БИОХИМИЯ, 2021, том 86, вып. 7, с. 964 - 984
УДК 578.834;578.832
COVID 19: МИФЫ И РЕАЛЬНОСТЬ
Обзор
© 2021
Л.В. Кордюкова1*, А.В. Шанько2,3
1 НИИ физико химической биологии имени А.Н. Белозерского, Московский государственный университет
имени М.В. Ломоносова, 119991 Москва, Россия; электронная почта: kord@belozersky.msu.ru
2 ООО ФОРТ, Департамент науки, 119435 Москва, Россия
3 НИИ вирусологии имени Д.И. Ивановского, «НИЦЭМ им. Н. Ф. Гамалеи»
Минздрава России, 123098 Москва, Россия
Поступила в редакцию 07.04.2021
После доработки 28.04.2021
Принята к публикации 28.04.2021
COVID 19, новое респираторное заболевание человека, жертвами которого за год пандемии стали около
3 млн человек, является глобальным вызовом здравоохранению во всем мире. Вызвавший его инфекцион
ный агент - новый коронавирус SARS CoV 2 - имеет ряд существенных структурных отличий от других ко
ронавирусов, что делает его более патогенным и способным быстро распространяться. В настоящем обзоре
мы рассмотрим важные характеристики основного поверхностного антигена SARS CoV 2 (шиповидного
S белка), такие как: (1) способность рецептор связывающего домена (RBD) переходить из положения
«стоя» («открытая» префузионная конформация) в положение «лежа» («закрытая» префузионная конфор
мация) для ускользания от нейтрализующих антител; (2) использование преимуществ высокого сродства
связывания RBD в открытой конформации с рецептором - ангиотензинпревращающим ферментом 2 чело
века для эффективного входа в клетку; (3) предактивация S белка внутриклеточными фуриноподобными
протеазами для распространения вируса по разным типам клеток. Мы рассмотрим взаимодействие S белка
с клеточными рецепторами, корецепторами и антагонистами и опишем гипотетический механизм дестаби
лизации гомотримерной структуры «шипа», запускающий слияние оболочки вириона с мембраной клетки
при физиологическом значении рН и опосредующий вход вирусного нуклеокапсида в цитоплазму. Крайне
важным с практической точки зрения, особенно при производстве вакцин, является обнаруженное свой
ство перехода префузионной конформации S белка в постфузионную на поверхности вирионов в случае
инактивации их такими реагентами, как β пропиолактон. Мы также сравним несколько особенностей
COVID 19 и S белка вируса SARS CoV 2 со вспышками тяжелого гриппа, вызванными высокопатогенны
ми штаммами вируса гриппа А подтипов H5 и H7, и структурными характеристиками молекул гемагглюти
нина этих штаммов. В конце обзора коротко перечислены перспективные и внедренные в практику анти
вирусные и патогенетические средства терапии COVID 19; рассмотрены традиционные и инновационные
вакцины, недавно одобренные для профилактики заболевания, и их молекулярно иммунологические осо
бенности.
КЛЮЧЕВЫЕ СЛОВА: COVID 19, SARS CoV 2, S белок, структура, вирус гриппа, гемагглютинин, вакцины.
DOI: 10.31857/S0320972521070022
ВВЕДЕНИЕ
же - SARS CoV 2 (severe acute respiratory syn
drome coronavirus 2). В феврале 2020 года Все
Эпидемия заболевания с симптомами ати
мирная организация здравоохранения назвала
пичной пневмонии началась в городе Ухань
данное заболевание «COVID 19» (COronaVIrus
(провинция Хубей, Китай) зимой 2019 года. При
Disease 2019). Эпидемия стала глобальной и пе
чиной позднее был признан новый коронавирус
решла в пандемию. Большинство пациентов,
человека (2019 nCoV, 2019 novel coronavirus), он
инфицированных вирусом SARS CoV 2, испы
Принятые сокращения: крио ЭМ - криоэлектронная микроскопия; крио ЭТ - криоэлектронная томография;
MHC - комплекс гистосовместимости; СТ - cytoplasmic tail (цитоплазматический домен); TM - трансмембранный;
ACE2 - angiotensin converting enzyme 2 (ангиотензинпревращающий фермент 2); COVID 19 - COronaVIrus Disease 2019
(новое коронавирусное заболевание 2019); ERGIC - компартмент эндоплазматического ретикулума и аппарата Гольджи;
HA - гемагглютинин; IFN - интерферон; RBD - рецептор связывающий домен; RBM - рецептор связывающий мотив;
SARS CoV 2 - severe acute respiratory syndrome coronavirus 2 (коронавирус, вызывающий тяжелый острый респираторный
синдром 2).
* Адресат для корреспонденции.
964
COVID 19: МИФЫ И РЕАЛЬНОСТЬ
965
тывают легкие или умеренные симптомы, кото
Тяжёлое течение COVID 19, характеризую
рые исчезают через 6-10 дней. Однако почти у
щееся совокупностью патогенетическиих фак
20% пациентов болезнь протекает с серьёзными
торов, имеет сходство с гриппом, но только не
осложнениями, включая атипичную двусторон
сезонным, а тяжёлым, вызываемым разнообраз
ную пневмонию и острый респираторный дист
ными представителями другого семейства обо
ресс синдром, что чревато высокой леталь
лочечных вирусов - Orthomyxoviridae, в част
ностью [1]. На 5 апреля 2021 г. людские потери
ности, пандемическими штаммами 2009 г. под
от COVID 19 исчисляются 2,85 млн при 131 млн
типа H1N1 и вызвавшими вспышки «птичьего»
заболевших (приблизительно 1-3% летальности
гриппа подтипами H5N1 и H7N9. При этом оче
по данным Всемирной Организации Здравоох
видны параллели в некоторых структурных эле
ментах поверхностных белков этих вирусов, ге
нения ежегодные вспышки сезонного гриппа,
магглютинина (HA) вируса гриппа и S белка
по некоторым оценкам, уносят 250 000-500 000
SARS CoV 2, а также в характере протекания
жизней ежегодно [3], а смертность во время
заболевания, которые будут отмечены.
пандемии гриппа подтипа H1N1 в 2009 году со
В конце обзора будут затронуты проблемы
ставила от 151 700 до 575 400 человек [4].
блокады вирусной инфекции и перечислены ис
Оболочечные РНК содержащие вирусы се
пользуемые стратегии терапии COVID 19. В от
мейства Coronaviridae, как болезнетворные
дельном разделе мы остановимся на принципах
агенты, впервые выделены у человека в 60 х го
вакцинопрофилактики COVID 19 и рассмотрим
дах прошлого века. Четыре представителя коро
особенности как классических, так и инноваци
навирусов, инфицирующие человека (HCoV
онных вакцин, одобренных для практического
NL63, HCoV 229E, HCoV OC43 и HKU1), вы
применения.
зывают в большинстве случаев мягкие заболева
ния верхних дыхательных путей, но могут быть
причиной серьёзных инфекций у маленьких де
ОБЩАЯ ХАРАКТЕРИСТИКА
тей и пожилых людей [5]. Смертельные вспыш
КОРОНАВИРУСОВ
ки, причиной которых явились члены этого се
мейства, зарегистрированы гораздо позже. Вы
Семейство Coronaviridae включает вирусы,
сокопатогенный вирус SARS CoV (severe acute
поражающие разных хозяев, включая птиц и
respiratory syndrome coronavirus) стал причиной
млекопитающих. Это семейство относится к по
вспышки атипичной пневмонии в 2002 г., а в
рядку Nidovirales, и в нем выделяют 2 подсемей
2012 г. вирус MERS CoV (Middle East respiratory
ства. Подсемейство Orthocoronavirinae включает
syndrome coronavirus) привел к возникновению
все виды коронавирусов, инфицирующих чело
нового заболевания - так называемого ближне
века, и имеет 4 рода: Alphacoronavirus, Betacorona
восточного респираторного синдрома. Смерт
virus, Gammacoronavirus, Deltacoronavirus (https://
ность, по некоторым данным, составила 9,5% в
talk.ictvonline.org/taxonomy/). Род Betacoronavirus
случае атипичной пневмонии и 34,4% - в случае
включает 4 подрода (по более ранней класифи
MERS [6]. К счастью, оба заболевания распро
кации - линии): Embecovirus (линия A), Sarbeco
странились на относительно небольшом геогра
virus (линия B), Merbecovirus (линия C) и Nobeco
фическом ареале.
virus (линия D) [8]. Вирусы HCoV 229E и HCoV
В настоящем обзоре мы коснемся классифи
NL63 (род Alphacoronavirus), HCoV OC43 и
кации коронавирусов и отличий SARS CoV 2 от
HCoV HKU1 (род Betacoronavirus) могут вызы
других коронавирусов человека; дадим основ
вать у человека респираторные заболевания
ные характеристики всех его структурных ком
средней тяжести. Вирус SARS CoV 2 принадле
понентов. Основное внимание будет уделено
жит к роду Betacoronavirus и подроду Sarbecovirus.
структурному анализу шиповидного (спайково
К этому же подроду относится близкородствен
го, S) белка SARS CoV 2, который является
ный вирус SARS CoV (SARS CoV 1), в то время
главной иммуногенной мишенью и поэтому на
как вирус MERS CoV принадлежит к подро
ходится в центре внимания при разработке вак
ду Merbecovirus.
цин [7]. Будут рассмотрены его взаимодействия
Сферические или слабо полиморфные вири
с клеточными рецепторами, корецепторами и
оны коронавирусов диаметром 80-120 нм окру
антагонистами, описан гипотетический меха
жены липопротеиновой оболочкой и имеют ши
низм дестабилизации, запускающий слияние
повидные выступы на поверхности [9, 10], по
оболочки вириона с мембраной клетки при фи
форме напоминающие зубцы царской короны,
зиологическом значении рН и проникновение
что и дало семейству соответствующее назва
генома вируса в цитоплазму для его последую
ние (рис. 1, а). По морфологии вирионы коро
щей репликации.
навирусов сходны с вирионами вируса гриппа,
БИОХИМИЯ том 86 вып. 7 2021
966
КОРДЮКОВА и др.
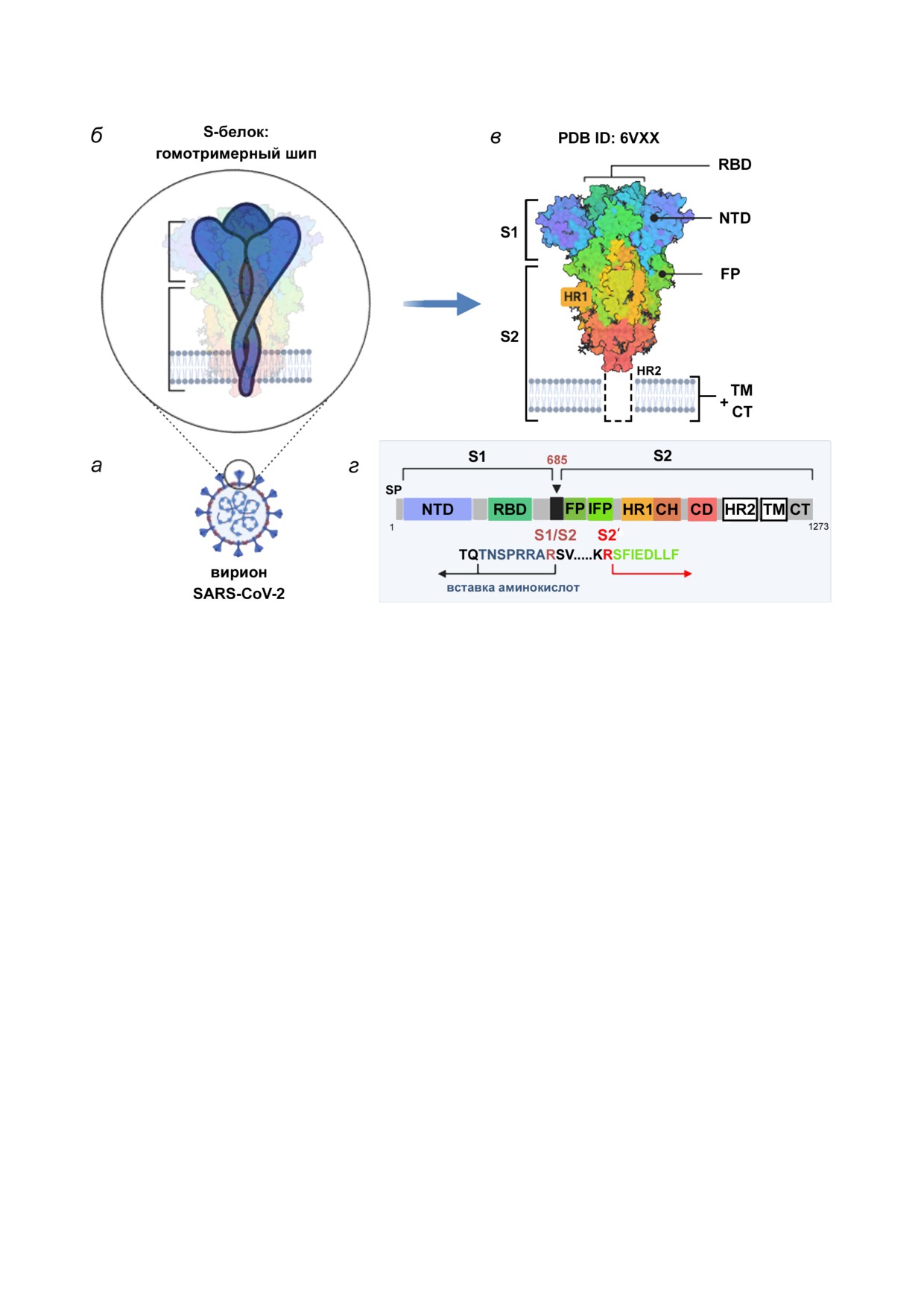
Рис. 1. Строение вириона SARS CoV 2 и структурные белки. а - Схема вириона с указанием локализации структурных
белков (S, N, M, E) и геномной одноцепочечной +РНК (ssRNA). Обозначения структурных белков: S - шиповидный бе
лок, E - белок оболочки, M - мембранный белок, N - нуклеокапсидный белок. б - Электрофоретический анализ в SDS
ПААГ очищенных вирионов. 1 - молекулярные веса белковых маркеров; 2 - мажорные структурные белки (S, его субъ
единицы S1, S2 после протеолитической активации; N). Адаптировано с разрешением из статьи Yao et al. [9]. Авторское
право Elsevier, 2020. (С цветными вариантами рисунков можно ознакомиться в электронной версии статьи на сайте:
но несут «шипы» только одного типа - гомотри
тропизма вируса; опосредует слияние мембраны
меры S белка [11, 12], в то время как два типа
вириона с мембраной клетки хозяина, обеспечи
шипов (гомотримеры гемагглютинина и гомо
вая вход вирусного генома в цитоплазму; участ
тетрамеры нейраминидазы (NA)) различают у
вует в сборке вирионов. На электрофореграмме
вирионов вирусов гриппа А и В [13, 14].
после разделения белков нативных вирионов в
Геном коронавирусов представляет собой од
денатурирующих условиях S белок представлен
нонитевую молекулу РНК позитивной поляр
частично в виде нерасщеплённого (неактивно
ности (+РНК) размером от ~27 до 32 Кб
го) S белка и в большей пропорции - в виде двух
(29,3 Кб - у SARS CoV 2). Это самый большой
его субъединиц (S1 и S2) (рис. 1, б). Эпитопы
и достаточно сложно организованный геном
S белка служат основными антигенами, кото
среди всех РНК содержащих вирусов (для срав
рые стимулируют образование нейтрализующих
нения геном вируса гриппа весит 13,5 Кб). РНК
антител и становятся важными мишенями для
коронавирусов кодирует 28-29 белков, в том
цитотоксических лимфоцитов.
числе 4 или 5 структурных белков. Вирусы чело
Белок M (мембранный) (~25-30 кДа) содер
века HCoV 229E, HCoV NL63, SARS CoV,
жит 3 ТМ домена [16, 17]. Он имеет небольшой
SARS CoV 2, MERS CoV имеют четыре струк
N концевой гликозилированный эктодомен и
турных белка: S - шиповидный белок (Spike),
гораздо более протяжённый С концевой эндо
E - белок оболочки (Envelope), M - мембран
домен, который уходит на 6-8 нм вглубь вирус
ный белок (Membrane), N - нуклеокапсидный
ной частицы [17, 18]. Мембранный протеин оп
белок (Nucleoprotein) (рис. 1, а). В составе вири
ределяет форму вириона и обеспечивает контакт
онов HCoV OC43 и HCoV HKU1 присутствует
между разными структурными белками при его
пятый структурный белок, HE - гемагглюти
сборке [16, 19]. Исследования с помощью крио
нин эстераза (Hemagglutinin Esterase).
электронной томографии (крио ЭТ) показали,
Белок S (шиповидный белок) коронавирусов с
что М белок существует в виде димера, может
молекулярной массой ~175 кДа [15] представля
принимать две разные конформации и способ
ет собой мембранный белок типа I, включаю
ствует искривлению мембраны, что важно при
щий большой гликозилированный N концевой
формировании дочерних вирионов.
эктодомен, один трансмембранный (ТМ) домен
Белок N (нуклеокапсидный) (~46-48 кДа) ас
и С концевой внутривирионный (цитоплазма
социирован с вирусной +РНК, образуя нуклео
тический, СТ) домен [11, 12]. S белок связыва
капсид. Этот белок может участвовать в регуля
ется с рецепторами на поверхности клеток и иг
ции синтеза РНК вируса и взаимодействует с
рает ключевую роль в определении тканевого
белком M при почковании вируса [16]. В N бел
БИОХИМИЯ том 86 вып. 7 2021
COVID 19: МИФЫ И РЕАЛЬНОСТЬ
967
ке предсказано несколько антигенных эпито
цию по прежнему остается спорным [27]. Неко
пов; он считается одним из важных антигенов
торые филогенетические исследования указы
коронавирусов [20].
вают, что промежуточным хозяином могут быть
Белок E (белок оболочки) (~8-12 кДа) транс
панголины. Так, показано, что рецептор связы
мембранный белок типа III, который имеет не
вающий мотив (RBM) шиповидного белка
большой N концевой эктодомен (~16 а.о.), один
SARS CoV 2 может быть получен путем реком
ТМ домен и СТ домен, включающий
бинации с вирусом панголина [28, 29].
~37-50 а.о. Этот белок сходен по функции с
Беспрецедентный анализ более 200 000 по
белком М2 вируса гриппа: он имеет випорино
следовательностей полных геномов SARS CoV 2
вую активность (образует пентамерный ионный
показал, что среди множества мутаций «очища
канал в липидной мембране), а также участвует
ющего» отбора (purifying selection) встречаются
в сборке вирусных частиц и их отпочковыва
несколько мутаций, которые можно объяснить
нии [16]. Три консервативных остатка цистеина
давлением положительного отбора (positive
S ацилированы, что важно для сборки вирио
selection). Это - замена 614G в шиповидном
нов [21-23]. Белки Е и М влияют на внутрикле
белке и несколько замен в нуклеокапсидном
точный транспорт белка S, его протеолиз и N
белке (в частности, 203К) [30]. Множественные
гликозилирование [24].
замены в N белке, по видимому, важны для
Белок HE (гемагглютинин эстераза) входит в
адаптации нового вируса SARS CoV 2 к челове
структуру только некоторых β коронавирусов, в
ку [30].
частности вирусов человека HCoV OC43 и
Интересны биоинформатические данные
HKU1, а также вирусов птиц рода Deltacorona
анализа так называемой внутренней неупорядо
virus. Часть HE белка связывается с нейрамино
ченности (intrinsic disorder) в N и M белках у
вой кислотой на поверхности клетки хозяина и,
разных коронавирусов [31]. Авторы показали,
вероятно, способствует начальной адсорбции
что SARS CoV 2 располагает удивительно
вируса мембраной. Эстераза отщепляет аце
«жёсткой» (состоящей из высокоупорядочен
тильные группы от нейраминовой кислоты. Ге
ных белков) защитной оболочкой. По видимо
ны, кодирующие HE белки коронавирусов,
му, из за таких характеристик вирион вируса
имеют гомологичные последовательности с ге
SARS CoV 2 очень устойчив в слюне и других
нами, кодирующими гликопротеин гемагглюти
жидкостях организма и во внешней среде [31].
нин эстераза фьюжен (HEF) вируса гриппа C.
Жизненный цикл коронавирусов начинается с
Геном SARS CoV 2 кодирует 16 неструктур
входа вируса в клетку. Предполагают два воз
ных белков (nsp1-nsp16) и 8 вспомогатель
можных пути: (1) слияние вирусной оболочки с
ных (accessory) белков, вовлечённых в биогенез
плазматической мембраной клетки (по видимо
новых вирусных частиц [25]. Среди них укажем
му, это основной путь); (2) эндоцитоз с последу
РНК зависимую РНК полимеразу (RdRp),
ющим слиянием вирусной оболочки с мембра
обеспечивающую репликацию вирусной РНК;
ной эндосомы. В обоих случаях рецепторную
две протеиназы - PLpro (papain like cysteine pro
функцию и функцию слияния мембран выпол
tease) и 3CLpro/Mpro (chemotrypsin like pro
няет белок S.
tease/main protease), отвечающие за аутолити
У вируса гриппа за вход в клетку отвечает ге
ческое расщепление вирусных полипротеинов
магглютинин, который, как и S белок корона
на функциональные фрагменты.
вирусов, является гликопротеином типа I. Гемаг
глютинин связывается с белками и липидами
клеточной поверхности, несущими сиаловые
ЭВОЛЮЦИЯ И ЖИЗНЕННЫЙ ЦИКЛ
кислоты. Известно, что перестройка HA, необ
КОРОНАВИРУСОВ
ходимая для слияния мембран, запускается в
эндолизосоме клетки при кислом значении рН.
Множественные выравнивания нуклеотидных
В отличие от вирусов гриппа у бетакоронавиру
последовательностей различных коронавирусов
сов слияние мембран происходит при физиоло
показывают, что наиболее близко родственны
гическом (нейтральном) значении рН. Соответ
ми для вируса SARS CoV 2 могут быть корона
ственно, триггером дестабилизации и пере
вирусы, инфицирующие летучую мышь. Однако
стройки S белка SARS CoV 2 должны быть
заражение человека вирусом летучей мыши на
иные факторы. Отдельные этапы, инициирую
прямую представляется маловероятным, по
щие этот процесс, и гипотетический механизм
скольку его связывание с человеческим рецеп
дестабилизации будут рассмотрены ниже.
тором ACE2 малоэффективно [26]. Вопрос о
Репликация коронавирусной РНК происхо
промежуточном хозяине при зоонозном заносе
дит в специализированных структурах вблизи
нового коронавируса в человеческую популя
ядра, включающих везикулы с двойной мембра
БИОХИМИЯ том 86 вып. 7 2021
968
КОРДЮКОВА и др.
ной, в которых при помощи крио ЭТ визуали
эпителии бронхов и экспрессируются в эндоте
зованы нити двуспиральной РНК [9, 32]. В зара
лии сосудов. На основании доступной структур
жённой клетке детектированы два типа органи
ной информации было постулировано, что два
зации рибонуклеопротеидных комплексов
аллеля ACE2 rs73635825 (S19P) и rs143936283
(РНП): шарообразные структуры, сгруппиро
(E329G) могут придавать устойчивость к инфек
ванные гексагонально по типу «яйца в гнезде»
ции SARS CoV 2 [35].
(один шар в центре, шесть вокруг него), либо в
Физиологическая функция ACE2 заключается
виде пирамиды из четырех шаров [9]. Статисти
в преобразовании ангиотензина I в ангиотензин
ческий анализ показал, что гексагональная и
(1 9) и ангиотензина II в ангиотензин (1 7). Ан
тетраэдральная упаковка соответствуют сфери
гиотензин (1 9) связывается с рецептором Mas,
ческим и эллипсоидным вирионам соответ
что приводит к расширению сосудов и противо
ственно. Отдельные шаровидные структуры со
воспалительному эффекту. Это действие проти
единены между собой более тонкими нитевид
воположно действию АСЕ, который превращает
ными перемычками. Такой способ упаковки
ангиотензин I в ангиотензин II. Ангиотензин II
при сохранении высокой стерической гибкости
вызывает сужение сосудов и провоспалитель
позволяет включить необычайно большой ге
ные эффекты через рецептор ангиотензина II
ном коронавируса во внутренний объём вирио
первого типа [33]. Можно предположить, что
на диаметром 80-100 нм [9, 32].
связывание вирусом молекул ACE2 будет сдви
Синтез N белка и растворимых неструктур
гать равновесие в сторону накопления ангио
ных белков коронавирусов происходит в цито
тензина II и, соответственно, сужения сосудов и
плазме, а синтез мембранных белков - на мемб
развития воспалительных реакций. Данные
ранах эндоплазматического ретикулума. Соб
процессы могут способствовать развитию тром
ранные в люменальных цистернах тримеры S
бозов, хотя точный механизм образования
белка, по всей видимости, участвуют в организа
тромбов в сосудах до сих пор неясен.
ции сайтов баддинга в промежуточном компарт
менте эндоплазматического ретикулума и аппа
рата Гольджи (ERGIC) [32]. Дочерние вирионы
СТРУКТУРНЫЕ ИССЛЕДОВАНИЯ
отпочковываются от цистерн ERGIC и далее
ШИПОВИДНОГО БЕЛКА SARS CoV 2
попадают во внеклеточную среду путём экзоци
тоза.
S белок SARS CoV 2 формирует гомотриме
Распознавание рецепторов является первым
ры по поверхности вирионов (рис. 2, a-в). Мо
этапом вирусной инфекции и ключевой детер
номер содержит 1273 а.о. (UniProt ID P0DTC2).
минантой тропизма клеток и тканей хозяина.
После протеолитического отщепления сигналь
Основным рецептором для SARS CoV 2, как и
ного пептида (SP) белок включает две функцио
для близко родственного вируса SARS CoV, яв
нальные субъединицы, S1 (а.о. 13-685) и S2
ляется белок ACE2 (angiotensin converting
(а.о. 686-1273).
enzyme 2); также в русскоязычной литературе
Дистальная субъединица S1 способствует ста
используется термин АПФ2 (ангиотензинпре
билизации префузионного (pre fusion, «до слия
вращающий фермент 2). АПФ2 - трансмемб
ния») состояния S2 субъединицы, заякоренной в
ранный белок типа I, фермент дипептидилкар
мембрану вириона. В то время как S1 отвечает за
боксипептидаза (КФ:3.4.17.23), который содер
рецепцию, S2 отвечает за слияние. Субъединица
жит около 805 а.о. и включает один цинк связы
S1 содержит N концевой домен (NTD) и С кон
вающий домен [33]. Полноразмерная молекула
цевой домен (CTD, он же рецептор связываю
ACE2 включает N концевой пептидазный до
щий домен, RBD, а.о. 319-541). NTD имеет то
мен и C концевой коллектрино подобный до
пологию галектинов человека - галактозосвя
мен, содержащий α спиральный ТМ домен и
зывающих лектинов [29]. RBD содержит две под
СТ домен (~40 а.о.).
области: основную структуру (5 цепочечный
Показано, что SARS CoV 2 связывается с
антипараллельный β лист) и собственно
ACE2 с 10-20 кратной аффинностью по сравне
RBM (а.о. 437-508), который определяет специ
нию с SARS CoV [11, 34]. Рецепторную функ
фичность связывания. Показано, что мотив
цию для коронавирусов выполняет пептидаз
(Gly Val Glu Gly) в составе RBM (а.о. 482-485)
ный домен, но пептидазная активность для это
обеспечивает более эффективный контакт
го не нужна
[33]. ACE2 преимущественно
RBD домена SARS CoV 2 с ACE2 по сравнению
экспрессируется в лёгких, сердце, почках, яич
с вирусом SARS CoV [37].
ках и желудочно кишечном тракте. В лёгких мо
Сайты активации S белка клеточными проте
лекулы ACE2 найдены в клетках альвеолярного
азами. Для обеспечения функции входа в клетку
эпителия типа II, они также присутствуют в
S белок должен быть активирован - расщеплён
БИОХИМИЯ том 86 вып. 7 2021
COVID 19: МИФЫ И РЕАЛЬНОСТЬ
969
Рис. 2. Структура шиповидного белка S. а - Схема вириона с «шипами» S белка; б - схема гомотримерного шипа, заяко
ренного в мембрану вириона; в
-
3D структура шипа в
«закрытой» конформации по данным крио
ЭМ (PDB ID 6VXX [12]); г - схема мономера S белка с указанием функциональных доменов и сайтов протеолитической
активации. S1 и S2 - субъединицы S белка; S1/S2 - фуриновый сайт расщепления; SP - сигнальный пептид; NTD - N
концевой домен; RBD - рецептор связывающий домен; FP - пептид слияния; IFP - «внутренний» пептид слияния, по
являющийся после расщепления S2 субъединицы в сайте S2′ [36]; HR1 и HR2 (Heptad Repeats 1 и 2) - специализирован
ные повторы в аминокислотной последовательности; TM - трансмембранный домен; CT - цитоплазматический домен.
Схема создана с использованием шаблона BioRender.com
протеазами на две субъединицы, S1 и S2, кото
ми, что видно по данным электрофоретического
рые в случае коронавирусов остаются нековалент
анализа, рис. 1, б). Это свойство значительно
но связанными. Наиболее заметная вариация,
повышает патогенный потенциал вируса, кото
отличающая аминокислотную последователь
рый приобретает способность инфицировать
ность S белка вируса SARS CoV 2 от шиповид
разные типы клеток в организме, помимо эпи
ного белка его «предшественника» (коронави
телия верхних дыхательных путей, где присут
руса летучих мышей BetaCoV/RaTG13/2013), а
ствует много экстраклеточных трипсин подоб
также от вируса SARS CoV, возникает из за
ных протеаз.
вставки дополнительных положительно заря
Поскольку фурин в больших количествах
женных аминокислотных остатков в сайт S1/S2
экспрессируется в лёгких, оболочечные вирусы,
(рис. 2, г) [12, 29, 36]. Вместо одиночного остат
инфицирующие дыхательные пути, могут ус
ка аргинина появляется сайт «(682)R R A
пешно использовать эту конвертазу для актива
R(685)↓», удобный для распознавания фурино
ции своего поверхностного гликопротеина. Фу
подобными протеазами, локализованными в ап
риновый сайт R X R/K R ранее обнаружен
парате Гольджи клетки [12, 36]. Гидролиз фури
между субъединицами HA1 и HA2 высокопато
ном происходит в процессе биосинтеза белка в
генных штаммов вируса гриппа А птиц и челове
клетке, и, таким образом, дочерние вирионы
ка подтипов Н5 и Н7 [38]. Полиосновные сайты
SARS CoV 2 получают пред активированный
протеолиза также найдены в поверхностных
S белок (но часть молекул в составе вириона
гликопротеинах некоторых вирусов семейства
могут остаться нерасщеплёнными - неактивны
Paramyxoviridae, инфицирующих человека (ви
БИОХИМИЯ том 86 вып. 7 2021
970
КОРДЮКОВА и др.
рус кори, паротита, респираторный синцити
10 ацилированных остатков цистеина (30 остат
альный вирус (RSV)), патогенного штамма ви
ков в гомотримере) в составе внутривирионного
руса болезни Ньюкасла (NDV); вирусов Эболы и
сегмента, включающего 39 а.о. [21] (рис. 3, а).
Марбург; вируса желтой лихорадки; HIV 1 и ря
Обнаружено, что остатки жирных кислот стаби
да опасных для человека вирусов из семейства
лизируют S белок. Кроме того, они организуют
Herpesviridae [38, 39].
домены со специфическим «рафтовым» соста
У всех коронавирусов S белок после связы
вом липидов, богатым сфингомиелинами и хо
вания с рецептором ACE2 расщепляется на
лестерином, уже в компартменте ERGIC, где
внутреннем участке S2′, расположенном непо
происходит созревание дочерних вирионов.
средственно перед последовательностью «S F I
В незаражённой клетке этот компартмент не со
E D L L F» - «внутренним» пептидом слияния
держит высоких концентраций холестери
[29, 36] (рис. 2, г). Расщепление в сайте S2′ явля
на [21]. Замена остатков цистеина в СТ домене
ется сигналом для необратимых конформаци
на остатки аланина приводит к драматическому
онных изменений S белка, промотирующих
изменению характеристик S белка и снижению
процесс слияния мембран. Протеолитическое
инфекционности вирусных частиц [21]. Наибо
расщепление осуществляет главным образом се
лее функционально значимыми оказались два
риновая мембранная протеиназа TMPRSS2
остатка цистеина, ближайшие к липидной
(transmembrane serine protease
2). Помимо
мембране. Предполагают, что их модификация
TMPRSS2, S белок вируса SARS CoV 2 может
облегчает навешивание остатков жирных кис
быть активирован и другими клеточными про
лот на удалённые от мембраны С концевые ос
теазами, в том числе катепсинами B и L (эндо
татки цистеина, поскольку её выполняет встро
сомальными цистеиновыми протеазами), фури
енный в мембрану фермент из семейства
ном, эластазой. Тем не менее считают, что толь
ZDHHC ацилтрансфераз [21]. Эта липидная
ко активность TMPRSS2 абсолютно необходима
модификация характерна и для других корона
для распространения вируса SARS CoV 2. Ранее
вирусов (число сайтов S ацилирования варьиру
была отмечена важная роль протеазы TMPRSS2
ет от 6 до 10). Есть данные о том, что модифика
в патогенезе вируса гриппа А [40].
ция жирными кислотами необходима для фор
Посттрансляционные модификации. Поверх
мирования полноценных вирионов [21, 45, 46].
ность гомотримерного шипа интенсивно глико
Ранее показано, что делеция всех трёх сайтов
зилирована [9, 11, 12, 41-43]. В молекуле S бел
ацилирования HA вируса гриппа А подавляет
ка SARS CoV 2 картированы 22 сайта N глико
репродукцию вируса, а удаление одного или
зилирования (66 сайтов в гомотримере), несу
двух сайтов, ближайших к С концу молекулы,
щие протяжённые углеводные цепочки, и 2 сай
приводит к его аттенуации [48]. Нашей группой
та O гликозилирования (T323/S325), которые
с помощью МАЛДИ времяпролётной масс
оказались на 99% немодифицированными в
спектрометрии было показано, что гемагглюти
природном белке [41, 42]. Состав сахаров в угле
нины трех типов вируса гриппа (А, В и С), а так
водных цепочках, связанных с N и О сайтами
же гликопротеины ряда других оболочечных ви
гликозилирования, определён с помощью масс
русов дифференциально S ацилированы двумя
спектрометрического анализа (LC MS/MS) [9,
типами высших жирных кислот: остаток стеара
43]. Он варьирует в зависимости от типа клеток,
та (С18:0) может присоединяться только к ос
где выращивали вирус, а также может разли
татку цистеина, расположенному на границе
чаться у природного и рекомбинантного бел
ТМ и СТ доменов, в то время как остатки цис
ков [9]. Разветвлённость цепочек оказалась даже
теина в СТ домене связывают исключительно
более высокой, чем было предсказано первона
остатки пальмитатов (С16:0) [49-52] (рис. 3, б).
чально [12]. Пять N связанных гликанов экспо
Сохраняется ли такая закономерность в случае
нированы в среду даже в постфузионной кон
S белка коронавирусов, еще предстоит опреде
формации шипа, которая не играет значитель
лить.
ную роль в «ускользании» вируса от иммунитета
3D структура «шипа» и его взаимодействие с
хозяина [44].
рецептором ACE2. На конец марта 2021 г. в базе
Помимо обильного гликозилирования, вли
данных PDB представлено более ста PDB entries
яющего на антигенные характеристики, S белок
3D структур S белка SARS CoV 2. Среди них -
содержит липидную модификацию, которая
структуры водорастворимого эктодомена, сво
вовлечена в процессы слияния мембран и сбор
бодного и в комплексе с рецептором ACE2;
ки вирионов - пальмитоилирование (S ацили
структуры полноразмерного рекомбинантного
рование) консервативных остатков цистеина ос
белка, а также структуры «шипов» на поверх
татками высших жирных кислот. Мономер S
ности вирионов. Детектированы несколько ва
белка вируса SARS CoV 2 содержит кластер из
риантов 3D структур префузионной конформа
БИОХИМИЯ том 86 вып. 7 2021
COVID 19: МИФЫ И РЕАЛЬНОСТЬ
971
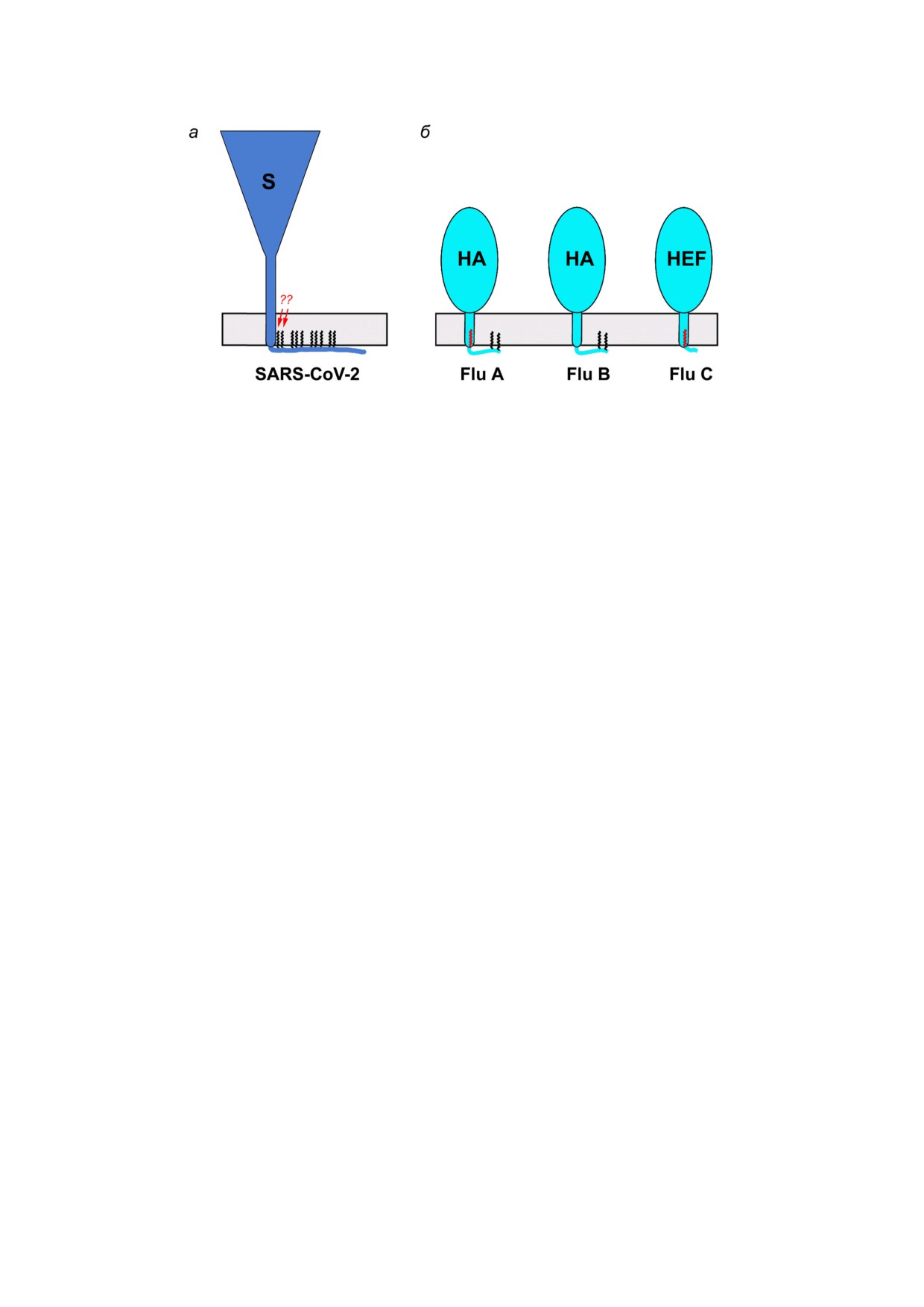
Рис. 3. Липидная модификация высшими жирными кислотами S белка SARS CoV 2 (а) и гемагглютининов вируса грип
па А, В и С (б). HA - гемагглютинин; HEF - гемагглютинин эстераза фьюжен. Остатки ковалентно связанных остатков
жирных кислот изображены как черные (пальмитаты, С16:0) либо красные (стеараты, С18:0) зигзагообразные линии для
одного мономера гомотримерного шипа. Стрелками указаны два остатка жирных кислот, которые гипотетически могут
связывать стеараты. Соотношения размеров HA и S приблизительно соблюдены (высота шипа S ~25 нм [32]; высота ши
па HA ~13,5 нм [47])
ции и одна постфузионная (post fusion, «после
один из RBD должен находиться в положе
слияния»). Для стабилизации префузионной
нии «стоя». Такой RBD может связаться с пеп
структуры изолированного S белка в S2 субъ
тидазным доменом молекулы ACE2. Это связы
единицу после последовательности HR1 вводят
вание инициирует «открывание» RBD соседнего
два остатка пролина в позициях 986 и 987, уда
мономера, его связывание со вторым ACE2 ре
ляют полиосновный сайт расщепления фури
цептором, а затем третьего RBD с третьим ре
ном, а последовательности ТМ и СТ доменов
цептором. Последовательные стадии связыва
заменяют на искусственный домен (foldon,
ния RBD доменов каждого из трех мономеров
trimerization domain) [11, 53].
шипа S белка с пептидазными доменами моле
Ключевой особенностью 3D структуры эк
кул ACE2 приводят в конечном итоге к «сбрасы
тодомена S белка в префузионной конформа
ванию» субъединиц S1, «раскрыванию» гомо
ции является подвижность его RBD домена, что
тримерного комплекса и экспозиции сайтов S2′
даёт возможность переключаться между «закры
в составе S2 субъединиц, до этого спрятанных в
той» конформацией шипа (RBD в положении
глубине шипа. Таким образом, появляется дос
«лежа»), имеющей значение для ускользания от
туп для мембранной протеазы TMPRSS2, кото
нейтрализующих антител [54], и «открытой»
рая расщепляет полипептидную цепь по этому
конформацией (один RBD в положении «стоя»),
сайту [55] и, как результат, высвобождается
инициирующей связывание с молекулой ACE2,
«внутренний» пептид слияния (рис. 2, г).
каскад перестроек S белка и, в конечном итоге,
Последующий структурный переход префу
слияние мембран. Разнообразие детектирован
зионной формы в постфузионную приводит к
ных префузионных структур особенно велико в
суперпозиции пептида слияния и ТМ домена на
работе, в которой анализировали смесь эктодо
одном конце длинной структуры, центрирован
мена S белка и водорастворимого фрагмента
ной вокруг трехспирального пучка [44]. Подроб
ACE2 [55]. Были описаны 9 различных конфор
но механизм слияния мембран при инфициро
маций гомотримерного шипа с RBD доменами
вании клеток SARS CoV 2 пока не изучен, но по
в разных положениях (до и после связывания с
аналогии с другими вирусами, использующими
рецептором ACE2), в том числе представлен эк
для входа в клетку фузионные белки I типа,
тодомен S белка с двумя открытыми/одним зак
можно предположить, что при встраивании
рытым RBD, хотя доля таких структур была не
пептидов слияния в мембрану мишень образу
высокой [55], а также детектирована S1 субъ
ется мембранная пора, которая последовательно
единица в комплексе с ACE2.
расширяется и пропускает вирусный нуклео
Предложена следующая гипотетическая схе
капсид в цитоплазму клетки [56].
ма взаимодействия гомотримерного шипа S
Корецепторы и антагонисты. Обнаружены и
белка на поверхности вириона с клеточными ре
другие связывающие детерминанты для глико
цепторами. Для инициации связывания хотя бы
протеина S на поверхности клетки, которые мо
БИОХИМИЯ том 86 вып. 7 2021
972
КОРДЮКОВА и др.
гут способствовать разработке эффективной
цептором ACE2 in vitro [58]. Интересно, что в
противовирусной терапии. Показано, что S бе
клетках человека добавка линолевой кислоты
лок вируса SARS CoV 2 взаимодействует с гепа
синергирует с препаратом ремдезивир [58].
рансульфатом посредством RBD, что способ
Топография шипов на поверхности вирионов.
ствует образованию «открытой» конформации
Формы шипов в конформациях до и после сли
шипа и, таким образом, усиливает его связыва
яния существенно различаются, что облегчает
ние с ACE2 [57]. Гепарансульфат можно позици
их распознавание на поверхности вирионов да
онировать в качестве корецептора («host attach
же при не очень высоком разрешении крио
ment factor»). Такой эффект не наблюдался в
ЭМ (рис. 4). В то время как в префузионном
случае других коронавирусов человека - SARS
состоянии шип имеет форму «цепа» (flail, пере
CoV 1 и MERS CoV [57].
вернутой пирамиды) (рис. 4, а), в постфузион
Антагонистом рецепции SARS CoV 2 на
ном состоянии он более узкий и имеет форму
клеточной мембране, как недавно было обнару
иглы (рис. 4, б).
жено, является незаменимая линолевая кисло
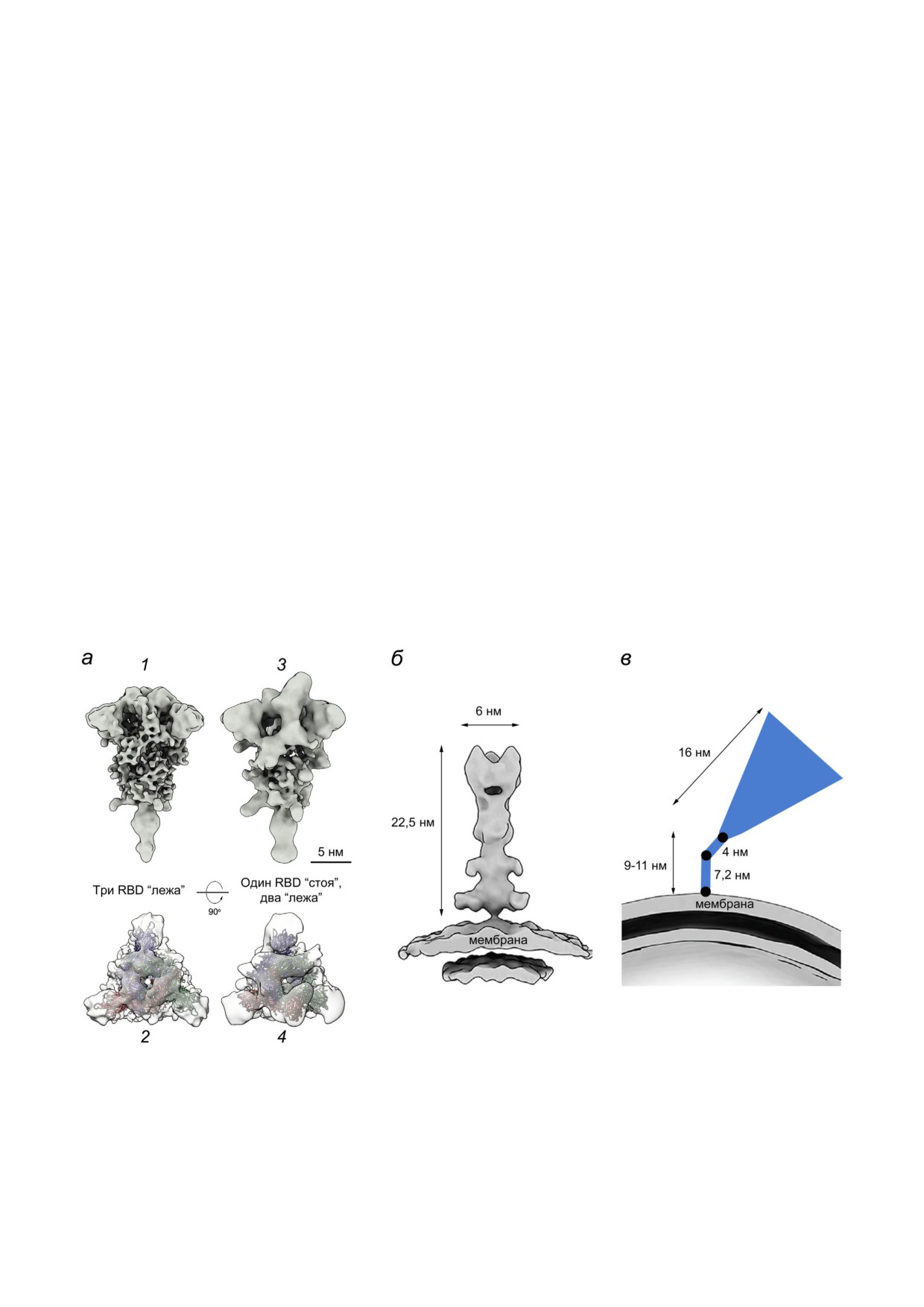
Вирионы несут в среднем 24 ± 9 тримеров S
та (linoleic acid, С18:2) [58]. С помощью крио
белка [10] (по другим оценкам: ~40 триме
электронной микроскопии (крио ЭМ) показано,
ров [41] и ~5-60 тримеров [9]). Приблизительно
что RBD домены акцептируют свободную ли
97% «шипов» на поверхности вирионов, фикси
нолевую кислоту в трех композитных связываю
рованных формалином (насыщенным раство
щих карманах. Примечательно, что в S белке,
ром формальдегида, сшивающего белки), нахо
связавшем линолевую кислоту, RBM упорядо
дились в конформации «пре фьюжен», а 3% - в
чен и спрятан на границе раздела между тремя
конформации «пост фьюжен» [10]. Эти кон
RBD доменами, тогда как в ранее описанных
формации не образовывали кластеры, а были
крио ЭМ структурах, в том числе в комплексе
распределены равномерно на поверхности ви
S белка с ACE2, он неупорядочен [11, 12]. Свя
рионов. Интересно, что после концентрирова
зывание линолевой кислоты стабилизирует
ния вирусных частиц по стандартному протоко
«закрытую» конформацию S белка, что приво
лу с применением ультрацентрифугирования на
дит к снижению взаимодействия вируса с ре
их поверхности детектировали шипы только в
Рис. 4. Крио ЭМ структуры гомотримерного шипа S белка на поверхности вирионов SARS CoV 2. а - Префузионная
конформация: 1 и 2 - «закрытая» («цеп»; три RBD в положении «лежа»); 3 и 4 - «открытая» (один RBD в положении
«стоя», два других - в положении «лежа»); показаны проекции сбоку (1, 3) и сверху (2, 4); б - постфузионная конформа
ция («игла»); в - схематичное изображение наклоненного шипа за счёт шарниров в тонком стебле (черные кружки) [41].
Размеры и масштабный штрих указаны согласно опубликованным ранее работам [9, 16, 41]. Адаптировано с разрешени
ем из статьи Yao et al. [9]. Авторское право Elsevier, 2020
БИОХИМИЯ том 86 вып. 7 2021
COVID 19: МИФЫ И РЕАЛЬНОСТЬ
973
закрытой префузионной конформации, но если
привела к возникновению новых генетических
содержащую вирус среду только осветляли при
вариантов вируса с увеличенным сродством
помощи низкоскоростного центрифугирова
к ACE2. На момент написания обзора таких ге
ния, то встречалась и закрытая, и открытая кон
нетических вариантов (variants of concern), вы
формации [10]. Вероятно, хрупкая открытая
зывающих серьезное беспокойство по поводу
конформация не выдерживает процедуры ультра
повышенной инфекционности и тяжести тече
центрифугирования и разрушается.
ния заболевания, зафиксировано несколько.
При инактивации вирионов β пропиолакто
Основные: (1) 202012/01 или 501Y.V1, так назы
ном, связывающим нуклеиновую кислоту виру
ваемый «британский» вариант, на 50% более
са, ситуация кардинально отличалась от опи
инфекционный; (2) 501Y.V2 - Южно Африкан
санной выше: доля шипов в префузионной кон
ский вариант, виновный в уменьшении эффек
формации составляла всего 24% против 76% в
тивности вакцин; (3) 501Y.V3 - Бразильский ва
конформации «пост фьюжен» [59]. Вирионы,
риант, имеющий профиль мутаций, близкий к
несущие на своей поверхности большое количе
Южно Африканскому варианту. Генетическая
ство шипов в постфузионной конформации,
линия B.1.351, к которой относится вариант
значительно ослаблены в плане возможности
501Y.V2, помимо мутации N501Y, несёт также
проникновения в клетку. Находясь в составе
мутации E484K и K417N. Все три мутации рас
инактивированных вакцин, такие вирионы,
положены в RBD [66]; а N501Y и E484K - в об
скорее всего, будут инициировать выработку так
ласти RBM и потенциально могут влиять на
называемых не нейтрализующих, либо субнейт
сродство вируса к ACE2 рецептору. Важной в
рализующих антител [60], не способных нейтра
контексте инфекционности вируса может быть
лизовать живой вирус. При наихудшем сцена
мутация P681H линии B.1.1.7, непосредственно
рии такие антитела могут способствовать разви
примыкающая к участку расщепления молеку
тию феномена антителозависимого усиления
инфекции (АЗУИ; в английской аббревиатуре -
preliminary genomic characterisation of an emergent
ADE, antibody dependent enhancement) при по
sars cov 2 lineage in the uk defined by a novel
падании нативного вируса в организм [59].
set of spike mutations/563). Проводятся иссле
Крио ЭТ анализ показал, что большинство
дования для понимания взаимосвязи возникаю
шипов S белка находятся под углом к мембране
щих мутаций с эффективностью вакцин. В част
вириона (в среднем угол составлял ~50° относи
ности, показано, что группа мутаций E484K +
тельно нормали к поверхности; иногда достигал
+ N501Y + D614G не понижает существенно
90°) [10, 41] (рис. 4 в). Такие наклоны во всех
способность антител, продуцируемых в резуль
направлениях могут обеспечиваться нескольки
тате действия вакцины BTN162b2, нейтрализо
ми достаточно гибкими шарнирами в области
вать вирус [67].
длинного (9-11 нм) и тонкого стебля шипа [41].
Подробный треккер вновь появляющихся
Высокая подвижность шипа должна помогать
мутаций и генетических вариантов SARS CoV 2,
вирусной частице «сканировать» поверхность
по результатам многочисленных данных, пред
клетки хозяина для поиска наиболее «подходя
щих» участков для прикрепления. Для сравне
nytimes.com/interactive/2021/health/coronavirus
ния ножка шипа HA вируса гриппа гораздо ко
variant tracker.html), а также в глобальной базе
роче (1,8-2 нм [47]). Подвижность шипа HA бы
филогенетических данных GISAID (www.
ла отмечена ранее [61, 62], но она менее выраже
gisaid.org). Анализ частоты быстроменяющихся
на по сравнению с шипом SARS CoV 2.
вариантов позволяет сделать предположение,
Мутационная изменчивость S белка. Возник
что вирус SARS CoV 2 в недалёком будущем
новение новых генетических линий и вариан
может приблизиться по разнообразию к сезон
тов вызывает наибольшее беспокойство, осо
ному гриппу, что не может не отразиться на
бенно при производстве вакцин, нацеленных
стратегии производства вакцин в сторону пре
на выработку иммунитета к определенной ан
обладания поливалентных.
тигенной детерминанте S белка. Так, мута
ция D614G, присутствующая во всех генетичес
ких линиях SARS CoV 2 [63] и приводящая к
ПАТОГЕНЕЗ COVID 19
исчезновению солевого мостика с остат
ком K854 [10], ведёт к увеличению скорости
Развитие COVID 19 у инфицированного че
репродукции вируса и заразности (увеличению
ловека можно разделить на три стадии [68]. Ста
показателя R0 - basic reproduction number) что,
дия I характеризуется первичным связыванием
вероятно, и привело к возникновению панде
вируса с ACE2 рецепторами клеток респиратор
мии [63-65]. Мутация N501Y в эпитопе S белка
ного эпителия в верхних дыхательных путях,
БИОХИМИЯ том 86 вып. 7 2021
974
КОРДЮКОВА и др.
участием протеазы TMPRSS2 в праймировании
Центральная парадигма иммунитета заклю
S белка; на этой стадии вирус может быть обна
чается в том, что опосредованные интерферона
ружен с помощью ПЦР теста в назальных смы
ми противовирусные реакции предшествуют
вах; детектируется низкая вирусная нагрузка и
провоспалительным, оптимизируя защиту хозя
слабый иммунный ответ. Стадия II характеризу
ина и сводя к минимуму коллатеральные по
ется высвобождением провоспалительных ци
вреждения [71]. По видимому, для COVID 19
токинов, интерферонов (IFN) β, γ, эпителиаль
эта парадигма неприменима. Исследуя времен
ными клетками. Во время стадии III отмечаются
ные паттерны IFN и воспалительных цитокинов
гипервоспаление, высокая вирусная нагрузка;
у 32 пациентов c COVID 19 средней и тяжёлой
вирус достигает альвеол в лёгких, наблюдается
степени, госпитализированных по поводу пнев
апоптоз эпителиальных клеток и в конечном
монии и наблюдаемых в связи с развитием ды
итоге развивается острый респираторный ди
хательной недостаточности, авторы обнаружи
стресс синдром (ОРДС) (acute respiratory distress
ли, что уровни IFN λ и IFN типа I одновремен
syndrome, ARDS), который характеризуется сис
но снижались, а их подъём откладывался и при
темной дисфункцией органов [68]. Наблюдается
этом индуцировался только у части пациентов,
снижение лимфоцитов в периферической кро
когда состояние больных становилось крити
ви - лимфопения [69].
ческим. Напротив, провоспалительные цитоки
Накопление фактических данных свидетель
ны, такие как TNF, IL 6 и IL 8, продуцирова
ствует о том, что несбалансированная и неконт
лись раньше интерферонов у всех пациентов и
ролируемая реакция врожденного иммунитета
сохранялись в течение длительного времени.
лежит в основе прогрессирования COVID 19.
Более высокие концентрации IFN λ у пациен
Эндотелий легочных капилляров после его ак
тов с COVID 19 коррелировали с более низкой
тивации начинает экспрессировать цитокины и
вирусной нагрузкой в бронхиальных аспиратах
молекулы сосудистой адгезии, которые могут
и более быстрым вирусным клиренсом. Для
усилить цитокиновый шторм и способствовать
сравнения у 16 больных гриппом, госпитализи
тромбозу сосудов. Среди провоспалительных
рованных по поводу пневмонии со сходными
цитокинов, гиперпродукция которых может
клинико патологическими характеристиками,
привести к дисфункции эндотелия, следует упо
как у пациентов с COVID 19, и у 24 не госпита
мянуть интерлейкин 6 (IL 6) и фактор некроза
лизированных пациентов, имеющих более лёг
опухоли (TNF) [1]. IL 6 способствует повыше
кие симптомы гриппа, IFN λ и IFN I типа были
нию проницаемости сосудов и секреции про
достоверно индуцированы ранее на более высо
воспалительных цитокинов самими эндотели
ких уровнях и независимо от тяжести заболева
альными клетками, тем самым усиливая высво
ния, в то время как провоспалительные цитоки
бождение цитокинов [1].
ны вырабатывались только в острой стадии. Из
В тяжёлых случаях COVID 19 массивная эн
мененные цитокиновые паттерны у пациентов с
дотелиальная дисфункция, обширная коагуло
COVID 19 коррелировали с более длительной
патия и тромбоз, вызванный системой компле
госпитализацией и более высокой частотой тя
мента, могут привести к развитию системной
жёлого заболевания и смертности по сравнению
микроангиопатии и тромбоэмболии; эти ослож
с гриппом. Эти данные указывают на разбалан
нения опасны для жизни и в конечном итоге мо
сированный противовирусный ответ у больных
гут привести к полиорганной недостаточности,
COVID 19, способствующий длительному при
включая миокардиты, сердечную недостаточ
сутствию вируса в организме, гипервоспалению
ность, отёки лёгочной ткани, гипоксию и пора
и дыхательной недостаточности [72].
жения почек [1]. Нарушение функции почек
Целый спектр тяжёлых патологических со
связано со значительно повышенным риском
стояний, таких как двусторонняя пневмония,
смерти у тяжелобольных пациентов [1]. Проте
поражения сердца и печени был ранее описан
канию COVID 19 с осложнениями могут способ
для вспышек заболевания гриппом A/H5N1
ствовать такие заболевания, как диабет и хрони
и A/H7N9 и во время пандемии A/H1N1 в
ческая обструктивная болезнь лёгких (ХОБЛ),
2009 году [14]. Новые данные указывают на то,
при которой наблюдается повышенная экспрес
что дисфункция эндотелия, индуцированная
сия рецепторов ACE2 [68]. Факторами риска
инфекцией SARS CoV 2, отличается от воздей
считают пожилой возраст и гиповитами
ствия вируса гриппа А/H1N1 пандемических
ноз D [70]. Работы на мышах указывают на гор
штаммов 2009 года. В случае COVID 19 развива
монально модулированную повышенную экс
ется пироптоз, приводящий к гибели эндотели
прессию ACE2 у самцов [70], что может указы
альных клеток и способствующий усилению
вать на возможность более тяжёлого течения
провоспалительных стимулов и тромбогенных
COVID 19 у мужчин.
событий [73]. Очевидно, что клиническая кар
БИОХИМИЯ том 86 вып. 7 2021
COVID 19: МИФЫ И РЕАЛЬНОСТЬ
975
тина тяжёлых форм гриппа отличается от разви
(Talampicillin, Lurasidone, Rubitecan, Loprazolam)
тия тяжёлого COVID 19 (хотя прослеживаются
[82]; триазавирин (Triazavirin) обладает проти
некоторые параллели, такие, как запуск цитоки
вовирусной активностью, подавляет Mpro
нового шторма). При сезонном гриппе тяжёлых
[83-85]. Изучается воздействие на SARS CoV 2
дисфункций иммунитета, как правило, не на
препаратов, заявленных как противовирусные
блюдается.
(Favipiravir, Ivermectin, Ribavirin) [64]; другие
Морфометрические исследования показали,
средства, на которые ранее возлагались надеж
что стенки лёгочных артерий у больных COVID 19
ды (Lopinavir, Ritonavir, Hydroxychloroquine), в
в два раза толще (а просвет сосудов, соответ
настоящий момент считаются неэффективными
ственно, тоньше), чем у больных пандемичес
и ушли из рассмотрения.
ким штаммом вируса гриппа H1N1 2009 года
Авторы ряда фундаментальных исследований
[74]. Появляются данные, что именно S белок
предлагают перспективные антивирусные стра
может запускать сигнальные каскады, разруша
тегии, которые пока позиционируются как идеи.
юще действующие на клетки [74], и способство
Так, с целью подавления слияния вирусной и
вать развитию тяжёлого патологического состо
клеточной мембран предлагают использовать
яния, называемого «гипертонией лёгочных ар
синтетический липопептид с аминокислотной
терий» (pulmonary arterial hypertension) [75].
последовательностью HR2 домена, который
Учитывая, что S белок SARS CoV 2 является
блокирует конформационную перестройку S
основным антигенным компонентом вакцин,
белка [86]; нокдаун ацилтрансферазы ZDHHC20,
крайне важно, по мнению авторов, провести
ацилирующей S белок, должен уменьшать ре
дальнейшие эксперименты для изучения его
продукцию вирусных частиц и их инфекцион
возможного воздействия на клетки лёгочных ар
ность [21]. Для ограничения инфекции назнача
терий и других тканей/органов, таких как сис
ют препараты интерферонов типа I; также с
темные сосуды, сердце и мозг [75].
целью снижения возможных побочных эффек
тов от их прямого введения предложена страте
гия активации так называемых интерферон
ТЕРАПИЯ COVID 19
стимулируемых генов (interferon stimulated
genes, ISGs), кодирующих, в частности, IFITM
Потенциальные антивирусные препараты и те
(IFN induced transmembrane proteins) или
рапевтические антитела. Антивирусные препара
CH25H (сholesterol 25 hydroxylase) [87].
ты, направленные на подавление других вирус
Новые терапевтические возможности от
ных инфекций, гипотетически могут быть пе
крывает факт ингибирования связывания S бел
репрофилированы для борьбы с COVID
ка с ACE2 экзогенным гепарином и его произ
19 [76, 77]. Ключевыми белковыми компонента
водными, не являющимися антикоагулянта
ми SARS CoV 2, по отношению к которым бы
ми [57]. Антагонисты связывания вируса с гепа
ли предприняты попытки подобрать антивирус
рансульфатом на поверхности клетки предлага
ный препарат, являются S белок, РНК зависи
ется использовать в комбинированной противо
мая РНК полимераза (RdRp) и вирусная проте
вирусной терапии [57, 88, 89].
аза Mpro. Наиболее известен препарат ремдеси
Нейтрализующие антитела были одними из
вир (Remdesivir) - синтетический аналог адено
первых противовирусных средств, использован
зина, который связывается с RdRp, блокируя
ных во время пандемии COVID 19 [12, 90, 91].
синтез вирусной РНК [78]. Препарат производ
Моноклональные антитела, выделенные от выз
ства компании «Gilead Sciences» (США) не по
доравливающего пациента, проявили нейтрали
казал каких либо заметных отклонений от пла
зующие способности, блокируя контакт между
цебо при лечении болезни средней и тяжёлой
RBD S белка и клеточным рецептором ACE2
степени [79], однако есть данные, подтверждаю
[91]. Недостатком терапии антителами может
щие, что применение ремдесивира способствует
быть возникновение мутаций в RBD [92]. Ис
улучшению на ранних стадиях COVID 19; пре
пользование взаимодополняющих антител, уз
парат разрешён для экстренного применения в
нающих разные эпитопы RBD, повышает бло
кирующий эффект при клиническом примене
nih.gov/therapeutic management/).
нии и минимизирует ускользание вируса от им
Препараты камистад мезилат (Camostat
мунного ответа [91].
mesylate) [80] и нафамистад (Nafamostat) [81]
Купирование патогенетических процессов.
ингибируют мембранную протеазу TMPRSS2.
Спустя год от начала пандемии можно конста
Сходный потенциал воздействия на протеазы
тировать, что большинство противовирусных
имеют коммерчески доступные препараты та
препаратов пока не смогли улучшить клиничес
лампициллин, лурасидон, рубитек, лопразолам
кие исходы при тяжёлом COVID 19; напротив,
БИОХИМИЯ том 86 вып. 7 2021
976
КОРДЮКОВА и др.
терапевтические вмешательства, нацеленные на
ют во внутриклеточной мРНК: КЭП структу
патологические реакции организма пациента,
ра (7 methyl guanosine); 5′ UTR; участок, коди
включающие гипериммунный ответ, активацию
рующий целевой белок (в случае COVID 19 - S
комплемента и системный тромбоз, оказались
белок вируса SARS CoV 2); 3′ UTR и участок
более перспективными [1]. Некоторые препара
полиаденирования. С момента попадания РНК
ты используются в клинической практике в Рос
вакцины в клетки организма и до формирова
сии.
ния иммунного ответа происходит множество
Дексаметазон (Dexamethasone) - синтетичес
сигнальных каскадов и событий, которые мож
кий глюкокортикостероидный препарат, ис
но разделить на три типа, каждый из которых
пользуемый для лечения ревматических заболе
требует определенного подхода и решений по
ваний, ряда кожных заболеваний, тяжёлых ал
его успешной реализации: (1) доставка РНК в
лергических состояний, астмы, ХОБЛ, отёка
клетки и преодоление клеточного барьера,
мозга, а также вместе с антибиотиками при ту
(2) индукция интерферонов или самоадъювант
беркулёзе [2, 64].
ный эффект и (3) процессирование антигенов
Артлегиа (Олокизумаб, Olokizumab) - препа
для систем комплекса гистосовместимости
рат производства компании «Р Фарма» (Рос
классов I и II (MHC I и MHC II).
сия), моноклональное антитело, непосред
На первом этапе прохождения мРНК через
ственно ингибирующее интерлейкин 6 (IL 6) и
плазматическую мембрану основную роль игра
первоначально разработанное компанией «UCB
ют системы защиты РНК от деградации РНКа
Pharma» (Бельгия) для терапии ревматоидного
зами в межклеточном пространстве и кровото
артрита [93].
ке, а также эффективность самой доставки.
Противовоспалительные антитела в сочета
РНК является отрицательно заряженным поли
нии с молекулами антикоагулянтов используют
анионом и не может пассивно диффундировать
для ограничения коагулопатий и разрушитель
через мембрану. Для преодоления этого барьера
ной цитокиновой сигнализации, запускаемой
необходим активный транспорт. Для его реали
SARS CoV 2 [94].
зации предложена доставка в виде липоплекса -
комплекса нуклеиновых кислот и липидов, по
падающего в клетку путем эндоцитоза [96-100].
ПРИНЦИПЫ
Липидные составы, которые были протестиро
ВАКЦИНОПРОФИЛАКТИКИ
ваны на доставку мРНК, обычно состоят из ка
тионного или ионизируемого липида и липидов
Вакцинопрофилактика на сегодняшний
для стабилизации структуры, таких как фосфо
день является единственной мерой по сдержи
липиды, холестерин или полиэтиленгликоль
ванию распространения коронавирусной ин
липид (ПЭГ липид). Катионные липиды обыч
фекции. По состоянию на февраль 2021 года в
но включают для обеспечения образования
мире насчитывалось около 289 кандидатных
комплексов с отрицательно заряженными моле
вакцин против SARS CoV 2, около 70 из кото
кулами мРНК, и их можно разделить в соответ
рых находятся на той или иной стадии клини
ствии с pKa аминогруппы на липиды посто
янного заряда, такие как DOTMA (1,2 ди O ок
blueprint/covid 19). В настоящий момент мы яв
тедеценил 3 триметиламинопропан), DOTAP
ляемся свидетелями новой технологической ре
(1,2 диолеилокси 3 триметиламинопропан),
волюции вакцинопроизводства, которая рас
DC холестерин (холестерил 3 бета карбокси
пространяется по всему миру [95]. Среди разре
амидоэтилен N диметиламин) и pH зависимые
шенных к применению вакцин против SARS
ионизируемые липиды. Последние в условиях
CoV 2 есть как традиционные инактивирован
низкого значения рН имеют положительный за
ные и рекомбинантные, так и инновационные,
ряд, что облегчает образование комплекса с мо
ранее не применявшиеся вакцины, основанные
лекулами РНК. В условиях физиологическо
на доставке генетического материала, кодирую
го pH заряд комплексов варьирует от нейтраль
щего целевой антиген [53]. Принцип использо
ного до умеренно катионного, что снижает
вания ДНК или РНК в качестве вакцин копиру
их токсичность, препятствует неспецифичес
ет естественный процесс транскрипции, транс
ким взаимодействиям с белками сыворотки
ляции и экспрессии протеинов клетки.
крови и продлевает время циркуляции в крови
Вакцины на основе РНК. За годы исследова
[101, 102].
ний по созданию эффективной РНК вакцины
Технология липофекции при внутрикожных
были опробованы различные варианты РНК,
инъекциях описана во множестве статей, в том
сохраняющие при этом структуру из нескольких
числе с использованием мРНК против вирусов
базовых элементов, которые также присутству
гриппа A/H10N8 и A/H7N9, для лечения диабе
БИОХИМИЯ том 86 вып. 7 2021
COVID 19: МИФЫ И РЕАЛЬНОСТЬ
977
та II го типа, против вируса Денге [103-105].
Основа этой технологии - получение липидных
наночастиц размером 80-120 нм. Структура
частиц липоплекса разнообразна и варьирует от
ламеллярной до сферической и гексагональной,
сохраняя при этом распределение нитей
ДНК/РНК и бислоёв липидов (рис. 5). Пионе
ром в производстве новой вакцины против ви
руса SARS CoV 2 на основе мРНК является
американская компания «Moderna», которая
уже создавала прототипы мРНК вакцин против
вируса Zika [98, 106], вируса гриппа [107] и
Рис. 5. Варианты структуры липоплекса. а - Многослой
RSV [108]. Для создания липоплекса липиды
ная ламеллярная структура; б - инвертированная гексаго
растворяют в спирте в соответствующих моляр
нальная структура. Адаптировано с разрешением из обзора
ных соотношениях и смешивают с РНК в буфе
Ewert et al. [109]. Авторское право Taylor & Francis, 2005
ре с низким значением pH (~4,0) с использова
нием миксера для микрофлюидики [98].
Основными мишенями при вакцинирова
ей «BioNTech» и американской компанией
нии должны стать антиген презентирующие
«Pfizer». По результатам клинических исследо
клетки, например дендритные клетки (ДК), т.к.
ваний, в которых приняло участие 43 998 паци
они являются связующим звеном между вос
ентов, эффективность вакцины BNT162b2
приятием антигенов и выработкой Т клеточно
составила около
95%
(«ClinicalTrials.Gov/
го и В клеточного иммунного ответа. Репрезен
NCT04368728, Pfizer/BioNTech»). Второй мРНК
тация антигенов дендритными клетками может
вакциной, разработанной компанией «Moderna»
происходить либо из цитоплазмы, что включает
совместно с Национальным Институтом Здра
презентацию антигенных пептидов в молекулах
воохранения NIH, является вакцина mRNA
главного комплекса гистосовместимости клас
1273. По результатам фазы III клинических ис
са I, либо из лизосом, которые несут фрагменты
пытаний, в которой участвовало 28 207 пациен
антигенов, захваченных с помощью эндоцито
тов, эффективность новой вакцины составила
за, - презентация MHC класса II. МНС I и II
94,5% («ClinicalTrials.Gov/NCT04470427, Moderna»).
классов затем распознаются рецепторами Т
Обе РНК вакцины являются двухдозовыми,
клеток CD8+ или CD4+ соответственно.
вводятся внутримышечно с промежутком 3-4
Главным триггером в череде иммуномодули
недели, температура транспортировки от -25 °C
рующих эффектов РНК вакцины после её попа
до -15 °C.
дания в клетку является индукция интерферо
Вакцины на основе аденовирусов (Ad vec
нов или самоадъювантный эффект, который, в
tored). С момента открытия возможности при
зависимости от последствий, может иметь двой
менения аденовирусов как генно инженерного
ственное значение. В целом острые реакции
средства в начале 90 х годов прошлого столетия,
IFN типа I опосредуют плейотропные и воспа
модифицированные аденовирусы широко при
лительные эффекты как при врождённом, так и
меняются при разработке эффективного спосо
при адаптивном иммунных ответах [110, 111].
ба внутриклеточной доставки генного материа
Известно, что передача сигналов IFN типа I ин
ла и создании прототипов вакцин [115]. В геном
дуцирует созревание ДК, улучшает процессинг
аденовируса с помощью методов молекулярного
и презентацию антигена и усиливает миграцию
клонирования встраивают участок ДНК, коди
ДК в зоны трансфецированных клеток [112], но
рующий целевой протеин, для выработки им
также может привести к тяжёлым аутоиммун
мунного ответа (в случае COVID 19 - это S бе
ным последствиям, таким как системная крас
лок вируса SARS CoV 2). Для этих целей ис
ная волчанка и диабет I го типа [113, 114]. Об
пользуют репликационно дефектные модифи
ход этой проблемы завязан на структуре мРНК,
цированные аденовирусы (векторы), основой
которая используется в качестве вакцины и яв
которых являются наименее распространен
ляется одним из ключевых элементов в балансе
ные (rare species) или отсутствующие в челове
уровня экспрессии целевых протеинов и опо
ческой популяции типы, например вирус чело
средованного IFN воспаления и аутоиммунной
века Ad26 или вирусы приматов (chimpanzees or
реакции.
non human primates), для обхода ранее приобре
Первой мРНК вакциной против новой ко
тенного иммунитета к аденовирусам. При ис
ронавирусной инфекции стала вакцина
пользовании аденовирусных векторов целевой
BNT162b2, разработанная немецкой компани
протеин экспрессируется в клетках организма в
3 БИОХИМИЯ том 86 вып. 7 2021
978
КОРДЮКОВА и др.
среднем две недели с пиком на второй-третий
COVID 19, а также хорошую переносимость у
день, что вполне достаточно для выработки им
более чем 16 500 участников клинических испы
мунного ответа [116].
таний [120].
Современные репликационно дефектные
Компания «AstraZeneka» совместно с Окс
аденовирусы могут размножаться только на спе
фордским университетом разработала и выпус
циальном клеточном субстрате, в специальных
тила на рынок вакцину AZD1222 на основе аде
клетках линий НЕК 293 или PER.C6 (packaging
новирусного вектора шимпанзе ChAdOx1. По
cell lines), которые несут всю 5′ концевую часть
результатам фазы III расширенных клинических
генома, включая E1, ITR, цис действующие по
исследований вакцина эффективна на 79% про
следовательности упаковки и последователь
тив заражения SARS CoV 2 и на 100% - против
ности белка IX. Существование аденовирусных
развития тяжёлых форм COVID 19 (https://
векторов в виде молекулярных клонов в плазми
clinicaltrials.gov/ct2/show/NCT04516746?term=
дах позволяет манипулировать с конструкцией,
NCT04516746&draw=2&rank=1). Однодозовые
а также наращивать их в клетках Escherichia coli,
вакцины разработаны и выпущены компани
а использование в конструкции сильного цито
ей «Janssen Biotech, Inc.» на базе вектора Ad26 c
мегаловирусного промотора позволяет достичь
эффективностью от 61% до 72% (в зависимости
значительного уровня экспрессии целевых ге
от страны, где проводились клинические иссле
нов по сравнению с другими генно инженерны
дования) и компанией «CanSino Biologics» на
ми способами внутриклеточной доставки коди
базе аденовирусного вектора Ad5 с эффектив
рующего материала [117].
ностью 65,28%.
После попадания в клетку хозяина аденови
Инактивированные цельновирионные и рас
русная частица распаковывается, а ДНК вируса
щéпленные вакцины. Ещё одним типом вакцин
посредством транспорта по микротрубочкам
против SARS CoV 2, разработанных и одобрен
попадает в ядро, где служит матрицей для само
ных региональными регуляторами, является
воспроизведения. Поскольку жизненный цикл
инактивированная цельновирионная или (как
аденовируса дикого типа является внехромо
усовершенствованный вариант) расщéпленная
сомным, Ad векторы, по умолчанию, воспри
вакцина, в которой инактивированные вирио
нимаются как неинтегрирующие. Однако ряд
ны вируса дополнительно разрушаются детер
модельных экспериментов на мышах показал
гентами, например Triton X 100. Несколько пре
возможность рекомбинации между вектором и
паратов производства компаний «Sinopharm»,
хромосомной ДНК [118]. Это направление ис
«Sinovac» и ФНЦ исследований и разработки
следований должно быть продолжено для изуче
иммунобиологических препаратов им. М.П. Чу
ния долгосрочных эффектов и биобезопасности
макова РАН разрешены для иммунизации огра
векторных вакцин.
ниченного контингента людей. Несмотря на
Особое внимание исследователей приковано
незначительные различия в технологическом
к иммунному ответу, приобретаемому к самой
процессе, основное внимание при производстве
аденовирусной частице. Существуют многочис
данного типа вакцин должно быть уделено кон
ленные данные, подтверждающие значительное
формации S белка в итоговом препарате в связи
падение выработки антител на целевой белок
с инактивацией живого вируса β пропиолакто
при повторной иммунизации одним и тем же
ном, о чём уже упомянуто выше. Так как некон
вектором [119]. Данное явление ставит серьёз
тролируемое применение инактивирующих
ные препоны для аденовирусных векторов при
агентов может в конечном счёте привести к вы
разработке стратегии вакцинации, особенно в
работке не нейтрализующих антител.
свете перспективы становления COVID 19 се
Пептидные вакцины. Первая синтетическая
зонным заболеванием с чередой сменяемости
пептидная вакцина «ЭпиВакКорона» против
активно циркулирующих вариантов.
нового коронавируса, которая представляет со
К моменту написания обзора в мире уже за
бой суспензию для внутримышечного введения,
регистрировано и выпущено на рынок несколь
содержащую композицию химически синтези
ко вакцин на основе аденовирусной доставки
рованных пептидных иммуногенов S белка
генного материала. В основе двухдозовой вак
SARS CoV 2, конъюгированных с белком но
цины «Спутник V», зарегистрированной в Рос
сителем и адсорбированых на гидроксиде алю
сии как «Гам КОВИД Вак», лежит гетерологич
миния, была представлена ФБУН ГНЦ ВБ
ный векторный подход к доставке целевого ге
«Вектор» Роспотребнадзора. Первые данные об
на, кодирующего S белок SARS CoV 2, на ос
этой вакцине недавно были опубликова
нове двух типов аденовируса: Ad26 и Ad5. По ре
ны [121]. В настоящее время проводятся I-II
зультатам фазы III клинических испытаний вак
фазы клинических испытаний вакцины. Пози
цина показала эффективность 91,6% против
ционируется, что вакцина «ЭпиВакКорона» на
БИОХИМИЯ том 86 вып. 7 2021
COVID 19: МИФЫ И РЕАЛЬНОСТЬ
979
основе пептидов имеет низкую реактогенность,
ем на организм и возможностями терапии требу
является иммуногенным и безопасным продук
ют дальнейшего изучения.
том. Дальнейшие исследования необходимы для
Пандемия COVID 19 внесла столько нового
оценки её эффективности.
в сферу глобального здравоохранения, сколько
никакая угроза до этого. Введены в практику но
вые типы вакцин, а само вакцинирование 7 мил
ЗАКЛЮЧЕНИЕ
лиардов человек является до сих пор невидан
ным вызовом человечеству по координации сов
Год назад внезапно возникшая и стремитель
местных усилий. Один из ключевых вопросов на
но развивающаяся пандемия COVID 19 породи
будущее заключается в том, насколько силен Т
ла много домыслов и слухов относительно пато
клеточный иммунитет у переболевших COVID
генности нового коронавируса и его происхож
19 и у вакцинированных пациентов. Данные о
дения. В настоящий момент специалисты пола
долгосрочных эффектах вакцинации пока нахо
гают, что вероятность появления SARS CoV 2 в
дятся на стадии накопления. Однако ранние
результате лабораторных манипуляций с
данные от пациентов, перенёсших новое коро
родственным коронавирусом ничтожно ма
навирусное заболевание, являются многообеща
ла [122]. Генетические данные неопровержимо
ющими. Они демонстрируют высокие показате
показывают, что SARS CoV 2 не является произ
ли памяти как у CD4+, так и CD8+ Т клеток по
водным от какого либо ранее использовавшего
отношению к нескольким белкам SARS CoV 2,
ся вирусного остова. К такому же выводу скло
включая не только S белок, но и нуклеопротеин
няются эксперты ВОЗ, опубликовавшие 30 мар
и мембранный белок [123]. Дальнейший мони
та 2021 г. отчетный доклад о результатах рассле
торинг выздоровевших пациентов должен дать
дования специальной комиссии в Китае (https://
представление о защитной способности иммун
ной памяти - гуморальной и клеточной. Имеет
global study of origins of sars cov 2 china part).
важное значение и понимание того, как форми
Основной гипотезой появления вспышки ново
руется Т клеточная память у пациентов, кото
го коронавирусного заболевания, лежавшей в
рые испытывают лёгкие симптомы COVID 19
основе пандемии, считают зооноз - перенос ви
по сравнению с тяжёлой болезнью.
руса от животного к человеку.
Заканчивая обзор на позитивной ноте, хо
За истекший 2020 год накоплен огромный
чется надеяться, что мутационная изменчивость
экспериментальный материал относительно ши
в будущем приведёт к ослаблению нового коро
повидного белка SARS CoV 2. Разнообразие по
навируса, и тогда по аналогии с гриппом вероят
лученных 3D структур позволяет выделить не
ны регулярные сезонные вспышки коронави
сколько ключевых особенностей S белка, опреде
русного заболевания с более лёгким течением, с
ляющих специфику патогенеза вируса: (1) «зак
которыми человечество сможет справляться без
рытая» префузионная конформация шипа может
таких серьёзных потерь, какие сопровождали
быть использована вирусом для ускользания от
сезон 2020-2021.
иммунитета; (2) высокое сродство связывания
RBD в «открытой» конформации с рецепто
ром ACE2 человека важно для эффективной ре
Финансирование. Работа выполнена при фи
цепции на клеточной поверхности; (3) предва
нансовой поддержке Российского фонда фунда
рительная активация (прайминг) S белка фури
ментальных исследований (грант № 20 14
ноподобными протеазами позволяет вирусу
50441).
распространяться по разным типам клеток. Эти
Конфликт интересов. Авторы заявляют об от
знания заложили основу как для поиска антиви
сутствии конфликта интересов.
русных препаратов, так и для конструирования
Соблюдение этических норм. Настоящая
вакцин. Тем не менее очень многие аспекты,
статья не содержит описания каких либо иссле
связанные с механизмами репликации вируса,
дований с участием людей или животных в каче
его разрушительным патогенетическим действи
стве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Perico, L., Benigni, A., Casiraghi, F., Ng, L. F. P., Renia, L.,
2.
Asselah, T., Durantel, D., Pasmant, E., Lau, G., and
and Remuzzi, G. (2021) Immunity, endothelial injury and
Schinazi, R. F. (2021) COVID 19: discovery, diagnostics
complement induced coagulopathy in COVID 19, Nat.
and drug development, J. Hepatol.,
74,
168184,
Rev. Nephrol., 17, 46 64, doi: 10.1038/s41581 020 00357 4.
doi: 10.1016/j.jhep.2020.09.031.
БИОХИМИЯ том 86 вып. 7 2021
3*
980
КОРДЮКОВА и др.
3.
Paget, J., Spreeuwenberg, P., Charu, V., Taylor, R. J.,
teins S, M and E, J. Gen. Virol., 86, 14231434,
Iuliano, A. D., et al. (2019) Global mortality associated
doi: 10.1099/vir.0.80671 0.
with seasonal influenza epidemics: new burden estimates
19.
Neuman, B. W., Kiss, G., Kunding, A. H., Bhella, D.,
and predictors from the GLaMOR Project, J. Glob. Health,
Baksh, M. F., et al. (2011) A structural analysis of M pro
9, 020421, doi: 10.7189/jogh.09.020421.
tein in coronavirus assembly and morphology, J. Struct.
4.
Dawood, F. S., Iuliano, A. D., Reed, C., Meltzer, M. I.,
Biol., 174, 11 22, doi: 10.1016/j.jsb.2010.11.021.
Shay, D. K., et al. (2012) Estimated global mortality asso
20.
Wang, J., Ji, J., Ye, J., Zhao, X., Wen, J., et al. (2003) The
ciated with the first 12 months of 2009 pandemic influenza
structure analysis and antigenicity study of the N protein of
A H1N1 virus circulation: a modelling study, Lancet Infect.
SARS CoV, Genomics Proteomics Bioinformatics, 1, 145
Dis., 12, 687 695, doi: 10.1016/S1473 3099(12)70121 4.
154, doi: 10.1016/s1672 0229(03)01018 0.
5.
Cui, J., Li, F., and Shi, Z. L. (2019) Origin and evolution
21.
Mesquita, F. S., Abrami, L., Sergeeva, O., Turelli, P.,
of pathogenic coronaviruses, Nat. Rev. Microbiol., 17, 181
Kunz, B., et al. (2021) S acylation controls SARS Cov 2
192, doi: 10.1038/s41579 018 0118 9.
membrane lipid organization and enhances infectivity,
6.
Petrosillo, N., Viceconte, G., Ergonul, O., Ippolito, G.,
BioRxiv, doi: 10.1101/2021.03.14.435299.
and Petersen, E. (2020) COVID 19, SARS and MERS: are
22.
Lopez, L. A., Riffle, A. J., Pike, S. L., Gardner, D., and
they closely related? Clin. Microbiol. Infect., 26, 729 734,
Hogue, B. G. (2008) Importance of conserved cysteine
doi: 10.1016/j.cmi.2020.03.026.
residues in the coronavirus envelope protein, J. Virol., 82,
7.
Henderson, R., Edwards, R. J., Mansouri, K.,
3000 3010, doi: 10.1128/JVI.01914 07.
Janowska, K., Stalls, V., et al. (2020) Controlling the
23.
Boscarino, J. A., Logan, H. L., Lacny, J. J., and Gallagher,
SARS CoV 2 spike glycoprotein conformation, Nat.
T. M. (2008) Envelope protein palmitoylations are crucial
Struct. Mol. Biol., 27, 925 933, doi: 10.1038/s41594 020
for murine coronavirus assembly, J. Virol., 82, 2989 2999,
0479 4.
doi: 10.1128/JVI.01906 07.
8.
Khan, M., Adil, S. F., Alkhathlan, H. Z., Tahir, M. N.,
24.
Boson, B., Legros, V., Zhou, B., Siret, E., Mathieu, C.,
Saif, S., et al. (2020) COVID 19: a global challenge with
et al. (2021) The SARS CoV 2 envelope and membrane
old history, epidemiology and progress so far, Molecules,
proteins modulate maturation and retention of the spike
26, 39, doi: 10.3390/molecules26010039.
protein, allowing assembly of virus like particles, J. Biol.
9.
Yao, H., Song, Y., Chen, Y., Wu, N., Xu, J., et al. (2020)
Chem., 296, 100111, doi: 10.1074/jbc.RA120.016175.
Molecular architecture of the SARS CoV 2 virus, Cell,
25.
V’kovski, P., Kratzel, A., Steiner, S., Stalder, H., and
183, 730 738.e13, doi: 10.1016/j.cell.2020.09.018.
Thiel, V. (2021) Coronavirus biology and replication:
10.
Ke, Z., Oton, J., Qu, K., Cortese, M., Zila, V., et al. (2020)
implications for SARS CoV 2, Nat. Rev. Microbiol., 19,
Structures and distributions of SARS CoV 2 spike proteins
155 170, doi: 10.1038/s41579 020 00468 6.
on intact virions, Nature, 588, 498 502, doi: 10.1038/
26.
Wrobel, A. G., Benton, D. J., Xu, P., Roustan, C., Martin,
s41586 020 2665 2.
S. R., et al. (2020) SARS CoV 2 and bat RaTG13 spike
11.
Wrapp, D., Wang, N., Corbett, K. S., Goldsmith, J. A.,
glycoprotein structures inform on virus evolution and
Hsieh, C. L., et al. (2020) Cryo EM structure of the 2019
furin cleavage effects, Nat. Struct. Mol. Biol., 27, 763 767,
nCoV spike in the prefusion conformation, Science, 367,
doi: 10.1038/s41594 020 0468 7.
1260 1263, doi: 10.1126/science.abb2507.
27.
Luan, J., Lu, Y., Jin, X., and Zhang, L. (2020) Spike pro
12.
Walls, A. C., Park, Y. J., Tortorici, M. A., Wall, A.,
tein recognition of mammalian ACE2 predicts the host
McGuire, A. T., and Veesler, D. (2020) Structure, function,
range and an optimized ACE2 for SARS CoV 2 infection,
and antigenicity of the SARS CoV 2 spike glycoprotein,
Biochem. Biophys. Res. Commun.,
526,
165169,
Cell, 181, 281 292.e6, doi: 10.1016/j.cell.2020.02.058.
doi: 10.1016/j.bbrc.2020.03.047.
13.
Harris, A., Cardone, G., Winkler, D. C., Heymann, J. B.,
28.
Li, X., Giorgi, E. E., Marichannegowda, M. H., Foley, B.,
Brecher, M., et al. (2006) Influenza virus pleiomorphy
et al. (2020) Emergence of SARS CoV 2 through recombi
characterized by cryoelectron tomography, Proc. Natl.
nation and strong purifying selection, Sci. Adv., 6,
Acad. Sci. USA, 103, 19123 19127, doi: 10.1073/pnas.
eabb9153, doi: 10.1126/sciadv.abb9153.
0607614103.
29.
Sun, J., He, W. T., Wang, L., Lai, A., Ji, X., et al. (2020)
14.
Jiang, C., Yao, X., Zhao, Y., Wu, J., Huang, P., et al. (2020)
COVID 19: epidemiology, evolution, and cross discipli
Comparative review of respiratory diseases caused by coro
nary perspectives, Trends Mol. Med.,
26,
483495,
naviruses and influenza A viruses during epidemic season,
doi: 10.1016/j.molmed.2020.02.008.
Microbes Infect., 22, 236 244, doi: 10.1016/j.micinf.2020.
30.
Rochman, N. D., Wolf, Y. I., Faure, G., Mutz, P.,
05.005.
Zhang, F., and Koonin, E. (2020) Ongoing global and
15.
Zeng, R., Ruan, H. Q., Jiang, X. S., Zhou, H., Shi, L.,
regional adaptive evolution of SARS CoV 2, BioRxiv,
et al. (2004) Proteomic analysis of SARS associated coron
doi: 10.1101/2020.10.12.336644.
avirus using two dimensional liquid chromatography mass
31.
Goh, G. K. M., Dunker, A. K., Foster, J. A., and Uversky,
spectrometry and one dimensional sodium dodecyl sul
V. N. (2020) Shell disorder analysis predicts greater
fate polyacrylamide gel electrophoresis followed by mass
resilience of the SARS CoV 2 (COVID 19) outside the
spectroemtric analysis, J. Proteome Res., 3, 549555,
body and in body fluids, Microb. Pathog., 144, 104177,
doi: 10.1021/pr034111j.
doi: 10.1016/j.micpath.2020.104177.
16.
Malik, Y. A. (2020) Properties of coronavirus and SARS
32.
Klein, S., Cortese, M., Winter, S. L., Wachsmuth
CoV 2, Malays. J. Pathol., 42, 3 11.
Melm, M., Neufeldt, C. J., et al. (2020) SARS CoV 2
17.
Mahtarin, R., Islam, S., Islam, Md. J., Ullah, M. O., Ali,
structure and replication characterized by in situ cryo elec
M. A., and Halim, M. A. (2020) Structure and dynamics of
tron tomography, Nat. Commun., 11, 5885, doi: 10.1038/
membrane protein in SARS CoV 2, J. Biomol. Struct.
s41467 020 19619 7.
Dyn., 1 14, doi: 10.1080/07391102.2020.1861983.
33.
Pillay, T. S. (2020) Gene of the month: the
2019
18.
Nal, B., Chan, C., Kien, F., Siu, L., Tse, J., et al. (2005)
nCoV/SARS CoV 2 novel coronavirus spike protein,
Differential maturation and subcellular localization of
J. Clin. Pathol., 73, 366 369, doi: 10.1136/jclinpath 2020
severe acute respiratory syndrome coronavirus surface pro
206658.
БИОХИМИЯ том 86 вып. 7 2021
COVID 19: МИФЫ И РЕАЛЬНОСТЬ
981
34.
Wang, Q., Zhang, Y., Wu, L., Niu, S., Song, C., et al.
50.
Kordyukova, L. V., Serebryakova, M. V., Baratova, L. A.,
(2020) Structural and functional basis of SARS CoV 2
and Veit, M. (2010) Site specific attachment of palmitate
entry by using human ACE2, Cell, 181, 894 904.e9,
or stearate to cytoplasmic versus transmembrane cysteines
doi: 10.1016/j.cell.2020.03.045.
is a common feature of viral spike proteins, Virology, 398,
35.
Hussain, M., Jabeen, N., Raza, F., Shabbir, S., Baig, A. A.,
49 56, doi: 10.1016/j.virol.2009.11.039.
Amanullah, A., and Aziz, B. (2020) Structural variations in
51.
Kordyukova, L. V., Serebryakova, M. V., Baratova, L. A.,
human ACE2 may influence its binding with SARS CoV 2
and Veit, M. (2008) S acylation of the hemagglutinin of
spike protein, J. Med. Virol., 92, 1580 1586, doi: 10.1002/
influenza viruses: mass spectrometry reveals site specific
jmv.25832.
attachment of stearic acid to a transmembrane cysteine,
36.
Coutard, B., Valle, C., de Lamballerie, X., Canard, B.,
J. Virol., 82, 9288 9292, doi: 10.1128/JVI.00704 08.
Seidah, N. G., and Decroly, E. (2020) The spike glycopro
52.
Serebryakova, M. V., Kordyukova, L. V., Baratova, L. A.,
tein of the new coronavirus 2019 nCoV contains a furin
and Markushin, S. G. (2006) Mass spectrometric sequenc
like cleavage site absent in CoV of the same clade, Antiviral
ing and acylation character analysis of C terminal anchor
Res., 176, 104742, doi: 10.1016/j.antiviral.2020.104742.
ing segment from influenza A hemagglutinin, Eur. J. Mass
37.
Shang, J., Ye, G., Shi, K., Wan, Y., Luo, C., et al. (2020)
Spectrom. (Chichester), 12, 51 62, doi: 10.1255/ejms.792.
Structural basis of receptor recognition by SARS CoV 2,
53.
Chakraborty, S., Mallajosyula, V., Tato, C. M., Tan, G. S.,
Nature, 581, 221 224, doi: 10.1038/s41586 020 2179 y.
and Wang, T. T. (2021) SARS CoV 2 vaccines in advanced
38.
Steinhauer, D. A. (1999) Role of hemagglutinin cleavage
clinical trials: where do we stand? Adv. Drug Deliv. Rev.,
for the pathogenicity of influenza virus, Virology, 258, 1
172, 314 338, doi: 10.1016/j.addr.2021.01.014.
20, doi: 10.1006/viro.1999.9716.
54.
Shang, J., Wan, Y., Luo, C., Ye, G., Geng, Q., et al. (2020)
39.
Klenk, H. D., and Garten, W. (1994) Host cell proteases
Cell entry mechanisms of SARS CoV 2, Proc. Natl. Acad.
controlling virus pathogenicity, Trends Microbiol., 2, 39
Sci. USA,
117,
1172711734, doi:
10.1073/pnas.
43, doi: 10.1016/0966 842x(94)90123 6.
2003138117.
40.
Bertram, S., Glowacka, I., Steffen, I., Kühl, A., and
55.
Benton, D. J., Wrobel, A. G., Xu, P., Roustan, C., Martin,
Pöhlmann, S. (2010) Novel insights into proteolytic cleav
S. R., et al. (2020) Receptor binding and priming of the
age of influenza virus hemagglutinin: proteolytic activation
spike protein of SARS CoV 2 for membrane fusion,
of influenza virus, Rev. Med. Virol.,
20,
298310,
Nature, 588, 327 330, doi: 10.1038/s41586 020 2772 0.
doi: 10.1002/rmv.657.
56.
Chernomordik, L. V., and Kozlov, M. M.
(2008)
41.
Turoňová, B., Sikora, M., Schürmann, C., Hagen,
Mechanics of membrane fusion, Nat. Struct. Mol. Biol., 15,
W. J. H., Welsch, S., et al. (2020) In situ structural analysis
675 683, doi: 10.1038/nsmb.1455.
of SARS CoV 2 spike reveals flexibility mediated by three
57.
Clausen, T. M., Sandoval, D. R., Spliid, C. B., Pihl, J.,
hinges, Science, 370, 203208, doi: 10.1126/science.
Perrett, H. R., et al. (2020) SARS CoV 2 infection
abd5223.
depends on cellular heparan sulfate and ACE2, Cell, 183,
42.
Watanabe, Y., Allen, J. D., Wrapp, D., McLellan, J. S., and
1043 1057. e15, doi: 10.1016/j.cell.2020.09.033.
Crispin, M. (2020) Site specific glycan analysis of the
58.
Toelzer, C., Gupta, K., Yadav, S. K. N., Borucu, U.,
SARS CoV 2 spike, Science, 369, 330 333, doi: 10.1126/
Davidson, A. D., et al. (2020) Free fatty acid binding pock
science.abb9983.
et in the locked structure of SARS CoV 2 spike protein,
43.
Shajahan, A., Supekar, N. T., Gleinich, A. S., and Azadi, P.
Science, 370, 725 730, doi: 10.1126/science.abd3255.
(2020) Deducing the N and O glycosylation profile of the
59.
Liu, C., Mendonça, L., Yang, Y., Gao, Y., Shen, C., et al.
spike protein of novel coronavirus SARS CoV 2,
(2020) The architecture of inactivated SARS CoV 2 with
Glycobiology, 30, 981 988, doi: 10.1093/glycob/cwaa042.
postfusion spikes revealed by Cryo EM and Cryo ET,
44.
Cai, Y., Zhang, J., Xiao, T., Peng, H., Sterling, S. M., et al.
Structure, 28, 1218 1224.e4, doi: 10.1016/j.str.2020.10.001.
(2020) Distinct conformational states of SARS CoV 2 spike
60.
Wan, Y., Shang, J., Sun, S., Tai, W., Chen, J., et al. (2020)
protein, Science, 369, 1586 1592, doi: 10.1126/science.
Molecular mechanism for antibody dependent enhance
abd4251.
ment of coronavirus entry, J. Virol., 94, e02015 19,
45.
Petit, C. M., Chouljenko, V. N., Iyer, A., Colgrove, R.,
doi: 10.1128/JVI.02015 19.
Farzan, M., et al. (2007) Palmitoylation of the cysteine
61.
Isin, B., Doruker, P., and Bahar, I. (2002) Functional
rich endodomain of the SARS coronavirus spike glycopro
motions of influenza virus hemagglutinin: a structure
tein is important for spike mediated cell fusion, Virology,
based analytical approach, Biophys. J., 82, 569581,
360, 264 274, doi: 10.1016/j.virol.2006.10.034.
doi: 10.1016/S0006 3495(02)75422 2.
46.
Veit, M. (2012) Palmitoylation of virus proteins, Biol. Cell,
62.
Benton, D. J., Nans, A., Calder, L. J., Turner, J., Neu, U.,
104, 493 515, doi: 10.1111/boc.201200006.
et al. (2018) Influenza hemagglutinin membrane anchor,
47.
Serebryakova, M. V., Kordyukova, L. V., Semashko, T. A.,
Proc. Natl. Acad. Sci. USA, 115, 10112 10117, doi: 10.1073/
Ksenofontov, A. L., Rudneva, I. A., et al. (2011) Influenza
pnas.1810927115.
virus hemagglutinin spike neck architectures and interac
63.
Plante, J. A., Liu, Y., Liu, J., Xia, H., Johnson, B. A., et al.
tion with model enzymes evaluated by MALDI TOF mass
(2021) Spike mutation D614G alters SARS CoV 2 fitness,
spectrometry and bioinformatics tools, Virus Res., 160,
Nature, 592, 116 121, doi: 10.1038/s41586 020 2895 3.
294 304, doi: 10.1016/j.virusres.2011.07.002.
64.
Mishra, S. K., and Tripathi, T. (2020) One year update on
48.
Veit, M., Serebryakova, M. V., and Kordyukova, L. V.
the COVID 19 pandemic: where are we now? Acta Trop.,
(2013) Palmitoylation of influenza virus proteins, Biochem.
214, 105778, doi: 10.1016/j.actatropica.2020.105778.
Soc. Trans., 41, 50 55, doi: 10.1042/BST20120210.
65.
Korber, B., Fischer, W. M., Gnanakaran, S., Yoon, H.,
49.
Kordyukova, L. V., Serebryakova, M. V., Polyansky, A. A.,
Theiler, J., et al. (2020) Tracking changes in SARS CoV 2
Kropotkina, E. A., Alexeevski, A. V., et al. (2011) Linker
spike: evidence that D614G increases infectivity of the
and/or transmembrane regions of influenza A/group 1,
COVID 19 virus, Cell, 182, 812 827.e19, doi: 10.1016/
A/group 2, and type B virus hemagglutinins are packed
j.cell.2020.06.043.
differently within trimers, Biochim. Biophys. Acta, 1808,
66.
Tegally, H., Wilkinson, E., Giovanetti, M., Iranzadeh, A.,
1843 1854, doi: 10.1016/j.bbamem.2011.03.005.
Fonseca, V., et al. (2021) Detection of a SARS CoV 2
БИОХИМИЯ том 86 вып. 7 2021
982
КОРДЮКОВА и др.
variant of concern in South Africa, Nature, 592, 438 443,
option for COVID 19, Antimicrob. Agents Chemother., 64,
doi: 10.1038/s41586 021 03402 9.
e00754 20, doi: 10.1128/AAC.00754 20.
67.
Xie, X., Liu, Y., Liu, J., Zhang, X., Zou, J., Fontes
82.
Elmezayen, A. D., Al Obaidi, A., Şahin, A. T., and
Garfias, C. R., et al. (2021) Neutralization of SARS CoV
Yelekçi, K. (2020) Drug repurposing for coronavirus
2 spike 69/70 deletion, E484K and N501Y variants by
(COVID 19): in silico screening of known drugs against
BNT162b2 vaccine elicited sera, Nat. Med., 27, 620 621,
coronavirus
3CL hydrolase and protease enzymes,
doi: 10.1038/s41591 021 01270 4.
J. Biomol. Struct. Dyn., 39, 2980 2992, doi: 10.1080/
68.
Loganathan, S., Kuppusamy, M., Wankhar, W.,
07391102.2020.1758791.
Gurugubelli, K. R., Mahadevappa, V. H., et al. (2021)
83.
Loginova, S. Ya., Borisevich, S. V., Rusinov, V. L.,
Angiotensin converting enzyme 2 (ACE2): COVID 19 gate
Ulomskii, U. N., Charushin, V. N., and Chupakhin, O. N.
way to multiple organ failure syndromes, Respir. Physiol.
(2014) Investigation of Triazaverin antiviral activity against
Neurobiol., 283, 103548, doi: 10.1016/j.resp.2020.103548.
tick borne encephalitis pathogen in cell culture, Antibiot.
69.
Lv, H., Wu, N. C., and Mok, C. K. P. (2020) COVID 19
Khimioter., 59, 3 5.
vaccines: knowing the unknown, Eur. J. Immunol., 50, 939
84.
Shahab, S., and Sheikhi, M. (2020) Triazavirin - poten
943, doi: 10.1002/eji.202048663.
tial inhibitor for 2019 nCoV Coronavirus M protease: a
70.
La Vignera, S., Cannarella, R., Condorelli, R. A.,
DFT study, Curr. Mol. Med., doi:
10.2174/
Torre, F., Aversa, A., and Calogero, A. E. (2020) Sex spe
1566524020666200521075848.
cific SARS CoV 2 mortality: among hormone modulated
85.
Wu, X., Yu, K., Wang, Y., Xu, W., Ma, H., et al. (2020)
ACE2 expression, risk of venous thromboembolism and
Efficacy and safety of triazavirin therapy for Coronavirus
hypovitaminosis D, Int. J. Mol. Sci.,
21,
2948,
disease
2019: a pilot randomized controlled trial,
doi: 10.3390/ijms21082948.
Engineering (Beijing), 6, 1185 1191, doi: 10.1016/j.eng.
71.
McNab, F., Mayer Barber, K., Sher, A., Wack, A., and
2020.08.011.
O’Garra, A. (2015) Type I interferons in infectious disease,
86.
Outlaw, V. K., Bovier, F. T., Mears, M. C., Cajimat, M. N.,
Nat. Rev. Immunol., 15, 87 103, doi: 10.1038/nri3787.
Zhu, Y., et al. (2020) Inhibition of coronavirus entry
72.
Galani, I. E., Rovina, N., Lampropoulou, V.,
in vitro and ex vivo by a lipid conjugated peptide derived
Triantafyllia, V., Manioudaki, M., et al. (2021) Untuned
from the SARS Cov 2 spike glycoprotein hrc domain,
antiviral immunity in COVID 19 revealed by temporal
mBio, 11, e01935 20, doi: 10.1128/mBio.01935 20.
type I/III interferon patterns and flu comparison, Nat.
87.
Zang, R., Case, J. B., Yutuc, E., Ma, X., Shen, S., et al.
Immunol., 22, 32 40, doi: 10.1038/s41590 020 00840 x.
(2020) Cholesterol 25 hydroxylase suppresses SARS CoV
73.
Nagashima, S., Mendes, M. C., Camargo Martins, A. P.,
2 replication by blocking membrane fusion, Proc. Natl.
Borges, N. H., Godoy, T. M., et al. (2020) Endothelial dys
Acad. Sci. USA, 117, 32105 32113, doi: 10.1073/pnas.
function and thrombosis in patients with COVID 19 -
2012197117.
brief report, Arterioscler. Thromb. Vasc. Biol., 40, 2404
88.
Wu, D., Koganti, R., Lambe, U. P., Yadavalli, T., Nandi,
2407, doi: 10.1161/ATVBAHA.120.314860.
S. S., and Shukla, D. (2020) Vaccines and therapies in
74.
Suzuki, Y. J., Nikolaienko, S. I., Dibrova, V. A., Dibrova,
development for SARS CoV 2 infections, J. Clin. Med., 9,
Y. V., Vasylyk, V. M., et al. (2021) SARS CoV 2 spike pro
1885, doi: 10.3390/jcm9061885.
tein mediated cell signaling in lung vascular cells, Vascul.
89.
Hu, Y., Meng, X., Zhang, F., Xiang, Y., and Wang, J.
Pharmacol., 137, 106823, doi: 10.1016/j.vph.2020.106823.
(2021) The in vitro antiviral activity of lactoferrin against
75.
Suresh, S. J., and Suzuki, Y. J. (2020) SARS CoV 2 spike
common human coronaviruses and SARS CoV 2 is medi
protein and lung vascular cells, J. Respir., 1, 40 48,
ated by targeting the heparan sulfate co receptor, Emerg.
doi: 10.3390/jor1010004.
Microbes Infect., 10, 317 330, doi: 10.1080/22221751.
76.
Zhirnov, O. P. (2020) Molecular targets in the chemother
2021.1888660.
apy of coronavirus infection, Biochemistry (Moscow), 85,
90.
Pinto, D., Park, Y. J., Beltramello, M., Walls, A. C.,
523 530, doi: 10.1134/S0006297920050016.
Tortorici, M. A., et al. (2020) Cross neutralization of
77.
Francés Monerris, A., Hognon, C., Miclot, T., Garc a
SARS CoV 2 by a human monoclonal SARS CoV anti
Iriepa, C., Iriepa, I., et al. (2020) Molecular basis of
body, Nature, 583, 290 295, doi: 10.1038/s41586 020
SARS Cov 2 infection and rational design of potential
2349 y.
antiviral agents: modeling and simulation approaches,
91.
Wu, Y., Wang, F., Shen, C., Peng, W., Li, D., et al. (2020)
J. Proteome Res., 19, 42914315, doi: 10.1021/acs.
A noncompeting pair of human neutralizing antibodies
jproteome.0c00779.
block COVID 19 virus binding to its receptor ACE2,
78.
Gordon, C. J., Tchesnokov, E. P., Feng, J. Y., Porter, D. P.,
Science, 368, 1274 1278, doi: 10.1126/science.abc2241.
and Götte, M. (2020) The antiviral compound remdesivir
92.
Widera, M., Wilhelm, A., Hoehl, S., Pallas, C.,
potently inhibits RNA dependent RNA polymerase from
Kohmer, N., et al. (2021) Bamlanivimab does not neutral
Middle East respiratory syndrome coronavirus, J. Biol.
ize two SARS CoV 2 variants carrying E484K in vitro,
Chem., 295, 47734779, doi: 10.1021/acs.jproteome.
MedRxiv, doi: 10.1101/2021.02.24.21252372.
0c00779.
93.
Kretsos, K., Golor, G., Jullion, A., Hickling, M.,
79.
Mahase, E. (2020) Covid 19: remdesivir probably reduces
McCabe, S., et al. (2014) Safety and pharmacokinetics of
recovery time, but evidence is uncertain, panel finds, BMJ,
olokizumab, an anti IL 6 monoclonal antibody, adminis
370, m3049, doi: 10.1136/bmj.m3049.
tered to healthy male volunteers: a randomized phase I
80.
Hoffmann, M., Kleine Weber, H., Schroeder, S.,
study, Clin. Pharmacol. Drug Dev.,
3,
388395,
Krüger, N., Herrler, T., et al. (2020) SARS CoV 2 cell
doi: 10.1002/cpdd.121.
entry depends on ACE2 and TMPRSS2 and is blocked by
94.
Mercurio, I., Tragni, V., Busto, F., De Grassi, A., and
a clinically proven protease inhibitor, Cell, 181,
271
Pierri, C. L. (2021) Protein structure analysis of the inter
280.e8, doi: 10.1016/j.cell.2020.02.052.
actions between SARS CoV 2 spike protein and the
81.
Hoffmann, M., Schroeder, S., Kleine Weber, H., Müller,
human ACE2 receptor: from conformational changes to
M. A., Drosten, C., and Pöhlmann, S. (2020) Nafamostat
novel neutralizing antibodies, Cell. Mol. Life Sci., 78, 1501
mesylate blocks activation of SARS CoV 2: new treatment
1522, doi: 10.1007/s00018 020 03580 1.
БИОХИМИЯ том 86 вып. 7 2021
COVID 19: МИФЫ И РЕАЛЬНОСТЬ
983
95. Verbeke, R., Lentacker, I., De Smedt, S. C., and
109. Ewert, K., Ahmad, A., Evans, H. M., and Safinya, C. R.
Dewitte, H. (2019) Three decades of messenger RNA vac
(2005) Cationic lipid DNA complexes for non viral gene
cine development, Nano Today, 28, 100766, doi: 10.1016/
therapy: relating supramolecular structures to cellular
j.nantod.2019.100766.
pathways, Expert Opin. Biol. Ther., 5, 33 53, doi: 10.1517/
96. Inoh, Y., Nagai, M., Matsushita, K., Nakanishi, M., and
14712598.5.1.33.
Furuno, T. (2017) Gene transfection efficiency into den
110. Hervas Stubbs, S., Perez Gracia, J. L., Rouzaut, A.,
dritic cells is influenced by the size of cationic lipo
Sanmamed, M. F., Le Bon, A., and Melero, I. (2011)
somes/DNA complexes, Eur. J. Pharm. Sci., 102, 230 236,
Direct effects of type I interferons on cells of the immune
doi: 10.1016/j.ejps.2017.03.023.
system, Clin. Cancer Res., 17, 2619 2627, doi: 10.1158/
97. Patel, S., Ashwanikumar, N., Robinson, E., DuRoss, A.,
1078 0432. CCR 10 1114.
Sun, C., et al. (2017) Boosting intracellular delivery of lipid
111.
Crouse, J., Kalinke, U., and Oxenius, A.
(2015)
nanoparticle encapsulated mRNA, Nano Lett., 17, 5711
Regulation of antiviral T cell responses by type I interfer
5718, doi: 10.1021/acs.nanolett.7b02664.
ons, Nat. Rev. Immunol., 15, 231 242, doi: 10.1038/
98. Hassett, K. J., Benenato, K. E., Jacquinet, E., Lee, A.,
nri3806.
Woods, A., et al. (2019) Optimization of lipid nanoparticles for
112. Longhi, M. P., Trumpfheller, C., Idoyaga, J., Caskey, M.,
intramuscular administration of mRNA vaccines, Mol. Ther.
Matos, I., et al. (2009) Dendritic cells require a systemic
Nucleic Acids, 15, 1 11, doi: 10.1016/j.omtn.2019.01.013.
type I interferon response to mature and induce CD4+ Th1
99. Koloskova, O. O., Nikonova, A. A., Budanova, U. A.,
immunity with poly IC as adjuvant, J. Exp. Med., 206,
Shilovskiy, I. P., Kofiadi, I. A., et al. (2016) Synthesis and
1589 1602, doi: 10.1084/jem.20090247.
evaluation of novel lipopeptide as a vehicle for efficient
113. Domeier, P. P., Chodisetti, S. B., Schell, S. L., Kawasawa,
gene delivery and gene silencing, Eur. J. Pharm. Biopharm.,
Y. I., Fasnacht, M. J., et al. (2018) B cell intrinsic type 1
102, 159 167, doi: 10.1016/j.ejpb.2016.03.014.
interferon signaling is crucial for loss of tolerance and the
100. Loseva, A. A., Budanova, U. A., and Sobyakin, Yu. L.
development of autoreactive B cells, Cell Rep., 24, 406
(2019) Synthesis of new guanidine containing amphiphiles
418, doi: 10.1016/j.celrep.2018.06.046.
and their pyrene analogue for liposomal delivery system
114. Marro, B. S., Legrain, S., Ware, B. C., and Oldstone,
and visualization in target cells, Russ. J. Org. Chem., 55,
M. B. A. (2019) Macrophage IFN I signaling promotes
1826 1831, doi: 10.1134/S1070428019120030.
autoreactive T cell infiltration into islets in type 1 diabetes
101. Rietwyk, S., and Peer, D. (2017) Next generation lipids in
model, JCI Insight, 4, e125067, doi: 10.1172/jci.insight.
RNA interference therapeutics, ACS Nano, 11, 7572 7586,
125067.
doi: 10.1021/acsnano.7b04734.
115. Tatsis, N., and Ertl, H. C. J. (2004) Adenoviruses as vac
102. Basha, G., Novobrantseva, T. I., Rosin, N., Tam, Y. Y.,
cine vectors, Mol. Ther., 10, 616629, doi: 10.1016/
Hafez, I. M., et al. (2011) Influence of cationic lipid com
j.ymthe.2004.07.013.
position on gene silencing properties of lipid nanoparticle
116. Bilbao, G., Contreras, J. L., Zhang, H. G., Pike, M. J.,
formulations of siRNA in antigen presenting cells, Mol.
Overturf, K., et al. (1999) Adenovirus mediated gene
Ther., 19, 2186 2200, doi: 10.1038/mt.2011.190.
expression in vivo is enhanced by the antiapoptotic bcl 2
103. Feldman, R. A., Fuhr, R., Smolenov, I., Ribeiro, A.,
gene, J. Virol., 73, 6992 7000, doi: 10.1128/JVI.73.8.6992
Panther, L., et al. (2019) mRNA vaccines against H10N8
7000.1999.
and H7N9 influenza viruses of pandemic potential are
117. Xiang, Z. Q., Yang, Y., Wilson, J. M., and Ertl, H. C. J.
immunogenic and well tolerated in healthy adults in phase
(1996) A replication defective human adenovirus recombi
1 randomized clinical trials, Vaccine, 37, 33263334,
nant serves as a highly efficacious vaccine carrier, Virology,
doi: 10.1016/j.vaccine.2019.04.074.
219, 220 227, doi: 10.1006/viro.1996.0239.
104. Gan, L. M., Lagerström Fermér, M., Carlsson, L. G.,
118. Stephen, S. L., Montini, E., Sivanandam, V. G., Al
Arfvidsson, C., Egnell, A. C., et al. (2019) Intradermal
Dhalimy, M., Kestler, H. A., et al. (2010) Chromosomal
delivery of modified mRNA encoding VEGF A in patients
integration of adenoviral vector DNA in vivo, J. Virol., 84,
with type 2 diabetes, Nat. Commun., 10, 871, doi: 10.1038/
9987 9994, doi: 10.1128/JVI. 00751 10.
s41467 019 08852 4.
119. Crystal, R. G. (2014) Adenovirus: the first effective in vivo
105. Roth, C., Cantaert, T., Colas, C., Prot, M.,
gene delivery vector, Hum. Gene Ther.,
25,
3 11,
Casadémont, I., et al. (2019) A modified mRNA vaccine
doi: 10.1089/hum.2013.2527.
targeting immunodominant NS epitopes protects against
120. Logunov, D. Y., Dolzhikova, I. V., Shcheblyakov, D. V.,
dengue virus infection in HLA class I transgenic mice,
Tukhvatulin, A. I., Zubkova, O. V., et al. (2021) Safety and
Front. Immunol., 10, 1424, doi: 10.3389/fimmu.2019.01424.
efficacy of an rAd26 and rAd5 vector based heterologous
106. Jagger, B. W., Dowd, K. A., Chen, R. E., Desai, P.,
prime boost COVID 19 vaccine: an interim analysis of a
Foreman, B., et al. (2019) Protective efficacy of nucleic
randomised controlled phase 3 trial in Russia, Lancet, 397,
acid vaccines against transmission of zika virus during
671 681, doi: 10.1016/S0140 6736(21)00234 8.
pregnancy in mice, J. Infect. Dis., 220, 15771588,
121. Ryzhikov, A. B., (2021) A single blind, placebo controlled
doi: 10.1093/infdis/jiz338.
randomized study of the safety, reactogenicity and
107. Liang, F., Lindgren, G., Lin, A., Thompson, E. A., Ols, S.,
immunogenicity of the “EpiVacCorona” vaccine for the
et al. (2017) Efficient targeting and activation of antigen
prevention of COVID 19, in volunteers aged 18 60 years
presenting cells in vivo after modified mRNA vaccine
(phase I II), Russ. J. Infect. Immunity, 11, 283296,
administration in rhesus macaques, Mol. Ther., 25, 2635
doi: 10.15789/2220 7619 ASB 1699.
2647, doi: 10.1016/j.ymthe.2017.08.006.
122. Andersen, K. G., Rambaut, A., Lipkin, W. I., Holmes,
108. Espeseth, A. S., Cejas, P. J., Citron, M. P., Wang, D.,
E. C., and Garry, R. F. (2020) The proximal origin of SARS
DiStefano, D. J., et al. (2020) Modified mRNA/lipid
CoV 2, Nat. Med., 26, 450 452, doi: 10.1038/s41591 020
nanoparticle based vaccines expressing respiratory syncy
0820 9.
tial virus F protein variants are immunogenic and protec
123. Chen, Z., and John Wherry, E. (2020) T cell responses in
tive in rodent models of RSV infection, NPJ Vaccines, 5,
patients with COVID 19, Nat. Rev. Immunol., 20, 529 536,
16, doi: 10.1038/s41541 020 0163 z.
doi: 10.1038/s41577 020 0402 6.
БИОХИМИЯ том 86 вып. 7 2021
984
КОРДЮКОВА и др.
COVID 19: MYTHS AND REALITY
Review
L. V. Kordyukova1* and A. V. Shanko2,3
1 Belozersky Institute of Physico Chemical Biology, Lomonosov Moscow State University,
119991 Moscow, Russia; E mail: kord@belozersky.msu.ru
2 FORT LLC, R&D Department, 119435 Moscow, Russia
3 Ivanovsky Institute of Virology, N. F. Gamaleya Federal Research Center
for Epidemiology and Microbiology, 123098 Moscow, Russia
COVID 19, a new human respiratory disease that has killed nearly 3 million people over a year since the start of the
pandemic, is a global public health challenge. Its infectious agent, SARS CoV 2, has several significant structural dif
ferences from other coronaviruses which make it more pathogenic and transmissible. In this review, we discuss some
important characteristics of the main surface antigen of SARS CoV 2, the spike (S) protein, such as (1) the ability of
the receptor binding domain (RBD) switching between a standing up position (“open” pre fusion conformation) for
receptor binding and a lying down position (“closed” pre fusion conformation) for immune evasion; (2) taking
advantage of the high binding affinity of RBD in an open conformation with the human ACE2 receptor for efficient
cell entry; (3) priming of S protein by intra cellular furin like proteases to spread the virus across different cell types.
We describe an interaction between S protein and cellular receptors, co receptors and antagonists as well as a hypo
thetical mechanism of destabilization of the homotrimeric spike structure that triggers the fusion of the viral envelope
with the cell membrane at physiological pH and mediates the entry of viral nucleocapsid into the cytoplasm. The
observed transition of pre fusion conformation of S protein to post fusion on the surface of virions after their treat
ment with some reagents such as β propiolactone is crucial, especially in relation to vaccine production. We also com
pare a few peculiarities of COVID 19 as well as S protein of SARS CoV 2 with those of outbreaks of severe Influenza
disease caused by highly pathogenic influenza A virus strains of H5 and H7 subtypes and structural features of hemag
glutinins of those viruses. In conclusion we touch on promising and rolled out COVID 19 anti viral and pathogenet
ic therapeutics, some recently approved conventional and innovative COVID 19 vaccines, and their molecular and
immunological features.
Keywords: COVID 19, SARS CoV 2, S protein, structure, influenza virus, hemagglutinin, vaccines
БИОХИМИЯ том 86 вып. 7 2021