane microdomains, Biochem. J.,
45. Frick, M., Bright, N. A., Riento, K., Bray, A.,
403, 313 322, doi: 10.1042/BJ20061686.
Merrified, C., and Nichols, B. J. (2007) Coassembly of
CRABP1 AND CRABP2 PROTEIN LEVELS DO NOT CORRELATE
WITH THE SENSITIVITY OF BREAST CANCER CELLS TO RETINOIC
ACID, BUT CORRELATE WITH EACH OTHER WITH CRABP2
BEING AN UPSTREAM REGULATOR OF CRABP1 PRODUCTION*
A. D. Enikeev, A. V. Komelkov**, M. E. Axelrod, S. A. Galetsky,
S. А. Kuzmichev, and Е. М. Tchevkina
Blokhin National Medical Research Center of Oncology, 115478 Moscow, Russia; E'mail: komelkov@gmail.com
Retinoic acid (RA) binding proteins, CRABP1 and CRABP2, are molecular chaperones that mediate intracellular
activity of RA, the key promoter of cell differentiation with tumor suppressor activity. One of the main functions of
CRABP2 is delivery and transfer of RA to the nuclear receptors RAR/RXR, which leads to activation of the tran
scription of a wide range of retinoid responsive genes. The functions of CRABP1 are less studied but are apparently
associated with sequestration of RA in cytoplasm and limitation of its transcriptional activity, suggesting involvement
of this protein in the development of RA resistance. The mechanisms regulating activity of CRABP1 are also poorly
understood. Comparison of the CRABP1 level in tumor cell lines of various origins, performed for the first time here,
showed absence of the CRABP1 protein in the cell lines of tumors considered to be RA resistant, and pronounced
production of this protein in the RA sensitive cells. However, analysis carried out with a panel of breast cancer cell
lines with different levels of RA sensitivity showed that there was no correlation between the production of CRABP1
protein and the sensitivity of the cells to RA. At the same time, we found strong correlation between the expression of
CRABP1 and CRABP2 proteins in all studied cell types, regardless of their origin and RA sensitivity/resistance.
Moreover, suppression of the CRABP1 level in both RA sensitive and RA resistant cells was shown in the cells with
cells with knockdown of CRABP2 gene. The revealed CRABP2 dependent regulation of CRABP1 production is a new
mechanism of the intracellular retinoic signaling system.
Keywords: retinoic acid, retinoic acid binding proteins, ATRA, CRABP1, CRABP2, proliferation, expression regu
lation
9 БИОХИМИЯ том 86 вып. 2 2021