БИОХИМИЯ, 2021, том 86, вып. 6, с. 787 - 804
УДК 57.052;57.053;57.054
АССОЦИИРОВАННЫЕ СО СТРЕССОМ ГИППОКАМПАЛЬНЫЕ
МОЛЕКУЛЯРНО КЛЕТОЧНЫЕ МЕХАНИЗМЫ,
ОБЩИЕ ДЛЯ ЭПИЛЕПСИИ И КОМОРБИДНЫХ
ДЕПРЕССИВНЫХ РАССТРОЙСТВ
Обзор
© 2021
Н.В. Гуляева1,2
1 Институт высшей нервной деятельности и нейрофизиологии Российской академии наук,
117485 Москва, Россия; электронная почта: nata_gul@ihna.ru
2 Научно'практический психоневрологический центр имени З.П. Соловьева, 115419 Москва, Россия
Поступила в редакцию 24.02.2021
После доработки 30.03.2021
Принята к публикации 30.03.2021
В обзоре рассматриваются молекулярно клеточные механизмы, общие для эпилептических и депрессивных
расстройств. Коморбидные эпилепсия височной доли и депрессия ассоциированы с нарушениями функцио
нирования гипоталамо гипофизарно адренокортикальной оси и вызванными избытком глюкокортикои
дов нарушениями функции и структуры гиппокампа, ключевого региона мозга для обучения/памяти и эмо
ций. Селективная чувствительность гиппокампа к стрессу, опосредованная рецепцией секретируемых при
стрессе глюкокортикоидных гормонов, является «ценой» высокой функциональной пластичности и
плейотропности этой лимбической структуры. Общие молекулярно клеточные механизмы включают нару
шения функции глюкокортикоидных рецепторов, нейротрансмиттерных систем, нейротрофических фак
торов и развитие нейровоспаления, приводящие к нейродегенерации и гибели нейронов гиппокампа. На
ряду с этим происходит нарушение нейрогенеза в субгранулярной нейрогенной нише и формирование
аберрантных нейронных сетей. Эти глюкокортикоид зависимые процессы лежат в основе измененной
стресс реактивности и развития ассоциированных с хроническим стрессом коморбидных патологий, в
частности височной эпилепсии и депрессивных расстройств.
КЛЮЧЕВЫЕ СЛОВА: эпилепсия, эпилепсия височной доли, депрессия, гиппокамп, глюкокортикоиды,
стресс, нейровоспаление, гипоталамо гипофизарно адренокортикальная ось, нейрогенез, нейронные сети.
DOI: 10.31857/S0320972521060038
ВВЕДЕНИЕ
и структурных изменений в мозге, позволяю
щих адаптироваться к условиям внешней среды,
Эпилепсия является одним из самых распро
обучаться, запоминать информацию, а также
страненных неврологических заболеваний, от
восстанавливать мозг после повреждения), в ос
которого в мире страдает около 70 млн человек.
нове которой лежат биохимические процессы в
Оно характеризуется периодическими спонтан
синапсах и других компартментах нейронов [1].
ными судорогами, обусловленными гипервозбу
Эпилепсия характеризуется судорожными
димостью и гиперсинхронизацией нейронов
припадками, но также часто может быть ассоции
мозга. Эпилептические расстройства ассоции
рована с коморбидными когнитивными и пси
рованы с различными этиологическими факто
хическими расстройствами [2]. Распространен
рами - генетическими, инфекционными, трав
ность сопутствующих психических заболеваний
матическими, сосудистыми, токсическими,
у пациентов с эпилепсией относительно высо
опухолевыми и др. Почти у трети пациентов
ка: каждый третий пациент в течение своей жиз
эпилепсия фармакорезистентна. Эпилептичес
ни испытывал психическое расстройство, как
кий мозг является примером хронического на
правило, расстройства настроения и тревож
рушения нейропластичности (функциональных
ность. Психические коморбидные заболевания
часто предшествуют возникновению судорож
Принятые сокращения: BDNF - нейротрофичес
ного расстройства и влияют на качество жизни
кий фактор мозга; ГГАКС - гипоталамо гипофизарно ад
пациентов и течение эпилепсии в нескольких
ренокортикальная система (ось); ГК - глюкокортикоиды;
ГР - глюкокортикоидные рецепторы; ЭВД - эпилепсия
аспектах. В частности, это худшая переноси
височной доли.
мость фармакотерапии противоэпилептически
787
3*
788
ГУЛЯЕВА
ми препаратами, развитие ятрогенных психиат
склерозом гиппокампа. Для пациентов с реф
рических симптомов от фармакологического и
рактерной эпилепсией характерна высокая час
хирургического лечения, повышенный риск
тота дисфункциональных личностных паттер
смертности и увеличение экономического бре
нов, различающихся в зависимости от эпилеп
мени для пациента, семьи и общества в целом.
тогенной зоны [7, 8]. Интересно, что к факторам
Важность проблемы такова, что идет обсужде
риска коморбидной депрессии при ЭВД, поми
ние среди специалистов, не следует ли включить
мо фармакорезистентности, относится и
сопутствующие психиатрические заболевания в
пол: женщины страдают чаще, что в целом ха
классификацию эпилепсий [3].
рактерно для депрессивных расстройств [9].
Депрессия - одно из наиболее инвалидизи
рующих из сопутствующих эпилепсии заболева
ний, она оказывает глубокое негативное влия
ПАТОГЕНЕТИЧЕСКАЯ КОНЦЕПЦИЯ
ние на качество жизни пациентов с эпилепсией.
КОМОРБИДНОСТИ ЭПИЛЕПСИИ
Связь между депрессией и эпилепсией, извест
И ДЕПРЕССИИ
ная с древних времен, до настоящего времени до
конца не изучена. Долгое время депрессивные
Существование общих патогенетических ме
расстройства рассматривались только как выра
ханизмов при первичных психических рас
жение реактивного процесса на стрессорные
стройствах и эпилепсии было постулировано в
факторы существования больного эпилепсией.
качестве одной из ведущих гипотез для объясне
Однако эпилепсия и сопутствующие психичес
ния их тесной взаимосвязи. Для проверки этой
кие заболевания имеют сложную взаимозависи
гипотезы применяют анализ нейробиологичес
мость, которая может проявляться как их высо
ких характеристик расстройств настроения и
кой коморбидностью, так и наличием двуна
эпилепсии [4]. На животных моделях были по
правленной связи, в результате чего не только
лучены данные о том, как нейробиологические
люди с эпилепсией подвергаются большему
аспекты расстройств настроения могут индуци
риску развития психических расстройств, но и
ровать эпилептогенный процесс. Анализ этих
пациенты с первичными психическими рас
данных помогает объяснить повышенный риск
стройствами подвергаются более высокому рис
развития эпилепсии, в частности фармакоре
ку развития эпилепсии [4]. Такая связь может
зистентной ЭВД, у пациентов с первичными
быть объяснена только наличием общих патоге
расстройствами настроения. Указанные иссле
нетических механизмов этих заболеваний.
дования подтверждают существование общих
Действительно, полученные за последние три
патогенетических механизмов, функционирую
десятилетия данные продемонстрировали об
щих как при депрессивных и, вероятно, тревож
щие для эпилепсии и депрессии биохимичес
ных расстройствах, так и при эпилепсии, и мо
кие, нейропатологические и нейрофизиологи
гут объяснить их относительно высокую комор
ческие изменения. Они подтверждены исследо
бидность, двунаправленную связь и худшее те
ваниями на животных моделях и опосредуют
чение судорожного расстройства, связанного с
как развитие расстройств настроения, так и раз
предшествующим анамнезом депрессии [2, 4].
витие судорожной активности.
Авторы этого анализа справедливо отмечают,
Среди множества различных форм эпилеп
что имеющиеся данные представляют собой
сии наиболее распространенным эпилептичес
лишь вершину айсберга, а исследования в этой
ким синдромом у взрослых является эпилепсия
области только начинаются.
височной доли (ЭВД). По разным данным от 30
При изучении моделей эпилепсии на живот
до 70% пациентов с ЭВД имеют нервно психи
ных депрессивно подобное поведение отчаяния
ческие осложнения, наиболее часто это рас
(неподвижность в тесте принудительного плава
стройства депрессивного спектра. Коморбид
ния) и ангедония (отсутствие предпочтения
ные расстройства настроения у пациентов с
подслащенной воды) были описаны на модели
ЭВД включают расстройства личности (особен
абсансной эпилепсии (линия крыс WAG/Rij), а
но обсессивно компульсивные расстройства) и
также на моделях ЭВД. Литий пилокарпиновая
«большую» депрессию (major depressive disorder),
модель эпилептического статуса с последующим
наиболее часто проявляющиеся при фармакоре
периодом эпилептогенеза в течение нескольких
зистентной (лекарственно устойчивой, рефрак
недель является одной из наиболее адекватных
терной) ЭВД с гиппокампальным (мезиальным
моделей ЭВД. У животных при этом наблюда
височным) склерозом [5, 6]. Частота депрессии
ются повреждения мозга, в первую очередь гип
колеблется от 20 до 55% у пациентов с рефрак
покампа, и изменения нейропластичности, на
терной эпилепсией, большинство которых сос
поминающие таковые при ЭВД у человека. Обе
тавляют пациенты с ЭВД, ассоциированной со
модели эпилепсии вызывают развитие поведен
БИОХИМИЯ том 86 вып. 6 2021
СТРЕСС, ГИППОКАМП, ЭПИЛЕПСИЯ И КОМОРБИДНАЯ ДЕПРЕССИЯ
789
ческих симптомов отчаяния, ангедонию и изме
покамп, структура лимбической системы с мак
нения в серотонинергических путях гиппокампа
симальной плотностью кортикостероидных ре
[10], а также ряд других биохимических измене
цепторов, является важной как для когнитив
ний. Эти изменения непосредственно или опо
ной функции (дорсальный/задний гиппокамп),
средованно затрагивают гиппокамп, включая
так и для эмоциональной сферы (вентраль
избыточную активацию гипоталамо гипофи
ный/передний гиппокамп). Соответственно,
зарно адренокортикальной системы (ГГАКС) и
именно регуляция функции гиппокампа гормо
провоспалительные процессы.
нами стресса становится ключевой для реакции
организма на стресс [21, 23]. Важно также, что в
отличие от других структур мозга, в гиппокампе
ЭПИЛЕПСИЯ КАК МОДЕЛЬ
базальная мембрана покрывает только 30% по
ХРОНИЧЕСКОГО СТРЕССА
верхности сосудов [24], что облегчает доставку
гормонов в нейроны. Сбалансированный ответ
Депрессивные расстройства относят к
в обеих фазах острого стресса, на пике ГК и при
стресс зависимым заболеваниям, для которых
последующем снижении их уровня, необходим
ключевая роль различных стрессорных факто
для преодоления стресса. Однако при хроничес
ров (в т.ч. стресса в раннем возрасте и хроничес
ком стрессе этот баланс нарушается, особенно у
кого стресса) и нарушения стресс реактивности
генетически уязвимых индивидов, и это приво
в патогенезе заболевания подтверждена много
дит к усилению проявления заболевания. Это
численными экспериментальными [11-14] и
касается и психических расстройств, и невроло
клиническими данными [15-20]. Однако роли
гических заболеваний, в частности эпилепсии.
стрессорных факторов в эпилептогенезе и при
Как системы стресс реактивности, так и судо
эпилепсии уделяется значительно меньше вни
рожная активность существенно зависят от эн
мания. Результаты клинических исследований,
догенных нейростероидов, обладающих важной
а также данные животных моделей о роли стрес
нейрофизиологической активностью. Нейро
сорных факторов в патогенезе эпилепсии свиде
стероиды взаимодействуют с глутамат , серото
тельствуют о том, что стресс является важным
нин , адрен и ГАМКергической системами, по
фактором развития эпилепсии путем ряда меха
тенциал зависимыми катионными и анионны
низмов, а само заболевание может быть рас
ми каналами, системами нейротрофинов и др.
смотрено как модель хронического стресса.
Нарушения уровней нейростероидов ассоции
Гормоны стресса глюкокортикоиды (ГК -
рованы с нейровоспалением и стресс индуци
кортизол у человека и кортикостерон у лабора
рованными заболеваниями, а также депрессией
торных грызунов), секретируемые в кровь в ре
и эпилепсией [25].
зультате функционирования ГГАКС, оказывают
К настоящему времени стало очевидным,
непрерывное действие на мозг. Естественные
что факторы стресса являются ключевыми в па
изменения их уровней происходят в результате
тогенезе эпилепсии на ранних ее стадиях. Судо
ультрадианных или циркадианных ритмов и в
рожная активность является одним из послед
стрессогенных ситуациях [21]. Клетки мозга
ствий раннего стресса; и хотя лежащие в основе
экспрессируют два типа кортикостероидных ре
этого феномена механизмы остаются малоизу
цепторов, глюкокортикоидные (ГР) и минера
ченными, имеются свидетельства вовлеченнос
локортикоидные (МР), различающиеся по регио
ти в этот процесс глутамат , ГАМК и адренер
нарному распределению и аффинности. Эти ре
гической систем [26].
цепторы опосредуют как быстрые негеномные,
Для исследования ассоциации эпилепсии и
так и более медленные геномные эффекты в
стресса у пациентов используется несколько
нейронах (мембранные и цитоплазматические
подходов. При опросе пациенты с эпилепсией
кортикостероидные рецепторы соответствен
зачастую сообщают, что именно стрессорные
но). Именно поэтому естественные, в частности
события запускают у них судорожные приступы.
вызванные стрессом, сдвиги в уровне ГК ассо
Тем не менее доклинические, эпидемиологичес
циированы со сложным временным и регионар
кие и клинические исследования связи стресса с
ным паттерном нейронной активности. Изме
эпилепсией дают довольно противоречивые ре
нения активности клеток мозга и соответствую
зультаты. Экспериментальные результаты боль
щих нейронных сетей приводят к когнитивным
шинства доклинических исследований подтвер
и эмоциональным сдвигам [22]. Непосредствен
ждают, что хронический стресс усугубляет про
но после стресса на пике концентрации ГК по
явления эпилепсии, а в некоторых случаях мо
вышается активность областей мозга, вовлечен
жет служить ее причиной [27, 28], однако дан
ных в эмоциональные ответы, а в последующий
ные клинических исследований не так одно
период - в высшие когнитивные функции. Гип
значны. Негативное влияние депрессии на
БИОХИМИЯ том 86 вып. 6 2021
790
ГУЛЯЕВА
контроль судорожных расстройств было задоку
Как было указано выше, наиболее частой ко
ментировано в различных исследованиях [29]. У
морбидностью эпилепсии являются депрессив
большинства пациентов с эпилепсией, которые
ные эпизоды, встречаемость которых, в зависи
отмечают стресс как индуктор приступов, обна
мости от исследуемых когорт, может превышать
руживаются тревожность, депрессия или детс
50% [34]. Депрессия относится к заболеваниям,
кая травма в анамнезе, в отличие от тех, которые
в основе патогенеза которых лежит неспособ
не считают стресс важным фактором начала
ность пациента адекватно справляться со стрес
приступов. При этом большинство пациентов
сорными нагрузками. Проявления депрессии
со стресс индуцированными судорогами, ис
при эпилепсии включают много взаимодейству
пользующих методы снижения уровня стресса
ющих нейробиологических и психосоциальных
для облегчения приступов, считают этот подход
детерминант, в т.ч. клинические проявления
эффективным [30]. Несмотря на то что механиз
эпилепсии (частоту и тип приступов, источник
мы проэпилептических эффектов стресса до сих
и его латерализацию), нейрохимические и ятро
пор остаются недостаточно изученными, веро
генные механизмы. Наиболее понятным и реа
ятно, для целей формализации анализа биоло
листичным (хотя лишь частичным) объяснени
гических механизмов патогенеза эпилепсии ее
ем депрессивной симптоматики при эпилепсии
целесообразно рассматривать как модель хро
является тот факт, что больной находится под
нического стресса.
действием хронического мультифакторного
Медиаторы стресса, гормоны и нейротранс
стресса. Неопределенность и непредсказуемость
миттеры связываются с рецепторами в различ
приступов вызывает чувства уныния, одиноче
ных структурах мозга, и в первую очередь в гип
ства, отчаяния, низкую самооценку и самоупре
покампе, чувствительном к стрессу и легко раз
ки, что может привести к социальной изоляции,
вивающем судорожную активность. Взаимодей
социальной стигматизации и бессилию. Депрес
ствие медиаторов стресса с рецепторами в гип
сия часто рассматривается как реакция на соци
покампе влияет на возбудимость отдельных ней
альную стигму и связанное с ней пониженное
ронов и нервных сетей. Важно, что в зависимос
качество жизни, а частота депрессии и суици
ти от многих факторов медиаторы демонстриру
дальности у пациентов с эпилепсией в 4-5 раз
ют уникальные пространственно временные
выше, чем в здоровой популяции [35]. Симпто
профили, определяющие природу сложных ком
мы тревожности также часто ассоциированы с
бинаторных эффектов на возбудимость сетей
эпилепсией, хотя в целом независимы от прис
гиппокампа и генерацию судорог [31]. Несмотря
тупов. Нередко они проявляются перииктально
на многочисленные клинические свидетельства
(«около» судорожного приступа) и классифици
того, что стресс может быть триггерным факто
руются в соответствии с этим на преиктальные,
ром развития приступов у пациентов с эпилеп
иктальные и постиктальные, хотя, как полагают,
сией, у этих пациентов мало изучено состояние
в целом происходят независимо от приступов.
ГГАКС, определяющее стресс реактивность.
Как и в случае депрессии, отмечается двусто
Невзирая на существенный разброс данных, ко
ронняя патофизиологическая связь между тре
торый может быть отчасти связан с приемом
вожным расстройством и эпилепсией. У некото
противоэпилептических препаратов, в большин
рых пациентов тревога инициирует приступы;
стве исследований выявлены ассоциации прис
для понимания и объяснения этого феномена
тупов с усилением стрессорной нагрузки и повы
используют концепции стресса и возбуждения
шением уровня кортизола [32]. При этом высо
(arousal) [36]. Приступы, источник которых на
кая частота приступов ассоциирована с повыше
ходится в лимбических сетях, включающих
нием уровнем кортизола и более низкой функ
амигдалу, могут иметь симптомологию, связан
циональной коннективностью мозга. Выражен
ную со страхом.
ность психологического дистресса, реализован
Анализ имеющихся к настоящему времени
ная в первую очередь в депрессивных и тревож
данных свидетельствует о том, что факторы
ных расстройствах, при эпилепсии существенно
стресса, действующие в раннем онтогенезе или
выше, чем при многих других заболеваниях. Ряд
на взрослый организм, могут являться эпилепто
исследователей предполагает, что эти коморбид
генными и инициировать развитие эпилепсии.
ные состояния вносят даже больший вклад в
Эти факторы могут быть хроническими и
ухудшение качества жизни пациентов, чем сама
действовать при эпилептогенезе и развившемся
эпилепсия [33]. Очевидно, что нейробиологи
заболевании, либо однократно, запуская моле
ческая связь между психиатрическими расстрой
кулярные процессы, лежащие в основе эпилеп
ствами и эпилепсией в существенной степени
сии и коморбидной депрессии (подробно рас
опосредована стрессом, вовлеченным в психо и
смотрены в статьях [27, 37, 38]). На пациента с
патофизиологические механизмы.
эпилепсией действует целый комплекс хрони
БИОХИМИЯ том 86 вып. 6 2021
СТРЕСС, ГИППОКАМП, ЭПИЛЕПСИЯ И КОМОРБИДНАЯ ДЕПРЕССИЯ
791
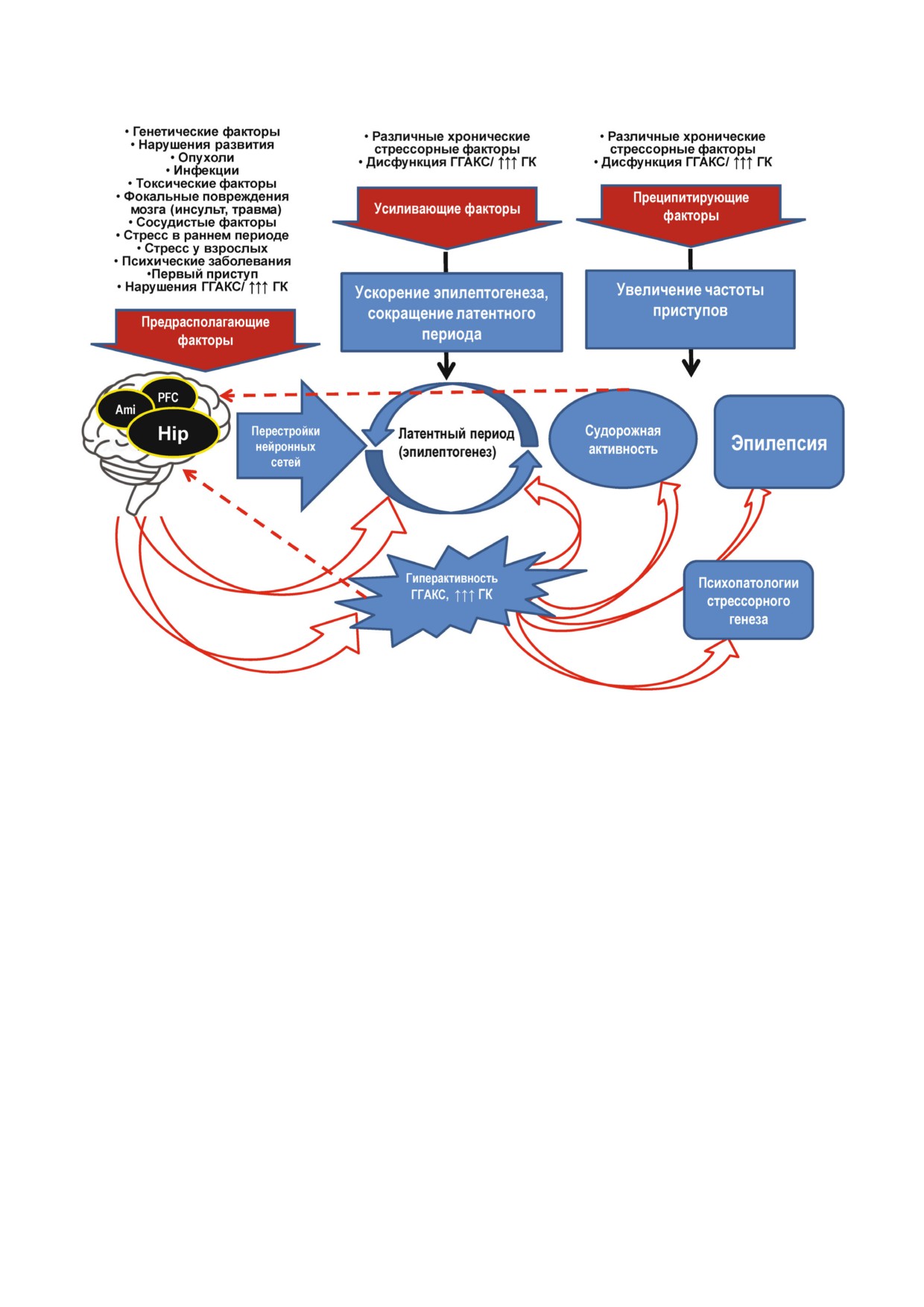
Рис. 1. Общие факторы развития эпилепсии и коморбидной психопатологии. Указаны группы факторов, предрасполага
ющих и усиливающих эпилептогенез, а также преципитирующих судорожные приступы. Схема составлена на основании
данных, приведенных в работах [27, 29, 37, 40]. ГК - глюкокортикоиды; ГГАКС - гипоталамо гипофизарно адренокор
тикальная система; Ami - амигдала; Hip - гиппокамп; PFC - префронтальная кора. (С цветными вариантами рисунков
ческих стрессорных факторов, психологических
При неконтролируемом хроническом стрес
и физических. По сути, коморбидные эпилеп
се гиперкортизолемия (в экспериментах на жи
сии, депрессивные и тревожные расстройства и
вотных - гиперкортикостеронемия) становится
подтверждают хронизацию неизбегаемого стрес
важнейшим компонентом при формировании
са. Представление об эпилепсии как модели хро
порочного круга коморбидности эпилепсии и
нического стресса схематически отражено на ри
депрессии, в основе которого лежат стрессор
сунке 1, где приведены стрессорные проэпилеп
ные и нейрогуморальные факторы [41]. След
тические факторы, действующие в разные пери
ствием развивающихся на фоне дисфункции
оды развития эпилепсии. Также показана их по
ГГАКС коморбидных депрессии и ЭВД стано
тенциальная связь с развитием психопатоло
вится усиление действия взаимно потенцирую
гии: указаны группы предрасполагающих, уси
щих хронических стрессорных факторов, в ка
ливающих эпилептогенез и преципитирующих
честве которых начинают выступать и сами за
судорожные приступы факторов. При этом ус
болевания, в частности осознание пациентом
тойчивые нарушения ГГАКС представляются
своего состояния. В основе такого порочного
ключевыми в развитии коморбидных эпилепсии
круга лежит тот факт, что ГГАКС контролирует
и депрессии. Вначале это избыточная активация
как стрессорный ответ, так и судорожную актив
системы, затем, на стадии хронической депрес
ность, потому что гормоны стресса, в первую
сии, наступает истощение ГГАКС [39].
очередь ГК, влияют и на возбудимость нейро
БИОХИМИЯ том 86 вып. 6 2021
792
ГУЛЯЕВА
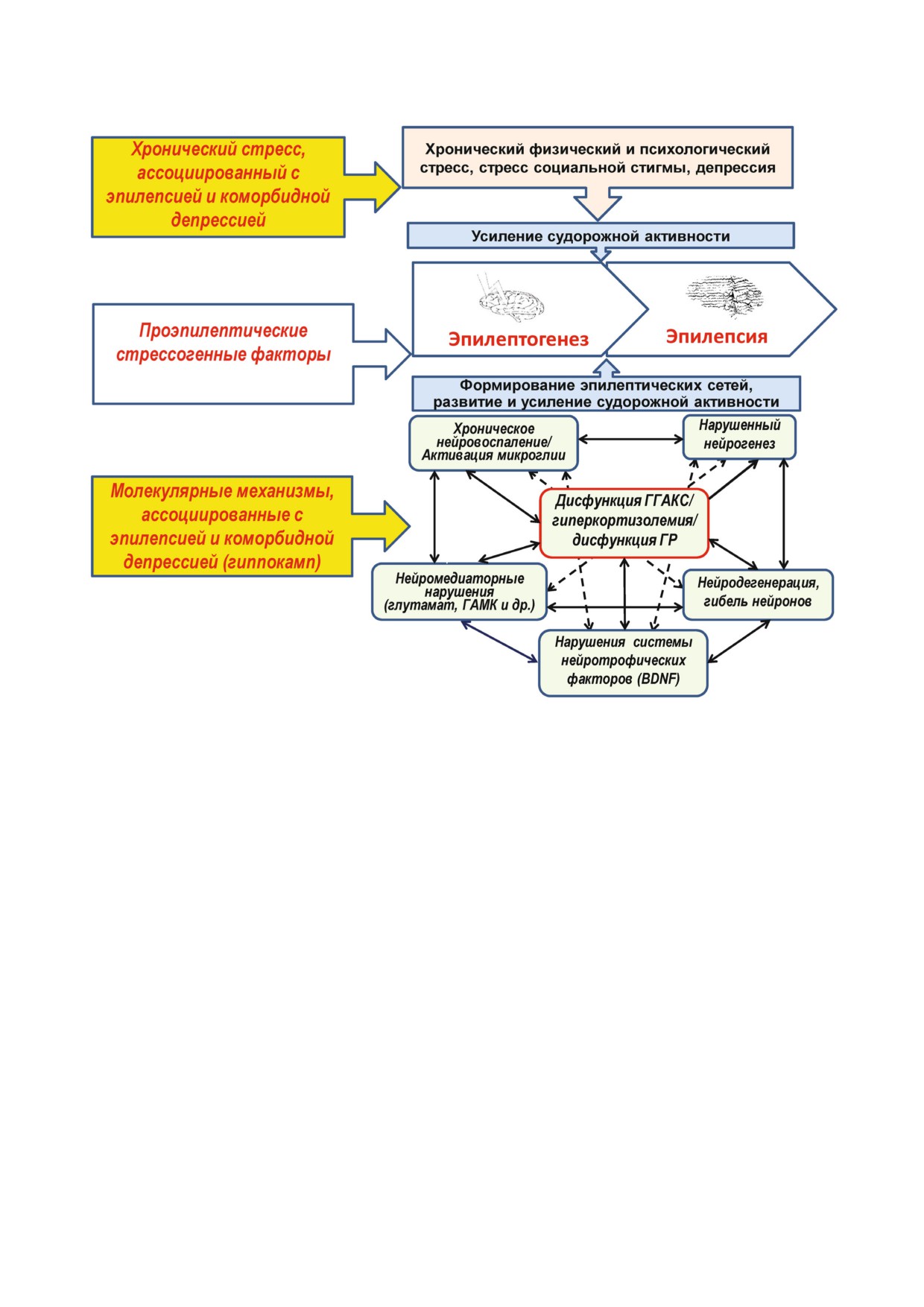
Рис. 2. Порочный круг коморбидности эпилепсии и депрессии, в основе которого лежат стрессорные и нейрогумораль
ные факторы. Следствия развивающихся на фоне дисфункции ГГАКС коморбидных депрессии и ЭВД представляют со
бой взаимно усиливающие коморбидную патологию стрессорные факторы, образующие порочный круг и способствую
щие развитию и усугублению обоих заболеваний. Хронический стресс (в т.ч. ассоциированный с эпилепсией) приводит к
дисфункции ГГАКС, что в свою очередь вызывает нарушение стресс реактивности, углубляя эту дисфункцию, результа
том которой является развитие депрессивного расстройства. Развитие депрессии, в свою очередь, усиливает дисфункцию
ГГАКС, выступая в качестве дополнительного хронического стрессорного фактора для пациента. При этом стресс посред
ством механизмов разного уровня приводит к запуску судорожных приступов и увеличению их частоты. Утяжеление эпи
лепсии также ассоциировано с усилением хронического стресса и коморбидной депрессии
нов, и на их чувствительность к судорожной ак
следить стадийность изменений нейропластич
тивности [42, 43] (рис. 2).
ности. Основными фазами, противоположными
по направлению, являются активный эпилепто
генез и стадия сформировавшейся ЭВД [44-46].
ГИППОКАМПАЛЬНЫЕ МОЛЕКУЛЯРНО
В период эпилептогенеза происходят события,
КЛЕТОЧНЫЕ МЕХАНИЗМЫ РАЗВИТИЯ
приводящие к формированию эпилептических
ЭВД, ЛЕЖАЩИЕ В ОСНОВЕ
нейронных сетей и соответствующей коннекто
КОМОРБИДНОСТИ ЭПИЛЕПСИИ
патии. Это изменения глутаматергических и
И ДЕПРЕССИИ
ГАМКергических нейронов (рис. 3), усиление
аберрантного нейрогенеза, аксональный спру
Стадийность изменений нейропластичности в
тинг и ремоделирование дендритов, в частности
гиппокампе при ЭВД. На примере ЭВД, наибо
за счет избыточного усиления системы нейро
лее распространенной и изученной формы эпи
трофического фактора мозга (BDNF, brain derived
лепсии у взрослых, затрагивающей структуру и
neurotrophic factor) в специфических областях
функционирование гиппокампа, можно про
гиппокампа. По мере развития эпилептогенеза
БИОХИМИЯ том 86 вып. 6 2021
СТРЕСС, ГИППОКАМП, ЭПИЛЕПСИЯ И КОМОРБИДНАЯ ДЕПРЕССИЯ
793
эта стадия аберрантной суперпластичности сме
также и при депрессивных расстройствах, ко
няется формированием эпилептогенного очага,
торые, как правило, также связаны с дисфунк
функционирование которого сопровождается
цией ГГАКС. Другие общие механизмы, вклю
противоположными событиями, приводящими
чая истощение BDNF, активацию микроглии,
к ограничению пластичности: истощением сис
нейровоспаление, нарушение нейрогенеза и
темы BDNF, нейровоспалением, повреждением
нейродегенерацию [40, 48], изменения в ней
аксонов, нейродегенерацией, гибелью нейро
ромедиаторных системах (глутамат , серото
нов, склерозом гиппокампа, подавлением суб
нин , моноамин и ГАМКергической) [49],
гранулярного нейрогенеза (рис. 3).
часто также связаны с ГГАКС прямо или опос
Центральная роль ГГАКС в коморбидности
редованно.
эпилепсии и депрессии. Перестройка синапти
Исследования последних лет подтверждают
ческих связей и нейронных сетей в процессе
наличие гиперактивной ГГАКС и повышенного
эпилептогенеза в значительной степени ини
уровня ГК у пациентов с ЭВД. Избыток ГК сти
циируется нарушениями ГГАКС. Контроль
мулирует возбудимость нейронов, эпилептиформ
большинства ключевых механизмов, опосреду
ную активность и предрасположенность к при
ющих эпилептогенез и формирование зрелых
падкам. Пациенты с ЭВД могут проявлять ано
эпилептических сетей при ЭВД, нарушается
мальные реакции на стресс, облегчающие раз
Рис. 3. Схематическое представление молекулярно клеточных гиппокампальных механизмов основных стадий патогене
за ЭВД [44-47]. По мере развития ЭВД в процессе эпилептогенеза на фоне избытка BDNF и усиления нейрогенеза в суб
гранулярной зоне зубчатой извилины формируется «аберрантная суперпластичность» и происходит созревание эпилеп
тических нейронных сетей. Затем на фоне дисфункции ГГАКС (избытка ГК и нарушения функционирования глюкокор
тикоидных и минералокортикоидных рецепторов) «гиперглутаматергический» сигналинг и активация нейровоспаления
приводят к подавлению нейрогенеза, нейродегенерации и гибели нейронов, что существенно снижает пластичность гип
покампа; этот процесс усугубляется в результате рекуррентной судорожной активности
БИОХИМИЯ том 86 вып. 6 2021
794
ГУЛЯЕВА
витие приступа и повышающие вероятность ко
В гиппокампе пациентов с фармакорезистент
морбидных психопатологий [37, 50]. Гиперак
ной ЭВД и коморбидной депрессией обнаруже
тивность ГГАКС отрицательно влияет на струк
но снижение плотности ГР [55]. Экспрессию ГР
туру и функцию височной доли мозга, значи
оценивали иммуногистохимически у пациентов
тельно вовлеченной и в ЭВД, и в расстройства
с ЭВД, перенесших нейрохирургическую опера
настроения. ГК через свои рецепторы на мемб
цию по поводу фармакорезистентной эпилеп
ранах и в цитоплазме глутаматергических си
сии, а также у контрольной группы (аутопсия).
напсов могут нарушать глутаматергическую
В контрольных образцах выявлена высокая
трансмиссию (подробнее в обзоре [51]). Избы
плотность ГР в гранулярных нейронах зубчатой
точная возбуждающая активность нейронов,
извилины и пирамидных клетках CA1, в то вре
сопровождающая судорожные приступы, пов
мя как экспрессия ГР у пациентов с ЭВД была
реждает структуры височных долей, еще больше
снижена, особенно при наличии депрессии. Со
растормаживая ГГАКС [43]. Гиппокамп особен
отношение между числом клеток ГР+/NeuN+ и
но чувствителен к хроническому стрессу и
экспрессия ГР в глиальных клетках были ниже у
действию ГК [41]. Эти факторы вызывают, в
пациентов с коморбидными ЭВД и депрессией в
частности, ретракцию дендритов - обратимую
сравнении с ЭВД и отрицательно коррелирова
форму пластичности, которая включает ре
ли с тяжестью депрессии, оцененной по психи
структуризацию дендритов. При ретракции
атрическому анамнезу. Таким образом, сниже
дендритов, которая может сохраняться в тече
ние экспрессии ГР в гиппокампе может быть
ние недель, месяцев или даже лет, вероятность
вовлечено в этиологию депрессии у пациентов с
повреждения гиппокампа увеличивается. Нали
ЭВД [55].
чие такого «временнóго окна» является одним
Клинические данные свидетельствуют о том,
из постулатов «гипотезы уязвимости гиппокам
что усиленная нейроэндокринная реакция на
па для ГК» [52].
стресс у пациентов с ЭВД имеет хронический
Дисфункция ГГАКС при эпилепсии требует
характер, потенциально способствуя как прог
более подробного механистического изучения,
рессированию заболевания, так и развитию со
потому что именно она является ключом к по
путствующей психопатологии. Не вполне ясно,
ниманию развития и прогрессирования ЭВД и
отражают нарушения ГГАКС прямые эффекты
коморбидных патологий, а также потенциаль
эпилептической патологии или вторичные эф
ной мишенью для терапии. Мета анализ соот
фекты бремени болезни, в частности социаль
ношения уровня кортизола и судорог у пациен
ное отчуждение, стигматизацию и другие типы
тов с эпилепсией был проведен в работе Cano
хронического стресса, сопутствующего эпилеп
López et al. [32] по данным 38 статей, в 14 из ко
сии. Моделирование на животных дает возмож
торых был проведен анализ базального уровня
ность разделить эти факторы для оценки вос
кортизола, в 8 - исследовалось влияние проти
произведения нейроэндокринных и поведен
воэпилептических препаратов, в 13 - оценива
ческих изменений, характерных для эпилепсии
лись эффекты судорог на уровень кортизола, в
человека. В работе Wulsin et al. [56] на самцах
3 - влияние стресса на этот показатель. Базаль
мышей линии FVB моделировали ЭВД при по
ный кортизол у пациентов оказался выше по
мощи пилокарпина, который вызывал эпилеп
сравнению с популяцией в исследованиях с на
тический статус и последующее развитие спон
иболее гомогенными группами (45% из 38).
танных рецидивирующих припадков. Базальные
Несмотря на гетерогенность данных по влия
уровни кортикостерона были повышены у мы
нию противоэпилептических препаратов, судо
шей, получавших пилокарпин, через 1, 7 и
роги были ассоциированы с повышением уров
10 недель после эпистатуса по сравнению с
ня кортизола в 77% исследований. Единствен
контрольной группой. Мыши с эпилепсией
ная работа с предъявлением острого стресса по
имели увеличенный вес надпочечников и ги
казала более высокую реактивность кортизола
персекрецию кортикостерона после острого им
при эпилепсии в сравнении с контролем [32]. В
мобилизационного стресса. Животные в модели
исследованиях с самооценкой стресса пациен
ЭВД демонстрировали усиленное ангедоничес
тами более высокая частота приступов была ас
кое поведение (показатель депрессивно подоб
социирована с повышенными уровнями корти
ного фенотипа), гиперактивность и тревож
зола и сниженной функциональной коннектив
ность. Таким образом, при моделировании ЭВД
ностью мозга. Суточный ритм эпилептических
на мышах развивается гиперактивность ГГАКС
припадков при разных типах эпилепсии сходен
и депрессивно подобное поведение. Эпилепти
с циркадианным ритмом кортизола, и эти ре
ческий статус индуцирует быструю гиперакти
зультаты подтверждают гипотезу зависимости
вацию ГГАКС, а дисфункция этой оси может
припадков от уровня ГК [53, 54].
усугублять вызванное судорожным приступом
БИОХИМИЯ том 86 вып. 6 2021
СТРЕСС, ГИППОКАМП, ЭПИЛЕПСИЯ И КОМОРБИДНАЯ ДЕПРЕССИЯ
795
повреждение головного мозга. На той же модели
торможения в гиппокампе и экстрагиппокам
ЭВД было показано, что введение животным
пальных областях [70]. Такие изменения также
антагониста ГР RU486 блокирует вызванное пи
сопровождаются нарушениями обучения, памя
локарпин индуцированным эпистатусом повы
ти и депрессией [71, 72]. Это не удивительно, по
шение уровня кортизола [57]. RU486 также пре
тому что нормальный нейрогенез важен как для
пятствовал развитию патологических измене
обучения/памяти, так и для поддержания нор
ний гиппокампа, включая потерю мшистых
мального эмоционального состояния [73]. По
клеток хилуса и патологическую пролиферацию
давление нейрогенеза в гиппокампе считают од
клеток в зубчатой фасции. Интересно, что поте
ним из ключевых механизмов развития депрес
ря мшистых клеток и накопление эктопических
сивных расстройств [74-77], поэтому снижение
новообразованных клеток в хилусе положитель
нейрогенеза при ЭВД потенциально является
но коррелировали с тяжестью судорожных
основой для коморбидности с депрессией. Для
приступов, что позволяет предположить суще
пациентов с фармакорезистентной ЭВД и ко
ствование потенциального антиэпилептогенно
морбидной депрессией, перенесших хирурги
го эффекта антагонистов ГК [57]. С использова
ческую операцию, характерен другой паттерн
нием другой модели ЭВД, пентилентетразоло
потери экспрессирующих кальбиндин нейронов
вого киндлинга (процесса, при котором повтор
в зубчатой извилине в сочетании с более высо
ные введения подпороговых доз конвульсанта
ким спрутингом аксонов этих нейронов по срав
приводили к развитию судорожной активнос
нению с пациентами с ЭВД [78]. Эти изменения
ти), мы ранее показали, что даже однократный
могут свидетельствовать о нарушениях созрева
судорожный приступ вызывает постепенное
ния гранулярных клеток, связанных с феноме
ухудшение функций обучения и памяти в гип
ном эпилептической гипервозбудимости.
покамп зависимых тестах [58]. По видимому,
Нейровоспаление. За последние годы было
кортикоид зависимый механизм повреждения
сформировано и подтверждено многочислен
гиппокампа может лежать и в основе когнитив
ными экспериментальными данными представ
ных нарушений, описанных у пациентов с ЭВД.
ление о ключевой роли процессов нейровоспа
Изменения нейрогенеза. Практически у всех
ления при эпилепсии, в т.ч. в гиппокампе при
млекопитающих, включая человека, на протя
ЭВД [79-83]. Данные другого пула исследова
жении всей жизни в гиппокампе реализуется
ний убедительно свидетельствуют о том, что
нейрогенез. Многочисленные исследования на
нейровоспаление в гиппокампе играет важную
животных моделях показывают, что нейрогенез
роль и в патогенезе депрессивных расстройств
в субгранулярной нише (зубчатой извилине гип
[84, 85]. Современное представление о роли ци
покампа) важен для таких функций мозга, как
токинов как нейромодуляторов в мозге при эпи
обучение, память и эмоции, при этом вновь об
лепсии и депрессии складывается из результатов
разованные нейроны потенциально более возбу
исследований с участием людей и эксперимен
димы, чем «старые». Нейрогенез в гиппокампе
тальных животных [86, 87]. Цитокиновые ре
взрослого мозга очень чувствителен к физиоло
цепторы присутствуют в нейронах и популяциях
гическим и патологическим раздражителям.
глиальных клеток в определенных областях моз
При этом ключевым механизмом влияния
га. В гиппокампе цитокины являются неотъем
стрессорных событий на нейрогенез является
лемыми участниками физиологических процес
его модуляция посредством ГК [59, 60]. Судо
сов, таких как память и обучение; они также
рожные приступы влияют на нейрогенез как ко
вовлечены в патологические состояния, такие
личественно, так и качественно [61]. Острые
как депрессия и эпилепсия. В норме интерлей
приступы, как правило, ассоциируются с усиле
кин 1β необходим для поддержания долговре
нием нейрогенеза и миграцией новорожденных
менной потенциации в гиппокампе, а интерлей
нейронов в эктопические области, такие как хи
кин 6 играет регуляторную роль в формирова
лус и молекулярный слой зубчатой извилины.
нии долговременной памяти, оказывая негатив
Аномально (эктопически) мигрирующие ново
ный эффект при избыточной концентрации.
рожденные нейроны, формирующиеся в резуль
Секретируемый астроцитами фактор некроза
тате острых приступов, эпилептического статуса
опухоли увеличивает поверхностную транслока
или черепно мозговой травмы, принимают
цию глутаматных АМРА рецепторов в глутама
участие в создании эпилептогенных нейронных
тергических синапсах, а хемокин CXCL12 может
сетей гиппокампа [62-69]. Напротив, рецидиви
подавлять тоническую активность нейронов Ка
рующие спонтанные припадки при хронической
халя-Ретциуса в гиппокампе. При ЭВД отмече
ЭВД ассоциированы со значительно снижен
но значительное повышение концентрации ин
ным нейрогенезом, аномальной реорганизацией
терлейкина 10, интерферона γ, ICAM1, CCL2 и
нейронных сетей, потерей функционального
CCL4 в гиппокампе больных [88].
БИОХИМИЯ том 86 вып. 6 2021
796
ГУЛЯЕВА
Глиальные клетки играют важнейшую роль в
лептогенез, так и в фармакорезистентность и
патогенезе эпилепсии, модулируя как воспали
психиатрическую коморбидность у пациентов с
тельные процессы (в первую очередь микрог
ЭВД [107, 108]. Эпилепсия и расстройства наст
лия, но также и астроциты), так и возбудимость
роения имеют также общие рецепторные и
нейронов (астроциты). Механизмы вовлечен
трансдукционные механизмы систем серотони
ности глиальных клеток в эти процессы подроб
на, дофамина и γ аминомасляной кислоты
но рассмотрены в обзоре [89]. Потеря нейронов
(ГАМК) [109].
гиппокампа, индуцированная эксайтотоксич
Система BDNF. Роль системы BDNF в гип
ностью и нейровоспалением, а также реактив
покампе подробно изучена как в эпилептогене
ный астроглиоз являются патологическими
зе при ЭВД ([110, 111], рис. 3), так и в развитии
признаками ЭВД. Функциональные изменения
депрессивных расстройств [112, 113]. В обоих
астроцитов гиппокампа у больных с ЭВД ассо
случаях недостаточность BDNF ассоциирована
циированы с психическими расстройствами и
со снижением нейрогенеза и усилением нейро
экспрессией связанных с астроглиозом генов
воспалительных процессов в гиппокампе. Сис
глиального фибриллярного кислого белка, ме
тема BDNF тесно связана с функционировани
таллотионеина I/II и аквапорина 4 [90].
ем нейронов различного типа, потенциально
Глутаматергическая система. Изменения глу
важных для коморбидности эпилепсии и деп
таматергической системы, основной системы
рессии; в первую очередь она осуществляет
возбуждающей нейротрансмиссии височной
мультифакторную реципрокную регуляцию с
доли, описаны как при эпилепсии, при ко
глутаматергической системой [114].
торой они являются ключевыми в развитии из
Общность молекулярных механизмов стрес
быточного возбуждения [91-96], так и при деп
сорного ответа и эпилептогенеза. Принципиаль
рессии [97-101]. Повышенная глутаматерги
но важно еще раз подчеркнуть, что все упомяну
ческая нейротрансмиссия (в последнее время
тые выше основные молекулярные механизмы,
часто используется термин «гиперглутаматерги
вовлеченные в повреждение гиппокампа при
ческая нейротрансмиссия») в гиппокампе, как
ответе на хронический стресс, ассоциирован
центральный фактор патогенеза эпилепсии,
ный с эпилептогенезом/эпилепсией и развити
депрессии и когнитивных нарушений [101], ин
ем коморбидного депрессивного расстройства,
дуцирует нейродегенеративные изменения и
являются взаимозависимыми. При этом цент
гибель нейронов в гиппокампе. Аберрантная
ральным интегрирующим не только на уровне
сигнализация затрагивает как ионотропные
мозга, но и целого организма, и модулирующим
[102, 103], так и метаботропные рецепторы
звеном является ГГАКС, нарушения которой
[104] и индуцирует развитие эксайтотоксичнос
проявляются в избытке ГК и в дисфункции ГР
ти и окислительного стресса, которые также
(рис. 4).
вносят вклад в коморбидность ЭВД и психиат
В соответствии со схемой, представленной
рических заболеваний [93, 105]. Важно отме
на рисунке 4, принципиально важно, что моле
тить, что к настоящему времени известны мно
кулярные механизмы стрессорного ответа и
гочисленные пути, по которым ГК регулируют
эпилептогенеза являются общими, и это обес
функционирование глутаматергических синап
печивает коморбидность эпилепсии и других
сов, на мембранах и в цитоплазме которых
индуцируемых стрессом заболеваний, в част
экспрессированы минералокортикоидные и
ности депрессивных/тревожных расстройств. В
глюкокортикоидные рецепторы [51]. Исследо
центре сложной системы взаимосвязанных мо
вания роли глутаматных рецепторов в комор
лекулярных процессов находится дисфункция
бидности эпилепсии и депрессии на животных
ГГАКС, связанная с нарушением баланса кор
моделях только начинаются. Например, на мо
тикостероидных рецепторов и приводящая к ги
дели флуоксетин резистентной эпилепсии, ас
перкортизолемии, определяющей второй важ
социированной с депрессивно подобным пове
нейший узел схемы. Системы возбуждающего
дением, было показано, что важным фактором
нейромедиатора глутамата и нейротрофина
развития коморбидной патологии является хро
BDNF принципиально важны для феноменов
ническое нарушение сигнальной трансдукции
клеточной и синаптической пластичности. Они
через NMDA рецептор, в частности сигнально
связаны многочисленными реципрокными свя
го каскада ERK/CREB/ELK1/BDNF/cFOS и
зями, обеспечивающими взаимную регуляцию
фосфорилирования CREB (cyclic AMP response
этих систем [114]. Именно сложная, но хорошо
element binding protein) [106]. Интересно отме
скоординированная природа этих взаимоотно
тить, что глутаматные синапсы содержат и кан
шений обеспечивает оптимальную клеточную и
набиноидные рецепторы, а нарушения эндо
синаптическую пластичность мозга в норме.
каннабиноидной системы вовлечены как в эпи
Обе системы связаны с патогенезом аффектив
БИОХИМИЯ том 86 вып. 6 2021
СТРЕСС, ГИППОКАМП, ЭПИЛЕПСИЯ И КОМОРБИДНАЯ ДЕПРЕССИЯ
797
Рис. 4. Основные молекулярные механизмы, вовлеченные в повреждение гиппокампа при ответе на хронический стресс,
ассоциированный с эпилептогенезом, эпилепсией и развитием коморбидного депрессивного расстройства. Центральным
интегрирующим и модулирующим звеном этих взаимосвязанных механизмов является ГГАКС, нарушения которой про
являются в избытке ГК и дисфункции ГР. Подробные объяснения в тексте статьи
ных расстройств, и нарушение тесных и сбалан
Функциональные и структурные повреждения
сированных взаимоотношений между ними вы
гиппокампа, селективно уязвимого при
зывает неблагоприятные изменения нейроплас
действии внешних факторов и отвечающего на
тичности, лежащие в основе судорожной актив
них повышением секреции цитокинов [115],
ности, депрессии, тревожности и других психи
составляют индивидуальную основу для разви
ческих заболеваний. Нейровоспаление, реали
тия как эпилепсии, так и депрессивно тревож
зуемое в ответ на дисфункцию ГГАКС в основ
ных расстройств.
ном активированной микроглией, также тесно
связано с упомянутыми выше системами и ин
дуцирует нейродегенерацию и гибель нейронов
ИЗМЕНЕНИЯ НЕЙРОННЫХ СЕТЕЙ,
лимбической системы мозга, а также подавле
ЛЕЖАЩИЕ В ОСНОВЕ КОМОРБИДНОСТИ
ние нейрогенеза. Избыток кортикостероидов,
ЭПИЛЕПСИИ И ДЕПРЕССИИ
секретированных в условиях ЭВД, особенно у
пациентов с исходно аномальным стрессорным
Описанные выше механизмы патогенеза
ответом в связи с дисфункцией ГГАКС, взаимо
эпилепсии на уровне молекул и клеток имеют
действует с ГР гиппокампа, индуцируя сигналь
место и при депрессивных расстройствах. Сос
ные пути, стимулирующие нейровоспаление и
тавляя основу коморбидности этих заболева
последующие события, включая нарушения
ний, они несомненно должны формировать ба
нейрогенеза и нейродегенерацию [21, 41, 60].
зу и для общих механизмов на сетевом и систем
БИОХИМИЯ том 86 вып. 6 2021
798
ГУЛЯЕВА
ном уровнях. Эпилепсия характеризуется как
Интересно, что нейротизм, основная черта
расстройство, связанное с повторяющимися
личности, характеризующаяся склонностью к
припадками, которые определяются как «прехо
переживанию негативных аффектов, выше у
дящее возникновение признаков и/или симпто
людей с ЭВД по сравнению с популяционным
мов вследствие аномальной чрезмерной или
уровнем. Нейротизм является известным пре
синхронной активности нейронов в головном
диктором депрессии и тревоги, которые также
мозге» [116]. Такая гиперактивность является
чаще встречаются у людей с ЭВД. По данным
результатом патологической активности в боль
работы Rivera Bonet et al. [120], при повышен
ших группах или сетях нейронов. Организация и
ном нейротизме у пациентов с депрессией и тре
потенциальные механизмы регуляции аберрант
вожными симптомами существуют как уникаль
ных нейронных сетей при ЭВД подробно рас
ные, так и общие паттерны аномальной функ
сматриваются в работе Alexander et al. [116] как
циональной коннективности сетей покоя между
на уровне «микросхем», т.е. отношений между
областями мезиальной височной и лобной до
отдельными нейронами или небольшими груп
лей. Например, повышенный нейротизм досто
пами нейронов, так и на уровне макросистем,
верно ассоциирован с гипосинхронией между
охватывающих большие области мозга. ЭВД -
правым гиппокампом и областью Бродмана - и
хорошая биологическая модель для установле
эта уникальная связь не зависит от симптомов
ния общей структурной основы между депрес
депрессии и тревоги.
сией и эпилепсией. Эти заболевания имеют
сходные свойства нейронных сетей, включаю
щих височные доли с гиппокампом, миндали
ЗАКЛЮЧЕНИЕ: ГИППОКАМП
ной и энторинальной корой и неокортексом;
КАК КОНТРАПУНКТ СТРЕССА,
лобные доли с поясной извилиной; подкорко
ЭПИЛЕПСИИ И ДЕПРЕССИИ
вые структуры, такие как базальные ганглии и
таламус, а также соединительные пути [117].
Эпилепсия - это неврологическое состоя
Иными словами, существуют клинические и
ние, связанное с судорогами, нейродегенераци
структурные доказательства того, что депрессия
ей, перестройкой нейронных сетей и другими
и ЭВД представляют собой эпифеномен, в осно
структурными и функциональными нарушени
ве которого лежат сходные свойства нейронных
ями. Последние могут приводить как к измене
сетей.
нию поведения и эмоционального состояния
Обычно при рассмотрении коморбидности
(тревога, депрессия), так и к когнитивным дис
заболеваний возникает неоднозначно решае
функциям (неспособность к обучению, ухудше
мый вопрос о том, какое из них первично. Счи
ние памяти) и ускоренному старению. Все боль
тают, что депрессия и тревожность при эпилеп
ше исследователей отмечает общие механизмы
сии отражают дисфункцию широко распреде
развития нейродегенеративных заболеваний,
ленных лимбических сетей, которая может быть
депрессивных расстройств и эпилепсии
причиной или следствием эпилептогенеза. При
[21, 121]. При этом когнитивные нарушения и
рефрактерной эпилепсии как судороги, так и
эмоционально личностные расстройства рас
некоторые противосудорожные препараты мо
сматривают как симптомы, а гиппокамп - как
гут нарушать функционирование центральных
принципиальную область поражения.
гомеостатических механизмов, лежащих в осно
К настоящему времени накоплен значитель
ве эмоций, и повышать риск последующих
ный пул клинических и экспериментальных
приступов [118]. При ЭВД нейронные сети, свя
данных, которые доказательно связывают ЭВД с
занные со страхом, ассоциированы с симптома
депрессией. При изучении механизмов патоге
ми тревоги и иктальной паники [6]; многие ис
неза этих коморбидных заболеваний использу
следователи рассматривают изменения в ней
ется множество подходов от ультраструктурного
ронных сетях при ЭВД как основу и для психи
уровня, например, анализа экспрессии клеточ
ческих расстройств [5]. Тем не менее обзор ней
ных рецепторов, до поведенческого уровня,
ровизуализационных данных взрослых пациен
например, исследования коморбидной депрес
тов с эпилепсией и депрессией показал их про
сии у животных при моделировании эпилепсии.
тиворечивость. В частности, некоторые авторы
Изучение нейробиологических аспектов
предполагают, что депрессия (по крайней мере
расстройств настроения при ЭВД, первичных
при ЭВД) связана с другим паттерном измене
расстройств настроения и патогенетических ме
ний мозга по сравнению с большой депрессией,
ханизмов эпилепсии, полученных в исследова
и это подтверждает представление ряда авторов
ниях с участием людей и на животных моделях,
о феноменологических особенностях депрессии
позволяет лучше понять сложную взаимосвязь
при эпилепсии [119].
между этими состояниями, а комплексные ис
БИОХИМИЯ том 86 вып. 6 2021
СТРЕСС, ГИППОКАМП, ЭПИЛЕПСИЯ И КОМОРБИДНАЯ ДЕПРЕССИЯ
799
следования позволяют выявить формирующую
астроциты вовлечены в различные процессы,
ся картину общих патофизиологических меха
играя роль, которая выходит далеко за рамки их
низмов.
участия в склерозе гиппокампа. Все эти меха
Обосновывая концепцию о континууме
низмы тесно переплетены и, вероятно, двунап
нейропластичности и нейропатологии [47], мы
равлены, так что структурные и функциональ
подчеркивали общность и плейотропность
ные изменения от одного заболевания увеличи
(вовлеченность как в реализацию нормальной
вают риск развития другого. Контрапунктом
адаптивной пластичности, так и в патогенез за
депрессии и эпилепсии является лимбическая
болевания мозга) механизмов на молекуляр
система, в первую очередь гиппокамп.
ном, синаптическом, клеточном и сетевом
Понимание общих нейробиологических ме
уровнях. Эти механизмы ассоциированы с вы
ханизмов патогенеза, ответственных за комор
сокой адаптивной пластичностью гиппокампа,
бидность эпилепсии и депрессии, имеет высо
ответственного за интегративную функцию
кую трансляционную значимость и потенциаль
мозга, в частности обучение, память и эмоции.
ное применение в клинической практике. Су
Однако ценой высокой пластичности является
ществование общих механизмов может влиять
селективная чувствительность гиппокампа к
на течение эпилепсии и реакцию на фармаколо
развитию патологических процессов. Сигнали
гическое и хирургическое лечение судорожных
зация путем секреции малых молекул характер
расстройств у пациентов с депрессией в анамне
на как для нервной, так и для эндокринной и
зе. Индивидуальная стресс реактивность паци
иммунной систем. Масштабы и значение взаи
ента является фактором, принципиально значи
модействия между этими системами организма
мым для развития эпилепсии и коморбидных
стали объектами научного исследования только
психических расстройств. К сожалению, доста
в последнее время. Нейромедиаторы, гормоны
точно простые и эффективные подходы к оцен
и цитокины воздействуют на клетки мишени
ке индивидуальной стресс реактивности, осно
через мембранные или внутриклеточные рецеп
ванные на уровнях маркеров стресса, не приме
торы, включающие каскадные молекулярные
няются у пациентов с эпилепсией. Маркеры мо
механизмы. Одной из ключевых центральных
гут быть проанализированы как в крови, так и
структур, регулирующих взаимодействие между
неинвазивно (в волосах, слюне и т.д.) [39]; ин
этими системами, несомненно является гиппо
формативность этих данных повышается при
камп.
использовании легкого стрессорного тестового
Появляется все больше доказательств того,
воздействия [123]. Представление о роли инди
что при эпилепсии и депрессии изменения во
видуальной стресс реактивности в развитии
взаимодействии серотонин и норадренергичес
эпилепсии и коморбидных заболеваний может
ких нейронов с глутаматергическими системами
быть экспериментально подтверждено и ис
связано с аномальными нейронными сетями и
пользовано в практической медицине для разра
повышенной возбудимостью. Гипервозбуди
ботки новых персонализированных стратегий
мость нейронов может вызывать судорожную
коррекции функционального состояния орга
активность, а также тревожные эмоции. Кроме
низма и контроля эффективности лечения.
того, изменение синаптических концентраций
нейромедиаторов и высокий уровень ГК влияют
на внутриклеточные сигнальные пути, вызывая
Финансирование. Работа выполнена при фи
нарушения системы нейротрофинов и других
нансовой поддержке Российского фонда фунда
нейротрофических факторов [122]. Коморбид
ментальных
исследований
(грант
ные механизмы могут быть ассоциированы с ат
№ 20 115 50019 Экспансия).
рофией гиппокампа, наблюдаемой при магнит
Конфликт интересов. Автор заявляет об отсут
но резонансной томографии, и нарушением па
ствии конфликта интересов.
мяти. Исследования с использованием позит
Соблюдение этических норм. Настоящая
ронно эмиссионной томографии дополнитель
статья не содержит описания выполненных ав
но указывают на гипометаболизм глюкозы в ви
тором экспериментальных исследований с учас
сочных и лобных долях. При ЭВД и депрессии
тием людей или животных в качестве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Gulyaeva, N. V. (2017) Molecular mechanisms of neuro
2.
Kanner, A. M. (2017) Psychiatric comorbidities in new
plasticity: an expanding universe, Biochemistry (Moscow),
onset epilepsy: should they be always investigated? Seizure,
82, 237 242, doi: 10.1134/S0006297917030014.
49, 79 82, doi: 10.1016/j.seizure.2017.04.007.
БИОХИМИЯ том 86 вып. 6 2021
800
ГУЛЯЕВА
3.
Kanner, A. M. (2016) Psychiatric comorbidities in epilep
17.
Larrieu, T., and Sandi, C. (2018) Stress induced depres
sy: should they be considered in the classification of epilep
sion: is social rank a predictive risk factor? Bioessays, 40,
tic disorders? Epilepsy Behav., 64 (Pt B), 306308,
e1800012, doi: 10.1002/bies.201800012.
doi: 10.1016/j.yebeh.2016.06.040.
18.
Slavich, G. M., and Sacher, J. (2019) Stress, sex hormones,
4.
Ribot, R., and Kanner, A. M. (2019) Neurobiologic prop
inflammation, and major depressive disorder: extending
erties of mood disorders may have an impact on epilepsy:
social signal transduction theory of depression to account
should this motivate neurologists to screen for this psychi
for sex differences in mood disorders, Psychopharmacology
atric comorbidity in these patients? Epilepsy Behav., 98 (Pt
(Berl),
236,
30633079, doi:
10.1007/s00213 019
B), 298 301, doi: 10.1016/j.yebeh.2019.01.026.
05326 9.
5.
Sair, A., Şair, Y. B., Saracoğlu, İ., Sevincok, L., and Akyol,
19.
Sotiropoulos, I., Silva, J. M., Gomes, P., Sousa, N., and
A. (2021) The relation of major depression, OCD, person
Almeida, O. F. X. (2019) Stress and the etiopathogenesis of
ality disorders and affective temperaments with temporal
Alzheimer’s disease and depression, Adv. Exp. Med. Biol.,
lobe epilepsy, Epilepsy Res., 171, 106565, doi: 10.1016/
1184, 241 257, doi: 10.1007/978 981 32 9358 8_20.
j.eplepsyres.2021.106565.
20.
LeMoult, J., Humphreys, K. L., Tracy, A., Hoffmeister,
6.
Melo, H. M., Guarnieri, R., Vascouto, H. D., Formolo,
J. A., Ip, E., and Gotlib, I. H. (2020) Meta analysis: expo
D. A., de Carvalho, C. R., et al. (2020) Ictal fear is associ
sure to early life stress and risk for depression in childhood
ated with anxiety symptoms and interictal dysphoric disor
and adolescence, J. Am. Acad. Child. Adolesc. Psychiatry,
der in drug resistant mesial temporal lobe epilepsy, Epilepsy
59, 842 855, doi: 10.1016/j.jaac.2019.10.011.
Behav., 115, 107548, doi: 10.1016/j.yebeh.2020.107548.
21.
Gulyaeva, N. V. (2019) Biochemical mechanisms and
7.
Kim, S. J., Lee, S. A., Ryu, H. U., Han, S. H., Lee, G. H.,
translational relevance of hippocampal vulnerability to dis
et al. (2020) Factors associated with obsessive compulsive
tant focal brain injury: the price of stress response,
symptoms in people with epilepsy, Epilepsy Behav., 102,
Biochemistry (Moscow), 84, 13061328, doi: 10.1134/
106723, doi: 10.1016/j.yebeh.2019.106723.
S0006297919110087.
8.
Novais, F., Franco, A., Loureiro, S., Andrea, M., Figueira,
22.
Joëls, M.
(2018) Corticosteroids and the brain,
M. L., Pimentel, J., and Pestana, L. C. (2019) Personality
J. Endocrinol., 238, R121 R130, doi: 10.1530/JOE 18
patterns of people with medically refractory epilepsy -
0226.
does the epileptogenic zone matter? Epilepsy Behav., 97,
23.
Gulyaeva, N. V. (2015) Ventral hippocampus, stress and
130 134, doi: 10.1016/j.yebeh.2019.05.049.
psychopathology: translational implications, Neurochem. J.,
9.
Ho, P. H., Leung, W. C. Y., Leung, I. Y. H., and Chang,
9, 85 94, doi: 10.1134/S1819712415020075.
R. S. K. (2020) Factors associated with depression in peo
24.
Licht, T., Sasson, E., Bell, B., Grunewald, M., Kumar, S.,
ple with epilepsy: a retrospective case control analysis,
et al. (2020) Hippocampal neural stem cells facilitate
Hong Kong Med. J.,
26,
311317, doi:
10.12809/
access from circulation via apical cytoplasmic processes,
hkmj198310.
Elife, 9, e52134, doi: 10.7554/eLife.52134.
10.
Sankar, R., and Mazarati, A. (2012) Neurobiology of
25.
Tuem, K. B., and Atey, T. M. (2017) Neuroactive steroids:
depression as a comorbidity of epilepsy, in Jasper’s Basic
receptor interactions and responses, Front. Neurol., 8, 442,
Mechanisms of the Epilepsies [Internet] (Noebels, J. L.,
doi: 10.3389/fneur.2017.00442.
Avoli, M., Rogawski, M. A., Olsen, R. W., and Delgado
26.
Saboory, E., Mohammadi, S., Dindarian, S., and
Escueta, A. V., eds) 4th Edn., Bethesda (MD): National
Mohammadi, H. (2019) Prenatal stress and elevated
Center for Biotechnology Information (US).
seizure susceptibility: molecular inheritable changes,
11.
Duric, V., Clayton, S., Leong, M. L., and Yuan, L. L.
Epilepsy Behav., 96, 122 131, doi: 10.1016/j.yebeh.2019.
(2016) Comorbidity factors and brain mechanisms linking
04.046.
chronic stress and systemic illness, Neural Plast., 2016,
27.
Galtrey, C. M., Mula, M., and Cock, H. R. (2016) Stress
5460732, doi: 10.1155/2016/5460732.
and epilepsy: fact or fiction, and what can we do about it?
12.
Du Preez, A., Eum, J., Eiben, I., Eiben, P., Zunszain,
Pract. Neurol., 16, 270 278, doi: 10.1136/practneurol
P. A., et al. (2020) Do different types of stress differentially
2015 001337.
alter behavioural and neurobiological outcomes associated
28.
Novakova, B., Harris, P. R., Ponnusamy, A., and
with depression in rodent models? A systematic review,
Reuber, M. (2013) The role of stress as a trigger for epilep
Front. Neuroendocrinol., 61, 100896, doi: 10.1016/j.yfrne.
tic seizures: a narrative review of evidence from human and
2020.100896.
animal studies, Epilepsia, 54, 1866 76, doi: 10.1111/epi.
13.
Khan, A. R., Geiger, L., Wiborg, O., and Czéh, B. (2020)
12377.
Stress induced morphological, cellular and molecular
29.
Kanner, A. M., Schachter, S. C., Barry, J. J., Hesdorffer,
changes in the brain lessons learned from the chronic mild
D. C., Mula, M., et al. (2012) Depression and epilepsy:
stress model of depression, Cells, 9, 1026, doi: 10.3390/
epidemiologic and neurobiologic perspectives that may
cells9041026.
explain their high comorbid occurrence, Epilepsy Behav.,
14.
Li, M., Fu, X., Xie, W., Guo, W., Li, B., et al. (2020) Effect
24, 156 68, doi: 10.1016/j.yebeh.2012.01.007.
of early life stress on the epigenetic profiles in depression,
30.
McKee, H. R., and Privitera, M. D. (2017) Stress as a
Front. Cell. Dev. Biol., 8, 867, doi: 10.3389/fcell.2020.00867.
seizure precipitant: Identification, associated factors, and
15.
Negrón Oyarzo, I., Aboitiz, F., and Fuentealba, P. (2016)
treatment options, Seizure, 44, 2126, doi: 10.1016/
Impaired functional connectivity in the prefrontal cortex: a
j.seizure.2016.12.009.
mechanism for chronic stress induced neuropsychiatric
31.
Gunn, B. G., and Baram, T. Z. (2017) Stress and seizures:
disorder, Neural Plast., 2016, 753906, doi: 10.1155/2016/
space, time and hippocampal circuits, Trends Neurosci.,
7539065.
40, 667 679, doi: 10.1016/j.tins.2017.08.004.
16.
Ross, R. A., Foster, S. L., and Ionescu, D. F. (2017) The
32.
Cano López, I., and González Bono, E. (2019) Cortisol
role of chronic stress in anxious depression, chronic stress
levels and seizures in adults with epilepsy: a systematic
(thousand oaks), Chronic Stress, 1, 2470547016689472,
review, Neurosci. Biobehav. Rev.,
103,
216229,
doi: 10.1177/2470547016689472.
doi: 10.1016/j.neubiorev.2019.05.023.
БИОХИМИЯ том 86 вып. 6 2021
СТРЕСС, ГИППОКАМП, ЭПИЛЕПСИЯ И КОМОРБИДНАЯ ДЕПРЕССИЯ
801
33.
Kotwas, I., McGonigal, A., Bastien Toniazzo, M.,
markers, Biomed. Res. Int., 2014, 932757, doi: 10.1155/
Bartolome, I. F., and Micoulaud Franchi, J. A. (2017)
2014/932757.
Stress regulation in drug resistant epilepsy, Epilepsy
49.
Kanner, A. M. (2008) Mood disorder and epilepsy: a neu
Behav., 71 (Pt A), 3950, doi: 10.1016/j.yebeh.2017.
robiologic perspective of their relationship, Dialogues Clin.
01.025.
Neurosci., 10, 39 45, doi: 10.31887/DCNS.2008.10.1/
34.
Yang, Y., Yang, M., Shi, Q., Wang, T., and Jiang, M. (2020)
amkanner.
Risk factors for depression in patients with epilepsy: a
50.
Pauli, E., and Stefan, H. (2009) Emotional and affective
meta analysis, Epilepsy Behav., 106, 107030, doi: 10.1016/
disorders, anxiety and personality disorders in epilepsies,
j.yebeh.2020.107030.
Nervenarzt, 80, 1440 1451, doi: 10.1007/s00115 009 2740 2.
35.
Błaszczyk, B., and Czuczwar, S. J. (2016) Epilepsy coex
51.
Gulyaeva, N. V. (2021) Glucocorticoid regulation of the
isting with depression, Pharmacol. Rep., 68, 1084 1092,
glutamatergic synapse: Mechanisms of stress dependent
doi: 10.1016/j.pharep.2016.06.011.
neuroplasticity, Ross. Fiziologicheskii Zhurnal im. I. M.
36.
Hingray, C., McGonigal, A., Kotwas, I., and Micoulaud
Sechenova (Sechenov Physiology Journal), 107, 518 532,
Franchi, J. A. (2019) The relationship between epilepsy
doi: 10.31857/S0869813921040099.
and anxiety disorders, Curr. Psychiatry Rep., 21, 40,
52.
Conrad, C. D. (2008) Chronic stress induced hippocam
doi: 10.1007/s11920 019 1029 9.
pal vulnerability: the glucocorticoid vulnerability hypothe
37.
Wulsin, A. C., Solomon, M. B., Privitera, M. D., Danzer,
sis, Rev. Neurosci., 19, 395 411, doi: 10.1515/revneuro.
S. C., and Herman, J. P. (2016) Hypothalamic pitui
2008.19.6.395.
tary adrenocortical axis dysfunction in epilepsy,
53.
Van Campen, J. S., Valentijn, F.A., Jansen, F. E., Joels, M.,
Physiol. Behav., 166, 22 31, doi: 10.1016/j.physbeh.2016.
and Braun, K. P. (2015) Seizure occurrence and the
05.015.
circadian rhythm of cortisol: a systematic review,
38.
Jones, N. C., and O’Brien, T. J. (2013) Stress, epilepsy, and
Epilepsy Behav., 47, 132 137, doi: 10.1016/j.yebeh.2015.
psychiatric comorbidity: how can animal models inform
04.071.
the clinic? Epilepsy Behav., 26, 363 369, doi: 10.1016/
54.
Van Campen, J. S., Hompe, E.L., Jansen, F. E., Velis,
j.yebeh.2012.09.002.
D.N., Otte, W.M., et al. (2016) Cortisol fluctuations relate
39.
Pochigaeva, K., Druzhkova, T., Yakovlev, A., Onufriev, M.,
to interictal epileptiform discharges in stress sensitive
Grishkina, M., et al. (2017) Hair cortisol as a marker of
epilepsy, Brain, 139 (Pt 6), 1673 1679, doi: 10.1093/
hypothalamic pituitary adrenal Axis activity in female
brain/aww071.
patients with major depressive disorder, Metab. Brain Dis.,
55.
D’Alessio, L., Mesarosova, L., Anink, J. J., Kochen, S.,
32, 577 583, doi: 10.1007/s11011 017 9952 0.
Sol s, P., et al. (2020) Reduced expression of the glucocor
40.
Kanner, A. M. (2017) Can neurochemical changes of
ticoid receptor in the hippocampus of patients with drug
mood disorders explain the increase risk of epilepsy or its
resistant temporal lobe epilepsy and comorbid depression,
worse seizure control? Neurochem. Res., 42, 2071 2076,
Epilepsia, 61, 1595 1605, doi: 10.1111/epi.16598.
doi: 10.1007/s11064 017 2331 8.
56.
Wulsin, A. C., Franco Villanueva, A., Romancheck, C.,
41.
Gulyaeva, N. V. (2019) Functional neurochemistry of the
Morano, R. L., Smith, B. L., et al. (2018) Functional dis
ventral and dorsal hippocampus: Stress, depression,
ruption of stress modulatory circuits in a model of tempo
dementia and remote hippocampal damage, Neurochem.
ral lobe epilepsy, PLoS One, 13, e0197955, doi: 10.1371/
Res., 44, 1306 1322, doi: 10.1007/s11064 018 2662 0.
journal.pone.0197955.
42.
Maguire, J., Salpekar, J. A. (2013) Stress, seizures, and
57.
Wulsin, A. C., Herman, J. P., and Danzer, S. C. (2016)
hypothalamic pituitary adrenal axis targets for the treat
RU486 mitigates hippocampal pathology following status
ment of epilepsy, Epilepsy Behav.,
26,
352362,
epilepticusm, Front. Neurol., 7, 214, doi: 10.3389/fneur.
doi: 10.1016/j.yebeh.2012.09.040.
2016.00214.
43.
Basu, T., Maguire, J., Salpekar, J. A. (2021) Hypothalamic
58.
Aniol, V. A., Ivanova Dyatlova, A. Y., Keren, O., Guekht,
pituitary adrenal axis targets for the treatment of epilepsy,
A. B., Sarne, Y., and Gulyaeva, N. V. (2013) A single
Neurosci. Lett., 746, 135618, doi: 10.1016/j.neulet.2020.
pentylenetetrazole induced clonic tonic seizure episode is
135618.
accompanied by a slowly developing cognitive decline in
44.
Gulyaeva, N. V. (2015) Neuronal plasticity and epilepsy:
rats, Epilepsy Behav., 26, 196 202, doi: 10.1016/j.yebeh.
modern concepts and mechanisms of epilepsy and depres
2012.12.006.
sion comorbidity, Zh. Nevrol. Psikhiatr. Im. S. S. Korsakova,
59.
Gulyaeva, N. V. (2010) Aberrant neurogenesis in adult
115, 148 153, doi: 10.17116/jnevro2015115112148 153.
epileptic brain: compensatory or pathologic, Neurochem. J.,
45.
Gulyaeva, N. V. (2016) Cerebral plasticity and con
4, 84 89, doi: 10.1134/S1819712410020029.
nectopathies: mechanisms of comorbidity of neurolog
60.
Podgorny, O. V., and Gulyaeva, N. V. (2021) Glucocorti
ical diseases and depression, Zh. Nevrol. Psikhiatr.
coid mediated mechanisms of hippocampal damage:
Im. S. S. Korsakova, 116, 157162, doi: 10.17116/
Contribution of subgranular neurogenesis, J. Neurochem.,
jnevro2016116111157 162.
157, 370 392, doi: 10.1111/jnc.15265.
46.
Gulyaeva, N. V. (2017) Staging of neuroplasticity alter
61.
Kuruba, R., Hattiangady, B., and Shetty, A. K. (2009)
ations during epileptogenesis (temporal lobe epilepsy as an
Hippocampal neurogenesis and neural stem cells in tem
example), Zh. Nevrol. Psikhiatr. Im. S. S. Korsakova, 117,
poral lobe epilepsy, Epilepsy Behav., 14 (Suppl 1), 65 73,
10 16, doi: 10.17116/jnevro20171179210 16.
doi: 10.1016/j.yebeh.2008.08.020.
47.
Gulyaeva, N. V. (2020) Physiological continuum of plastic
62.
Parent, J. M., and Lowenstein, D. H. (2002) Seizure
ity and pathology of the nervous system [in Russian],
induced neurogenesis: are more new neurons good for an
Integr. Physiol., 1, 294 302, doi: 10.33910/2687 1270
adult brain? Prog. Brain. Res., 135, 121 131, doi: 10.1016/
2020 1 4 294 302.
S0079 6123(02)35012 X.
48.
Stepanichev, M., Dygalo, N. N., Grigoryan, G.,
63.
Parent, J. M. (2002) The role of seizure induced neuroge
Shishkina, G. T., and Gulyaeva, N. (2014) Rodent models
nesis in epileptogenesis and brain repair, Epilepsy Res., 50,
of depression: neurotrophic and neuroinflammatory bio
179 189, doi: 10.1016/s0920 1211(02)00078 5.
4 БИОХИМИЯ том 86 вып. 6 2021
802
ГУЛЯЕВА
64.
Parent, J. M. (2007) Adult neurogenesis in the intact and
epilepsy, Curr. Neurol. Neurosci. Rep., 20, 5, doi: 10.1007/
epileptic dentate gyrus, Prog. Brain Res., 163, 529 540,
s11910 020 1025 9.
doi: 10.1016/S0079 6123(07)63028 3.
82.
Mukherjee, S., Arisi, G. M., Mims, K., Hollingsworth, G.,
65.
Kokaia, M. (2011) Seizure induced neurogenesis in the
O’Neil, K., and Shapiro, L. A. (2020) Neuroinflam
adult brain, Eur. J. Neurosci., 33, 1133 1138, doi: 10.1111/
matory mechanisms of post traumatic epilepsy,
j.1460 9568.2011.07612.x.
J. Neuroinflammation, 17, 193, doi: 10.1186/s12974 020
66.
Jessberger, S., and Parent, J. M. (2015) Epilepsy and adult
01854 w.
neurogenesis, Cold Spring Harb Perspect. Biol., 7, a020677,
83.
Sanz, P., and Garcia Gimeno, M. A. (2020) Reactive glia
doi: 10.1101/cshperspect.a020677.
inflammatory signaling pathways and epilepsy, Int. J. Mol.
67.
Scharfman, H. E. (2004) Functional implications of
Sci., 21, 4096, doi: 10.3390/ijms21114096.
seizure induced neurogenesis, Adv. Exp. Med. Biol., 548,
84.
Meyer, J. H., Cervenka, S., Kim, M. J., Kreisl, W. C.,
192 212, doi: 10.1007/978 1 4757 6376 8_14.
Henter, I. D., and Innis, R. B. (2020) Neuroinflammation
68.
Scharfman, H. E., and Gray, W. P. (2007) Relevance of
in psychiatric disorders: PET imaging and promising new
seizure induced neurogenesis in animal models of epilepsy
targets, Lancet Psychiatry, 7, 1064 1074, doi: 10.1016/
to the etiology of temporal lobe epilepsy, Epilepsia, 48
S2215 0366(20)30255 8.
(Suppl 2), 33 41, doi: 10.1111/j.1528 1167.2007.01065.x.
85.
Troubat, R., Barone, P., Leman, S., Desmidt, T.,
69.
Siebzehnrubl, F. A., and Blumcke, I. (2008) Neurogenesis
Cressant, A., et al. (2021) Neuroinflammation and depres
in the human hippocampus and its relevance to temporal
sion: a review, Eur. J. Neurosci.,
53,
151171,
lobe epilepsies, Epilepsia,
49 (Suppl
5),
5565,
doi: 10.1111/ejn.14720.
doi: 10.1111/j.1528 1167.2008.01638.x.
86.
Ting, E. Y., Yang, A. C., and Tsai, S. J. (2020) Role of
70.
Hattiangady, B., and Shetty, A. K. (2008) Implications of
interleukin 6 in depressive disorder, Int. J. Mol. Sci., 21,
decreased hippocampal neurogenesis in chronic temporal
2194, doi: 10.3390/ijms21062194.
lobe epilepsy, Epilepsia,
49 (Suppl
5),
2641,
87.
Tsai, S. J. (2021) Role of interleukin 8 in depression and
doi: 10.1111/j.1528 1167.2008.01635.x.
other psychiatric disorders, Prog. Neuropsychopharmacol.
71.
Danzer, S. C. (2012) Depression, stress, epilepsy and adult
Biol. Psychiatry, 106, 110173, doi: 10.1016/j.pnpbp.2020.
neurogenesis, Exp. Neurol., 233, 22 32, doi: 10.1016/j.exp
110173.
neurol.2011.05.023.
88.
Arisi, G. M. (2014) Nervous and immune systems signals
72.
Braun, S. M., and Jessberger, S. (2014) Adult neurogenesis
and connections: cytokines in hippocampus physiology
and its role in neuropsychiatric disease, brain repair and
and pathology, Epilepsy Behav., 38, 43 47, doi: 10.1016/
normal brain function, Neuropathol. Appl. Neurobiol., 40,
j.yebeh.2014.01.017.
3 12, doi: 10.1111/nan.12107.
89.
Devinsky, O., Vezzani, A., Najjar, S., De Lanerolle, N. C.,
73.
Abrous, D. N., Koehl, M., and Le Moal, M. (2005) Adult
and Rogawski, M. A. (2013) Glia and epilepsy: excitability
neurogenesis: from precursors to network and physiology,
and inflammation, Trends Neurosci.,
36,
174184,
Physiol. Rev., 85, 523 569, doi: 10.1152/physrev.00055.2003.
doi: 10.1016/j.tins.2012.11.008.
74.
Hayashi, Y., Jinnou, H., Sawamoto, K., and Hitoshi, S.
90.
Lu, J., Huang, H., Zeng, Q., Zhang, X., Xu, M., et al.
(2018) Adult neurogenesis and its role in brain injury and
(2019) Hippocampal neuron loss and astrogliosis in medi
psychiatric diseases, J. Neurochem.,
147,
584594,
al temporal lobe epileptic patients with mental disorders,
doi: 10.1111/jnc.14557.
J. Integr. Neurosci., 18, 127 132, doi: 10.31083/j.jin.2019.
75.
Lino de Oliveira, C., Bolzan, J. A., Surget, A., and
02.16.
Belzung, C. (2020) Do antidepressants promote neurogen
91.
Albrecht, J., and Zielińska, M. (2017) Mechanisms of
esis in adult hippocampus? A systematic review and meta
excessive extracellular glutamate accumulation in and tem
analysis on naive rodents, Pharmacol. Ther., 210, 107515,
poral lobe epilepsy, Neurochem. Res., 42, 1724 1734,
doi: 10.1016/j.pharmthera.2020.107515.
doi: 10.1007/s11064 016 2105 8.
76.
Patel, D., Kas, M. J., Chattarji, S., and Buwalda, B. (2019)
92.
Barker Haliski, M., and White, H. S. (2015) Glutamat
Rodent models of social stress and neuronal plasticity: rel
ergic mechanisms associated with seizures and epilepsy,
evance to depressive like disorders, Behav. Brain Res., 369,
Cold Spring Harb. Perspect. Med., 5, a022863, doi: 10.1101/
111900, doi: 10.1016/j.bbr.2019.111900.
cshperspect.a022863.
77.
Planchez, B., Surget, A., and Belzung, C. (2020) Adult hip
93.
Eid, T., Gruenbaum, S. E., Dhaher, R., Lee, T. W.,
pocampal neurogenesis and antidepressants effects, Curr. Opin.
Zhou, Y., and Danbolt, N. C. (2016) The glutamate-glut
Pharmacol., 50, 88 95, doi: 10.1016/j.coph.2019.11.009.
amine cycle in epilepsy, Adv. Neurobiol., 13, 351 400,
78.
D’Alessio, L., Konopka, H., Sol s, P., Scévola, L., Lima,
doi: 10.1007/978 3 319 45096 4_14.
M. F., et al. (2019) Depression and temporal lobe epilepsy:
94.
Nicolo, J. P., O’Brien, T. J., and Kwan, P. (2019) Role of
expression pattern of calbindin immunoreactivity in
cerebral glutamate in post stroke epileptogenesis,
hippocampal dentate gyrus of patients who underwent
Neuroimage Clin., 24, 102069, doi: 10.1016/j.nicl.2019.
epilepsy surgery with and without comorbid depression,
102069.
Behav. Neurol., 2019, 7396793, doi: 10.1155/2019/7396793.
95.
O’Neil, D. A., Nicholas, M. A., Lajud, N., Kline, A. E.,
79.
Vezzani, A., Balosso, S., and Ravizza, T.
(2019)
and Bondi, C. O. (2018) Preclinical models of traumatic
Neuroinflammatory pathways as treatment targets and bio
brain injury: emerging role of glutamate in the pathophysi
markers in epilepsy, Nat. Rev. Neurol., 15, 459472,
ology of depression, Front. Pharmacol.,
9,
579,
doi: 10.1038/s41582 019 0217 x.
doi: 10.3389/fphar.2018.00579.
80.
Vezzani, B., Carinci, M., Patergnani, S., Pasquin, M. P.,
96.
Cui, W., Ning, Y., Hong, W., Wang, J., Liu, Z., and Li,
Guarino, A., et al. (2020) The dichotomous role of inflam
M. D. (2019) Crosstalk between inflammation and gluta
mation in the CNS: a mitochondrial point of view,
mate system in depression: signaling pathway and molecu
Biomolecules, 10, 1437, doi: 10.3390/biom10101437.
lar biomarkers for ketamine’s antidepressant effect, Mol.
81.
Sharma, A. A., and Szaflarski, J. P. (2020) In vivo imaging
Neurobiol., 56, 34843500, doi: 10.1007/s12035 018
of neuroinflammatory targets in treatment resistant
1306 3.
БИОХИМИЯ том 86 вып. 6 2021
СТРЕСС, ГИППОКАМП, ЭПИЛЕПСИЯ И КОМОРБИДНАЯ ДЕПРЕССИЯ
803
97. Fullana, N., Gasull Camós, J., Tarrés Gatius, M.,
111.
McNamara, J. O., and Scharfman, H. E. (2012) Temporal
Castañé, A., Bortolozzi, A., and Artigas, F.
(2020)
lobe epilepsy and the BDNF receptor, TrkB, in Jasper’s
Astrocyte control of glutamatergic activity: downstream
Basic Mechanisms of the Epilepsies
[Internet]
effects on serotonergic function and emotional behavior,
(Noebels, J. L., Avoli, M., Rogawski, M. A., Olsen, R. W.,
Neuropharmacology, 166, 107914, doi: 10.1016/j.neuropharm.
and Delgado Escueta, A. V., eds.) 4th Edn., Bethesda
2019.107914.
(MD): National Center for Biotechnology Inform
98. Henter, I. D., de Sousa, R. T., and Zarate, C. A., Jr.
ation (US).
(2018) Glutamatergic modulators in depression, Harv.
112. Brunoni, A. R., Lopes, M., and Fregni, F. (2008) A sys
Rev. Psychiatry,
26,
307319, doi:
10.1097/HRP.
tematic review and meta analysis of clinical studies on
0000000000000183.
major depression and BDNF levels: implications for the
99. Li, C. T., Yang, K. C., and Lin, W. C. (2019) Glutamatergic
role of neuroplasticity in depression, Int. J.
dysfunction and glutamatergic compounds for major psychi
Neuropsychopharmacol., 11, 11691180, doi: 10.1017/
atric disorders: evidence from clinical neuroimaging studies,
S1461145708009309.
Front. Psychiatry, 9, 767, doi: 10.3389/fpsyt.2018.00767.
113. Martinowich, K., Manji, H., and Lu, B. (2007) New
100. Reus, G. Z., de Moura, A. B., Silva, R. H., Resende,
insights into BDNF function in depression and anxiety,
W. R., and Quevedo, J. (2018) Resilience dysregulation in
Nat. Neurosci., 10, 1089 1093, doi: 10.1038/nn1971.
major depressive disorder: focus on glutamatergic imbal
114. Gulyaeva, N. V. (2017) Interplay between brain BDNF and
ance and microglial activation, Curr. Neuropharmacol., 16,
glutamatergic systems: a brief state of the evidence and
297 307, doi: 10.2174/1570159X15666170630164715.
association with the pathogenesis of depression,
101. Gulyaeva, N. V. (2021) Hippocampal hyperglutamatergic
Biochemistry (Moscow),
82,
301307, doi:
10.1134/
signaling matters: early targeting glutamate neurotransmis
S0006297917030087.
sion as a preventive strategy in Alzheimer’s disease: an edi
115. Bolshakov, A. P., Tret’yakova, L. V., Kvichansky, A. A., and
torial highlight for “riluzole attenuates glutamatergic tone
Gulyaeva, N. V. (2021) Glucocorticoids: Dr. Jekyll
and cognitive decline in AβPP/PS1 mice” on https://
and Mr. Hyde of hippocampal neuroinflammation,
doi.org/10.1111/jnc.15224, J. Neurochem., 156, 399 402,
Biochemistry (Moscow),
86,
156167, doi:
10.1134/
doi: 10.1111/jnc.15238.
S0006297921020048.
102. Hanada, T. (2020) Ionotropic glutamate receptors in
116. Alexander, A., Maroso, M., and Soltesz, I.
(2016)
epilepsy: a review focusing on AMPA and NMDA recep
Organization and control of epileptic circuits in temporal
tors, Biomolecules, 10, 464, doi: 10.3390/biom10030464.
lobe epilepsy, Prog. Brain Res.,
226,
127154,
103. Mihály, A. (2019) The reactive plasticity of hippocampal
doi: 10.1016/bs.pbr.2016.04.007.
ionotropic glutamate receptors in animal epilepsies, Int. J.
117. Valente, K. D., and Busatto, F. G. (2013) Depression and
Mol. Sci., 20, 1030, doi: 10.3390/ijms20051030.
temporal lobe epilepsy represent an epiphenomenon shar
104. Qian, F., and Tang, F. R. (2016) Metabotropic glutamate
ing similar neural networks: clinical and brain structural
receptors and interacting proteins in epileptogenesis,
evidences, Arq. Neuropsiquiatr., 71, 183 190, doi: 10.1590/
Curr. Neuropharmacol.,
14,
551562, doi:
10.2174/
s0004 282x2013000300011.
1570159x14666160331142228.
118. Krishnan, V. (2020) Depression and anxiety in the epilep
105. Miladinovic, T., Nashed, M. G., and Singh, G. (2015)
sies: from bench to bedside, Curr. Neurol. Neurosci. Rep.,
Overview of glutamatergic dysregulation in central
20, 41, doi: 10.1007/s11910 020 01065 z.
pathologies, Biomolecules, 5, 3112 3141, doi: 10.3390/
119. Elkommos, S., and Mula, M. (2020) A systematic review of
biom5043112.
neuroimaging studies of depression in adults with epilepsy,
106. Sadeghi, M. A., Hemmati, S., Mohammadi, S., Yousefi
Epilepsy Behav., 115, 107695, doi: 10.1016/j.yebeh.
Manesh, H., Vafaei, A., et al. (2021) Chronically altered
2020.107695.
NMDAR signaling in epilepsy mediates comorbid depres
120. Rivera Bonet, C. N., Hwang, G., Hermann, B., Struck,
sion, Acta Neuropathol. Commun., 9, 53, doi: 10.1186/
A. F., Cook, C. J., et al. (2020) Neuroticism in temporal
s40478 021 01153 2.
lobe epilepsy is associated with altered limbic frontal lobe
107. Scarante, F. F., Vila Verde, C., Detoni, V. L., Ferreira
resting state functional connectivity, Epilepsy Behav., 110,
Junior, N. C., Guimarães, F. S., and Campos, A. C. (2017)
107172, doi: 10.1016/j.yebeh.2020.107172.
Cannabinoid modulation of the stressed hippocampus,
121. Szot, P. (2012) Common factors among Alzheimer’s dis
Front. Mol. Neurosci., 10, 411, doi: 10.3389/fnmol.
ease, Parkinson’s disease, and epilepsy: possible role of the
2017.00411.
noradrenergic nervous system., Epilepsia, 53 (Suppl 1), 61
108. Cheung, K. A. K., Peiris, H., Wallace, G., Holland, O. J.,
66, doi: 10.1111/j.1528 1167.2012.03476.x.
and Mitchell, M. D. (2019) The interplay between the
122. Kondziella, D., Alvestad, S., Vaaler, A., and
endocannabinoid system, epilepsy and cannabinoids,
Sonnewald, U. (2007) Which clinical and experimental
Int. J. Mol. Sci., 20, 6079, doi: 10.3390/ijms20236079.
data link temporal lobe epilepsy with depression?
109. Rocha, L., Alonso Vanegas, M., Orozco Suárez, S.,
J. Neurochem., 103, 2136 2152, doi: 10.1111/j.1471 4159.
Alcántara González, D., Cruzblanca, H., and Castro, E.
2007.04926.x.
(2014) Do certain signal transduction mechanisms explain
123. Druzhkova, T., Pochigaeva, K., Yakovlev, A.,
the comorbidity of epilepsy and mood disorders? Epilepsy
Kazimirova, E., Grishkina, M., et al. (2019) Acute stress
Behav., 38, 25 31, doi: 10.1016/j.yebeh.2014.01.001.
response to a cognitive task in patients with major depres
110. Koyama, R., and Ikegaya, Y. (2005) To BDNF or not to
sive disorder: potential metabolic and proinflammatory
BDNF: that is the epileptic hippocampus, Neuroscientist,
biomarkers, Metab. Brain. Dis., 34, 621 629, doi: 10.1007/
11, 282 287, doi: 10.1177/1073858405278266.
s11011 018 0367 3.
БИОХИМИЯ том 86 вып. 6 2021
4*
804
ГУЛЯЕВА
STRESS ASSOCIATED SHARED MOLECULAR AND CELLULAR
HIPPOCAMPAL MECHANISMS OF EPILEPSY
AND COMORBID DEPRESSIVE DISORDERS
Review
N. V. Gulyaeva1,2
1 Institute of Higher Nervous Activity and Neurophysiology, Russian Academy of Sciences,
117485 Moscow, Russia; e'mail: nata_gul@ihna.ru
2 Research and Clinical Center for Neuropsychiatry of Moscow Healthcare Department, 115419 Moscow, Russia
The review discusses molecular and cellular mechanisms common to the temporal lobe epileptogenesis/epilepsy and
depressive disorders. Comorbid temporal lobe epilepsy and depression are associated with dysfunction of the hypo
thalamic pituitary adrenocortical axis. Excessive glucocorticoids disrupt the function and impair the structure of the
hippocampus, a brain region key to learning, memory, and emotions. Selective vulnerability of the hippocampus to
stress, mediated by the reception of glucocorticoid hormones secreted during stress, is the price of the high function
al plasticity and pleiotropy of this limbic structure. Common molecular and cellular mechanisms include the dys
function of glucocorticoid receptors, neurotransmitters, and neurotrophic factors, development of neuroinflamma
tion, leading to neurodegeneration and loss of hippocampal neurons, as well as disturbances in neurogenesis in the
subgranular neurogenic niche and formation of aberrant neural networks. These glucocorticoid dependent processes
underlie altered stress response and the development of chronic stress induced comorbid pathologies, in particular,
temporal lobe epilepsy and depressive disorders.
Keywords: epilepsy, temporal lobe epilepsy, depression, hippocampus, glucocorticoids, stress, neuroinflammation,
hypothalamic pituitary adrenocortical axis, neurogenesis, neural networks
БИОХИМИЯ том 86 вып. 6 2021