БИОХИМИЯ, 2022, том 87, вып. 2, с. 230 - 257
УДК 575.113
ИССЛЕДОВАНИЯ МОЛЕКУЛЯРНОЙ РЕГУЛЯЦИИ ПРОЦЕССОВ
РАЗВИТИЯ НИЗШИХ МНОГОКЛЕТОЧНЫХ НА ПРИМЕРЕ
СТРЕКАЮЩИХ С ИСПОЛЬЗОВАНИЕМ ТЕХНОЛОГИЙ
ВЫСОКОПРОИЗВОДИТЕЛЬНОГО СЕКВЕНИРОВАНИЯ
Обзор
© 2022
Т.В. Ерофеева1,2, А.П. Григоренко1,2*, Ф.Е. Гусев1,2,
И.А. Косевич1,3, Е.И. Рогаев1,2,3,4
1 Научный центр генетики и наук о жизни, направление генетика, АНО ВО «Научно$технологический
университет “Сириус”», 354349 Краснодарский край, Сочи, Россия; электронная почта: anast1998@mail.ru
2 Институт общей генетики им. Н.И. Вавилова РАН, 119991 Москва, Россия
3 Московский государственный университет имени М.В. Ломоносова,
биологический факультет, 119234 Москва, Россия
4 Медицинская школа Чен Массачусетского университета, департамент психиатрии, 01545 Шрусбери, США
Поступила в редакцию 28.10.2021
После доработки 13.12.2021
Принята к публикации 17.01.2022
Исключительный набор особенностей и характеристик представителей типа Cnidaria (Стрекающие) делает
их модельным объектом для широкого круга исследований. Особый научный интерес представляют плас#
тичность жизненного цикла и связанные с ним процессы клеточной дифференцировки и развития целост#
ного многоклеточного организма. Новый уровень развития молекулярно#генетических методов, в том чис#
ле использование методов широкомасштабного секвенирования геномов, транскриптомов и эпигеномов,
как на уровне целого организма, так и на уровне отдельных клеток, делает возможным получение детальной
картины развития этих животных. В представленном обзоре рассматриваются современные подходы и дос#
тижения с использованием методов широкомасштабного секвенирования в реконструкции процессов он#
тогенеза Cnidaria путём изучения регуляторных путей клеточной трансдукции и их взаимодействий.
КЛЮЧЕВЫЕ СЛОВА: Cnidaria, развитие, методы секвенирования, геномика, сигнальные каскады.
DOI: 10.31857/S0320972522020075
ВВЕДЕНИЕ
вотных (Bilateria) [1]. Cnidaria, являясь двухслой#
ными (Diploblastica) по организации животны#
Многоклеточные организмы, содержащие
ми, обладают отличительной морфологической
различные дифференцированные типы клеток,
характеристикой - наличием специализирован#
подразделяются на низшие и высшие. К «низ#
ных стрекательных клеток, «книдоцитов», кото#
шим многоклеточным» относят пластинчатых
рые используются для захвата добычи, защиты и
(Placozoa), губок (Porifera), гребневиков
передвижения [2]. Разнообразные представите#
(Ctenophora) и стрекающих (Cnidaria), в то время
ли Cnidaria - коралловые полипы, актинии,
как к «высшим» относят всех трехслойных била#
сцифо# и гидромедузы, жгучие сифонофоры ти#
терально#симметричных многоклеточных жи#
па Португальского кораблика и морские осы из
группы кубомедуз представляют собой заметную
Принятые сокращения: ATAC#seq - анализ доступ# и важную часть экосистем океанов и морей. От#
ного для транспозазы хроматина с использованием секве#
дельную немаловажную роль играют представи#
нирования (Assay for Transposase#Accessible Chromatin using
sequencing); ChIP#seq - иммунопреципитация хроматина с
тели актиний и кораллов и их эндосимбионты -
последующим секвенированием (Chromatin immunopre#
одноклеточные водоросли. Успешный симбиоз
cipitation followed by sequencing); MARS#Seq - массивное обусловливает рост и выживание рифовых ко#
параллельное секвенирование транскриптомов единичных раллов в неблагоприятных условиях с обеднён#
клеток (Massively parallel single cell RNA#Seq); scRNA#
ным содержанием питательных веществ [3].
seq - секвенирование транскриптомов единичных клеток
(single cell RNA#sequencing); UMI - уникальный молеку#
В свою очередь, кораллы вступают в симбиоз с
лярный идентификатор (Unique molecular identifier).
макроорганизмами, обеспечивая безопасную
* Адресат для корреспонденции.
среду обитания для большого количества морс#
230
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
231
ких животных (рыбы, двустворчатые моллюски,
состоящие из нейритов (отростков нейронов),
ракообразные) [4]. Согласно систематике меж#
кроме того, у многих медуз есть специализиро#
дународной базы данных WoRMS, cложная фи#
ванные органы чувств - светочувствительные
логенетическая организация Cnidaria в настоя#
глазки и органы равновесия [10, 11]. У прикреп#
щее время характеризуется делением на следую#
лённых полипов нервная система организована
щие классы: Anthozoa, Cubozoa, Hydrozoa,
как сеть из нейронов, располагающаяся в осно#
Myxozoa, Scyphozoa и Staurozoa [5].
вании эпителиев по всему телу, с некоторой кон#
Стрекающие являются объектом активного
центрацией нейронов в оральной и аборальной
научного изучения. Относительная простота ор#
областях [10, 11]. Определённая регионализация
ганизации, высокая способность к регенерации,
нервной системы, связанная с формированием
пластичность жизненного цикла и сложная фи#
мезентериальных нервных тяжей (располагаю#
логенетическая структура делают Cnidaria уни#
щихся вдоль энтодермальных септ#мезентерий),
кальными представителями мировой фауны. Ис#
как у полипов Anthozoa [12], и нервных колец,
следования, связанные с изучением транскрип#
как у некоторых Hydra и медуз, наблюдается у
томов и эпигеномов, позволяющие охарактери#
Стрекающих, что, однако, не нарушает основ#
зовать как целый организм, так и его клеточное
ной принцип организации нервной системы для
строение, демонстрируют сложную организа#
этого типа [13]. Движение представителей Стре#
цию представителей Cnidaria со специфичными
кающих обеспечивается наличием эпителиаль#
особенностями строения и онтогенеза.
но#мышечных клеток с сократительными отро#
стками в базальной части. Эти клетки составля#
ют основу обоих эпителиев. Сократительные от#
ХАРАКТЕРИСТИКА ТИПА СТРЕКАЮЩИХ
ростки эпителиально#мышечных клеток эпи#
дермы и гастродермы полипов относятся пре#
Тип Стрекающие (Cnidaria) объединяет раз#
имущественно к гладкому типу. У медуз на внут#
нообразных представителей водной среды оби#
ренней стороне колокола сократительные отро#
тания с преобладанием внешней радиальной
стки эпителиально#мышечных клеток относят#
симметрии, рассматриваемый как сестринская
ся к поперечнополосатому типу [14]. Существу#
группа билатерально#симметричных животных.
ют вариации строения мышечной системы у
Стрекающие - это двухслойные животные, на#
Стрекающих, подробно описанные в обзоре
ходящиеся на уровне эпителиальной организа#
Leclère и Röttinger [14]. Здесь следует упомянуть
ции [6]. Их тело построено двумя эпителиями -
несколько особенностей строения в разных
эпидермой и гастродермой, которые разделены
группах Стрекающих. У многих Hydrozoa в щу#
межклеточным матриксом или мезоглеей.
пальцах энтодермальные эпителиальные клетки
У Стрекающих отсутствуют органы. Пищевари#
не содержат миофиламентов, а у многих поли#
тельная система представлена ротовым отверс#
пов Medusozoa имеются мощные продольные
тием, которое ведёт в единую замкнутую гаст#
сократительные пучки эктодермального проис#
ральную полость. Гастральная полость может
хождения, миоциты которых полностью распо#
быть представлена сложно#разветвлённой сис#
лагаются в мезоглее и не связанны ни с одним из
темой каналов и называется гастроваскулярной
эпителиев стенки тела. [14]. У Scyphozoa, как
системой [7]. Ротовое отверстие - единственное
правило, отсутствуют миофибриллы во внеш#
отверстие пищеварительной системы, использу#
нем слое колокола (эксумбрелле) [15]. У боль#
емое как для заглатывания пищи, так и для вы#
шинства Anthozoa, в отличие от Medusozoa, хо#
броса непереваренных остатков. Самым важ#
рошо развиты эпителиально#мышечные клетки
ным отличительным признаком Стрекающих
энтодермы, составляющие круговую мускулату#
является наличие стрекательных клеток или
ру стенки тела, продольные париетальные мыш#
книдоцитов со специализированными органел#
цы у основания септ и ретракторные мышцы на
лами, которые представляют собой капсулы с
одной стороне каждой септы. А эктодермальные
выстреливающими нитями, использующимися
мышцы большинства Anthozoa развиты лишь в
для охоты, защиты и прикрепления [2]. Книдо#
зоне щупалец и ротового отверстия. Кроме того,
циты вместе с сенсорными клетками, которые
на примере Nematostella vectensis было показано
имеют чувствительные реснички на апикальных
наличие в щупальцах мышечных клеток, обо#
частях, выходящих на поверхность эпителиев, и
собленных от эпителия в отдельный слой кле#
сетью ганглиозных клеток формируют нервную
ток, не несущий эпителиальной функции [16].
систему Стрекающих, которая различается по
У паразитических Endocnidozoa отсутствуют
строению в зависимости от жизненной формы
эпителиально#мышечные клетки: миоциты
организма [8, 9]. У свободноплавающих медуз
(гладкомышечного типа) расположены отдель#
по краю колокола имеются нервные кольца,
но от эпителия в мезоглее [17].
БИОХИМИЯ том 87 вып. 2 2022
232
ЕРОФЕЕВА и др.
ют и отклонения от общей схемы жизненного
цикла. Например, у Hydrozoa могут быть реду#
цированы стадии полипа или медузы. Так, у
подкласса Trachylina полипы либо очень ма#
ленького размера, либо отсутствуют [23]. В то же
время у представителей подкласса Hydroidolina
может отсутствовать стадия медузы (например,
у некоторых Leptothecata [24]). У ряда предста#
вителей семейств Bougainvillidae, Hydractiniidae
и Rathkeidae вегетативное размножение может
происходить не только на стадии полипа, но и
на стадии медузы [25]. У некоторых сцифоид#
ных медуз в жизненном цикле также отсутствует
стадия полипа, например, у Pelagia [26]. Парази#
тические Endocnidozoa включают животных с
разнообразными жизненными циклами.
Myxozoa - это микроскопические эндопарази#
ты, колонизирующие беспозвоночных и позво#
ночных, жизненный цикл которых реализуется
с участием двух хозяев. Polypodiozoa проходят в
жизненном цикле через свободноживущую ста#
дию и паразитических личинок осетровых рыб,
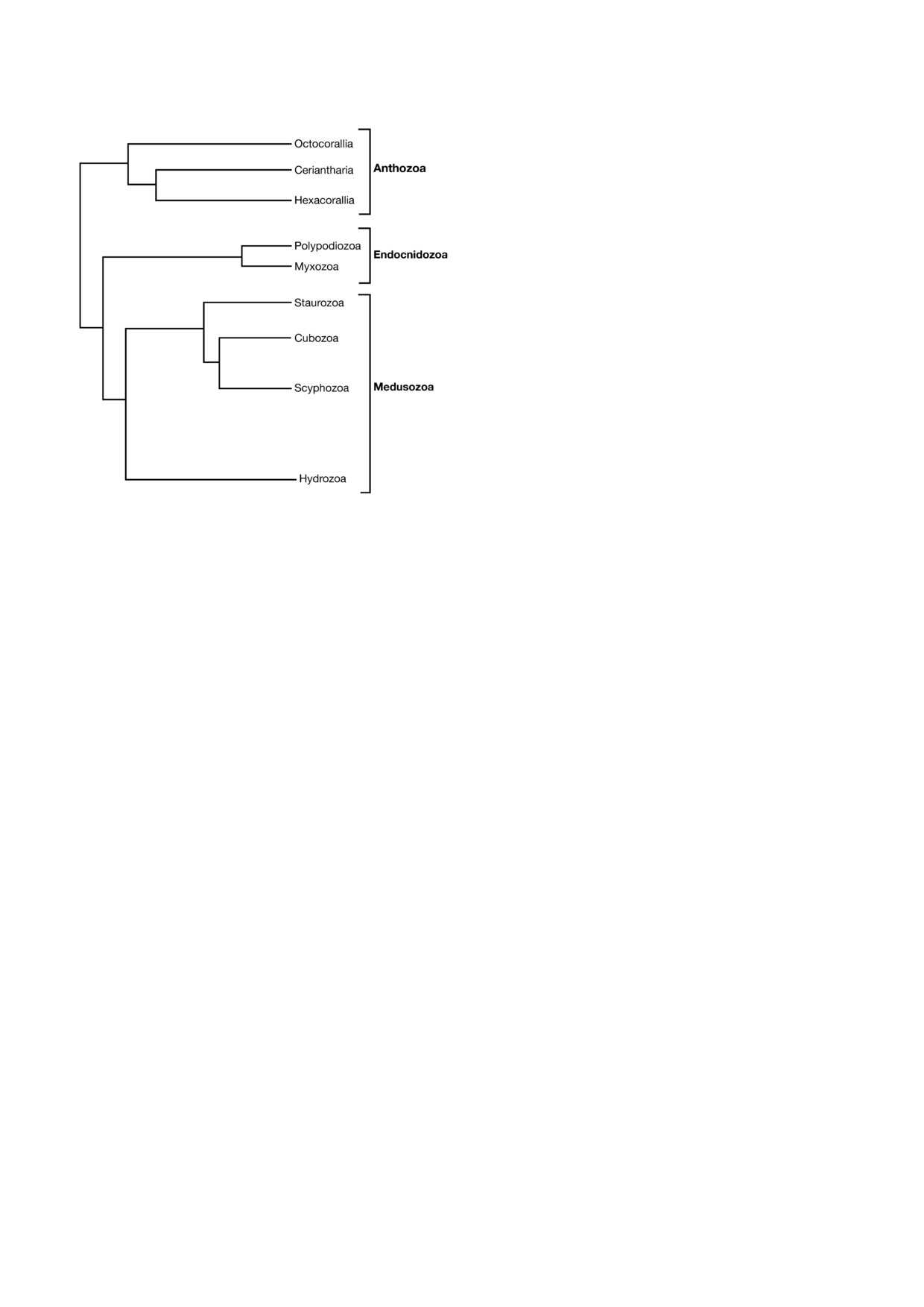
Рис. 1. Филогенетическое дерево, отображающее тополо#
гию основных классов Стрекающих, построенное на осно#
соответственно, имеют одного хозяина в жиз#
вании последних опубликованных данных полногеномно#
ненном цикле [17].
го секвенирования [18, 19, 21]
Отдельного рассмотрения заслуживает онто#
генез представителей Medusozoa. Помимо опи#
санного выше классического жизненного цик#
Тип Стрекающие представлен следующими
ла, у них было обнаружено уникальное явление
классами: Anthozoa, Cubozoa, Hydrozoa,
«обратного развития», которое заключается в
Myxozoa, Scyphozoa, Staurozoa [5]. Филогенети#
переходе организма определённой стадии жиз#
ческая структура Cnidaria является предметом
ненного цикла не в следующую, а в предыдущую
дискуссий, но, по последним данным, она опи#
стадию («возврат к предыдущей стадии»). Счи#
сывается делением на 3 монофилетические груп#
тается, что данный процесс вызывается стрессо#
пы - Anthozoa (шести# и восьмилучевые корал#
выми ситуациями, например голоданием или
лы, актинии), Medusozoa, объединяющая клас#
изменением условий среды обитания. «Обрат#
сы Cubozoa, Hydrozoa, Scyphozoa и Staurozoa, и
ное развитие» было зафиксировано у несколь#
Endocnidozoa, объединяющая паразитических
ких представителей Hydrozoa на медузоидной
Myxozoa и Polypodiozoa (рис. 1) [18-21].
стадии [25]. Одним из самых известных иссле#
Стрекающие демонстрируют большое раз#
дований является эксперимент с гидромедузами
нообразие жизненных циклов, отличающихся у
Turritopsis spp. (Hydrozoa), в котором в результа#
разных групп. У Anthozoa полипоидная стадия
те воздействия факторов, вызывающих стресс,
формирует гаметы, и жизненный цикл включает
таких как резкое повышение температуры, из#
стадии зародыша → личинки (планулы) → ре#
менение солёности воды или механическое по#
продуктивного полипа. Медузоидная стадия в
вреждение колокола медузы, организм полово#
жизненном цикле Anthozoa отсутствует. У
зрелой медузы регрессировал и образовывал ша#
представителей Medusozoa жизненный цикл
рообразное скопление клеток (цисту), из кото#
осуществляется по схеме: полипоидная стадия
рого впоследствии формировался новый по#
(которая размножается только вегетативным
лип [27]. Недавно подобное явление «обратного
путём) → медузоидная стадия (формирующая
развития» было описано и для сцифоме#
половые продукты)
→ гаметы
→ заро#
дуз (Scyphozoa) Aurelia aurita [28].
дыш → планула → полип. Исключение состав#
ляет Hydra, у которой отсутствуют стадии меду#
зы и планулы, и которая размножается преиму#
СИГНАЛЬНЫЕ ПУТИ, ЗАДЕЙСТВОВАННЫЕ
щественно вегетативным путём (почкованием),
В ПРОЦЕССАХ РАЗВИТИЯ У СТРЕКАЮЩИХ
а половое размножение реализуется по схеме:
гаметы (развивающиеся в основании эпидер#
Закладка орально#аборальной оси, форми#
мы) → зародыш → новый полип [22]. Существу#
рование разных типов тканей и органов, про#
БИОХИМИЯ том 87 вып. 2 2022
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
233
цессы регенерации, бесполое и половое раз#
TGF#β (таблица). У обоих классов (Myxozoa и
множение регулируются клеточными молеку#
Polypodiozoa) присутствуют ключевые составля#
лярными каскадами, среди которых ряд сиг#
ющие пути передачи сигналов Notch [18]. Авто#
нальных путей, являющихся эволюционно кон#
ры предполагают, что основной причиной отсут#
сервативными для Стрекающих и билатераль#
ствия полноценных сигнальных путей разви#
но#симметричных животных (Bilateria). Рядом
тия является уменьшение размера генома в свя#
исследований было показано, что у Стрекаю#
зи с переходом к паразитизму. Эксперименталь#
щих присутствуют сигнальные пути, которые
ные данные по функциям вышеупомянутых
также регулируют раннее эмбриональное раз#
сигнальных каскадов у паразитических Cnidaria
витие у Bilateria. К таким ключевым сигналь#
отсутствуют. Имеющиеся данные демонстриру#
ным клеточным путям относятся: Wnt#,
ют эволюционную консервативность сигналь#
Hedgehog#, Notch#, TGF#β# и RTK#путь [29].
ных каскадов, задействованных в развитии
В данном разделе будут рассмотрены молеку#
Cnidaria.
лярно#генетические характеристики структуры
Системы сигнальных каскадов играют важ#
сигнальных путей и их роль в развитии различ#
ную роль в процессах клеточной дифференци#
ных представителей Стрекающих.
ровки и развития живого организма. Далее будет
Большое количество научных исследований,
рассмотрена функциональная роль каждого мо#
связанных с изучением молекулярных сигналь#
лекулярного пути в ключевых биологических
ных каскадов в онтогенезе Стрекающих, демон#
процессах развития разных Стрекающих.
стрирует эволюционную консервативность этих
Формирование осевого паттерна вдоль ораль=
путей у Cnidaria и высших Metazoa. Данные по
но=аборальной оси и образование вторичной оси.
секвенированию геномов, транскриптомов и
Среди Стрекающих обнаруживаются как живот#
функциональные исследования роли путей кле#
ные с радиальной симметрией, имеющие одну
точной трансдукции в развитии Стрекающих
ось полярности (большая часть Hydrozoa и
показывают, что основные компоненты пере#
Medusozoa), так и организмы c элементами би#
численных выше сигнальных путей открыты
латеральной симметрии (кораллы и актинии
практически в каждом классе Cnidaria. Здесь
Anthozoa) с направляющей осью симметрии,
нужно отметить, что наибольшее количество
перпендикулярной орально#аборальной оси те#
исследований структуры молекулярных каска#
ла. Например, у N. vectensis вторичная ось тела
дов проведено на активно использующихся в
определяется щелевидной формой глотки и ас#
научных экспериментах моделях Anthozoa и
симетричным расположением ретракторных
Hydrozoa. В геномах Hydra и N. vectensis были
мышц на септах [38]. Однако вопрос о гомоло#
найдены все вышеперечисленные сигнальные
гичности данного варианта билатеральной сим#
пути [30, 31]. В базе данных генов и гено#
метрии таковому у Bilateria остаётся откры#
мов KEGG, объединяющей геномную и систем#
тым [39-43]. Эволюционно консервативной
ную функциональную информацию о разных
сигнальной системой, определяющей общность
организмах, можно найти подробные схемы
развития Стрекающих и высших Metazoa, явля#
сигнальных путей Wnt, Notch и TGF#β у Hydra и
ется молекулярный путь Wnt, который опосре#
N. vectensis [32]. У Scyphozoa молекулярная
дует формирование осевого паттерна вдоль
структура различных каскадов детектируется
орально#аборальной оси тела в раннем эмбрио#
наличием отдельных компонентов путей на ге#
генезе [43-47]. Впервые это было показано на
номном уровне [33, 34], при этом в работе
примере Hydra: после выделения ортологов ос#
Brekhman et al. [35] предложена модель пути
новных компонентов канонического сигналь#
Wnt/β#catenin у A. aurita. Меньше всего инфор#
ного пути Wnt/β#catenin с помощью гибридиза#
мации о структуре сигнальных путей имеется по
ции in situ определили участки локальной
представителям Cubozoa и Staurozoa [33, 36, 37].
экспрессии ортологов лиганда Wnt3 и эффек#
В геномах паразитических стрекающих
торного транскрипционного фактора Tcf/Lef в
Myxozoa, в отличие от других классов Cnidaria и
апикальных частях у взрослых полипов и у
паразитических Polypodiozoa (сестринский так#
вновь образующихся в процессе почкования.
сон Myxozoa с типичным для свободноживущих
Экспрессия гена β$catenin была повышенной в
Стрекающих строением тела), отсутствуют пол#
зоне образования будущей почки - месте зак#
ноценные сигнальные каскады, связанные с
ладки вторичной оси тела, а ортологов белка
процессами дифференцировки, развития и
Dishevelled и киназы гликогенсинтазы#3
межклеточных коммуникаций [18]. Среди ком#
(GSK3) - конститутивно на низком уровне по
понентов Wnt#пути у них обнаружены только
всему телу Hydra. Формирование апикальной
некоторые эффекторные белки, а также некото#
части Hydra сопровождалось экспрессией Wnt и
рые элементы сигнальных путей Hedgehog и
Tcf/Lef и в процессе регенерации головы, и в
6 БИОХИМИЯ том 87 вып. 2 2022
234
ЕРОФЕЕВА и др.
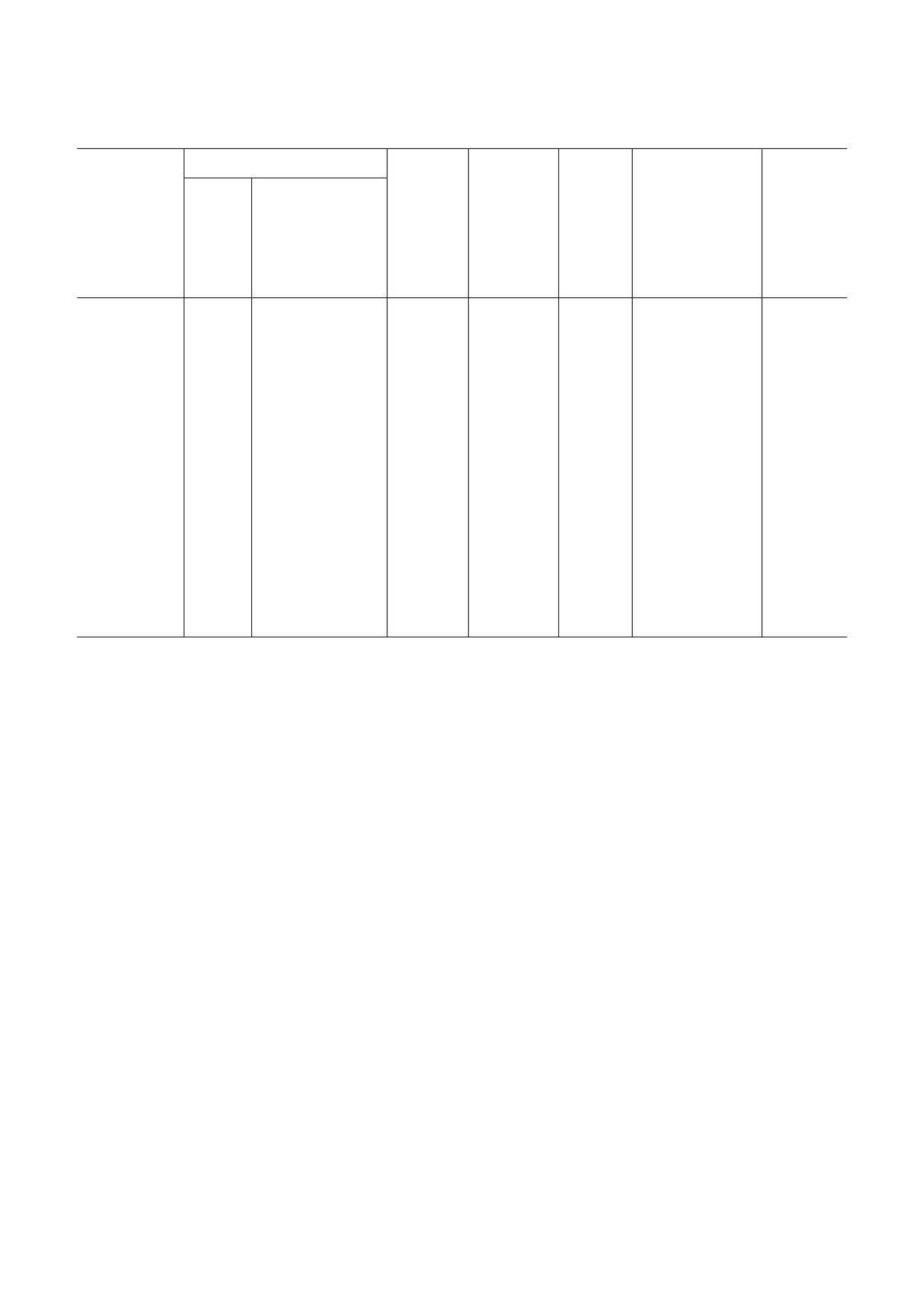
Сигнальные пути и гомологи белков, задействованные в процессах развития у различных представителей Cnidaria
Сигнальный
Hydrozoa
путь/Класс
(представители)
Medusozoa
(Hydractinia, Clytia
Scyphozoa
Anthozoa
Cubozoa
Myxozoa (Kudoa
Polypodiozoa
Hydra sp.
hemisphaerica,
(Aurelia,
(Nematostella
(Morbakka
iwatai, Myxobolus
(Polypodium
Cladonema pacificum,
Nemopilema)
vectensis)
virulenta)
cerebralis)
hydriforme)
Podocoryne carnea,
Dynamena pumila,
Ectopleura larynx)
Wnt
[30, 47-53,
[45, 46, 67, 68, 78,
[45, 46, 67,
[33#35, 100]
[33]
лиганды и рецеп#
[18]
79, 83,
87, 89, 123, 124, 135,
68, 78, 87,
торы пути утраче#
110-115,
138, 139, 144, 188]
89, 123, 124,
ны, но есть экс#
118, 131,
135, 138,
прессия эффектор#
169, 181]
139, 144, 188]
ных белков канони#
ческого и некано#
нического путей
Hedgehog
[30, 119]
[89]
[89]
[100]
нет
[18]
[18]
данных
Notch
[30, 54,
[99, 188]
[99, 188]
нет данных
нет
[18]
[18]
84, 117]
данных
TGF#β
[30, 85,
[89, 188]
[89, 188]
[33, 100]
[36]
[18]
[18]
118, 131,
181]
RTK
[30, 86,
[125, 137]
[125, 137]
нет данных
нет
нет данных
нет данных
106, 118,
данных
131, 181,
191]
Примечание. Таблица составлена по данным экспрессии, геномным и функциональным данным и базе данных KEGG [32].
процессе de novo образования головной части в
представителя Anthozoa N. vectensis выявило оп#
клеточных агрегатах из диссоциированной кле#
ределённые закономерности формирования
точной суспензии [47]. Дальнейшие функцио#
первичной оси тела, схожие с Hydra, но со свои#
нальные исследования на Hydra определили
ми особенностями: большое разнообразие орто#
ключевую роль пути Wnt в процессе формирова#
логов лигандов Wnt (подсемейства генов WntA
ния орально#аборальной оси тела с определени#
и Wnt1-11) экспрессировалось вдоль оси плану#
ем апикальной части организма - головного ор#
лы с ограничением экспрессии около бластопо#
ганизатора и вторичной оси тела, предшествую#
ра в процессе гаструляции и в зоне ротового
щей эвагинации почек [48-53]. Кроме того, в
участка у полипов. Экспрессия лигандов Wnt
процессе организации головной части Hydra,
была ограничена одним слоем ткани (эктодер#
участвующей в формировании оси тела, также
мой или энтодермой) [55]. Дальнейшие иссле#
может быть задействован сигнальный путь
дования определили, что лиганды Wnt, Dsh а
Notch, регулирующий работу генов лиган#
также β#catenin, являются основными регулято#
дов (Wnt7) и транскрипционных факторов#ми#
рами развития осевого паттерна N. vectensis в
шеней пути Wnt (Sp5 и Tcf) [54]. Анализ диффе#
раннем эмбриогенезе и в регенеративных про#
ренциальной экспрессии генов по всему геному
цессах [43, 44, 56-59]. β#Catenin, локализую#
Hydra после фармакологического ингибирова#
щийся в ядрах клеток бластомеров, демонстри#
ния каскада Notch выявил снижение экспрес#
рует первый молекулярный сайт осевой асси#
сии Wnt7, Tcf и Sp5 с последующим восстановле#
метрии на стадии бластулы [56]. Dsh, стабилизи#
нием до нормального уровня после удаления
рующий β#catenin, локализуется в анимальном
ингибитора Notch [54].
полюсе ооцита, дающем начало закладки ораль#
Исследования формирования орально#або#
ного полюса взрослого организма, и затем в
ральной оси тела на других моделях Стрекаю#
бластомерах [57]. В установлении осевого пат#
щих также демонстрируют важную роль молеку#
терна у N. vectensis на стадии планулы также
лярного каскада Wnt. Изучение эмбриогенеза
включается консервативная система генов гомео#
БИОХИМИЯ том 87 вып. 2 2022
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
235
боксовых транскрипционных факторов Hox,
рассмотрена роль сигнальных каскадов в регу#
важных регуляторов позиционной идентичнос#
ляции гаструляции.
ти вдоль передне#задней оси билатерий [44].
Из всех изучаемых Стрекающих больше все#
Филогенетически среди Hox#генов N. vectensis
го исследований молекулярной регуляции гаст#
представлены передние (NvAx6, NvAx6a, NvAx7
руляции посвящено Anthozoa. Подробные
и NvAx8) и центральные#задние (NvAx1, NvAx1a)
функциональные исследования N. vectensis вы#
гены [44]. NvAx6 и NvAx1 экспрессируются в
явили, что в процессе гаструляции активно ра#
оральной и аборальной областях соответственно,
ботает сигнальный каскад Wnt, который также
в процессе эмбрионального развития N. vecten$
детерминирует формирование первичной оси у
sis, что, возможно, опосредует регуляцию фор#
планулы, что связано с экспрессией множества
мирования основной оси тела с участием пере#
Wnt#лигандов в зоне бластопора (эктодерме или
дачи сигнала Wnt [44]. NvAx1a, NvAx6a, NvAx7,
энтодерме после завершения гаструляции) [55].
NvAx8 и NvAx8a, а также Gbx (ген подкласса Hox$
Разные компоненты канонического и некано#
like) экспрессируются вдоль направительной
нического сигнальных путей Wnt опосредуют
оси [42, 60] с задействованием каскада BMP в
процессы спецификации энтодермы [57], фор#
регуляции паттерна направительной оси [61].
мирования паттерна будущих энтомезодер#
Показано, что, помимо Wnt#пути, в формирова#
мальных и эктодермальных клеток c участием
нии осевого паттерна в процессе развития
сигнальных каскадов BMP, FGF
[71] и
N. vectensis задействованы сигнальные каскады
Hedgehog [72], образования архентерона и оп#
FGF, TGF#β и Hedgehog [62]. TGF#β#каскад че#
ределения клеточной судьбы во время гаструля#
рез передачу сигнала BMP задействован и в
ции [73], а также формирования паттерна
формировании направляющей оси тела
орального полюса [58, 65]. Ингибирование ка#
N. vectensis [59, 61, 63, 64], зависящей также от
нонического пути Wnt за счёт прямого отключе#
работы Wnt#каскада до установления направля#
ния трансляции β#catenin полностью блокиро#
ющей оси [59, 65].
вало гаструляцию [56, 74], однако нокдаун гена,
Отдельные компоненты пути Wnt определя#
кодирующего транскрипционный фактор Tcf,
ют развитие осевого паттерна в раннем эмбрио#
не влиял на прохождение гаструляции, но пре#
генезе и поддержание осевой идентичности в
дотвращал образование нормальной глот#
процессе онтогенеза у представителей гидро#
ки [71]. Важным моментом является то, что об#
идных медуз (лиганды Wnt, рецептор Frizzled,
разование архентерона, регулирующееся путём
Tcf) [45, 66-68]. Для представителей сцифоид#
Wnt/PCP, может происходить независимо от эн#
ных медуз рода Aurelia была показана неодно#
томезодермальной спецификации судьбы кле#
родная экспрессия лигандов Wnt на разных
ток, регулирующейся каскадом Wnt/β#catenin,
стадиях жизненного цикла: со спецификой для
что говорит о независимой эволюции этих двух
стадии полипа и тканеспецифичной экспрес#
процессов [75].
сией лигандов на стадии медузы, что демон#
У гидроидных распространены различные
стрирует консервативный механизм определе#
пути гаструляции, однако даже в случае аполяр#
ния передне#задней оси тела и тканевой иден#
ного пути, характеризующегося отсутствием вы#
тичности [33].
раженной морфологической полярности эмбри#
Регуляция сегрегации зародышевых листков.
она [70], на уровне гаструлы формируется пат#
Процесс образования зародышевых листков,
терн орально#аборальной оси, детерминирую#
являющийся ключевым этапом эмбриогенеза
щийся градиентом сигнального каскада Wnt
для всех Metazoa, ассоциирован с гаструляцией.
[46]. В то же время у некоторых гидроидных
У Стрекающих гаструляция опосредует образо#
морфогенетические процессы гаструляции и
вание двух слоёв клеток тела - эктодермы и эн#
спецификации клеточной судьбы не связаны с
тодермы. В общем случае инициация формиро#
молекулярной осью, опосредованной передачей
вания зародышевого слоя начинается с опреде#
сигнала Wnt [76, 77]. Удлинённая форма плану#
ления эмбрионального участка начала сегрега#
лы у гидроидной медузы Clytia образуется во
ции и определения тканевой идентичности
время гаструляции и опосредуется Wnt/РСР#
листка (определение клеточной судьбы), сопро#
каскадом [78]. Функциональная роль Wnt#пути в
вождающегося морфогенетическими движения#
эмбриогенезе Hydra, в отличие от взрослого по#
ми энтомезодермы (участка, отделённого от эк#
липа, пока не установлена, однако показана
тодермы и недифференцированного в энто# или
экспрессия ортологов Frizzled, β$catenin и Tcf, но
мезодерму) [69]. С подробным описанием меха#
не лиганда Wnt, транскрипты которого детекти#
низмов гаструляции у разных Стрекающих мож#
руются после завершения гаструляции. β$Сatenin
но ознакомиться в обзорах Technau [69], а также
и Tcf экспрессируются равномерно по всему
Kraus и Markov [70]. В данном подразделе будет
эмбриону [79].
БИОХИМИЯ том 87 вып. 2 2022
6*
236
ЕРОФЕЕВА и др.
Регуляция образования половых клеток. Стре#
гетативном полюсе, Wnt3 - в зоне анимального
кающие демонстрируют большое разнообразие
полюса [87]. Функциональные исследования де#
жизненных циклов и связанных с ними репро#
монстрируют изменения полярности ооцита
дуктивных систем [80]. Различные сигнальные
при нарушении пути передачи сигнала Wnt
каскады задействованы в процессе образования
(подробно описано в обзоре Houliston et al. [88]).
половых клеток. У Hydra гаметогенез начинает#
У Hydractinia symbiolongicarpus оогенез начинает#
ся в субпопуляции интерстициальных стволо#
ся в герминальной зоне тела гонозооида и про#
вых клеток в эктодерме. В процессе оогенеза
должается в гонофоре. Сперматогенез пол#
после нескольких этапов делений интерстици#
ностью проходит в мужском гонофоре. В про#
альных клеток с образованием кластеров, а за#
цессе гаметогенеза включается путь Hedgehog,
тем единого клеточного агрегата определяется
гомологи лигандов которого экспрессируются в
единственный ооцит. Оставшиеся клетки диф#
гастродермисе мужских и женских гонофо#
ференцируются во вспомогательные (эндоци#
ров [89]. Кроме того, может быть задействован
ты), которые после апоптоза фагоцитируются
сигнальный каскад TGF#β через BMP: экспрес#
растушим ооцитом [81, 82]. На ранних этапах
сия предполагаемого рецептора BMP
оогенеза, ещё в процессе формирования класте#
(BmpR_54452) и белка Capicua (репрессора тар#
ров интерстициальных клеток, увеличивается
гетных генов пути) была детектирована в разви#
экспрессия киназы GSK#3β, негативного регу#
вающихся ооцитах женских гонофоров и гаст#
лятора пути Wnt, которая снижается к моменту
родермисе мужских гонофоров [89].
установления ооцита. В то же время в процессе
Гаметогенез Anthozoa происходит в энтодер#
сперматогенеза увеличение экспрессии GSK#3β
ме мезентериев. В процессе роста и созревания
было зафиксировано лишь на поздней стадии в
ооциты постепенно выпячиваются в мезоглею,
базальной части сперматогония [83]. Функцио#
сохраняя контакт с гастродермой через трофоне#
нальные исследования показали, что GSK#3β
му, сформированную специализированными со#
необходима для инициации апоптоза в эндоци#
матическими клетками гастродермы, трофоци#
тах в процессе оогенеза [83]. Кроме того, в ооци#
тами. Мужские половые клетки в процессе спер#
те была показана экспрессия генов Frizzled, β$
матогенеза агрегируются с образованием фолли#
catenin и Tcf, но не Wnt [79]. Сигнальный каскад
кула, окружённого слоем мезоглеи. В дальней#
Notch также играет важную роль в оогенезе у
шем половые продукты попадают в гастральную
Hydra: нарушение пути передачи сигнала приво#
полость через разрывы эпителия мезентериев и
дит к ингибированию дифференциации интер#
выводятся наружу через рот [90, 91]. Wnt#каскад
стициальных клеток#предшественников ооци#
регулирует образование и поддержание ани#
та, что приводит к образованию обширных кле#
мально#вегетативной оси ооцита за счёт аккуму#
точных кластеров [84]. Определены гены#мише#
ляции белка Dishevelled на анимальном полюсе и
ни пути Notch в эндоцитах Hydra [54]. Путь кле#
возможной деградации на вегетативном [57].
точной трансдукции TGF#β/Smad также может
Молекулярный путь Hedgehog необходим для
быть задействован в оогенезе, но не в спермато#
формирования кластеров первичных половых
генезе Hydra: высокая экспрессия ортолога
клеток у N. vectensis, которые локализуются в ме#
Smad1 была детектирована у предшественников
зентериях в зоне между экспрессией лиганда
ооцита, а также на разных стадиях оогенеза,
Hedgehog1 и доменами рецептора Patched [72].
включая агрегацию и фагоцитоз [85]. Новая ре#
В результате световой индукции высвобождения
цепторная тирозинкиназа Lemon была открыта
ооцитов у N. vectensis была охарактеризована це#
у Hydra в процессе изучения гаметогенеза, что
лая сеть молекулярных каскадов, по данным
может свидетельствовать о включении пу#
транскриптомов, включающая увеличение
ти RTK: увеличение экспрессии наблюдалось в
экспрессии компонентов пути Wnt и RTK [92].
агрегированных интерстициальных клетках до
У склерактиниевого коралла Euphyllia ancora бы#
формирования мужской и женской гамет [86].
ла обнаружена активная экспрессия гомоло#
У Medusozoa гаметогенез происходит в гоно#
га Notch - сигнального Notch#каскада, опосреду#
зооидах - прикреплённых полипах с мужскими
ющего межклеточные взаимодействия в незре#
и женскими выростами#гонофорами или же в
лых и зрелых ооцитах [93].
гонадах у плавающих медуз [80]. У гидроидных
Регуляция нейрогенеза. Сенсорные клетки
медуз каскад Wnt задействован в формировании
эктодермы с апикальной ресничкой, ганглиоз#
полярности ооцита - анимально#вегетативной
ные клетки базальной части эктодермы и нема#
оси: экспрессия Frizzled1 локализуется в зоне
тоциты (книдоциты) составляют массив нерв#
анимального полюса около ядра ооцита в про#
ных клеток Стрекающих. У Hydrozoa нейроге#
цессе роста. Во время созревания ооцита
нез начинается в интерстициальных стволовых
Frizzled3 аккумулируется в зоне кортекса на ве#
клетках эктодермы, как у Hydra, или в энтодер#
БИОХИМИЯ том 87 вып. 2 2022
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
237
ме (гидроидные медузы) с миграцией в эктодер#
ного органа у планулы N. vectensis [103, 104] а
му. У Hydra клетки#предшественники нейронов
молекулярный каскад MAPK, опосредующий
и нематоцитов мигрируют к одному из полюсов
путь RTK, стимулирует нейрогенез за счёт регу#
тела (оральному или аборальному) и затем диф#
ляции экспрессии нейрогенных факторов тран#
ференцируются в разные типы клеток в зависи#
скрипции [105]. Пути передачи сигналов VEGF
мости от положения вдоль главной оси тела.
и FGF, проходящие через MAPK, регулируются
При этом существуют вариации клеточного
каскадом Wnt/β#catenin и могут участвовать в
нейрогенеза у Hydra и других гидроидных [94].
развитии нервной системы у Hydra, однако не#
У N. vectensis нейрогенез начинается с нейраль#
обходимы более подробные функциональные
ных клеток#предшественников в бластуле. Диф#
исследования [106].
ференциация клеток происходит в клетках экто#
• Эффекторы сигнального пути TGF#β/Smad -
дермы во время гаструляции, а затем в энтодер#
белки Smad - задействованы у Hydra в процес#
ме [95, 96]. У сцифоидных медуз дифференци#
сах дифференцировки нематоцитов. У N. vecten$
рующиеся нервные клетки обнаруживаются в
sis TGF#β#каскад через ВМР, индуцирующийся
эктодерме на стадии планулы [97]. Обе группы
путём Wnt/β#catenin, необходим для поддержа#
гидроидных и сцифоидных медуз проходят этап
ния паттерна сети нейронов в зоне рта вдоль
метаморфоза в жизненном цикле, при котором
орально#аборальной и направляющей осей
происходит дегенерация части зародышевых
[101].
нейронов и новый этап дифференциации и миг#
Регуляция процессов регенерации и морфоло=
рации нервных клеток [9]. Нейрогенез у взрос#
гических особенностей строения Стрекающих.
лых медуз протекает в манубриуме и бульбусах
Стрекающие обладают широким регенератив#
щупалец [98]. Молекулярная регуляция нейро#
ным потенциалом, в связи с чем их активно ис#
генеза Стрекающих консервативна и включает
пользуют в качестве моделей для изучения реге#
работу упомянутых выше сигнальных каскадов
нерации животных. Механизмы регенерации и
(Wnt, Notch, TGF#β, FGF, Hedgehog) [94]. Под#
их молекулярная структура подробно описаны в
робная характеристика компонентов молеку#
различных обзорах [107-109]. В данном подраз#
лярных каскадов описана в обзорах Galliot и
деле будет обобщена информация по молеку#
Quiquand [9], Rentzsch et al. [94] и Galliot et al.
лярной регуляции с помощью сигнальных кас#
[98], по данным которых и по последним науч#
кадов (Wnt, Notch, Hedgehog, RTK, TGF#β) про#
ным исследованиям можно выделить ряд зако#
цессов регенерации и роста организменных осо#
номерностей молекулярной регуляции нейроге#
бенностей строения тела Стрекающих.
неза у Стрекающих.
Путь передачи сигнала Wnt является самым
• Каскад Notch регулирует дифференцировку
консервативным и распространённым каска#
предшественников нематоцитов в зрелые нема#
дом, опосредующим регенеративные процессы
тоциты у Hydra
[54,
84] и др. Hydrozoa
Стрекающих. У Hydra путь Wnt включается в
(Hydractinia echinata [99]), количество эпители#
разных моделях регенерации:
альных предшественников нервных клеток и
• при установлении главной оси тела в про#
книдогенез у Anthozoa (N. vectensis).
цессе регенерации тканей в экспериментах с
• Экспрессия компонентов пути Hedgehog
трансплантацией разных частей тела поли#
наблюдается у N. vectensis в клетках предполага#
па [53];
емых нервных предшественников [62], у A. auri$
• при регенерации участка оральной области
ta (Scyphozoa) - в ропалиях развивающейся
тела путём активации Wnt3 в интерстициальных
эфиры [100].
клетках участка регенерации [110] и последую#
• Молекулярный каскад Wnt/β#catenin участ#
щей активации пула лигандов Wnt [50] с форми#
вует в формирования паттерна нервной системы
рованием петли обратной связи [50];
у N. vectensis в апикальной части (так называе#
• роль β#catenin в регенерации головной части
мой оральной нервной сети): путь регулирует
полипа также была продемонстрирована в экс#
экспрессию нейрогенных транскрипционных
периментах с ингибированием и оверэкспрес#
факторов на стадии бластулы и необходим для
сией β#catenin [111];
определения клеточной судьбы нейральных
• при регенерации тканей подошвы, что было
предшественников нервных клеток в апикаль#
показано в экспериментах с ингибированием и
ной части тела [101]. У гидроидной медузы
оверэкспрессией β#catenin [111];
H. echinata путь передачи сигнала Wnt может
• при регенерации частей тела de novo в кле#
включаться в процессе дифференцировки ней#
точных агрегатах - организатора головной части
ронов и нематоцитов [102].
за счёт Wnt3 [47, 112], в том числе в малых клас#
• Сигнальный путь RTK через FGF задейство#
терах клеток [113], при механическом растяже#
ван в развитии сенсорного апикального реснич#
нии ткани Hydra в сфероидах из клеток, где
БИОХИМИЯ том 87 вып. 2 2022
238
ЕРОФЕЕВА и др.
Wnt3 активируется в процессе регенерации го#
ний [123]. Помимо Wnt, в разных по морфологии
ловной части за счёт зависимой от растяжения
и функциям полипах единой колонии экспрес#
транскрипционной активации его гена [114].
сируются лиганд каскада Hedgehog (гомолог
Помимо регенеративных процессов, у Hydra
Hedgehog) - в гастродермисе мужских и женских
сигнальный каскад Wnt/PCP при активации ка#
гонофоров, рецептор BMP (BmpR_54452) - в оо#
нонического пути задействован в процессах
цитах и гастродермисе мужских гонофорах, что
дифференциации эпителиальных стволовых
может быть связано с гаметогенезом [89]. В пре#
клеток в зоне образования щупалец и по#
делах одного организма Hydractinia путь Wnt ре#
чек [49]. Кроме пути передачи сигнала Wnt, у
гулирует количество и положение щупалец на
Hydra в процессах регенерации включаются и
полипе [124]. Морфогенез щупалец у Hydractinia
другие молекулярные пути. Сигнальный кас#
в процессе регенерации апикальной части тела и
кад MAPK, активирующийся при заживлении
в процессе метаморфоза планулы опосредуется
раны, модулирует Wnt при рассечении Hydra
сигнальным каскадом Notch [99]. Ветвление
пополам с возможным механизмом транскрип#
щупалец Cladonema pacificum, связанное с агре#
ционной активации лигандов Wnt, необходи#
гацией интерстициальных клеток в области
мых для формирования регенеративного про#
формирования ветви, регулируемой каска#
цесса [115], а также активирует процесс апопто#
дом MAPK, опосредуется молекулярным пу#
за, необходимого для регенерации головной
тём RTK, задействованным в удлинении вет#
части Hydra [116]. Путь Notch необходим для
вей [125].
восстановления центра организации головной
Подробное описание механизмов регуля#
части в процессе регенерации и модуляции пат#
ции регенерации и функциональное значение
терна закладки щупалец [117], а передача сиг#
каждого каскада у представителей Anthozoa
налов через VEGF и FGF играет важную роль в
описано в обзоре Röttinger [108]. Здесь следует
регенерации гипостома и щупалец [106]. Кас#
выделить важные моменты. Путь передачи сиг#
кад BMP, регулируемый канонический пу#
нала Notch и каскад MAPK у N. vectensis вклю#
тём Wnt, задействован в базальной регенерации
чаются в процессе регенерации глотки полипа
тела Hydra [118]. Путь Hedgehog может участво#
при её ампутации, при этом после нанесения
вать в регенерации апикальной и базальной
раневого повреждения проколом эпителия в за#
частей тела Hydra при рассечении: ортолог ли#
живлении раны также участвует MAPK, что
ганда Hedgehog дифференциально экспресси#
сопровождается активацией апоптоза в области
ровался в эндодермальных клетках гипостома и
повреждения [126]. Каскад Wnt регулирует оп#
базальной части тела Hydra, в зоне подошвы в
ределение, формирование и поддержание рото#
интактных животных и при рассечении тела по#
вого паттерна полипа [58]. Путь FGF, по дан#
перек - в экто# и энтодерме апикальной и ба#
ным экспрессии, активируется через 20 часов
зальной зон с усилением экспрессии в верхней
при регенерации отсечённого от ротового отве#
части тела [119].
рстия до подошвы куска тела у актинии
Регенеративные процессы представителей
Calliactis polypus, что может свидетельствовать о
Medusozoa также связаны с включением каска#
начале нейрогенеза, по аналогии с экспрессией
да Wnt. У гидроидной медузы Hydractinia каскад
компонентов FGF при нейрогенезе у планулы
функционально задействован в регенерации
N. vectensis [103, 127]. При регенерации поли#
апикальной части полипа с образованием блас#
пов колониальных склерактиниевых кораллов
темы и одновременным ингибированием роста
из недифференцированных кусочков ткани
столона [67]. Регенерация столона требует, в
включаются каскады Wnt и FGF по данным
свою очередь, ингибирования каскада Wnt [120].
экспрессии разных компонентов каждого пути
У гидроидной медузы Clytia путь Wnt активирует#
на разных этапах регенеративного процесса,
ся в процессе ремоделирования тканей купола
что свидетельствует о возможном перекрёстном
медузы, установления бластемы и регенерации
взаимодействии двух каскадов. При этом у дан#
манубриума [121]. Каскад Wnt в том числе опос#
ных кораллов сохранялась тенденция регенера#
редует морфологические особенности строения
ции головной части, подобно Hydra при попе#
отдельных Medusozoa. Так, у колониальных гид#
речном иссечении и N. vectensis при поврежде#
роидов сигнальный каскад Wnt задействован в
нии, а также активация апоптотических про#
регуляции структуры колоний, связанной с по#
цессов с последующей активацией каноничес#
ложением зачатков полипов в колонии, специ#
кого пути Wnt [128]. С подробным описанием
фической для столонов экспрессией отдельных
механизмов молекулярной регуляции регенера#
компонентов каскада в пределах одной колонии,
ции у немодельных представителей Anthozoa
что подробно описано в обзоре Cartwright
можно ознакомиться в обзоре van der Burg и
et al. [122], и с морфогенезом ветвления коло#
Prentis [129].
БИОХИМИЯ том 87 вып. 2 2022
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
239
Регуляция жизненного цикла Стрекающих.
процессе метаморфоза [99]. Лиганд VEGF, путь
Стрекающие демонстрируют большое разнооб#
передачи которого опосредуется RTK#каска#
разие жизненных циклов, включающее подвиж#
дом, но не рецептор пути VEGFR, активно
ные и прикреплённые стадии, наличие вегета#
экспрессируется в процессе метаморфоза пла#
тивного и полового процессов с вариациями по#
нулы в клетках энтодермы Podocoryne carnea.
ла (гонохоризм, гермафродитизм) [80]. Ан#
Также лиганд и рецептор экспрессируются в эн#
самбль молекулярных каскадов участвует в регу#
тодерме развивающейся медузы P. carnea на ста#
ляции перехода из стадии в стадию.
дии почки - выроста тела полипа, затем в разви#
У Hydra вегетативное размножение происхо#
вающихся радиальных каналах, клетках попе#
дит за счёт процесса почкования: эвагинация
речнополосатых мышц и бульбусах щупалец.
почки из стенки тела, сопровождающаяся про#
При этом экспрессия VEGFR в этих областях
лиферацией и тканевыми перестройками в со#
медузы низкая [137].
ответствии орально#аборальной оси нового ор#
На примере P. carnea была показана актива#
ганизма, приводит к образованию дочернего по#
ция дифференциальной экспрессии генов ка#
липа, который впоследствии отделяется и ста#
нонического пути Wnt на стадии репродуктив#
новится автономным. Сигнальные каскады
ного полипа (Wnt, Frizzled, LRP5/6, Dishevelled,
FGFR и Notch функционально задействованы в
CK2, APC, Tcf/Lef), что может быть связано с
формировании границы между родительским
дальнейшим развитием медуз на гонофо#
полипом и дочерним зачатком почки с возмож#
рах [138]. Также была детектирована коэкспрес#
ным включением пути Wnt (экспрессия Tcf и β$
сия генов лиганда Wnt3 и мембранных рецепто#
catenin показана в зоне начала эвагинации поч#
ров Frizzled (1 и 3) на дистальном и оральном
ки), что подробно описано в обзоре Böttger и
концах осей последовательного развития медуз
Hassel [130]. Размеры тела Hydra контролируют#
и полипов [138], что демонстрирует универ#
ся как Wnt#, так и TGF#β#каскадами, притом
сальную роль сигнального каскада Wnt в про#
Wnt#путь непосредственно влияет на экспрес#
цессе развития разных стадий жизненного цик#
сию компонентов TGF#β#пути. TGF#β сигналь#
ла. У представителя Стрекающих с отсутствую#
ный путь контролирует, в свою очередь, образо#
щей стадией личинки#планулы в жизненном
вание почки при вегетативном размножении,
цикле (Aplanulata) Ectopleura larynx Wnt#путь
что формирует определённый переключатель
опосредует развитие как полипов, так и ранних
развития между фазой роста и репродуктивной
мужских и женских гонофоров, представляю#
фазой, определяющий количество клеток орга#
щих собой недоразвитых медуз, прикреплён#
низма Hydra [131].
ных к полипам. При этом экспрессия компо#
У представителей класса Hydrozoa группы
нентов Wnt#пути коррелирует с половым ди#
Medusozoa отдельные молекулярные каскады
морфизмом гонофоров E. larynx, связанным с
задействованы в процессах развития поло#
образованием зачатков щупалец в оральной зо#
вой (медуза) и бесполой (полип) стадий жизнен#
не женского гонофора и шарообразной струк#
ного цикла. Медуза развивается из группы про#
туры, не разделённой на щупальца, в оральной
лиферирующих клеток, называемой энтокодо#
зоне мужского гонофора [139]. Кроме того, ав#
ном, который образуется путём инвагинации
торы исследования предполагают, что ингиби#
участка эктодермы к энтодерме с формировани#
рование экспрессии элементов пути Wnt может
ем полости [132]. Полип развивается из планулы
быть вовлечено в процессы нарушения полно#
путём метаморфоза или морфологической пере#
ценного развития медузы E. larynx [139].
стройки, сопровождающейся апоптозом и ре#
У сцифоидных медузы развиваются из апи#
дифференциацией отдельных типов кле#
кальной части полипа в процессе стробиляции
ток [133, 134]. В процессе метаморфоза планулы
(сегментации), а полип, в свою очередь, - из
включается каскад Wnt. У Clytia и Hydractinia в
планулы в результате метаморфоза, связанного с
зоне орального полюса эктодермы планулы
элиминацией энтодермы планулы и образова#
экспрессируется Wnt3, при этом у Hydractinia та#
нием вторичной энтодермы из клеток эктодер#
кая экспрессия связана с предотвращением
мы планулы с последующим развитием щупалец
апоптоза [135], что, предположительно, может
полипа
[140]. У Aurelia дифференциальная
быть и у Clytia [136]. Кроме того, у Hydractinia в
экспрессия разных компонентов каскада Wnt,
процессе метаморфоза путь Wnt/Tcf опосредует
специфическая главным образом для стадии ли#
формирование и поддержание орального пат#
чинки#планулы, и несколько лигандов со спе#
терна с редукцией развития столонов при экто#
цифической экспрессией для медузы и строби#
пической активации сигнального каскада [67].
лы были детектированы на разных стадиях жиз#
Путь Notch не задействован функционально в
ненного цикла [35]. В процессе стробиляции
формировании нервной системы Hydractinia в
Aurelia включается несколько молекулярных
БИОХИМИЯ том 87 вып. 2 2022
240
ЕРОФЕЕВА и др.
каскадов. Wnt11a экспрессируются в эктодерме
СОВРЕМЕННЫЕ МЕТОДЫ
оральной части каждого развивающегося сег#
СЕКВЕНИРОВАНИЯ В ИССЛЕДОВАНИИ
мента - будущей эфиры, что сопровождается
РАЗВИТИЯ СТРЕКАЮЩИХ
экспрессией Bmp5/8 в энтодерме развивающей#
ся гастральной полости [33]. Гиперактивация
Использование новейших молекулярно#ге#
пути Wnt приводит к нарушению стробиляции,
нетических методов, применяемых для изуче#
связанной с отсутствием границ между сегмен#
ния процессов развития, позволяет проводить
тами
[33]. Компоненты сигнального пу#
детальные исследования не только молекуляр#
ти Hedgehog экспрессируются на стадии строби#
ной структуры и локализации включения регу#
лы в сегментах и развивающейся эфиры (рецеп#
ляционных сигнальных каскадов в пределах ор#
тор Patched и лиганды SHH1, SHH2) Aurelia.
ганизма, но и их функциональной значимости в
Ингибирование этого каскада приводит к нару#
отдельных процессах онтогенеза. Изучение
шению процесса сегментации полипа в процес#
функций путей клеточной трансдукции, рас#
се стробиляции [100].
смотренных в предыдущем разделе, осущест#
У N. vectensis в результате метаморфоза пла#
вляется с помощью спектра методов, связан#
нулы теряется подвижность с оседанием на дно,
ных с генной манипуляцией, к которым в насто#
формируется ротовое отверстие, окружённое за#
ящий момент относятся фармакологические
чатками щупалец, и развиваются первые мезен#
методы модулирования активности молекуляр#
терии [96]. Несколько генов#лигандов каскада
ных каскадов и методы обратной генетики
TGF#β (Bmp2/4 и Gdf5$like) начинают экспрес#
(RNAi#, морфолино#опосредованный нокдаун,
сироваться на стадии бластулы и далее в мезен#
CRISPR/Cas9, TALEN/Fok1#опосредованный
териях N. vectensis в процессе метаморфоза [40,
нокаут, гиперэкспрессия мРНК, использование
61, 63, 141]. Сигнальный путь FGF играет важ#
индуцибельных промоторов, активируемых теп#
ную роль в процессе состояния компетентнос#
ловым шоком). Подробное описание методик
ти, то есть способности планулы задерживать
изучения процессов развития на известных мо#
метаморфоз до обнаружения определённых сиг#
делях Стрекающих рассмотрено в обзорах [22,
налов среды обитания, и метаморфоза планулы
96, 108]. В данном разделе будут рассмотрены
рифового коралла Acropora millepora, что было
последние данные по использованию методов
показано с помощью ингибироваия рецептора
широкомасштабного секвенирования при изу#
пути FGFR1 в личинках в состоянии компетен#
чении процессов развития Стрекающих.
ции [142].
Применение методов широкомасштабного сек=
Данных по изучению молекулярных каска#
венирования геномов для изучения механизмов
дов, задействованных в процессах развития у
развития и распространения Стрекающих. Техно#
представителей Cubozoa, почти нет. Известно,
логии полногеномного секвенирования активно
что в геноме Morbakka virulenta было найдено
используются для изучения разнообразных ас#
14 генов семейства Wnt [33], однако отсутствует
пектов развития и распространения Cnidaria [30,
информация по функциональной составляю#
31, 33, 34, 37, 144, 145]. Последние исследования
щей Wnt#пути.
геномов Стрекающих выявляют интересные
Процессы развития Cnidaria опосредуются
особенности этого типа в плане развития и адап#
теми же ключевыми молекулярными путями,
тации к условиям окружающей среды. Так, учё#
что и у высших Metazoa: Wnt, Hedgehog, Notch,
ные определили геномные характеристики ко#
TGF#β и RTK. Функциональная составляющая
ралловых полипов Acropora (Anthozoa) и генети#
каждого сигнального каскада индивидуальна
ческие детерминанты их приспособленности к
для отдельных классов Cnidaria, однако можно
изменению условий окружающей среды [146].
выделить закономерности включения опреде#
Молекулярное датирование позволило опреде#
лённых сигнальных путей, определяющих одни
лить эволюционную историю развития Acropora,
и те же процессы развития у представителей
выявив, что их предки возникли около 55,8 млн
Стрекающих разных классов. Так, Wnt#путь
лет назад и пережили поэтапное повышение и
опосредует формирование орально#аборальной
понижение глобальной температуры. Возможно,
оси у всех Cidaria, а путь Notch задействован в
это привело к увеличению видового разнообра#
книдогенезе у Hydra, Nematostella и Hydractinia.
зия Acropora [146]. Кроме того, была установлена
Помимо описанных выше классических сиг#
предполагаемая причина высокой чувствитель#
нальных путей, в процессах развития Cnidaria
ности кораллов Acropora, приводящая к обесцве#
задействовано большое разнообразие молеку#
чиванию, связанная с нарушением у них синтеза
лярных каскадов, специфичных именно для
цистеина, ведущего к зависимости от симбиоти#
Стрекающих. Данные сигнальные пути более
ческих водорослей. Также, по геномным данным,
подробно описаны в литературе [92, 100, 143].
были определены основные механизмы адапта#
БИОХИМИЯ том 87 вып. 2 2022
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
241
ции кораллов к стрессовым условиям окружаю#
что процесс стробиляции инициирует сигналь#
щей среды, унаследованные от далеких предков
ный путь ретиноевой кислоты, запускающийся
Acropora. Это специфичный механизм высоко#
при добавлении ретинола в среду. Также были
скоростной кальцификации кораллов, специфи#
определены индуктор стробиляции - 5#меток#
ческая тандемная дупликация генов ответа на
си#2#метилиндол (индуцировал стробиляцию
стресс (например гены, кодирующие малые цис#
через 48 часов инкубации) - и гормон естест#
теин#богатые пептиды и коралловые каспазы),
венной стробиляции, специфичный для A. auri$
увеличение количества генов диметилсульфо#
ta [147]. С помощью транскриптомного профи#
пропионат лиазы (фермента цикла серы в океане,
лирования полного жизненного цикла A. aurita
который опосредует обратную связь между ат#
были более подробно охарактеризованы основ#
мосферой и океаном и может влиять на регули#
ные сигнальные каскады, опосредующие специ#
рование местного климата), произошедшее ещё
фические для разных стадий процессы [35, 100],
у предков Acropora в результате горизонтального
и специфические для каждого этапа жизненно#
переноса генов от симбиотических водорослей,
го цикла транскрипционные факторы [35], что
и положительный отбор генов, кодирующих ан#
было также продемонстрировано на представи#
тимикробные пептиды, и генов симбиоза [146].
теле Scyphozoa Rhopilema esculentum [148]. На
Секвенирование генома гигантской медузы
стадии планулы у аурелии экспрессировались
Nemopilema nomurai (Scyphozoa) выявило инди#
основные компоненты пути Wnt (Wnt2, $3, $5b,
видуальные генетические особенности этого ви#
$8 и #16a), Frizzled, GSK$3β и Axin. В то же время
да, связанные с мобильностью и активным хищ#
на других стадиях экспрессировалась лишь
ничеством, которые опосредуются увеличением
часть лигандов: Wnt9/10, Wnt11a и Wnt3 - на
числа генов, кодирующих миозины II типа, ней#
стадии медузы, Wnt16b - на стадии строби#
ротрансмиттеры и белки ядов [34].
лы [35]. Подобная дифференциальная экспрес#
Применение широкомасштабного секвенирова=
сия демонстрирует активное включение каска#
ния для исследования процессов прямого и обрат=
да Wnt в раннем эмбриональном развитии ауре#
ного развития Стрекающих. Молекулярно#гене#
лии. В исследовании развития Hydrozoa, прове#
тические детерминанты особенностей жизнен#
дённом Leclère et al. [144], была охарактеризова#
ного цикла существенно важны для исследова#
на молекулярная структура жизненного цикла
ния процессов развития Стрекающих. Как было
Clytia hemisphaerica с определением специфичес#
описано ранее, некоторые представители
ких маркёрных генов и транскрипционных фак#
Medusozoa обладают возможностью направлять
торов для каждой стадии, а также проведён
своё развитие в обратную сторону (по сравнению
сравнительный анализ процессов развития у
с типичным жизненным циклом) с переходом на
Hydrozoa, Scyphozoa и Anthozoa. Сборка тран#
более раннюю стадию развития. Описание эта#
скриптома de novo и последующий биоинформа#
пов классического пути жизненного цикла и пе#
тический анализ позволили сформировать но#
реключение на «обратное развитие» на уровне
вую высококачественную базу транскриптом#
клеточных процессов представляет особый науч#
ных данных представителя Cubozoa Tripedalia
ный интерес в области биологии развития.
cystophora, которая была использована для иден#
Больше всего молекулярно#генетических
тификации специфических для Стрекающих
исследований проводится в отношении прямого
нейропептидов, что расширило современные
развития Cnidaria. Так, у Scyphozoa на примере
представления о нейроанатомии представите#
A. aurita был охарактеризован процесс перехода
лей Cubozoa [149]. Молекулярные пути и меха#
от стадии полипа к медузе и проведён анализ ре#
низмы, лежащие в основе полового размноже#
гуляции жизненного цикла. Исследователи раз#
ния, были подробно описаны с использованием
делили процесс перехода на 3 этапа: индукция
транскриптомных данных у представителя
метаморфоза понижением температуры, стро#
Anthozoa N. vectensis [92]. Авторы определили
биляция (сегментирование тела полипа с апи#
ключевые процессы, активирующиеся при вы#
кальной части и по направлению к проксималь#
свобождении ооцитов в результате световой и
ной) и развитие новых медуз из отделившихся в
тепловой индукции, к которым относятся свето#
процессе стробиляции сегментов#эфир [147].
восприятие (экспрессия фоторецепторов), орга#
Было установлено, что в процессе стробиляции
низация внеклеточного матрикса, регуляция ак#
происходит выработка молекулярных веществ#
тинового цитоскелета и компоненты циркадных
индукторов метаморфоза, которые концентри#
ритмов. При этом экспрессия компонентов ме#
руются в сегментированной части стробилы и
таболических путей, таких как липидный мета#
отсутствуют у полипов и эфир. По данным тран#
болизм, и компонентов клеточного цикла была
скриптомного секвенирования стадий полипа,
подавлена. Половое размножение у исследуе#
медузы и эфиры, исследователи определили,
мых актиний более чувствительно к температур#
БИОХИМИЯ том 87 вып. 2 2022
242
ЕРОФЕЕВА и др.
ным сдвигам, чем к световым изменениям [92].
но меняется, что является важной характеристи#
Таким образом, высокопроизводительное сек#
кой для изучения регуляторных функций гено#
венирование транскриптомов Стрекающих поз#
ма [151]. Одним из полногеномных методов изу#
воляет подробно описать молекулярно#генети#
чения метилирования ДНК является бисульфит#
ческую структуру развития, а также определить
ное секвенирование, объединяющее разные под#
механизмы развития и влияние условий окружа#
ходы к изучению метилирования, большая часть
ющей среды на жизненный цикл.
из которых основана на бисульфитной конвер#
Эпигенетические подходы к изучению разви=
сии ДНК для обнаружения неметилированных
тия Стрекающих. Применение методов высо#
цитозинов (рис. 2). В процессе подготовки биб#
копроизводительного секвенирования для изу#
лиотеки инкубация с бисульфитом приводит к
чения процессов развития Стрекающих позво#
химической конверсии всех неметилированных
ляет проводить более детальные исследования,
цитозинов на урацил, который в процессе секве#
касающиеся структуры генома и генной
нирования идентифицируется как тимин. При
экспрессии с функциональной аннотацией. По#
последующей обработке высчитывают процент
мимо активно использующихся методов секве#
метилированных цитозинов по данным прочте#
нирования тотальных геномов и транскрипто#
ний после секвенирования [154, 155].
мов, нужно выделить эпигенетические исследо#
Данных по исследованию ДНК#метилирова#
вания, используемые для изучения генетичес#
ния у Стрекающих не так много, однако они де#
ких модификаций, не связанных с изменениями
монстрируют важную роль метилирования в
в первичной последовательности ДНК, и при#
процессах регуляции развития. На примере сим#
менение технологии секвенирования геномов,
биотического модельного организма Aiptasia
транскриптомов и эпигеномов единичных кле#
(Anthozoa) было показано существование генов с
ток в исследуемом образце для структурного и
повышенным уровнем метилирования ДНК по
функционального описания различных типов
CpG#сайтам в кодирующей области гена и пони#
клеток одного организма.
жением уровня метилирования ДНК в областях
Эпигенетические изменения являются вари#
начала и конца транскрипции [157]. Исследова#
антом реакции генома живого организма на ва#
тели предполагают, что такое специфическое ме#
риабельные условия окружающей среды [150].
тилирование играет важную роль для повыше#
В настоящее время изучение эпигенетических
ния точности транскрипции у Aiptasia и поддер#
модификаций ограничено доступными метода#
жания транскрипционного гомеостаза, отвечаю#
ми, такими как анализ ДНК#метилирования,
щего за симбиоз [157]. В то же время нанопоро#
исследования открытых участков хроматина,
вое секвенирование полногеномного профиля
ДНК#белковых взаимодействий и различных
ДНК#метилирования у симбиотического корал#
модификаций гистонов.
ла Anthopleura elegantissima (Anthozoa), который
Механизм ДНК#метилирования, заключаю#
может существовать и как апосимбионт, проде#
щийся в переносе метильной группы в положе#
монстрировало схожие профили метилирования
ние C5 на цитозине с образованием 5#метилци#
у симбиотической водоросли Elliptochloris marina
тозина, регулирует экспрессию генов путём
и у апосимбиотического A. elegantissima, что мо#
привлечения белков#репрессоров или путём ин#
жет быть обусловлено особенностями симбиоза
гибирования связывания транскрипционных
конкретных организмов [158]. У разных предста#
факторов с ДНК [151]. Существует несколько ва#
вителей Anthozoa CpG#метилирование в кодиру#
риантов метилирования в зависимости от поло#
ющей области генов связано с пластичностью
жения цитозина, которые можно разделить на
генной экспрессии в разных средах и популяци#
2 группы. К первой группе относится самый
ях [159], а также с акклиматизацией [160], адап#
распространённый вариант метилирования, воз#
тацией к низким уровням рН [160] и эволюцией
никающий на цитозинах, предшествующих гуа#
кодонов [161]. Данные по метилированию ДНК у
ниновым нуклеотидам (CpG#сайты)
[151].
Hydra ещё нуждаются в функциональной оценке,
Ко второй группе относятся редко встречающи#
однако есть некоторые исследования по тоталь#
еся варианты метилирования не по CpG#сайтам,
ному уровню метилирования ДНК в целом орга#
а на цитозинах, предшествующих другим типам
низме. В геноме Hydra показано низкое содержа#
нуклеотидов (A, T, C), что было детектировано у
ние GC [162], поэтому уровень метилирования в
растений [152]. В геномах беспозвоночных мети#
CpG#сайтах значительно ниже, чем в других об#
лирование ДНК относительно редко встречается
ластях генома. Авторы данного обзора предпола#
в нуклеотидном контексте CG, однако оно часто
гают, что метилирование аденозина в положении
детектируется в активно транскрибируемых ге#
N6 (m6dA) может способствовать началу тран#
нах [153]. В процессе развития живого организма
скрипции, необходимой в непрерывно делящих#
уровень метилирования ДНК в геноме постоян#
ся и дифференцирующихся клетках тела Hydra, а
БИОХИМИЯ том 87 вып. 2 2022
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
243
Рис. 2. Принцип полногеномного бисульфитного секвенирования. Обработка образца ДНК бисульфитом приводит к хи#
мическому преобразованию неметилированных цитозинов в урацилы, которые при проведении амплификации опреде#
ляются как тиминовые основания. Бисульфит#конвертированная ДНК используется для подготовки библиотеки секве#
нирования, которая заключается в лигировании адаптеров, сиквенсных праймеров и амплификации библиотеки. По по#
лученным данным секвенирования проводится анализ сайтов гипер# и гипометилирования по всему геному. Схема - на
основе протокола бисульфитной конверсии набора EZ DNA Methylation#Gold™ Kit («Zymo Research», США). Описание
к рисунку составлено на основе источников [154, 156]
метилирование цитозина в положении C5 может
Следующим важным методом эпигенетичес#
динамически регулироваться в процессе диффе#
ких исследований является изучение доступнос#
ренцировки ооцитов [163]. У представителей эн#
ти хроматина в геноме по непосредственной де#
допаразитов группы Myxo#sporea (Myxozoa), по
текции открытых областей, чувствительных к
данным бисульфитного секвенирования, пол#
нуклеазам, а также областей, в которых наблю#
ностью отсутствует метилирование ДНК по ци#
дается низкая плотность нуклеосом или же они
тозину, что, по мнению авторов, может быть свя#
отсутствуют. Открытый хроматин называют ещё
зано с малым размером генома [164].
«активным» по способности к экспрессии ге#
БИОХИМИЯ том 87 вып. 2 2022
244
ЕРОФЕЕВА и др.
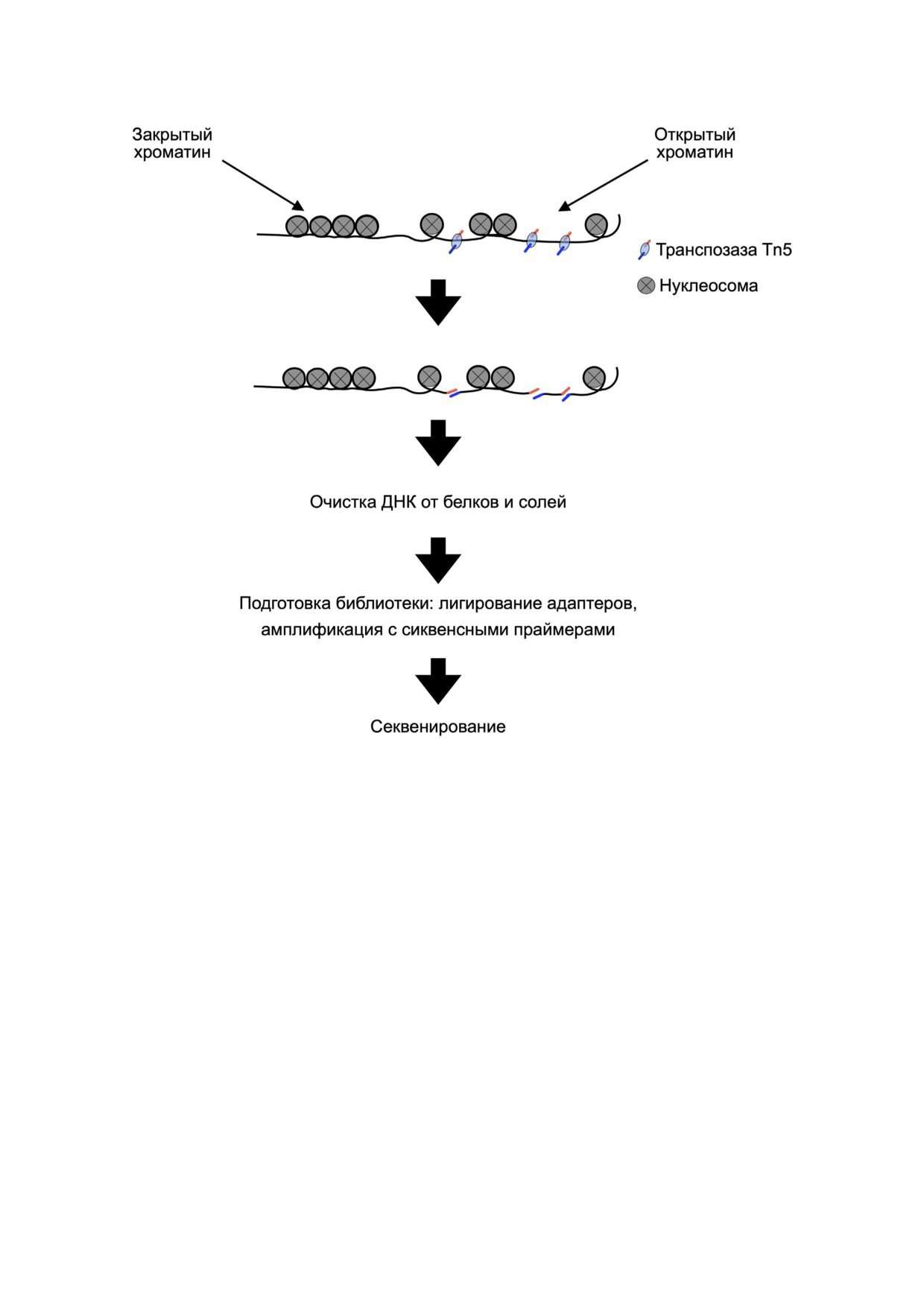
Рис. 3. Принцип метода ATAC#seq (Assay for Transposase#Accessible Chromatin using sequencing). Гиперактивная транспоза#
за Tn5 инкорпорирует специфические адаптеры в области открытого хроматина в месте разреза ДНК. Очищенная фраг#
ментированная по открытым участкам ДНК используется для подготовки библиотеки секвенирования. По полученным
данным секвенирования проводят анализ открытых участков генома, детектируют положение нуклеосом и закономер#
ности расположения транскрипционных факторов [165]
нов. Одним из часто используемых эпигенети#
Исследования с использованием ATAC#seq на
ческих подходов является анализ доступного
N. vectensis продемонстрировали взаимосвязь
для транспозазы хроматина с помощью секве#
открытых участков хроматина и транскрипции
нирования или ATAC#seq (Assay for Transposase#
генов циркадного ритма, изменяющейся в зави#
Accessible Chromatin using sequencing) [165].
симости от режима светового культивирования.
Принцип метода заключается в использовании
По результатам исследования показано, что бо#
высокоактивного фермента транспозазы Tn5,
лее половины промоторов известных циркадных
которая фрагментирует ДНК по открытым
генов находились в участках открытого хромати#
участкам, одновременно интегрируя праймеры
на. Кроме того, в работе были идентифицирова#
для секвенирования (рис. 3) [165].
ны активные энхансеры, специфичные для раз#
Секвенирование библиотеки проводится на
ных режимов светового культивирования [166].
стандартных платформах, например, на плат#
Эксперименты с тепловым воздействием (37 °С)
форме «Illumina», США. Анализ данных позво#
на N. vectensis, выращенную в лабораторных ус#
ляет идентифицировать открытые участки хро#
ловиях и собранную в полевых условиях, пока#
матина в геноме, а также использовать эти дан#
зали, что разные условия выращивания влияют
ные для изучения регуляции экспрессии генов.
на эпигеном N. vectensis, демонстрируя участие
БИОХИМИЯ том 87 вып. 2 2022
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
245
различных транскрипционных факторов и от#
состояние транскрипции генов локального
личный для разных условий транскрипционный
участка генома. К основным хорошо известным
ответ на повышение температуры. Функцио#
типам модификаций относятся: ацетилирование,
нальный анализ состава дифференциально
метилирование, фосфорилирование и убиквити#
экспрессируемых генов позволил определить,
нирование. Существуют также и другие недавно
что в ответ на тепловое воздействие включаются
открытые типы модификаций, например, деими#
разные сигнальные клеточные пути, зависящие
нация или пролиновая изомеризация [174].
от условий культивирования: в полевых услови#
В настоящее время известно большое количество
ях - пути, относящиеся к контрольным точкам
данных по модификациям гистонов и их функ#
клеточного цикла, репликации, репарации ДНК
циям в хроматине [175], однако к основным гис#
и структурированию гомеостаза, а в лаборатор#
тоновым меткам, широко использующимся в
ных условиях - пути, связанные с метаболичес#
эпигенетических исследованиях, относятся: мо#
кими процессами (метаболизмом аминокислот,
нометилирование по четвёртому остатку лизина
амидов), везикулярным транспортом и стабили#
гистона H3 (H3K4me1) и ацетилирование гисто#
зацией белков. Таким образом, данное исследо#
на H3 по лизину 27 (H3K27ac), связанные с эн#
вание демонстрирует прямое воздействие усло#
хансерными областями; метка промоторных ре#
вий окружающей среды на эпигенетическую ре#
гионов - триметилирование гистона H3 по чет#
гуляцию ответа на стрессовые условия [167].
вёртому остатку лизина (H3K4me3); триметили#
Разная эпигеномная регуляция в ответ на тепло#
рование гистона H3 по лизину 36 (H3K36me3) -
вой стресс была показана у вышеупомянутого
метка транскрибируемых участков в теле генов;
симбиотического анемона Aiptasia [168]. Описа#
триметилирование гистона H3 по лизину 27
на эпигенетическая регуляция в процессе реге#
(H3K27me3) - метка транскрипционного реп#
нерации оральной части с щупальцами у Hydra.
рессорного комплекса Polycomb; триметилирова#
По данным ATAC#seq, у Hydra в верхней части
ние гистона H3 по лизину 9 (H3K9me3) - метка
тела на стадии регенерации идентифицировали
гетерохроматина [173].
более 20 000 открытых хроматиновых участков.
Данные ChIP#seq расширили понимание
В сопоставлении с другим эпигенетическим ме#
эпигеномной регуляции регенерации частей те#
тодом ChIP#seq (описанным ниже) удалось уста#
ла Hydra. Как было упомянуто выше, комбини#
новить положение доступных участков хромати#
рование с данными ATAC#seq позволило опреде#
на: более 3000 из них находились в межгенных
лить положение доступных участков хроматина,
областях, более 800 открытых регионов пере#
а также идентифицировать кандидаты активных
крывались с интронами и более 200 участков - с
проксимальных промоторов, энхансерных реги#
экзонами. Таким образом, впервые был пред#
онов и транскрипционных факторов
[169].
ставлен анализ элементов открытого хроматина
В процессе регенерации тканей апикальной час#
в геноме Стрекающих в контексте процесса раз#
ти тела Hydra было отмечено динамическое ре#
вития [169].
моделирование регуляторных элементов, боль#
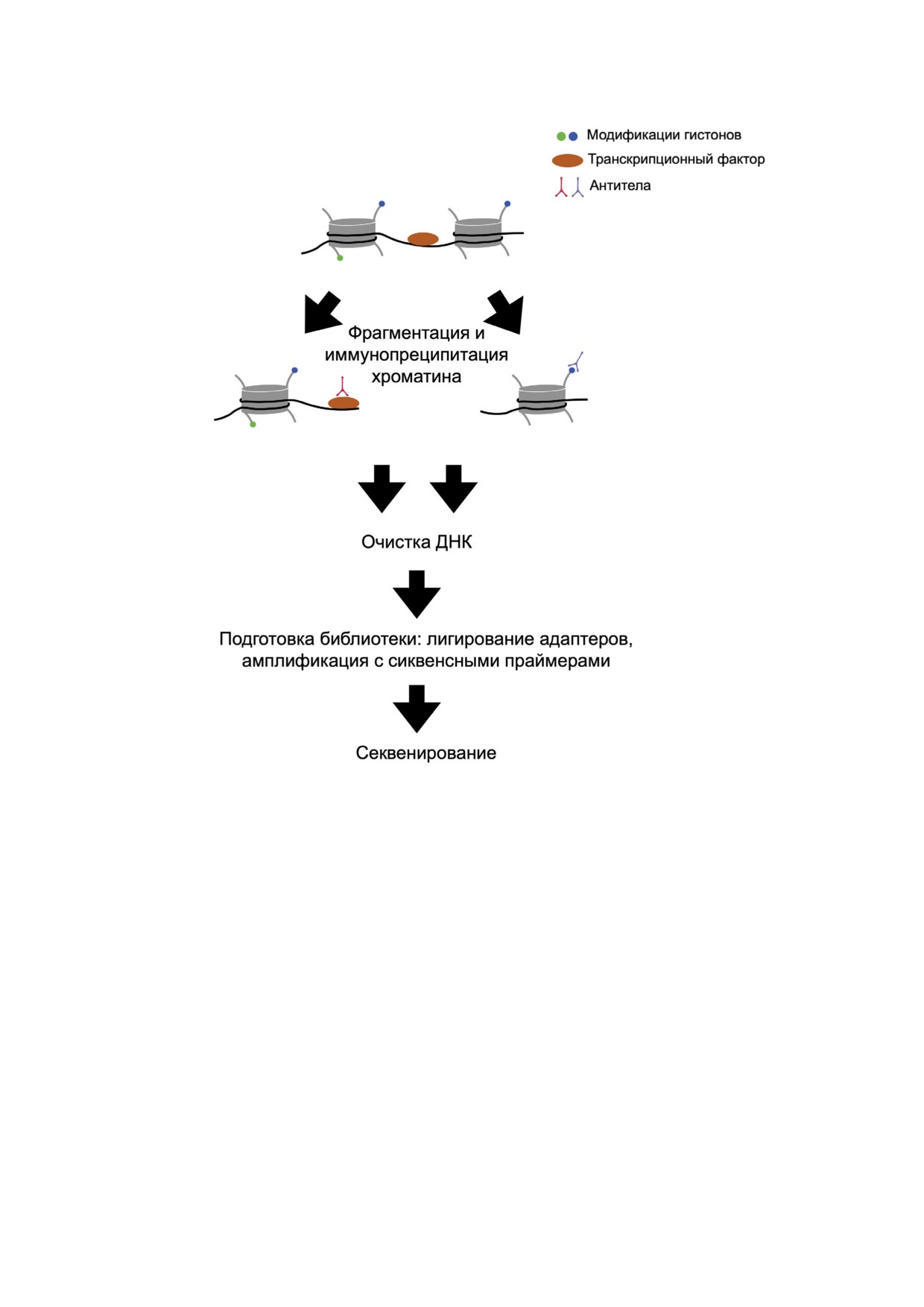
Анализ ДНК#белковых взаимодействий яв#
шинство которых теряло свою доступность при
ляется важнейшим из эпигенетических методов,
регенерации. При этом определённые энхансер#
позволяющим довольно точно детектировать
ные и промоторные области активировались,
регуляторные элементы генома. Одним из рас#
например, один промоторный и два регулятор#
пространённых методов считается ChIP#seq
ных сайта гена Wnt3, который, как известно, за#
(Chromatin immunoprecipitation followed by
действован в организации апикальной части
sequencing) или иммунопреципитация хромати#
Hydra в процессе регенерации [169]. При экто#
на с последующим секвенированием [170].
пической активации пути Wnt были детектиро#
Принцип заключается в использовании специ#
ваны межгенные участки - потенциальные эн#
фических антител, которые при инкубации им#
хансеры рядом с генами ключевых транскрипци#
мунопреципитируют ДНК#связанные белки, за#
онных факторов (Brachyury1, Cngsc, Pitx1) и ли#
тем ДНК осаждается, очищается от белков и
гандов Wnt#пути (Wnt5a, Wnt11), регулирующих
секвенируется (рис. 4). Этот метод позволяет де#
развитие головной части Hydra [176].
тектировать области активных промоторов, сай#
В геноме N. vectensis было идентифицирова#
ты связывания транскрипционных факторов и
но более 5000 энхансерных областей, подтверж#
других регуляторных элементов по всему гено#
дённых также в экспериментах in vivo [177]. Так#
му, а также в сочетании с РНК#секвенировани#
же у N. vectensis была определена ассоциация
ем и ДНК#метилированием устанавливать регу#
транскрипционного кофактора p300 с энхансе#
ляторные сети генов [170, 171].
рами и показана их активация непосредственно
Кроме того, данный метод используется для
в процессе гаструляции и на стадии планулы.
анализа модификаций гистонов, определяющих
Функциональная характеристика генов, распо#
БИОХИМИЯ том 87 вып. 2 2022
246
ЕРОФЕЕВА и др.
Рис. 4. Принцип метода ChIP#seq (Chromatin immunoprecipitation followed by sequencing). Фиксация хроматина, связанная
с обработкой клеток формальдегидом, in vivo приводит к перекрёстному сшиванию ДНК и целевых белков. При изучении
нативного хроматина этот шаг опускается. Далее хроматин фрагментируется ферментативно или ультразвуковой обработ#
кой. Последующая преципитация целевых ДНК#связывающих белков осуществляется путём использования специфич#
ных антител к данным белкам. Очистка ДНК заключается в длительном нагреве и ферментативной обработке протеина#
зами и рибонуклеазами с последующей очисткой на мембранных колонках или путём фенол#хлороформной экстракции.
Готовая ДНК используется для подготовки библиотеки. По полученным данным секвенирования проводят анализ откры#
тых участков генома, положения транскрипционных факторов и сайтов модификаций гистонов [171-173]
ложенных вблизи энхансерных областей, вы#
тия живых организмов, что позволило проводить
явила превалирование генов регуляции тран#
более детальные исследования разных типов кле#
скрипции, сигнальных клеточных путей и про#
ток в пределах одного организма и изучать кле#
цессов развития у N. vectensis. При этом гены,
точную дифференциацию на разных этапах жиз#
кодирующие транскрипционные факторы, бы#
ненного цикла. В данном подразделе будут опи#
ли связаны с несколькими энхансерами чаще,
саны исследования на Стрекающих с использо#
чем гены домашнего хозяйства [177].
ванием секвенирования единичных клеток.
Изучение развития Стрекающих на уровне еди=
Первым и фундаментальным научным ис#
ничных клеток. Методы секвенирования гено#
следованием по секвенированию отдельных
мов, транскриптомов и эпигеномов отдельных
клеток Стрекающих является работа научных
клеток положили начало новой эре молекуляр#
групп Marlow и Tanay [178]. В качестве модель#
но#генетических исследований процессов разви#
ного объекта использовали представителя акти#
БИОХИМИЯ том 87 вып. 2 2022
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
247
ний N. vectensis, на котором были проведены экс#
gans: ретракторные мышцы и гастродермис
перименты по секвенированию транскриптомов
N. vectensis соответствуют мышцам кишечника и
единичных клеток (по#другому называют single
стенки тела C. elegans; недифференцированные
cell RNA#sequencing или scRNA#seq) и открытого
клетки#предшественники N. vectensis - клеткам
хроматина методом ATAC#seq. Для секвенирова#
зародышевой линии и клеткам#предшественни#
ния транскриптомов использовали метод мас#
кам у C. elegans со специфической экспрессией
сивного параллельного секвенирования единич#
генов хромосомной организации, ассоцииро#
ных клеток (MARS#Seq), который заключается в
ванной с митозом, репликацией и клеточной
сортировке клеток с помощью проточного сор#
пролиферацией; нейроны N. vectensis - нейро#
тирующего цитофлуориметра (FACS) и их иммо#
нам C. elegans. Дополнительно исследователи ус#
билизации на луночном планшете (каждая клет#
тановили модули транскрипционных факторов,
ка в отдельной лунке). После лизиса клетки на
регулирующие разнообразие клеток у N. vecten$
полиА#последовательность мРНК отжигается
sis, а также тканеспецифичные регуляторные
олиго(dT)#праймер с уникальным молекуляр#
элементы в геноме N. vectensis [178]. Таким обра#
ным олигонуклеотидным идентификатором
зом, данное исследование демонстрирует новые
(UMI), индивидуальным для каждого праймера
знания об эволюции типов клеток животных и
и лунки, одинаковым для всех праймеров в каж#
тканеспецифической геномной регуляции.
дой лунке клеточным баркодом, адаптером
Исследование на стволовых клетках Hydra с
«Illumina» и промотором Т7. С олиго(dT)#прай#
применением секвенирования транскриптомов
мера начинается обратная транскрипция с обра#
единичных клеток позволило описать разнооб#
зованием первой цепи комплементарной
разные типы клеточных линий и их изменения в
ДНК (кДНК). После этого все клеточные лизаты
процессе дифференцировки [181]. Ученые при#
смешиваются, образуя единую группу для даль#
менили другую технологию секвенирования,
нейшей амплификации. Синтезируется вторая
которая называется «полногеномное профили#
цепь кДНК с последующей транскрипцией. По#
рование экспрессии отдельных клеток с исполь#
лученная РНК фрагментируется, и к ней лигиру#
зованием нанолитровых капель» или Drop#seq
ют адаптер «Illumina» для секвенирования с до#
(Droplet#sequencing) (рис. 6) [182].
полнительным баркодом клеточной смеси
Принцип метода заключается в генерации за
(группы амплификации). После обратной тран#
счёт тонкой микрофлюидики одноклеточной
скрипции готовая баркодированная кДНК ис#
эмульсии, состоящей из водно#масляной капли,
пользуется для подготовки библиотеки для сек#
шариков из смолы с баркодированными прай#
венирования (рис. 5) [179, 180].
мерами и клеток исследуемого образца, таким
Комбинируя транскриптомные и эпигеном#
образом, что все фрагменты от одной клетки
ные данные единичных клеток, исследователям
имеют общий баркод. Полученная эмульсия ис#
удалось создать целый атлас различных типов
пользуется в дальнейшем для создания стандарт#
клеток у взрослого организма N. vectensis: были
ных библиотек с короткими фрагментами для
идентифицированы кластеры разных клеток и
секвенирования на распространённых приборах
гены со специфической для каждого кластера
фирмы
«Illumina». Подготовка библиотеки
экспрессией, то есть маркёрные гены для каж#
включает этап тагментации - фрагментации
дого типа клеток. Например, было показано,
ДНК транспосомами с одновременным лигиро#
что в книдоцитах экспрессируются белки капсу#
ванием адаптерных последовательностей для
лы (миниколлагены, нематогалектины) и
последующей амплификации с сиквенсными
ядов (NEP). Также исследователи охарактеризо#
праймерами «Illumina». После секвенирования
вали разные типы клеток у стадии планулы, от#
специальное программное обеспечение исполь#
метив при этом два плануло#специфических
зует идентификационные баркоды для сопо#
кластера клеток - нейроны и клетки апикально#
ставления прочтений и их количества с их ядром
го органа. Сравнительный анализ репертуаров
или клеткой [182]. Исследователи отсеквениро#
генов, специфичных для разных типов клеток,
вали около 25 000 транскриптомов единичных
показал, что высоко консервативные гены име#
клеток. По результатам анализа все клетки были
ют низкую специфичность к типу клеток, а ге#
разделены на 3 кластера, представляющие от#
ны, кодирующие специфичные белки Стрекаю#
дельные клеточные линии: эктодерму, эндодер#
щих, демонстрируют высокую тканевую специ#
му и интерстициальные клетки. В каждом клас#
фичность. Кроме того, сравнение кластеров
тере были определены свои популяции клеток в
клеток актинии и модельной нематоды
широком диапазоне состояний дифференци#
Caenorhabditis elegans выявило 3 типа клеток
ровки. Так, например кластеры дифференциро#
N. vectensis, схожих по специфической генной
ванных эпителиальных клеток головы и подош#
экспрессии с разными типами клеток у C. ele$
вы связаны с соответствующими кластерами
БИОХИМИЯ том 87 вып. 2 2022
248
ЕРОФЕЕВА и др.
Рис. 5. Принцип метода MARS#seq (Massively parallel single cell RNA#Seq). Особенностью метода является возможность
использования трёхуровневого баркодирования: молекулярной метки (UMI), клеточной метки и метки клеточной смеси
для более точного анализа количества молекул. Метод позволяет учитывать только 3′#транскрипты мРНК с индивидуаль#
ными баркодами. По полученным данным секвенирования анализируют спектр активной мРНК во множестве отдельных
клеток. Описание к рисунку составлено на основе источников [179, 180]
БИОХИМИЯ том 87 вып. 2 2022
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
249
Рис. 6. Принцип метода Drop#seq (Droplets). Формат баркодирования схож с таковым у MARS#seq. Разница заключается
в том, что баркоды прикреплены к микрочастицам (шарикам). Каждая микрочастица содержит множество олиго(dT)#
праймеров с одним и тем же клеточным баркодом (Cell_bc), адаптером «Illumina» (IA1) и разными UMI для цифрового
подсчёта молекул мРНК. Как и в случае MARS#seq, при обработке данных учитываются только 3′#транскрипты. По полу#
ченным данным секвенирования анализируют транскрипты мРНК во множестве отдельных клеток. Схема - на основе
работы Macosko et al. [182] c дополнениями на основе протокола Bageritz и Raddi [183] и протокола подготовки библио#
тек Nextera XT DNA Library Prep Reference Guide («Illumina, Inc.»)
стволовых клеток тела полипа, а кластеры интер#
пространственную экспрессию эпителиальных
стициальных стволовых клеток связаны как с
генов вдоль орально#аборальной оси тела Hydra
предшественниками нейронов, так и с немато#
с определением ранее неохарактеризованных ге#
цитами. Кроме того, были определены кластеры
нов, кодирующих компоненты сигнальных пу#
дифференцированных нейронов, железистых
тей развития (Wnt, BMP и FGF). С помощью де#
клеток, нематоцитов и половых клеток, принад#
тальных исследований интерстициальной линии
лежащих интерстициальной линии [181]. Анализ
клеток исследователи идентифицировали попу#
дифференциальной экспрессии на уровне еди#
ляцию мультипотентных интерстициальных
ничных клеток позволил идентифицировать
стволовых клеток, установив у них низкий уро#
7 БИОХИМИЯ том 87 вып. 2 2022
250
ЕРОФЕЕВА и др.
вень или отсутствие экспрессии генов диффе#
стояние 3 (зрелые клетки с водорослями), а за#
ренцировки. Этот кластер клеток использовали
тем через промежуточное состояние 4 - в состо#
для определения траектории развития интерсти#
яние 5 (постэндосимбиотические клетки). В об#
циальной линии, что выявило дифференциров#
разцах, проходящих регенерацию, обнаружен
ку нейронов и железистых клеток через единое
более высокий процент клеток в состояниях 1
общее клеточное состояние, отличное от пути
и 2, в отличие от не регенерирующих вариантов,
дифференцировки нематоцитов [181]. Примене#
где оказалось больше клеток в состояниях 3-5.
ние метода ATAC#seq и его комбинирование с
Показано, что пре#эндосимбиотические клетки
scRNA#seq позволило определить экспрессию
экспрессируют лиганды молекулярного каска#
транскрипционных факторов, специфичную для
да Wnt (Wnt7b, Wnt11), которые могут быть за#
разных типов клеток и на разных этапах диффе#
действованы в регуляции пролиферации и диф#
ренцировки. Так, например, было показано, что
ференцировки клеток#предшественников [184].
гомеобоксный транскрипционный фактор RX
Данное исследование демонстрирует новую ха#
задействован в развитии базального диска, а
рактеристику эндосимбиоза у кораллов, а также
фактор транскрипции RFX функционирует в
его необходимость в процессе регенерации.
спецификации железистых клеток. Используя
Новые данные по секвенированию единич#
флуоресцентную гибридизацию in situ, научная
ных клеток у представителя Hydrozoa C. hemi$
группа сконструировала подробную молекуляр#
sphaerica показывают пластичность разных типов
ную карту нервной системы Hydra, определив
клеток в условиях голодания [187]. С использо#
12 различных подтипов нейронов [181].
ванием технологии «10X Genomics Ins.» было
Интересное исследование с использованием
определено 36 клеточных популяций, составля#
секвенирования транскриптомов единичных
ющих 8 основных кластеров клеток C. hemi$
клеток демонстрирует молекулярно#генетичес#
sphaerica, соответствующих эпидермису, гастро#
кую структуру эндосимбиоза между восьмилуче#
дермису и производным мультипотентных интер#
вым кораллом Xenia (Anthozoa) и одноклеточны#
стициальных стволовых клеток (ISC). Функцио#
ми динофлагеллятами [184]. Учёные использова#
нальная аннотация кластеров клеток с исполь#
ли технологию scRNA#seq на платформе компа#
зованием уже опубликованных данных scRNA#
нии «10X Genomics Ins.», США. Технология сек#
seq и экспериментальных данных гибридизации
венирования схожа с описанным выше Drop#seq.
in situ генов в целом организме позволила вы#
Исследователи определили 16 кластеров клеток с
явить новые типы клеток - биолюминесцент#
генами#маркёрами для каждого кластера. От#
ные клетки щупалец, экспрессирующие специ#
дельное внимание было уделено идентификации
фические эндогенные зелёные флуоресцентные
клеток, задействованных в эндосимбиозе. Для
белки (GFP), а также эпителиально#мышечные
этого с помощью цитометра были отсортирова#
клетки гладкого и поперечнополосатого типа,
ны клетки с водорослями и клетки без них. На
выстилающие нижний слой колокола и велум,
данных клетках провели тотальное транскрип#
6 подтипов пищеварительных клеток в гастро#
томное секвенирование, и затем полученные
дерме, соответствующие пищеварительным
данные комбинировали с данными scRNA#seq.
участкам в гастроваскулярной системе (желу#
Дополнительное применение гибридизации
док, гонады и бульбусы щупалец), предполагае#
in situ (метода RNAscope) и микроскопии крио#
мые механосенсорные клетки с экспрессией го#
консервированных срезов тканей позволили,
мологов компонентов механосенсорного аппа#
предположительно, установить кластер клеток
рата, характерного для волосковых клеток поз#
гастродермиса, участвующий в эндосимбио#
воночных. Особенностью медузы C. hemisphaeri$
зе [184]. Исследование процесса регенерации
ca является возможность постоянной генерации
эндосимбиотической линии Xenia после физи#
разных типов клеток, особенно нейронов и не#
ческого удаления щупалец с использованием
матоцитов, из пулов ISC эпидермиса бульбусов
вычислительных методов для установления
щупалец. Анализ траекторий дифференцировки
псевдовремени (меры того, как далеко продви#
клеток интерстициальной линии с изучением
нулась клетка в ходе биологического прогрес#
генной экспрессии выявил потенциально спе#
са) [185] и динамики экспрессии (ожидаемого
цифичные для C. hemisphaerica маркёрные гены.
изменения экспрессии генов в ближайшем буду#
Эксперимент с реакцией на стрессовые условия
щем) [186] показало, что эндосимбиотические
в виде голодания показал, что организм не гене#
клетки существуют в пяти динамических состоя#
рирует новые типы клеток в ответ на голодание,
ниях между гомеостатическими условиями и
а формирует ответ за счёт изменения состояния
процессом регенерации: состояние 1 - преэндо#
существующих клеток. Эффект голодания был
симбиотический предшественник, который че#
вариабельным для разных типов клеток, однако
рез промежуточное состояние 2 переходит в со#
наибольшие изменения происходили в клетках
БИОХИМИЯ том 87 вып. 2 2022
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
251
гастроваскулярной системы. Исследователи
этим животным, которые активно используются
составили целую систему из кластеров клеток, в
в качестве различных модельных объектов. Ог#
которых была детектирована дифференциаль#
ромное количество исследований посвящено
ная экспрессия генов в процессе голодания,
изучению процессов раннего развития разнооб#
например генов ферментов оксидоредуктаз, ди#
разных представителей Cnidaria. Использование
оксигеназ или генов лизосомального транспор#
технологий широкомасштабного высокопроиз#
та (Npc2$like), что позволило определить про#
водительного секвенирования поспособствовало
цессы, на которые влияет голодание, как, на#
выявлению эволюционно консервативных меха#
пример метаболизм жирных кислот в клетках
низмов молекулярно#генетического контроля
гастродермиса. Кроме того, высокая экспрессия
этапов раннего развития Стрекающих и возмож#
генов голодания была зафиксирована в ранних
ности их сопоставления с эмбриональным раз#
ооцитах. Таким образом, голодание вызвало ре#
витием высших Metazoa. Развитие технологий
организацию гастродермы и популяции клеток
секвенирования нового поколения в области ге#
ооцитов. При этом изменения в организации
номики, транскриптомики и эпигеномики как
гастродермы и профилей транскрипции, выз#
на уровне тканей, так и единичных клеток позво#
ванные голоданием, опосредовали процессы са#
лило проводить более глубокие исследования
мопереваривания тканей и мобилизацию гаст#
структуры и регуляторных функций генома, осо#
родермальных клеток из гонад [187].
бенностей экспрессии определённых генов и
Технология секвенирования транскриптомов
разных типов клеток в пределах одного организ#
и эпигеномов отдельных клеток является мощ#
ма. Использование методов широкомасштабно#
ным инструментом для изучения процессов раз#
го секвенирования вывело научные знания в об#
вития живых организмов. Метод позволяет не
ласти характеристики особенностей приспособ#
только качественно охарактеризовывать разные
ленности Стрекающих к стрессовым условиям
типы клеток и конструировать карты регулятор#
окружающей среды, изучения жизненных цик#
ных элементов геномов (на примере актинии
лов (в частности, уникального обратного разви#
N. vectensis), но и моделировать траектории раз#
тия), исследования стволовых клеток и механиз#
вития отдельных клеточных линий (например,
мов развития на новый уровень. Эти данные не#
интерстициальных клеток у Hydrozoa). Кроме
сут в себе важную информацию не только с точ#
того, разрешение секвенирования на уровне
ки зрения фундаментального научного интереса,
единичных клеток позволяет более детально
но и с точки зрения развития активно использу#
рассмотреть роль симбиотических отношений
ющихся в прикладных областях технологий ши#
между разными организмами в процессах разви#
рокомасштабного секвенирования.
тия и регенерации (на примере коралла Xenia), а
также установить клеточные и регуляторные ме#
ханизмы процессов, происходящих в отдельных
Финансирование. Исследование выполнено
организмах в ответ на стрессовые условия (на
при поддержке НТУ «Сириус» и Российского
примере гидромедузы C. hemisphaerica).
фонда фундаментальных исследований в рамках
научного проекта № 19#315#51015. Гусев Ф.Е.
(описание эпигенетических подходов) был под#
ЗАКЛЮЧЕНИЕ
держан Научно#технологическим университе#
том «Сириус».
Стрекающие представляют собой уникаль#
Конфликт интересов. Авторы заявляют об от#
ную группу животных с особенными характерис#
сутствии конфликта интересов.
тиками, связанными со строением, эволюцией,
Соблюдение этических норм. Настоящая
жизненными циклами, регенеративными спо#
статья не содержит описания каких#либо иссле#
собностями и процессами развития. Данные осо#
дований с участием людей или животных в каче#
бенности обусловливают научный интерес к
стве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Schierwater, B., Eitel, M., Jakob, W., Osigus, H.#J.,
charge, Toxicon, 54, 1038#1045, doi: 10.1016/j.toxicon.
Hadrys, H., et al. (2009) Concatenated analysis sheds light
2009.03.006.
on early metazoan evolution and fuels a modern “urmeta#
3.
Davy, S. K., Allemand, D., and Weis, V. M. (2012) Cell bio#
zoon” hypothesis, PLoS Biol., 7, e20#e20, doi: 10.1371/
logy of cnidarian#dinoflagellate symbiosis, Microbiol. Mol.
journal.pbio.1000020.
Biol. Rev., 76, 229#261, doi: 10.1128/MMBR.05014#11.
2.
Ozbek, S., Balasubramanian, P. G., and Holstein, T. W.
4.
Goulet, T. L., and Goulet, D. (2021) Climate change leads
(2009) Cnidocyst structure and the biomechanics of dis#
to a reduction in symbiotic derived cnidarian biodiversity
БИОХИМИЯ том 87 вып. 2 2022
7*
252
ЕРОФЕЕВА и др.
on coral reefs, Front. Ecol. Evol., 9, doi: 10.3389/fevo.2021.
22.
Technau, U., and Steele, R. E. (2011) Evolutionary cross#
636279.
roads in developmental biology: Cnidaria, Development,
5.
Horton, T., Kroh, A., Ahyong, S., Bailly, N., Bieler, R.,
138, 1447#1458, doi: 10.1242/dev.048959.
et al. (2021) World Register of Marine Species (WoRMS),
23.
Osadchenko, B. V., and Kraus, Y. A. (2018) Trachylina:
WoRMS Editorial Board.
The group that remains enigmatic despite 150 years of
6.
Diller, W. F. (1940) The Invertebrates: The Invertebrates:
investigations, Russ. J. Dev. Biol.,
49,
134#145,
Protozoa through Ctenophora (Hyman, L. B., ed.) 726 pp.
doi: 10.1134/S1062360418030074.
McGraw#Hill Publications in the Zoological Sciences.
24.
Maronna, M. M., Miranda, T. P., Peña Cantero, Á. L.,
A. Franklin Shull, consulting editor, Science, 92, 219#220,
Barbeitos, M. S., and Marques, A. C. (2016) Towards a
doi: 10.1126/science.92.2384.219.b.
phylogenetic classification of Leptothecata (Cnidaria,
7.
Folino#Rorem, N. C. (2015) Chapter 9 - Phylum Cnidaria.
Hydrozoa), Sci. Rep., 6, 18075, doi: 10.1038/srep18075.
in Thorp and Covich’s Freshwater Invertebrates (Fourth
25.
Piraino, S., Vito, D. D., Schmich, J. R., Bouillon, J. P.,
Edition) (Thorp, J. H., and Rogers, D. C., eds.) Academic
and Boero, F. (2004) Reverse development in Cnidaria,
Press, Boston, pp. 159#179.
Can. J. Zool., 82, 1748#1754, doi: 10.1139/z04#174.
8.
Grimmelikhuijzen, C. J., and Westfall, J. A. (1995) The
26.
Ballesteros, A., Östman, C., Sant n, A., Marambio, M.,
nervous systems of cnidarians, EXS,
72,
7#24,
Narda, M., et al. (2021) Cnidome and morphological fea#
doi: 10.1007/978#3#0348#9219#3_2.
tures of Pelagia noctiluca (Cnidaria: Scyphozoa) through#
9.
Galliot, B., and Quiquand, M. (2011) A two#step process
out the different life cycle stages, Front. Mar. Sci., 8, 1059,
in the emergence of neurogenesis, Eur. J. Neurosci., 34,
doi: 10.3389/fmars.2021.714503.
847#862, doi: 10.1111/j.1460#9568.2011.07829.x.
27.
Piraino, S., Boero, F., Aeschbach, B., and Schmid, V.
10.
Khabibulina, V. R., and Starunov, V. V.
(2020)
(1996) Reversing the life cycle: Medusae transforming into
FMRFamide immunoreactive nervous system in the adult
polyps and cell transdifferentiation in Turritopsis nutricula
Cassiopeia xamachana scyphopolyp and at the early stages
(Cnidaria, Hydrozoa), Biol. Bull.,
190,
302#312,
of planuloid formation, Invertebr. Zool., 17, 371#384,
doi: 10.2307/1543022.
doi: 10.15298/invertzool.17.4.03.
28.
He, J., Zheng, L., Zhang, W., and Lin, Y. (2015) Life Cycle
11.
Rentzsch, F., Juliano, C., and Galliot, B. (2019) Modern
Reversal in Aurelia sp. 1 (Cnidaria, Scyphozoa), PLoS One,
genomic tools reveal the structural and cellular diversity of
10, e0145314, doi: 10.1371/journal.pone.0145314.
cnidarian nervous systems, Curr. Opin. Neurobiol., 56, 87#
29.
Babonis, L. S., and Martindale, M. Q. (2017) Phylogenetic
96, doi: 10.1016/j.conb.2018.12.004.
evidence for the modular evolution of metazoan signalling
12.
Tournière, O., Dolan, D., Richards, G. S., Sunagar, K.,
pathways, Philos. Trans. R. Soc. B Biol. Sci., 372,
Columbus#Shenkar, Y. Y., et al. (2020) NvPOU4/Brain3
20150477, doi: 10.1098/rstb.2015.0477.
Functions as a terminal selector gene in the nervous system
30.
Chapman, J. A., Kirkness, E. F., Simakov, O., Hampson,
of the cnidarian Nematostella vectensis, Cell Rep., 30, 4473#
S. E., Mitros, T., et al. (2010) The dynamic genome of
4489.e4475, doi: 10.1016/j.celrep.2020.03.031.
Hydra, Nature, 464, 592#596, doi: 10.1038/nature08830.
13.
Watanabe, H., Fujisawa, T., and Holstein, T. W. (2009)
31.
Putnam, N. H., Srivastava, M., Hellsten, U., Dirks, B.,
Cnidarians and the evolutionary origin of the nervous sys#
Chapman, J., et al. (2007) Sea anemone genome reveals
tem, Dev. Growth Differ., 51, 167#183, doi: 10.1111/j.1440#
ancestral eumetazoan gene repertoire and genomic orga#
169X.2009.01103.x.
nization, Science, 317, 86#94, doi: 10.1126/science.
14.
Leclère, L., and Röttinger, E. (2016) Diversity of cnidari#
1139158.
an muscles: Function, anatomy, development and regener#
32.
Kanehisa, M., and Goto, S. (2000) KEGG: kyoto encyclo#
ation, Front. Cell Dev. Biol., 4, 157, doi: 10.3389/fcell.
pedia of genes and genomes, Nucleic Acids Res., 28, 27#30,
2016.00157.
doi: 10.1093/nar/28.1.27.
15.
Berzins, I. K., Yanong, R. P. E., LaDouceur, E. E. B., and
33.
Khalturin, K., Shinzato, C., Khalturina, M., Hamada, M.,
Peters, E. C. (2021) Cnidaria, in Invertebrate Histology,
Fujie, M., et al. (2019) Medusozoan genomes inform the
John Wiley & Sons, Inc., Hoboken, New Jersey, pp. 55#86.
evolution of the jellyfish body plan, Nat. Ecol. Evol., 3,
16.
Cole, A. G., Kaul, S., Jahnel, S. M., Steger, J.,
811#822, doi: 10.1038/s41559#019#0853#y.
Zimmerman, B., et al. (2020) Muscle cell type diversifica#
34.
Kim, H. M., Weber, J. A., Lee, N., Park, S. G., Cho, Y. S.,
tion facilitated by extensive gene duplications, bioRxiv,
et al. (2019) The genome of the giant Nomura’s jellyfish
doi: 10.1101/2020.07.19.210658.
sheds light on the early evolution of active predation, BMC
17.
Okamura, B., and Gruhl, A. (2021) Evolution, Origins and
Biol., 17, 28, doi: 10.1186/s12915#019#0643#7.
Diversification of Parasitic Cnidarians, in The Evolution
35.
Brekhman, V., Malik, A., Haas, B., Sher, N., and Lotan, T.
and Fossil Record of Parasitism: Identification and
(2015) Transcriptome profiling of the dynamic life cycle of
Macroevolution of Parasites (De Baets, K., and Huntley,
the scypohozoan jellyfish Aurelia aurita, BMC Genom., 16,
J. W., eds.) Springer International Publishing, Cham,
74, doi: 10.1186/s12864#015#1320#z.
pp. 109#152.
36.
Avila Soria, G. (2009) Molecular characterization of
18.
Chang, E. S., Neuhof, M., Rubinstein, N. D.,
Carukia barnesi and Malo kingi, Cnidaria; Cubozoa;
Diamant, A., Philippe, H., et al. (2015) Genomic insights
Carybdeidae. PhD thesis, James Cook University.
into the evolutionary origin of Myxozoa within Cnidaria,
37.
Ohdera, A., Ames, C. L., Dikow, R. B., Kayal, E.,
Proc. Natl. Acad. Sci. USA,
112,
14912#14917,
Chiodin, M., et al. (2019) Box, stalked, and upside#down?
doi: 10.1073/pnas.1511468112.
Draft genomes from diverse jellyfish (Cnidaria, Acraspeda)
19.
Kayal, E., Bentlage, B., Sabrina Pankey, M., Ohdera,
lineages: Alatina alata (Cubozoa), Calvadosia cruxmeliten$
A. H., Medina, M., et al. (2018) Phylogenomics provides a
sis (Staurozoa), and Cassiopea xamachana (Scyphozoa),
robust topology of the major cnidarian lineages and
Gigascience, 8, giz069, doi: 10.1093/gigascience/giz069.
insights on the origins of key organismal traits, BMC Evol.
38.
Martindale, M. Q., Finnerty, J. R., and Henry, J. Q. (2002)
Biol., 18, 68, doi: 10.1186/s12862#018#1142#0.
The Radiata and the evolutionary origins of the bilaterian
20.
Kayal, E., Roure, B., Philippe, H., Collins, A. G., and
body plan, Mol. Phylogenet. Evol.,
24,
358#365,
Lavrov, D. V. (2013) Cnidarian phylogenetic relationships
doi: 10.1016/S1055#7903(02)00208#7.
as revealed by mitogenomics, BMC Evol. Biol., 13, 5,
39.
Arendt, D., Tosches, M. A., and Marlow, H. (2016) From
doi: 10.1186/1471#2148#13#5.
nerve net to nerve ring, nerve cord and brain - evolution of
21.
Zapata, F., Goetz, F. E., Smith, S. A., Howison, M.,
the nervous system, Nat. Rev. Neurosci., 17, 61#72,
Siebert, S., et al. (2015) Phylogenomic analyses support
doi: 10.1038/nrn.2015.15.
traditional relationships within Cnidaria, PLoS One, 10,
40.
Finnerty, J. R., Pang, K., Burton, P., Paulson, D., and
e0139068, doi: 10.1371/journal.pone.0139068.
Martindale, M. Q. (2004) Origins of bilateral symmetry:
БИОХИМИЯ том 87 вып. 2 2022
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
253
Hox and Dpp expression in a sea anemone, Science, 304,
the anthozoan cnidarian, Nematostella vectensis, is mediat#
1335#1337, doi: doi:10.1126/science.1091946.
ed by Dishevelled, Dev. Biol., 310, 169#186, doi: 10.1016/
41.
Genikhovich, G., and Technau, U. (2017) On the evolu#
j.ydbio.2007.05.040.
tion of bilaterality, Development,
144,
3392#3404,
58.
Trevino, M., Stefanik, D. J., Rodriguez, R., Harmon, S.,
doi: 10.1242/dev.141507.
and Burton, P. M. (2011) Induction of canonical Wnt sig#
42.
He, S., Del Viso, F., Chen, C. Y., Ikmi, A., Kroesen, A. E.,
naling by alsterpaullone is sufficient for oral tissue fate dur#
et al. (2018) An axial Hox code controls tissue segmenta#
ing regeneration and embryogenesis in Nematostella
tion and body patterning in Nematostella vectensis, Science,
vectensis, Dev. Dyn., 240, 2673#2679, doi: 10.1002/dvdy.
361, 1377#1380, doi: 10.1126/science.aar8384.
22774.
43.
Lebedeva, T., Aman, A. J., Graf, T., Niedermoser, I.,
59.
Kirillova, A., Genikhovich, G., Pukhlyakova, E.,
Zimmermann, B., et al. (2021) Cnidarian#bilaterian com#
Demilly, A., Kraus, Y., et al. (2018) Germ#layer commit#
parison reveals the ancestral regulatory logic of the beta#
ment and axis formation in sea anemone embryonic cell
catenin dependent axial patterning, Nat. Commun., 12,
aggregates, Proc. Natl. Acad. Sci. USA, 115, 1813#1818,
4032, doi: 10.1038/s41467#021#24346#8.
doi: 10.1073/pnas.1711516115.
44.
DuBuc, T. Q., Stephenson, T. B., Rock, A. Q., and
60.
Ryan, J. F., Mazza, M. E., Pang, K., Matus, D. Q.,
Martindale, M. Q. (2018) Hox and Wnt pattern the prima#
Baxevanis, A. D., et al. (2007) Pre#bilaterian origins of the
ry body axis of an anthozoan cnidarian before gastrulation,
Hox cluster and the Hox code: Evidence from the sea
Nat. Commun., 9, 2007, doi: 10.1038/s41467#018#04184#x.
anemone, Nematostella vectensis, PLoS One, 2, e153,
45.
Momose, T., Derelle, R., and Houliston, E.
(2008)
doi: 10.1371/journal.pone.0000153.
A maternally localised Wnt ligand required for axial pat#
61.
Genikhovich, G., Fried, P., Prünster, M. M., Schinko,
terning in the cnidarian Clytia hemisphaerica, Development,
J. B., Gilles, A. F., et al. (2015) Axis patterning by BMPs:
135, 2105#2113, doi: 10.1242/dev.021543.
Cnidarian network reveals evolutionary constraints, Cell
46.
Plickert, G., Jacoby, V., Frank, U., Muller, W. A., and
Rep., 10, 1646#1654, doi: 10.1016/j.celrep.2015.02.035.
Mokady, O. (2006) Wnt signaling in hydroid development:
62.
Matus, D. Q., Magie, C. R., Pang, K., Martindale, M. Q.,
formation of the primary body axis in embryogenesis and
and Thomsen, G. H. (2008) The Hedgehog gene family of
its subsequent patterning, Dev. Biol., 298, 368#378,
the cnidarian, Nematostella vectensis, and implications for
doi: 10.1016/j.ydbio.2006.06.043.
understanding metazoan Hedgehog pathway evolution,
47.
Hobmayer, B., Rentzsch, F., Kuhn, K., Happel, C. M.,
Dev. Biol., 313, 501#518, doi: 10.1016/j.ydbio.2007.09.032.
von Laue, C. C., et al. (2000) WNT signalling molecules
63.
Saina, M., Genikhovich, G., Renfer, E., and Technau, U.
act in axis formation in the diploblastic metazoan Hydra,
(2009) BMPs and chordin regulate patterning of the direc#
Nature, 407, 186#189, doi: 10.1038/35025063.
tive axis in a sea anemone, Proc. Natl. Acad. Sci. USA, 106,
48.
Broun, M., Gee, L., Reinhardt, B., and Bode, H. R.
18592#18597, doi: 10.1073/pnas.0900151106.
(2005) Formation of the head organizer in hydra involves
64.
Leclère, L., and Rentzsch, F. (2014) RGM regulates BMP#
the canonical Wnt pathway, Development, 132, 2907#2916,
mediated secondary axis formation in the sea anemone
doi: 10.1242/dev.01848.
Nematostella vectensis, Cell Rep.,
9,
1921#1930,
49.
Philipp, I., Aufschnaiter, R., Ozbek, S., Pontasch, S.,
doi: 10.1016/j.celrep.2014.11.009.
Jenewein, M., et al. (2009) Wnt/beta#catenin and non#
65.
Kraus, Y., Aman, A., Technau, U., and Genikhovich, G.
canonical Wnt signaling interact in tissue evagination in the
(2016) Pre#bilaterian origin of the blastoporal axial orga#
simple eumetazoan Hydra, Proc. Natl. Acad. Sci. USA, 106,
nizer, Nat. Commun.,
7,
11694, doi:
10.1038/
4290#4295, doi: 10.1073/pnas.0812847106.
ncomms11694.
50.
Lengfeld, T., Watanabe, H., Simakov, O., Lindgens, D.,
66.
Momose, T., and Schmid, V. (2006) Animal pole determi#
Gee, L., et al. (2009) Multiple Wnts are involved in Hydra
nants define oral-aboral axis polarity and endodermal cell#
organizer formation and regeneration, Dev. Biol., 330, 186#
fate in hydrozoan jellyfish Podocoryne carnea, Dev. Biol.,
199, doi: 10.1016/j.ydbio.2009.02.004.
292, 371#380, doi: 10.1016/j.ydbio.2006.01.012.
51.
Nakamura, Y., Tsiairis, C. D., Özbek, S., and Holstein,
67.
Duffy, D. J., Plickert, G., Kuenzel, T., Tilmann, W., and
T. W. (2011) Autoregulatory and repressive inputs localize
Frank, U. (2010) Wnt signaling promotes oral but sup#
Hydra Wnt3 to the head organizer, Proc. Natl. Acad. Sci.
presses aboral structures in Hydractinia metamorphosis and
USA, 108, 9137#9142, doi: 10.1073/pnas.1018109108.
regeneration, Development, 137, 3057#3066, doi: 10.1242/
52.
Reddy, P. C., Gungi, A., Ubhe, S., Pradhan, S. J.,
dev.046631.
Kolte, A., et al. (2019) Molecular signature of an ancient
68.
Momose, T., and Houliston, E. (2007) Two oppositely
organizer regulated by Wnt/β#catenin signalling during
localised frizzled RNAs as axis determinants in a cnidarian
primary body axis patterning in Hydra, Commun. Biol., 2,
embryo, PLoS Biol., 5, e70, doi: 10.1371/journal.pbio.
1#11, doi: 10.1038/s42003#019#0680#3.
0050070.
53.
Wang, R., Steele, R. E., and Collins, E.#M. S. (2020) Wnt
69.
Technau, U. (2020) Gastrulation and germ layer formation
signaling determines body axis polarity in regenerating
in the sea anemone Nematostella vectensis and other
Hydra tissue fragments, Dev. Biol.,
467,
88#94,
cnidarians, Mech. Dev., 163, 103628, doi: 10.1016/j.mod.
doi: 10.1016/j.ydbio.2020.08.012.
2020.103628.
54.
Moneer, J., Siebert, S., Krebs, S., Cazet, J., Prexl, A., et al.
70.
Kraus, Y. A., and Markov, A. V. (2017) Gastrulation in
(2021) Differential gene regulation in DAPT#treated
Cnidaria: The key to an understanding of phylogeny or the
Hydra reveals candidate direct Notch signalling targets,
chaos of secondary modifications? Biol. Bull. Rev., 7, 7#25,
J. Cell Sci., 134, jcs258768, doi: 10.1242/jcs.258768.
doi: 10.1134/S2079086417010029.
55.
Kusserow, A., Pang, K., Sturm, C., Hrouda, M.,
71.
Röttinger, E., Dahlin, P., and Martindale, M. Q. (2012) A
Lentfer, J., et al. (2005) Unexpected complexity of the Wnt
framework for the establishment of a cnidarian gene regu#
gene family in a sea anemone, Nature, 433, 156#160,
latory network for “endomesoderm” specification: The
doi: 10.1038/nature03158.
inputs of β#catenin/TCF signaling, PLoS Genet., 8,
56.
Wikramanayake, A. H., Hong, M., Lee, P. N., Pang, K.,
e1003164, doi: 10.1371/journal.pgen.1003164.
Byrum, C. A., et al. (2003) An ancient role for nuclear β#
72.
Chen, C. Y., McKinney, S. A., Ellington, L. R., and
catenin in the evolution of axial polarity and germ layer
Gibson, M. C. (2020) Hedgehog signaling is required for
segregation, Nature,
426,
446#450, doi:
10.1038/
endomesodermal patterning and germ cell development in
nature02113.
the sea anemone Nematostella vectensis, Elife, 9, e54573,
57.
Lee, P. N., Kumburegama, S., Marlow, H. Q., Martindale,
doi: 10.7554/eLife.54573.
M. Q., and Wikramanayake, A. H. (2007) Asymmetric
73.
Wijesena, N. M. (2012) Wnt Signaling in the Cnidarian
developmental potential along the animal-vegetal axis in
Nematostella vectensis: Insighits into the Evolution of
БИОХИМИЯ том 87 вып. 2 2022
254
ЕРОФЕЕВА и др.
Gastrulation. PhD thesis, University of Miami, Coral
90. Eckelbarger, K. J., Hand, C., and Uhlinger, K. R. (2008)
Gables.
Ultrastructural features of the trophonema and oogenesis
74.
Leclère, L., Bause, M., Sinigaglia, C., Steger, J., and
in the starlet sea anemone, Nematostella vectensis
Rentzsch, F. (2016) Development of the aboral domain in
(Edwardsiidae), Invertebrate Biol.,
127,
381#395,
Nematostella requires β#catenin and the opposing activities
doi: 10.1111/j.1744#7410.2008.00146.x.
of Six3/6 and Frizzled5/8, Development, 143, 1766#1777,
91. Бочарова Е. С., Косевич И. А. (2011) Варианты раз#
doi: 10.1242/dev.120931.
множения актиний (CNIDARIA, ANTHOZOA), Зо$
75.
Wijesena, N., Sun, H., Kumburegama, S., and
ологический журнал, 90, 1283#1295.
Wikramanayake, A. H. (2021) Distinct Frizzled receptors
92. Reuven, S., Rinsky, M., Brekhman, V., Malik, A.,
independently mediate endomesoderm specification and
Levy, O., et al. (2021) Cellular pathways during spawning
primary arch on invagination during gastrulation in
induction in the starlet sea anemone Nematostella vectensis,
Nematostella, Dev. Biol., 481, 215#225, doi: 10.1016/
Sci. Rep., 11, 15451, doi: 10.1038/s41598#021#95033#3.
j.ydbio.2021.11.002.
93. Chiu, Y.#L., Shikina, S., Yoshioka, Y., Shinzato, C., and
76.
Vetrova, A. A., Lebedeva, T. S., Saidova, A. A., Kupaeva,
Chang, C.#F. (2020) De novo transcriptome assembly from
D. M., Kraus, Y. A., et al. (2021) From apolar gastrula to
the gonads of a scleractinian coral, Euphyllia ancora: mol#
polarized larva: Embryonic development of a marine
ecular mechanisms underlying scleractinian gametogene#
hydroid, Dynamena pumila, Dev. Dyn., 1=31, doi: 10.1002/
sis, BMC Genomics, 21, 732, doi: 10.1186/s12864#020#
dvdy.439.
07113#9.
77.
Kraus, Y., Flici, H., Hensel, K., Plickert, G., Leitz, T., and
94. Rentzsch, F., Layden, M., and Manuel, M. (2017) The cel#
Frank, U. (2014) The embryonic development of the
lular and molecular basis of cnidarian neurogenesis,
cnidarian Hydractinia echinata, Evol Dev, 16, 323#338,
WIREs Dev. Biol., 6, e257, doi: 10.1002/wdev.257.
doi: 10.1111/ede.12100.
95. Nakanishi, N., Renfer, E., Technau, U., and Rentzsch, F.
78.
Momose, T., Kraus, Y., and Houliston, E. (2012) A con#
(2012) Nervous systems of the sea anemone Nematostella
served function for Strabismus in establishing planar cell
vectensis are generated by ectoderm and endoderm and
polarity in the ciliated ectoderm during cnidarian larval
shaped by distinct mechanisms, Development, 139, 347#
development,
Development,
139,
4374#4382,
357, doi: 10.1242/dev.071902.
doi: 10.1242/dev.084251.
96. Al#Shaer, L., Havrilak, J., and Layden, M.
(2021)
79.
Fröbius, A. C., Genikhovich, G., Kürn, U., Anton#
Nematostella vectensis as a Model System. in Handbook of
Erxleben, F., and Bosch, T. C. G. (2003) Expression of
Marine Model Organisms in Experimental Biology, CRC
developmental genes during early embryogenesis of Hydra,
Press, Boca Raton, pp. 107#128.
Dev. Genes Evol., 213, 445#455, doi: 10.1007/s00427#003#
97. Yuan, D., Nakanishi, N., Jacobs, D. K., and
0344#6.
Hartenstein, V. (2008) Embryonic development and meta#
80.
Siebert, S., and Juliano, C. E. (2017) Sex, polyps, and
morphosis of the scyphozoan Aurelia, Dev. Genes Evol.,
medusae: Determination and maintenance of sex in
218, 525#539, doi: 10.1007/s00427#008#0254#8.
cnidarians, Mol. Reprod. Dev., 84, 105#119, doi: 10.1002/
98. Galliot, B., Quiquand, M., Ghila, L., de Rosa, R.,
mrd.22690.
Miljkovic#Licina, M., et al. (2009) Origins of neurogene#
81.
Technau, U., Miller, M. A., Bridge, D., and Steele, R. E.
sis, a cnidarian view, Dev. Biol.,
332,
2#24,
(2003) Arrested apoptosis of nurse cells during Hydra
doi: 10.1016/j.ydbio.2009.05.563.
oogenesis and embryogenesis, Dev. Biol., 260, 191#206,
99. Gahan, J. M., Schnitzler, C. E., DuBuc, T. Q., Doonan,
doi: 10.1016/S0012#1606(03)00241#0.
L. B., Kanska, J., et al. (2017) Functional studies on the
82.
Айзенштадт Т. Б. (1980) Исследование оогенеза у гид#
role of Notch signaling in Hydractinia development,
ры. IV. Фагоцитарная активность ооцитов, Онтогенез,
Dev. Biol., 428, 224#231, doi: 10.1016/j.ydbio.2017.06.006.
11, 31#38.
100. Wang, W. (2013) Regulation of metamorphosis and the evolu$
83.
Rentzsch, F., Hobmayer, B., and Holstein, T. W. (2005)
tion of life cycles: insights from the common moon jelly A.
Glycogen synthase kinase 3 has a proapoptotic function in
aurita. PhD thesis, Christian#Albrechts#Universität zu
Hydra gametogenesis, Dev. Biol., 278, 1#12, doi: 10.1016/
Kiel, Kiel.
j.ydbio.2004.10.007.
101. Watanabe, H., Kuhn, A., Fushiki, M., Agata, K.,
84.
Kasbauer, T., Towb, P., Alexandrova, O., David, C. N.,
Özbek, S., et al. (2014) Sequential actions of β#catenin and
Dall’armi, E., et al. (2007) The Notch signaling pathway in
Bmp pattern the oral nerve net in Nematostella vectensis,
the cnidarian Hydra, Dev. Biol.,
303,
376#390,
Nat. Commun., 5, 5536, doi: 10.1038/ncomms6536.
doi: 10.1016/j.ydbio.2006.11.022.
102. Duffy, D. J., and Frank, U. (2011) Modulation of COUP#
85.
Hobmayer, B., Rentzsch, F., and Holstein, T. W. (2001)
TF expression in a cnidarian by ectopic Wnt signalling and
Identification and expression of HySmad1, a member of
allorecognition, PLoS One, 6, e19443, doi: 10.1371/journal.
the R#Smad family of TGFbeta signal transducers, in the
pone.0019443.
diploblastic metazoan Hydra, Dev. Genes Evol., 211, 597#
103. Matus, D. Q., Thomsen, G. H., and Martindale, M. Q.
602, doi: 10.1007/s00427#001#0198#8.
(2007) FGF signaling in gastrulation and neural develop#
86.
Miller, M. A., and Steele, R. E. (2000) Lemon encodes an
ment in Nematostella vectensis, an anthozoan cnidarian,
unusual receptor protein#tyrosine kinase expressed during
Dev. Genes Evol., 217, 137#148, doi: 10.1007/s00427#006#
gametogenesis in Hydra, Dev. Biol.,
224,
286#298,
0122#3.
doi: 10.1006/dbio.2000.9786.
104. Rentzsch, F., Fritzenwanker, J. H., Scholz, C. B., and
87.
Amiel, A., and Houliston, E. (2009) Three distinct RNA
Technau, U. (2008) FGF signalling controls formation of
localization mechanisms contribute to oocyte polarity
the apical sensory organ in the cnidarian Nematostella
establishment in the cnidarian Clytia hemisphaerica,
vectensis, Development, 135, 1761#1769, doi: 10.1242/dev.
Dev. Biol., 327, 191#203, doi: 10.1016/j.ydbio.2008.12.
020784.
007.
105. Layden, M. J., Johnston, H., Amiel, A. R., Havrilak, J.,
88.
Houliston, E., Leclère, L., Munro, C., Copley, R., and
Steinworth, B., et al. (2016) MAPK signaling is necessary
Momose, T. (2021) Past, present and future of Clytia hemis$
for neurogenesis in Nematostella vectensis, BMC Biol., 14,
phaerica as a laboratory jellyfish, hal#03346217.
61, doi: 10.1186/s12915#016#0282#1.
89.
Sanders, S. M., Shcheglovitova, M., and Cartwright, P.
106. Turwankar, A., and Ghaskadbi, S. (2019) VEGF and FGF
(2014) Differential gene expression between functionally
signaling during head regeneration in hydra, Gene, 717,
specialized polyps of the colonial hydrozoan Hydractinia
144047, doi: 10.1016/j.gene.2019.144047.
symbiolongicarpus (Phylum Cnidaria), BMC Genomics, 15,
107. Fujita, S., Kuranaga, E., and Nakajima, Y.#I.
(2021)
406, doi: 10.1186/1471#2164#15#406.
Regeneration potential of jellyfish: Cellular mechanisms
БИОХИМИЯ том 87 вып. 2 2022
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
255
and molecular insights, Genes, 12, 758, doi: 10.3390/
125. Hou, S., Zhu, J., Shibata, S., Nakamoto, A., and
genes12050758.
Kumano, G. (2021) Repetitive accumulation of interstitial
108. Röttinger, E. (2021) Nematostella vectensis, an emerging
cells generates the branched structure of Cladonema
model for deciphering the molecular and cellular mecha#
medusa tentacles, Development, 148, doi: 10.1242/dev.
nisms underlying whole#body regeneration, Cells, 10,
199544.
2692, doi: 10.3390/cells10102692.
126. DuBuc, T. Q., Traylor#Knowles, N., and Martindale,
109. Vogg, M. C., Buzgariu, W., Suknovic, N. S., and
M. Q. (2014) Initiating a regenerative response; cellular
Galliot, B. (2021) Cellular, metabolic, and developmental
and molecular features of wound healing in the cnidarian
dimensions of whole#body regeneration in Hydra, Cold
Nematostella
vectensis,
BMC Biol.,
12,
24,
Spring Harb. Perspect. Biol., 13, doi: 10.1101/cshperspect.
doi: 10.1186/1741#7007#12#24.
a040725.
127. Stewart, Z. K., Pavasovic, A., Hock, D. H., and Prentis,
110. Chera, S., Ghila, L., Dobretz, K., Wenger, Y., Bauer, C.,
P. J. (2017) Transcriptomic investigation of wound healing
et al. (2009) Apoptotic cells provide an unexpected source
and regeneration in the cnidarian Calliactis polypus, Sci.
of Wnt3 signaling to drive hydra head regeneration, Dev.
Rep., 7, 41458, doi: 10.1038/srep41458.
Cell, 17, 279#289, doi: 10.1016/j.devcel.2009.07.014.
128. Luz, B. L. P., Miller, D. J., and Kitahara, M. V. (2021)
111.
Gufler, S., Artes, B., Bielen, H., Krainer, I., Eder, M. K.,
High regenerative capacity is a general feature within colo#
et al. (2018) β#Catenin acts in a position#independent
nial dendrophylliid corals (Anthozoa, Scleractinia), J. Exp.
regeneration response in the simple eumetazoan Hydra,
Zool. B Mol. Dev. Evol., 336, 281#292, doi: 10.1002/jez.
Dev. Biol., 433, 310#323, doi: 10.1016/j.ydbio.2017.09.005.
b.23021.
112. Vogg, M. C., Beccari, L., Iglesias Ollé, L., Rampon, C.,
129. Van der Burg, C. A., and Prentis, P. J.
(2021)
Vriz, S., et al. (2019) An evolutionarily#conserved Wnt3/β#
The Tentacular Spectacular: Evolution of regeneration in
catenin/Sp5 feedback loop restricts head organizer activity
sea anemones, Genes,
12,
1072, doi:
10.3390/
in Hydra, Nat. Commun., 10, 312, doi: 10.1038/s41467#
genes12071072.
018#08242#2.
130. Böttger, A., and Hassel, M. (2012) Hydra, a model system
113. Technau, U., Cramer von Laue, C., Rentzsch, F., Luft, S.,
to trace the emergence of boundaries in developing
Hobmayer, B., et al. (2000) Parameters of self#organization
eumetazoans, Int. J. Dev. Biol., 56, 583#591, doi: 10.1387/
in Hydra aggregates, Proc. Natl. Acad. Sci. USA, 97, 12127#
ijdb.113454ab.
12131, doi: 10.1073/pnas.97.22.12127.
131. Mortzfeld, B. M., Taubenheim, J., Klimovich, A. V.,
114. Ferenc, J., Papasaikas, P., Ferralli, J., Nakamura, Y.,
Fraune, S., Rosenstiel, P., et al. (2019) Temperature and
Smallwood, S., et al. (2020) Wnt3 expression as a readout
insulin signaling regulate body size in Hydra by the Wnt
of tissue stretching during Hydra regeneration, bioRxiv,
and TGF#beta pathways, Nat. Commun., 10, 3257,
doi: 10.1101/2020.12.22.423911.
doi: 10.1038/s41467#019#11136#6.
115. Tursch, A., Bartsch, N., and Holstein, T. W. (2020) MAPK
132. Seipel, K., and Schmid, V. (2006) Mesodermal anatomies
signaling links the injury response to Wnt#regulated pat#
in cnidarian polyps and medusae, Int. J. Dev. Biol., 50,
terning in Hydra regeneration, bioRxiv, doi: 10.1101/
589#599, doi: 10.1387/ijdb.062150ks.
2020.07.06.189795.
133. Pennati, R., Dell’Anna, A., Pagliara, P., Scar , G.,
116. Chera, S., Ghila, L., Wenger, Y., and Galliot, B. (2011)
Piraino, S., et al. (2013) Neural system reorganization dur#
Injury#induced activation of the MAPK/CREB pathway
ing metamorphosis in the planula larva of Clava multicornis
triggers apoptosis#induced compensatory proliferation in
(Hydrozoa, Cnidaria), Zoomorphology, 132, 227#237,
hydra head regeneration, Dev. Growth Differ., 53, 186#201,
doi: 10.1007/s00435#013#0188#1.
doi: 10.1111/j.1440#169X.2011.01250.x.
134. Seipp, S., Schmich, J., Will, B., Schetter, E., Plickert, G.,
117. Münder, S., Tischer, S., Grundhuber, M., Büchels, N.,
et al. (2010) Neuronal cell death during metamorphosis of
Bruckmeier, N., et al. (2013) Notch#signalling is required
Hydractina echinata (Cnidaria, Hydrozoa), Invertebrate
for head regeneration and tentacle patterning in Hydra,
Neurosci., 10, 77#91, doi: 10.1007/s10158#010#0109#7.
Dev. Biol., 383, 146#157, doi: 10.1016/j.ydbio.2013.08.022.
135. Stumpf, M., Will, B., Wittig, K., Kasper, J., Fischer, B.,
118. Wenger, Y., Buzgariu, W., Perruchoud, C., Loichot, G.,
et al. (2010) An organizing region in metamorphosing
and Galliot, B. (2019) Generic and context#dependent
hydrozoan planula larvae - stimulation of axis formation
gene modulations during Hydra whole body regeneration,
in both larval and in adult tissue, Int. J. Dev. Biol., 54, 795#
bioRxiv, doi: 10.1101/587147.
802, doi: 10.1387/ijdb.082738ms.
119. Kaloulis, K. (2001) Molecular Basis of Morphogenetic
136. Krasovec, G., Pottin, K., Rosello, M., Quéinnec, É., and
Events in Hydra: Study of the CREB and Hedgehog Pathways
Chambon, J.#P. (2021) Apoptosis and cell proliferation
during Budding and Regeneration. PhD thesis, Université de
during metamorphosis of the planula larva of Clytia hemi$
Genève, Genève.
sphaerica (Hydrozoa, Cnidaria), Dev. Dyn., 250, 1739#
120. Bradshaw, B., Thompson, K., and Frank, U. (2015)
1758, doi: 10.1002/dvdy.376.
Distinct mechanisms underlie oral vs aboral regeneration
137. Seipel, K., Eberhardt, M., Müller, P., Pescia, E.,
in the cnidarian Hydractinia echinata, eLife, 4, e05506,
Yanze, N., et al. (2004) Homologs of vascular endothelial
doi: 10.7554/eLife.05506.
growth factor and receptor, VEGF and VEGFR, in the jel#
121. Sinigaglia, C., Peron, S., Eichelbrenner, J., Chevalier, S.,
lyfish Podocoryne carnea, Dev. Dyn., 231, 303#312,
Steger, J., et al. (2020) Pattern regulation in a regenerating
doi: 10.1002/dvdy.20139.
jellyfish, Elife, 9, e54868, doi: 10.7554/eLife.54868.
138. Sanders, S. M., and Cartwright, P. (2015) Patterns of Wnt
122. Cartwright, P., Travert, M. K., and Sanders, S. M. (2021)
signaling in the life cycle of Podocoryna carnea and its
The evolution and development of coloniality in hydro#
implications for medusae evolution in Hydrozoa
zoans, J. Exp. Zool. B Mol. Dev. Evol., 336, 293#299,
(Cnidaria), Evol. Dev., 17, 325#336, doi: 10.1111/ede.
doi: 10.1002/jez.b.22996.
12165.
123. Bagaeva, T. S., Kupaeva, D. M., Vetrova, A. A., Kosevich,
139. Nawrocki, A. M., and Cartwright, P. (2013) Expression of
I. A., Kraus, Y. A., et al. (2019) cWnt signaling modulation
Wnt pathway genes in polyps and medusa#like structures of
results in a change of the colony architecture in a hydro#
Ectopleura larynx (Cnidaria: Hydrozoa), Evol. Dev., 15,
zoan, Dev. Biol., 456, 145#153, doi: 10.1016/j.ydbio.
373#384, doi: 10.1111/ede.12045.
2019.08.019.
140. Gold, D. A., Nakanishi, N., Hensley, N. M.,
124. Muller, W., Frank, U., Teo, R., Mokady, O., Guette, C.,
Hartenstein, V., and Jacobs, D. K. (2016) Cell tracking
et al. (2007) Wnt signaling in hydroid development: ectopic
supports secondary gastrulation in the moon jellyfish
heads and giant buds induced by GSK#3beta inhibitors,
Aurelia, Dev. Genes Evol., 226, 383#387, doi: 10.1007/
Int. J. Dev. Biol., 51, 211#220, doi: 10.1387/ijdb.62247wm.
s00427#016#0559#y.
БИОХИМИЯ том 87 вып. 2 2022
256
ЕРОФЕЕВА и др.
141. Rentzsch, F., Anton, R., Saina, M., Hammerschmidt, M.,
158. Dimond, J. L., Nguyen, N., and Roberts, S. B. (2021)
Holstein, T. W., et al. (2006) Asymmetric expression of the
DNA methylation profiling of a cnidarian#algal symbiosis
BMP antagonists chordin and gremlin in the sea anemone
using nanopore sequencing, G3 (Bathesda), 11, jkab148,
Nematostella vectensis: Implications for the evolution of
doi: 10.1093/g3journal/jkab148.
axial patterning, Dev. Biol., 296, 375#387, doi: 10.1016/
159. Dixon, G. B., Bay, L. K., and Matz, M. V. (2014) Bimodal
j.ydbio.2006.06.003.
signatures of germline methylation are linked with gene
142. Strader, M. E., Aglyamova, G. V., and Matz, M. V. (2018)
expression plasticity in the coral Acropora millepora, BMC
Molecular characterization of larval development from fer#
Genomics, 15, 1109, doi: 10.1186/1471#2164#15#1109.
tilization to metamorphosis in a reef#building coral, BMC
160. Dixon, G., Liao, Y., Bay, L. K., and Matz, M. V. (2018)
Genom., 19, 17, doi: 10.1186/s12864#017#4392#0.
Role of gene body methylation in acclimatization and
143. Steele, R. E. (2002) Developmental signaling in Hydra:
adaptation in a basal metazoan, Proc. Natl. Acad. Sci. USA,
What does it take to build a “simple” animal? Dev. Biol.,
115, 13342#13346, doi: 10.1073/pnas.1813749115.
248, 199#219, doi: doi.org/10.1006/dbio.2002.0744.
161. Dixon, G. B., Bay, L. K., and Matz, M. V. (2016)
144. Leclère, L., Horin, C., Chevalier, S., Lapebie, P., Dru, P.,
Evolutionary consequences of DNA methylation in a basal
et al. (2019) The genome of the jellyfish Clytia hemisphaer$
metazoan, Mol. Biol. Evol., 33, 2285#2293, doi: 10.1093/
ica and the evolution of the cnidarian life#cycle, Nat. Ecol.
molbev/msw100.
Evol., 3, 801#810, doi: 10.1038/s41559#019#0833#2.
162. Hassel, M., Cornelius, M. G., Vom Brocke, J., and
145. Lisenkova, A. A., Grigorenko, A. P., Tyazhelova, T. V.,
Schmeiser, H. H. (2010) Total nucleotide analysis of Hydra
Andreeva, T. V., Gusev, F. E., et al. (2017) Complete mito#
DNA and RNA by MEKC with LIF detection and 32P#
chondrial genome and evolutionary analysis of Turritopsis
postlabeling, Electrophoresis, 31, 299#302, doi: 10.1002/
dohrnii, the “immortal” jellyfish with a reversible life#
elps.200900458.
cycle, Mol. Phylogenet. Evol., 107, 232#238, doi: 10.1016/
163. Pillai, A., Gungi, A., Reddy, P. C., and Galande, S. (2021)
j.ympev.2016.11.007.
Epigenetic regulation in Hydra: Conserved and divergent
146. Shinzato, C., Khalturin, K., Inoue, J., Zayasu, Y.,
roles, Front. Cell Dev. Biol., 9, 663208, doi: 10.3389/fcell.
Kanda, M., et al. (2020) Eighteen coral genomes reveal the
2021.663208.
evolutionary origin of acropora strategies to accommodate
164. Kyger, R., Luzuriaga#Neira, A., Layman, T., Milkewitz
environmental changes, Mol. Biol. Evol., 38, 16#30,
Sandberg, T. O., Singh, D., et al. (2021) Myxosporea
doi: 10.1093/molbev/msaa216.
(Myxozoa, Cnidaria) lack DNA cytosine methylation, Mol.
147. Fuchs, B., Wang, W., Graspeuntner, S., Li, Y., Insua, S.,
Biol. Evol., 38, 393#404, doi: 10.1093/molbev/msaa214.
et al. (2014) Regulation of polyp#to#jellyfish transition in
165. Buenrostro, J. D., Giresi, P. G., Zaba, L. C., Chang, H. Y.,
Aurelia aurita, Curr. Biol., 24, 263#273, doi: 10.1016/
and Greenleaf, W. J. (2013) Transposition of native chro#
j.cub.2013.12.003.
matin for fast and sensitive epigenomic profiling of open
148. Ge, J., Liu, C., Tan, J., Bian, L., and Chen, S. (2018)
chromatin, DNA#binding proteins and nucleosome position,
Transcriptome analysis of scyphozoan jellyfish Rhopilema
Nat. Methods, 10, 1213#1218, doi: 10.1038/nmeth.2688.
esculentum from polyp to medusa identifies potential genes
166. Weizman, E. N., Tannenbaum, M., Tarrant, A. M.,
regulating strobilation, Dev. Genes Evol., 228, 243#254,
Hakim, O., and Levy, O. (2019) Chromatin dynamics
doi: 10.1007/s00427#018#0621#z.
enable transcriptional rhythms in the cnidarian
149. Nielsen, S. K. D., Koch, T. L., Hauser, F., Garm, A., and
Nematostella vectensis, PLoS Genet.,
15, e1008397,
Grimmelikhuijzen, C. J. P. (2019) De novo transcriptome
doi: 10.1371/journal.pgen.1008397.
assembly of the cubomedusa Tripedalia cystophora, includ#
167. Weizman, E., Rinsky, M., Simon#Blecher, N., Lampert#
ing the analysis of a set of genes involved in peptidergic
Karako, S., Yaron, O., et al. (2021) Chromatin dynamics
neurotransmission, BMC Genomics, 20, 175, doi: 10.1186/
and gene expression response to heat exposure in field#
s12864#019#5514#7.
conditioned versus laboratory#cultured Nematostella
150. Duncan, E. J., Gluckman, P. D., and Dearden, P. K.
vectensis, Int. J. Mol. Sci., 22, 7454, doi: 10.3390/
(2014) Epigenetics, plasticity, and evolution: How do we
ijms22147454.
link epigenetic change to phenotype?, J. Exp. Zool. B Mol.
168. Weizman, E., and Levy, O. (2019) The role of chromatin
Dev. Evol., 322, 208#220, doi: 10.1002/jez.b.22571.
dynamics under global warming response in the symbiotic
151. Moore, L. D., Le, T., and Fan, G. (2013) DNA methyla#
coral model Aiptasia, Commun. Biol., 2, 282, doi: 10.1038/
tion and its basic function, Neuropsychopharmacology, 38,
s42003#019#0543#y.
23#38, doi: 10.1038/npp.2012.112.
169. Murad, R., Macias#Muñoz, A., Wong, A., Ma, X., and
152. Henderson, I. R., and Jacobsen, S. E. (2007) Epigenetic
Mortazavi, A. (2021) Coordinated gene expression and
inheritance in plants, Nature, 447, 418#424, doi: 10.1038/
chromatin regulation during hydra head regeneration,
nature05917.
Genome Biol. Evol., 13, doi: 10.1093/gbe/evab221.
153. Sarda, S., Zeng, J., Hunt, B. G., and Yi, S. V. (2012)
170. Robertson, G., Hirst, M., Bainbridge, M., Bilenky, M.,
The evolution of invertebrate gene body methylation,
Zhao, Y., et al. (2007) Genome#wide profiles of STAT1
Mol. Biol. Evol., 29, 1907#1916, doi: 10.1093/molbev/
DNA association using chromatin immunoprecipitation
mss062.
and massively parallel sequencing, Nat. Methods, 4, 651#
154. Illumina. Methylation Sequencing, URL: https://
657, doi: 10.1038/nmeth1068.
171. Illumina. Chromatin Immunoprecipitation Sequencing
sequencing.html, Accessed 25.10.2021.
155. Frommer, M., McDonald, L. E., Millar, D. S., Collis, C.
sequencing/dna#sequencing/chip#seq.html, Accessed
M., Watt, F., et al. (1992) A genomic sequencing protocol
25.10.2021.
that yields a positive display of 5#methylcytosine residues in
172. Park, P. J. (2009) ChIP#seq: Advantages and challenges of
individual DNA strands, Proc. Natl. Acad. Sci. USA, 89,
a maturing technology, Nat. Rev. Genet., 10, 669#680,
1827#1831, doi: 10.1073/pnas.89.5.1827.
doi: 10.1038/nrg2641.
156. Masser, D. R., Hadad, N., Porter, H., Stout, M. B.,
173. Nakato, R., and Sakata, T. (2021) Methods for ChIP#seq
Unnikrishnan, A., et al. (2018) Analysis of DNA modifica#
analysis: a practical workflow and advanced applications,
tions in aging research, Geroscience,
40,
11#29,
Methods, 187, 44#53, doi: 10.1016/j.ymeth.2020.03.005.
doi: 10.1007/s11357#018#0005#3.
174. Abcam plc. Histone modifications, URL: https://
157. Li, Y., Liew, Y. J., Cui, G., Cziesielski, M. J., Zahran, N.,
et al. (2018) DNA methylation regulates transcriptional
Accessed 03.12.2021.
homeostasis of algal endosymbiosis in the coral model
175. Cell Signaling Technology, I. Histone Modification Table,
Aiptasia, Sci. Adv., 4, eaat2142, doi: 10.1126/sciadv.aat2142.
БИОХИМИЯ том 87 вып. 2 2022
НОВЫЕ ТЕХНОЛОГИИ В ИССЛЕДОВАНИЯХ СТРЕКАЮЩИХ
257
reference#tables/histone#modification#table, Accessed
Sequencing and Proteomics (Proserpio, V. ed.) Springer
03.12.2021.
New York, New York, pp. 73#85.
176. Reddy, P. C., Gungi, A., Ubhe, S., and Galande, S. (2020)
184. Hu, M., Zheng, X., Fan, C. M., and Zheng, Y. (2020)
Epigenomic landscape of enhancer elements during Hydra
Lineage dynamics of the endosymbiotic cell type in the soft
head organizer formation, Epigenet. Chromatin, 13, 43,
coral Xenia, Nature, 582, 534#538, doi: 10.1038/s41586#
doi: 10.1186/s13072#020#00364#6.
020#2385#7.
177. Schwaiger, M., Schonauer, A., Rendeiro, A. F., Pribitzer,
185. Qiu, X., Mao, Q., Tang, Y., Wang, L., Chawla, R., et al.
C., Schauer, A., et al. (2014) Evolutionary conservation of
(2017) Reversed graph embedding resolves complex single#
the eumetazoan gene regulatory landscape, Genome Res.,
cell trajectories, Nat. Methods, 14, 979#982, doi: 10.1038/
24, 639#650, doi: 10.1101/gr.162529.113.
nmeth.4402.
178. Sebe#Pedros, A., Saudemont, B., Chomsky, E., Plessier, F.,
186. La Manno, G., Soldatov, R., Zeisel, A., Braun, E.,
Mailhe, M. P., et al. (2018) Cnidarian cell type diversity
Hochgerner, H., et al. (2018) RNA velocity of single cells,
and regulation revealed by whole#organism single#cell
Nature, 560, 494#498, doi: 10.1038/s41586#018#0414#6.
RNA#Seq, Cell, 173, 1520#1534.e1520, doi: 10.1016/j.cell.
187. Chari, T., Weissbourd, B., Gehring, J., Ferraioli, A.,
2018.05.019.
Leclère, L., et al. (2021) Whole animal multiplexed single#
179. Jaitin, D. A., Kenigsberg, E., Keren#Shaul, H., Elefant,
cell RNA#Seq reveals plasticity of Clytia medusa cell types,
N., Paul, F., et al. (2014) Massively parallel single#cell
bioRxiv, doi: 10.1101/2021.01.22.427844.
RNA#seq for marker#free decomposition of tissues into cell
188. Lapebie, P., Ruggiero, A., Barreau, C., Chevalier, S.,
types, Science,
343,
776#779, doi:
10.1126/science.
Chang, P., et al. (2014) Differential responses to Wnt and
1247651.
PCP disruption predict expression and developmental
180. Keren#Shaul, H., Kenigsberg, E., Jaitin, D. A., David, E.,
function of conserved and novel genes in a cnidarian, PLoS
Paul, F., et al. (2019) MARS#seq2.0: An experimental and
Genet., 10, e1004590, doi: 10.1371/journal.pgen.1004590.
analytical pipeline for indexed sorting combined with sin#
189. Marlow, H., Roettinger, E., Boekhout, M., and
gle#cell RNA sequencing, Nat. Protoc., 14, 1841#1862,
Martindale, M. Q. (2012) Functional roles of Notch sig#
doi: 10.1038/s41596#019#0164#4.
naling in the cnidarian Nematostella vectensis, Dev. Biol.,
181. Siebert, S., Farrell, J. A., Cazet, J. F., Abeykoon, Y.,
362, 295#308, doi: 10.1016/j.ydbio.2011.11.012.
Primack, A. S., et al. (2019) Stem cell differentiation tra#
190. Matus, D. Q., Thomsen, G. H., and Martindale, M. Q.
jectories in Hydra resolved at single#cell resolution,
(2006) Dorso/ventral genes are asymmetrically expressed
Science, 365, eaav9314, doi: 10.1126/science.aav9314.
and involved in germ#layer demarcation during cnidarian
182. Macosko, E. Z., Basu, A., Satija, R., Nemesh, J.,
gastrulation, Curr. Biol., 16, 499#505, doi: 10.1016/j.cub.
Shekhar, K., et al. (2015) Highly parallel genome#wide
2006.01.052.
expression profiling of individual cells using nanoliter
191. Suryawanshi, A., Schaefer, K., Holz, O., Apel, D.,
droplets, Cell, 161, 1202#1214, doi: 10.1016/j.cell.2015.
Lange, E., et al. (2020) What lies beneath: Hydra provides
05.002.
cnidarian perspectives into the evolution of FGFR docking
183. Bageritz, J., and Raddi, G. (2019) Single$Cell RNA
proteins, Dev. Genes Evol., 230, 227#238, doi: 10.1007/
Sequencing with Drop$Seq. in Single Cell Methods:
s00427#020#00659#4.
STUDYING OF MOLECULAR REGULATION OF DEVELOPMENTAL
PROCESSES OF LOWER METAZOANS EXEMPLIFIED
BY Cnidaria USING HIGH=THROUGHPUT SEQUENCING
Review
T. V. Erofeeva1,2, A. P. Grigorenko1,2*, F. E. Gusev1,2,
I. A. Kosevich1,3, and E. I. Rogaev1,2,3,4
1 Department Research Center for Genetics and Life Sciences, Sirius University of Science and Technology,
354349 Sochi, Krasnodar Region, Russia; e$mail: anast1998@mail.ru
2 Vavilov Institute of General Genetics, Russian Academy of Sciences, 119991 Moscow, Russia
3 Faculty of Biology, Lomonosov Moscow State University, 119234 Moscow, Russia
4 Department of Psychiatry, UMass Chan Medical School, Shrewsbury, MA 01545, USA
A unique set of features and characteristics of species of the Cnidaria phylum is the one reason that makes them a
model for a various studies. The plasticity of a life cycle and the processes of cell differentiation and development of
an integral multicellular organism associated with it are of a specific scientific interest. A new stage of development of
molecular genetic methods, including methods for deep genome, transcriptome, and epigenome sequencing, both at
the level of the whole organism and at the level of individual cells, makes it possible to obtain a detailed picture of the
development of these animals. This review examines some modern approaches and advances in the reconstruction of
the processes of ontogenesis of Cnidaria by studying the regulatory signal transduction pathways and their interactions.
Keywords: Cnidaria, development, sequencing technologies, genomics, signaling pathways
БИОХИМИЯ том 87 вып. 2 2022