БИОХИМИЯ, 2022, том 87, вып. 9, с. 1318 - 1333
УДК 5.57.577
ДИСФУНКЦИЯ ГЕНОВ, АССОЦИИРУЕМЫХ
С КАНЦЕРОГЕНЕЗОМ И АПОПТОЗОМ, РАЗВИВАЮЩАЯСЯ
В ГИПОТАЛАМУСЕ САМЦОВ МЫШЕЙ ПОД ВЛИЯНИЕМ
ХРОНИЧЕСКОГО СОЦИАЛЬНОГО СТРЕССА
© 2022 А.Г. Галямина1#, Д.А. Смагин1#, И.Л. Коваленко1, О.Е. Редина1,
В.Н. Бабенко1, Н.Н. Кудрявцева1,2*
1 ФИЦ Институт цитологии и генетики СО РАН,
630090 Новосибирск, Россия; электронная почта: natnik@bionet.nsc.ru
2 Институт физиологии им. И.П. Павлова РАН, 199034 Санкт-Петербург, Россия
Поступила в редакцию 15.08.2022
После доработки 25.08.2022
Принята к публикации 25.08.2022
Хронический социальный стресс, вызванный ежедневными агонистическими взаимодействиями,
приводит к развитию смешанного тревожно/депрессивного расстройства, которое сопровождается
формированием психогенного иммунодефицита и стимуляцией онкологических процессов, а также
многими нейрогеномными изменениями в мозге самцов мышей. Целью исследования было выявить
дифференциально экспрессирующиеся гены (ДЭГ), ассоциируемые с механизмами канцерогенеза и
апоптоза в гипоталамусе депрессивных мышей и, для сравнения, у агрессивных самцов мышей с по-
зитивным социальным опытом в межсамцовых конфронтациях. Для получения животных с 20-днев-
ным альтернативным социальным опытом (побед и поражений) была использована модель хрониче-
ского социального конфликта. Результаты транскриптомного анализа (RNA-Seq) выявили сходные
изменения многих ДЭГ у агрессивных и депрессивных животных по отношению к контрольному
состоянию, однако число генов и выраженность изменений их экспрессии были существенно мень-
ше у агрессивных, чем у депрессивных самцов. Можно полагать, что однонаправленное изменение
экспрессии генов, ассоциированных с процессами онкогенеза и апоптоза, может быть следствием
длительного стрессирующего воздействия разной выраженности в гипоталамусе самцов обеих экс-
периментальных групп. Кроме того, были выделены 26 генов, не изменивших экспрессию у агрес-
сивных животных, которые могут рассматриваться как провоцирующие развитие канцерогенеза или
ингибирующие апоптоз. В результате анализа данных были выделены гены Akt1, Bag6, Foxp4, Mapk3,
Mapk8, Nol3, Pdcd10 и Xiap, экспрессия которых в наибольшей степени была скоррелирована с экс-
прессией других ДЭГ, что позволяет выдвинуть их на роль генов, кодирующих белки, обеспечиваю-
щие скоординированность нейрогеномных изменений в гипоталамусе. Дальнейшее изучение роли
этих генов может быть полезным для разработки методов фармакологической коррекции психосома-
тических патологий.
КЛЮЧЕВЫЕ СЛОВА: апоптоз, канцерогенез, RNA-Seq, ДЭГ, гипоталамус, хронический социальный
стресс.
DOI: 10.31857/S032097252209010X, EDN: BBFBTO
ВВЕДЕНИЕ
вых агонистических взаимодействиях, приво-
дит к формированию смешанного тревожно/
В результате многолетних исследований
депрессивного расстройства у мышей [1-3],
было показано, что хронический социальный
которое сопровождается развитием иммуно-
стресс, вызванный повторным негативным
супрессии, проявляющейся в снижении об-
опытом поражений в ежедневных межсамцо- щей резистентности и многих показателей гу-
морального и клеточного иммунитета [4-11],
что позволяло говорить о развитии психоген-
Принятые сокращения: ДЭГ
- дифференциально
ного иммунодефицита [10] в этих условиях.
экспрессирующиеся гены.
На фоне сниженного иммунитета у депрессив-
* Адресат для корреспонденции.
ных животных наблюдалось усиление мета-
# Авторы внесли равный вклад в работу.
стазирования прививаемых опухолевых клеток
1318
ДИСФУНКЦИЯ ГЕНОВ ПРИ ХРОНИЧЕСКОМ СОЦИАЛЬНОМ СТРЕССЕ
1319
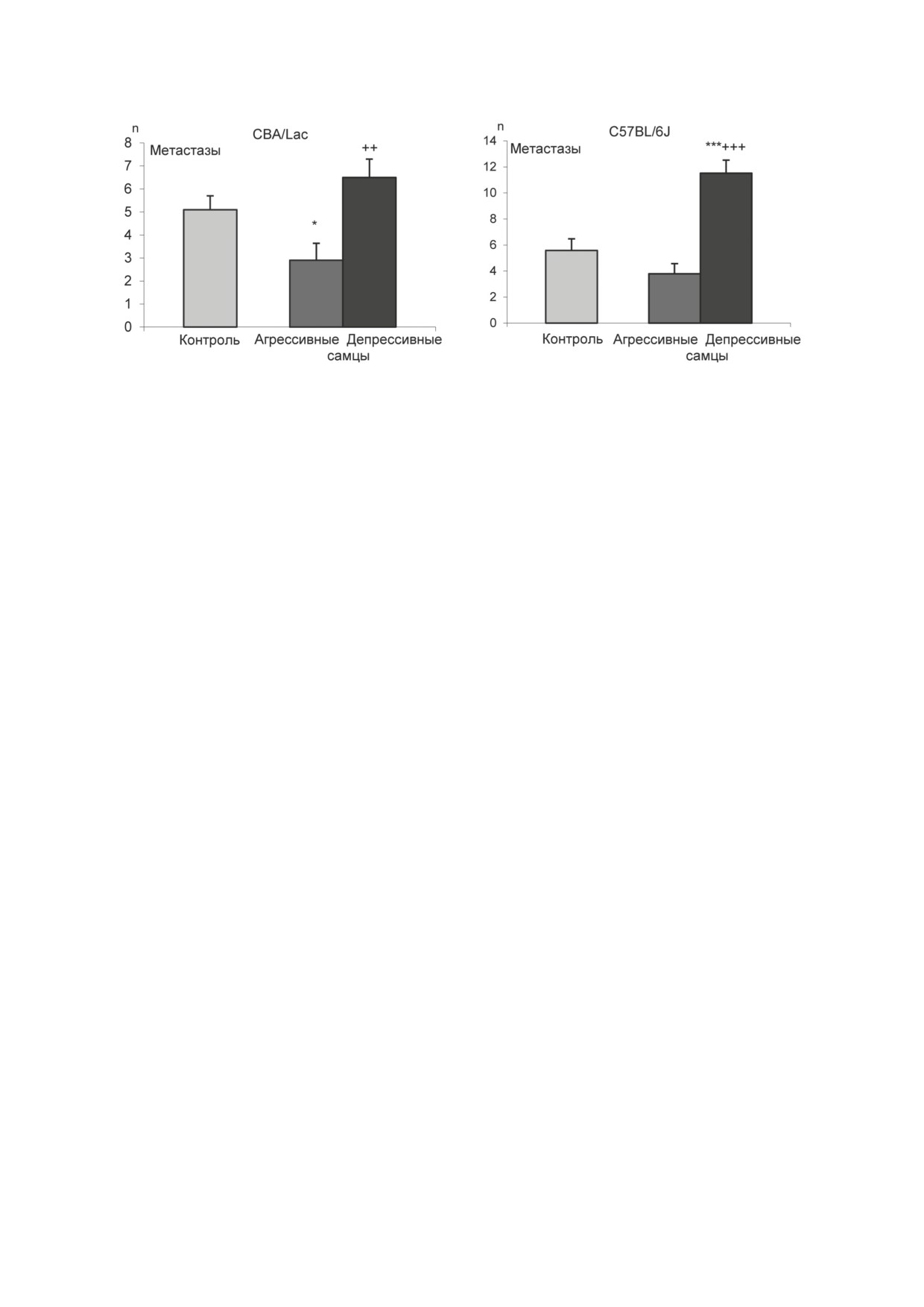
Рис. 1. Метастазы (n) в лёгких у агрессивных и депрессивных самцов мышей. Опухолевые клетки гепатокарциномы-29
вводили в хвостовую вену мышей линии СВА/Lac (контрольные, агрессивные и депрессивные самцы). Аналогичным
образом вводили соответствующим группам мышей линии С57BL/6 опухолевые клетки карциномы лёгких Льюиса.
На 16-й день после инъекции опухолевых клеток подсчитывали число метастазов в лёгких. * p < 0,05; *** p < 0,001 по
сравнению с контролем; ++ p < 0,01; +++ p < 0,001 по сравнению с агрессивными мышами [14]
по сравнению с контрольными животными
положение, что различный уровень метаста-
и агрессивными самцами с повторным пози-
зирования у животных с альтернативным со-
тивным социальным опытом [12-14] (рис. 1).
циальным опытом может быть как следствием
Различная интенсивность метастазирования
различий в периферической регуляции иммун-
первичных опухолей Кребс-2, карциномы
ных процессов, так и возникать на геномном
лёгких Льюиса (Levis lung carcinoma, LLC) и
уровне в мозге.
гепатокарциномы-29 (Н-29) у животных экс-
В качестве отдела мозга для изучения взаи-
периментальных групп свидетельствовала о
мообусловленных патофизиологических взаи-
выраженном и хорошо воспроизводимом вли-
мосвязей хронического социального стресса,
янии социального опыта у мышей.
канцерогенеза и апоптоза интерес представляет
В контексте взаимосвязи депрессивной
гипоталамус, вовлечённый как в нервную, так и
симптоматики с процессами снижения имму-
в гуморальную регуляцию поведения, а также в
нитета и, как следствие, повышенного онкоге-
иммунные процессы [29]. Предполагается, что
неза было актуальным рассмотреть взаимосвязь
участие гипоталамуса в механизмах развития де-
процессов апоптоза и канцерогенеза, общие
прессии может осуществляться на разных уров-
механизмы которых были показаны много-
нях [30]. Дискуссия о роли нейровоспаления в
кратно у людей и лабораторных животных [15,
этом отделе мозга при депрессии идёт уже в тече-
16]. Так, известно, что процессы роста опухо-
ние многих лет, и современные данные подтвер-
ли, как правило, сопровождаются интенсив-
ждают их взаимосвязь [29]. Кроме того, известно,
ной клеточной пролиферацией в сочетании со
что нарушения в функционировании гипота-
снижением интенсивности апоптоза, что в со-
ламо-гипофизарно-надпочечниковой системы
вокупности приводит к нарушению тканевого
наблюдаются при онкологических заболевани-
гомеостаза [17]. При депрессии существует по-
ях [31], сопровождающихся изменением процес-
вышенная вероятность развития опухоли [18,
сов апоптоза нейронов в гипоталамусе [32]. По-
19], о чем свидетельствуют также и наши дан-
казано также, что полиморфизм по некоторым
ные, полученные на депрессивных животных в
гипоталамическим генам связан с повышенным
сравнении с контрольным состоянием. Показа-
риском депрессии [33]. Очевидно, что гипотала-
но, что хронический социальный стресс может
мус можно рассматривать в качестве отдела моз-
стимулировать процессы апоптоза, в частности
га, в котором происходит пересечение патофизио-
в головном мозге (например, вызывать гибель
логических механизмов психоэмоциональных
нейронов в гиппокампе и миндалине) [20, 21].
расстройств и связанных с ними иммунных нару-
Тем важнее исследовать молекулярные меха-
шений, и онкологическиx заболеваний.
низмы, связывающие хронический социаль-
Целью данной работы было выявление из-
ный стресс, апоптоз и канцерогенез.
менений в профиле экспрессии генов, ассоци-
Поскольку многократно было показано
ированных с канцерогенезом и апоптозом, в
нами, что хронический социальный стресс вы-
гипоталамусе депрессивных самцов мышей со
зывает специфические изменения экспрессии
сниженным под влиянием хронического со-
генов в мозге [22-28], было высказано пред-
циального стресса иммунным статусом [10] в
БИОХИМИЯ том 87 вып. 9 2022
1
320
ГАЛЯМИНА и др.
сравнительном аспекте с агрессивными сам-
му за перегородкой. Взаимодействие самцов
цами с позитивным опытом в агонистических
прекращали, если интенсивные атаки со сто-
взаимодействиях и с интактным контролем.
роны нападающей особи во время агрессивных
Выявление геномных механизмов, лежащих в
столкновений длились не более трёх минут и
основе коморбидности психоэмоциональных
меньше, устанавливая между ними перегородку
расстройств, онкологических заболеваний и
и предотвращая физическое повреждение сам-
иммунного статуса, может помочь найти новые
цов, терпящих поражение.
эффективные фармакологические мишени для
Для нейрогеномных исследований отби-
более успешной терапии этих заболеваний. Ис-
рали животных с наиболее выраженным фе-
следование проводилось в рамках развиваемого
нотипом поведения, сформированным в еже-
коллективом направления
«Функциональная
дневных агонистических взаимодействиях.
нейрогеномика патологических состояний».
В эксперименте были исследованы три группы
животных: 1) депрессивные самцы, потерпев-
шие социальные поражения в течение 20 дней,
МАТЕРИАЛЫ И МЕТОДЫ
у которых под влиянием хронического социаль-
ного стресса развивается смешанное тревожно/
Животные. Эксперименты проводили
депрессивное расстройство с признаками вы-
на самцах мышей линии С57BL/6 в возрасте
раженного депрессивноподобного состояния,
2,5 мес. с массой тела 26-28 г. Животные были
а также тревоги и страха, оцениваемое мно-
привезены из Питомника лабораторных живот-
гими поведенческими тестами [1-3]; 2) агрес-
ных Института биоорганической химии РАН
сивные самцы с повторным опытом агрессии
(Пущино, Московская область). Эксперимент
в течение 20 дней, которые демонстрировали
был проведён в конвенциональном виварии
ежедневно гиперактивность и повышенную
Института цитологии и генетики СО РАН (Но-
агрессивность, не поддающуюся коррекции си-
восибирск, Россия). Животные имели посто-
туационными факторами [36]; 3) контрольные
янный неограниченный доступ к корму (гра-
животные - особи без последовательного опы-
нулам) и воде и содержались при 12-часовом
та агонистических взаимодействий. Животные
цикле свет/темнота.
экспериментальных групп были декапити-
Формирование патологий поведения у сам-
рованы на следующий день после последней
цов мышей. Хронический социальный стресс у
конфронтации одновременно с контрольными
мышей формировали с применением модели
особями. Гипоталамус извлекали в соответ-
сенсорного контакта [34], переименованной в
ствии с анатомическим атласом мозга (Allen
последующем в модель хронического социаль-
ного конфликта [35]. Методика подробно опи-
static/atlas). Все образцы помещали в раствор
сана в статьях и обзоре Kudryavtseva et al. [36]. В
RNAlater («Life Technologies», США) и хранили
ежедневных агонистических взаимодействиях
при температуре -70 °С до секвенирования.
животные приобретали альтернативный опыт
RNA-Seq-анализ. Образцы гипоталамуса
социального поведения - позитивный опыт
самцов мышей были отправлены в ЦКП «Гено-
побед в агонистических взаимодействиях и
аналитика» для секвенирования транскрипто-
хронический негативный опыт социальных по-
ражений c агрессивным партнёром.
В каждой группе было проанализировано по
Для этого животных попарно помещали в
3 животных. Все образцы секвенировали от-
экспериментальные клетки (28 × 14 × 10 см),
дельно. При анализе использовали более 20 млн
разделённые пополам прозрачной перегород-
прочтений ДНК. мРНК экстрагировали с ис-
кой с отверстиями, позволявшей мышам ви-
пользованием Dynabeads RNA Purificatio nKit
деть, слышать, воспринимать запахи друг дру-
(«Ambion», США). Секвенирование библиотек
га (сенсорный контакт), но предотвращавшей
кДНК было выполнено на платформе Illumina
физическое взаимодействие. Ежедневно во
Hiseq 1500 («Illumina Sequencing», США). Про-
второй половине дня (14:00-17:00 ч) убирали
грамма Cufflinks была использована для оцен-
перегородку на 10 минут, что приводило к меж-
ки уровня экспрессии генов в единицах FPKM
самцовым конфронтациям. Во время первых
(fragments per kilobase of transcript per million
2-3 дней в поведенческих тестах выявляли по-
mapped reads). Более 30 000 генов были про-
бедителей и особей, терпящих поражения при
анализированы. Только аннотированные гены
взаимодействии с одним и тем же партнёром.
были взяты для последующего анализа.
В дальнейшем ежедневно после теста побеж-
Дифференциально экспрессирующими-
дённого самца пересаживали в новую клетку к
ся считали гены, уровень экспрессии которых
незнакомому агрессивному партнёру, сидяще-
статистически значимо различался у контроль-
БИОХИМИЯ том 87 вып. 9 2022
ДИСФУНКЦИЯ ГЕНОВ ПРИ ХРОНИЧЕСКОМ СОЦИАЛЬНОМ СТРЕССЕ
1321
ных и подопытных мышей (p < 0,05). Решение о
поталамусе депрессивных и агрессивных мы-
статистически значимых различиях экспрессии
шей с использованием базы данных (http://
принималось также с использованием поправ-
ки на множественные сравнения (q-значения -
apocand/browse.php). Были отобраны 230 генов
скорректированные p-значения по методу
для анализа. У агрессивных самцов (табл. 1;
Беньямини-Хохберга (FDR)). Для биоинфор-
табл. 1 в Приложении) под влиянием позитив-
матической обработки исследуемого массива
ного социального опыта в агонистических взаи-
транскриптомных данных был использован
модействиях у 40 генов изменялась экспрессия
также корреляционный анализ.
по отношению к контролю, из них увеличение
Была проанализирована экспрессия генов,
экспрессии отмечали у 27 генов (68,5%), сни-
ассоциируемых с механизмами апоптоза и кан-
жение - у 13 генов (32,5%). У депрессивных
церогенеза. Гены для анализа были отобраны c
животных под влиянием хронического соци-
использованием баз данных, связанных с про-
ального стресса экспрессия была изменена
у 62 генов, из них у 38 генов (61%) она была
повышена по отношению к контролю, у 24 ге-
php), а также по литературным данным (табл. 1;
нов (39%) - снижена.
табл. 1 в Приложении). Для исследования были
Общими для обеих экспериментальных
взяты только гены, экспрессия которых была
групп являются 36 ДЭГ (табл. 1; табл. 1 в При-
изменена в гипоталамусе у агрессивных и/или
ложении) с однонаправленным изменением
депрессивных животных по отношению к кон-
показателей FPKM. Из них у 26 генов экспрес-
трольному состоянию.
сия была увеличена, у 10 - снижена. Исключе-
Категории генных онтологий списков диф-
нием были гены Casp2, Foxp2, Opa1 и Tnfrsf22,
ференциально экспрессирующихся генов (ДЭГ)
уровень экспрессии которых отличается от
определяли с использованием биоинформати-
контрольного только у агрессивных животных.
ческого интернет-ресурса DAVID Bioinformatics
Экспрессия гена Casp2 возрастает, в то время
как генов Foxp2, Opa1 и Tnfrsf22 - снижается.
лиз функций генов, а также биологических про-
Проведён анализ по выявлению ДЭГ, из-
цессов, ассоциируемых с этими генами, прово-
менивших экспрессию только у депрессив-
дили при помощи базы данных KEGG pathway
ных самцов мышей. Для этого были выделены
34 ДЭГ с изменённой экспрессией при стати-
стическом уровне значимости q < 0,05 у депрес-
деления кластеров ДЭГ, позволяющих оценить
сивных животных и р < 0,05 - у агрессивных,
взаимосвязь нейрогеномных процессов у экспе-
отличающиеся по показателям FPKM от кон-
риментальных групп по отношению к контро-
трольных животных (табл. 2 в Приложении).
лю, была применена программа XLStat (https://
Оказалось, что только у 13 генов специфически
была изменена экспрессия у депрессивных сам-
Поиск аналогов генов, ассоциируемых с кан-
цов: из них у четырёх генов экспрессия повы-
церогенезом и апоптозом, вовлечённых в разви-
шалась (Akt2, Сlnd5, Nos3 и Siva1), а у 9 - сни-
тие карциномы лёгких у людей (по литературным
жалась (Aifm1, Anxa1, Casp8, Fastkd2, Mcl1, Nol4,
данным). С целью поиска возможных перифе-
Pycard, Sfrp1 и Stk17b). При этом большинство
рических маркеров изменённой экспрессии
генов со сниженной экспрессией вовлечены в
генов канцерогенеза и апоптоза в гипоталаму-
механизмы апоптоза. Таким образом, на фоне
се у агрессивных и депрессивных мышей был
снижения экспрессии генов, кодирующих бел-
проведён сравнительный анализ состояния
ки, ассоциируемые с апоптозом, может проис-
генов у больных раком лёгких с использовани-
ходить усиление канцерогенеза, что и было по-
казано у депрессивных животных.
card/lung_cancer?search=lung%20cancer%20genes),
В то же время было выделено 26 генов, экс-
а также генов, изменивших экспрессию в лёгких
прессия которых была изменена у депрессив-
ных, но оставалась неизменной у агрессивных
org/search/results?query=lung+cancer+metastasis+
самцов по отношению к контролю: Aifm1, Ak2,
gene).
Akt2, Anxa1, Bcl2l13, Bnip3, Casp8, Ccar1, Сldn5,
Cldn12, Cyld, Dpf2, Fastkd2, Mcl1, Nol3, Nol4,
Nomo1, Nos1, Nos3, Pdcd5, Ppargc1b, Pycard, Sfrp1,
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Siva1, Stk17b и Tnfrsf8 (табл. 1). Из них у 12 ге-
нов экспрессия повышалась (Ak2, Akt2, Bcl2l13,
Были исследованы ДЭГ, ассоциируемые с
Сldn5, Dpf2, Nol3, Nomo1, Nos1, Nos3, Ppargc1b,
механизмами апоптоза и канцерогенеза в ги-
Siva1 и Tnfrsf8), а у 14 - снижалась (Aifm1, Anxa1,
11
БИОХИМИЯ том 87 вып. 9 2022
1
322
ГАЛЯМИНА и др.
Bnip3, Casp8, Ccar1, Cldn12, Cyld, Fastkd2, Mcl1, большинство генов вовлечены в механизмы
Nol4, Pdcd5, Pycard, Sfrp1 и Stk17b). При этом апоптоза (табл. 1 в Приложении).
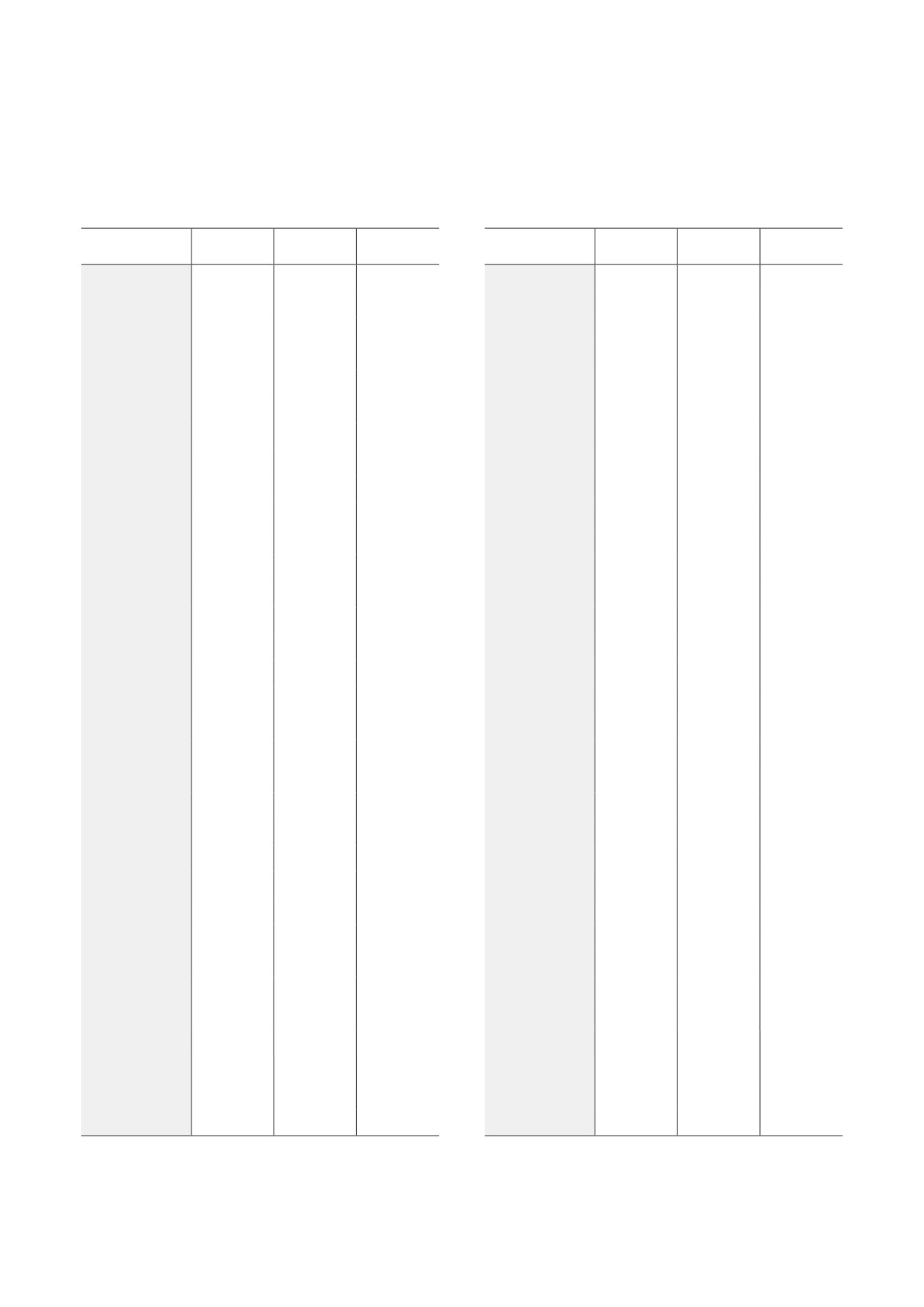
Таблица 1. ДЭГ, ассоциируемые с канцерогенезом и апоптозом в гипоталамусе самцов мышей с альтернативным
социальным опытом
Ген
К vs А
К vs Д
А vs Д
Ген
К vs А
К vs Д
А vs Д
Aifm1
=
↓↓*
Hrk
↑
↑
Ak2
=
↑
Mapk1
↓↓
↓↓*
Akt1
↑
↑↑↑*
Mapk3
↑↑
↑↑↑*
↑
Akt2
=
↑↑*
Mapk8
↓
↓↓*
Alkbh7
↑
↑↑↑*
↑↑
Mcl1
=
↓↓*
Anxa1
=
↓↓*
↓↓↓
Nfkbib
↑↑
↑↑↑*
Arc
↑↑
↑↑↑*
↑↑↑*
Nol3
=
↑
Avp
↑↑↑*
↑↑↑*
Nol4
=
↓↓*
Avpi1
↑
↑
Nol8
↓
↓↓↓*
Bag3
↑↑↑*
↑↑↑*
Nomo1
=
↑
Bag6
↑↑
↑↑↑*
Nos1
=
↑
Bax
↑↑
↑↑↑*
↑
Nos3
=
↑↑*
Bbc3
↑
↑
Nosip
↑
↑↑↑*
↑
Bcl2l1
↑↑
↑↑↑*
Opa1
↓
=
Bcl2l13
=
↑
Pdcd5
=
↓
Bnip3
=
↓
Pdcd10
↓↓*
↓↓↓*
Bok
↑↑*
↑↑↑*
Pomc
↑↑↑*
↑↑↑*
Casp2
↑
=
↓
Ppargc1b
=
↑
Casp8
=
↓↓*
↓↓
Pycard
=
↓↓*
↓↓
Casp8ap2
↓↓↓*
↓↓↓*
Sfrp1
=
↓↓↓*
Ccar1
=
↓
Sfrp5
↑↑↑*
↑↑↑*
Сldn5
=
↑↑*
Sh3glb1
↓
↓
Cldn12
=
↓
Siva1
=
↑↑*
Clip3
↑
↑↑*
Stk3
↓↓
↓↓*
Cyld
=
↓
Stk17b
=
↓↓*
↓
Dpf1
↑↑
↑↑↑*
Stk19
↑
↑↑↑*
Dpf2
=
↑
Stk39
↓↓*
↓↓*
Fasn
↑↑↑*
↑↑↑*
Tnfaip8
↓
↓
Fastk
↑
↑↑↑*
Tnfrsf1a
↑↑
↑
Fastkd2
=
↓↓*
Tnfrsf8
=
↑
Foxp2
↓
=
Tnfrsf22
↓
=
Foxp4
↑↑↑*
↑↑↑*
Tnfrsf25
↑↑↑*
↑↑↑*
Gapdh
↑↑
↑↑↑*
Xiap
↓↓↓*
↓↓↓*
Примечание. К - контроль; А - агрессивные мыши; Д - депрессивные мыши. ↓ - Снижение экспрессии; ↑ - увеличе-
ние экспрессии; = - экспрессия не изменена; ↓ p < 0,05; ↓↓ p < 0,01; ↓↓↓ p < 0,001; ↑ p < 0,05; ↑↑ p < 0,01; ↑↑↑ p < 0,001;
* q < 0,05.
БИОХИМИЯ том 87 вып. 9 2022
ДИСФУНКЦИЯ ГЕНОВ ПРИ ХРОНИЧЕСКОМ СОЦИАЛЬНОМ СТРЕССЕ
1323
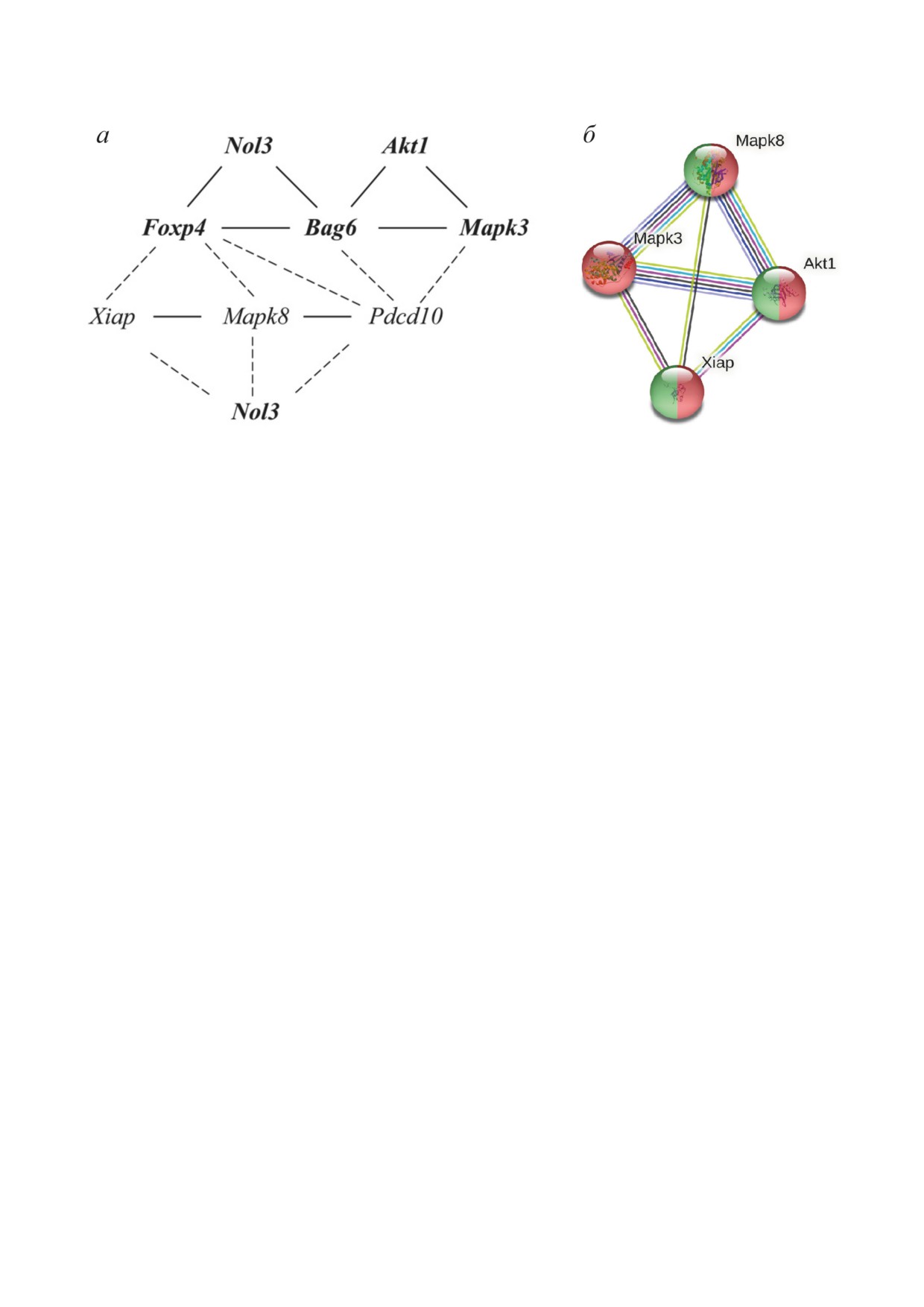
Рис. 2. Корреляционные взаимосвязи функциональной активности ДЭГ. а - Пунктирная линия - отрицательная взаи-
мосвязь; сплошная линия - положительная взаимосвязь; обычным шрифтом обозначено снижение экспрессии у кон-
кретного гена; жирным шрифтом - повышение экспрессии. Ген Nol3 представлен на рисунке дважды для удобства
представления. б - Наибольшее число взаимосвязей, согласно базе данных String (string-db.org), было установлено для
генов Akt1, Mapk3, Mapk8 и Xiap
Также было проведено сравнение выра-
группы коррелируют между собой положи-
женности экспрессии генов у агрессивных и
тельно. Между уровнями экспрессии генов,
депрессивных мышей (табл. 1 в Приложении)
относящихся к разным кластерам, наблюдает-
по отношению друг к другу. Различия по это-
ся отрицательная корреляция. Показаны от-
му показателю были обнаружены у 10 ДЭГ.
рицательные корреляции с генами Mapk8, Xiap
По сравнению с агрессивными животными,
и Pdcd10 и положительные - с Foxp4 и Bag6.
у депрессивных самцов гены Arc, Bax и Nosip,
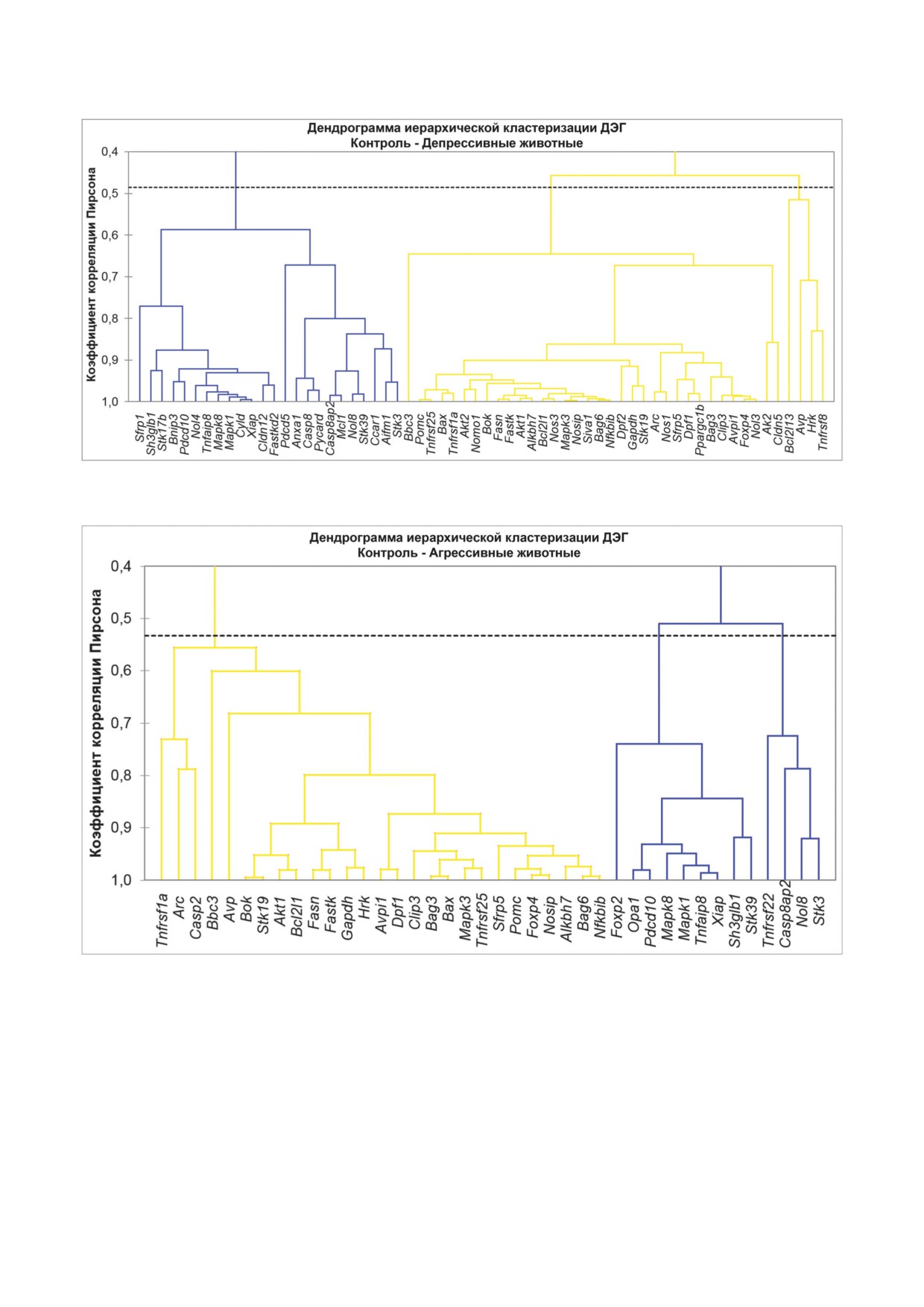
Агломеративная иерархическая кластериза-
кодирующие регуляторные молекулы, и гены
ция ДЭГ. Был проведён анализ по визуализации
Alkbh7 и Mapk3, кодирующие белки-ферменты,
иерархической кластеризации ДЭГ в группах
демонстрируют повышенный уровень экспрес-
агрессивных и депрессивных мышей в срав-
сии; а гены Casp2, Casp8 и Stk17b, кодирующие
нительном аспекте с контрольной группой.
белки-ферменты, а также Anxa1, кодирующий
В результате анализа данных по депрессивным
регуляторный белок, и Pycard, кодирующий
животным были выделены 3 кластера (рис. 3;
транскрипционный фактор, - пониженный.
табл. 4 в Приложении), включающие кластер из
Корреляционный анализ. В табл. 3 в Прило-
24 ДЭГ со сниженной экспрессией и 2 класте-
жении приведено общее число корреляцион-
ра генов, экспрессия которых повышалась (34
ных зависимостей между ДЭГ у трёх групп -
и 4 гена). В кластер генов со сниженной экс-
контроль, агрессивные и депрессивные самцы.
прессией попадают Pdcd10, Mapk8 и Xiap, кото-
Для дальнейшего рассмотрения были отобра-
рые могут рассматриваться как координирую-
ны гены Akt1, Bag6, Foxp4, Mapk3, Mapk8, Nol3,
щие взаимодействие других генов. В кластер с
Pdcd10 и Xiap, для которых было показано мак-
генами, у которых экспрессия повышалась, по-
симальное количество корреляций с другими
падают Akt1, Bag6, Foxp4, Mapk3 и Nol3, и, что
генами (при r ≥ 0,95 и r ≤ -0,95). Эти гены в
характерно, все они относятся к одному класте-
дальнейшем анализе рассматривались в каче-
ру из двух (большему).
стве возможных кандидатов на роль координи-
У агрессивных животных были также выде-
рующих генов, участвующих в формировании
лены 3 кластера (рис. 4; табл. 4 в Приложении),
психосоматических патологий под влиянием
включающие кластер из 27 генов с повышенной
хронического социального конфликта. В табл. 2
экспрессией и 2 кластера, экспрессия у которых
представлены ДЭГ, имеющие хотя бы одну кор-
снижалась - 4 и 9 ДЭГ. Что касается «коорди-
реляцию с одним из высоко коррелирующих
нирующих» генов, то здесь наблюдается то же,
генов. Кроме того, на рис. 2 можно видеть, что
что и у депрессивных животных: Pdcd10, Mapk8
выделяются 2 группы координирующих генов:
и Xiap попадают в один кластер снизивших
Akt1, Bag6, Foxp4, Mapk3 и Nol3, экспрессия ко-
экспрессию ДЭГ. В кластер с увеличившими
торых была увеличена; и Mapk8, Pdcd10, Xiap,
экспрессию генами попадают Akt1, Bag6, Foxp4
экспрессия которых снижалась. Гены каждой
и Mapk3.
БИОХИМИЯ том 87 вып. 9 2022
11*
1
324
ГАЛЯМИНА и др.
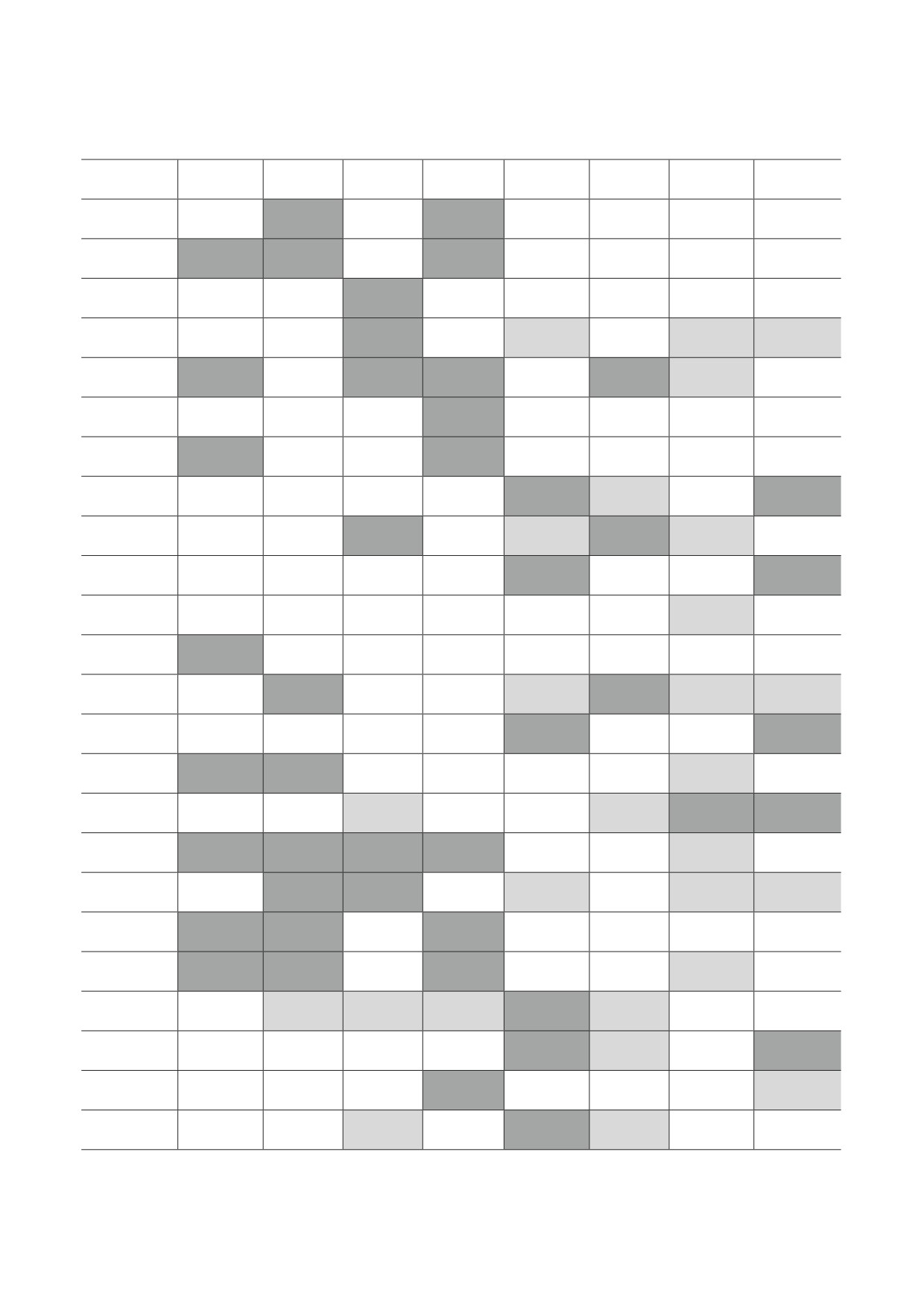
Таблица 2. Коэффициенты корреляций между экспрессией генов, кодирующие белки, вовлечённые в координацию ме-
ханизмов канцерогенеза и апоптоза
Akt1
Bag6
Foxp4
Mapk3
Mapk8
Nol3
Pdcd10
Xiap
Akt1
1,000
0,958
0,896
0,975
-0,844
0,864
-0,944
-0,864
Alkbh7
0,982
0,952
0,883
0,956
-0,838
0,870
-0,915
-0,853
Avpi1
0,844
0,876
0,954
0,849
-0,871
0,888
-0,908
-0,865
Bag3
0,872
0,930
0,957
0,935
-0,963
0,926
-0,970
-0,950
Bag6
0,958
1,000
0,961
0,983
-0,930
0,954
-0,988
-0,918
Bax
0,919
0,928
0,874
0,958
-0,908
0,864
-0,931
-0,905
Bcl2l1
0,966
0,940
0,864
0,952
-0,814
0,825
-0,942
-0,839
Cldn12
-0,803
-0,886
-0,944
-0,896
0,985
-0,952
0,913
0,955
Clip3
0,842
0,925
0,985
0,916
-0,968
0,982
-0,956
-0,945
Cyld
-0,850
-0,901
-0,930
-0,932
0,979
-0,945
0,920
0,981
Dpf1
0,884
0,928
0,929
0,893
-0,857
0,868
-0,950
-0,818
Fasn
0,979
0,937
0,835
0,949
-0,786
0,819
-0,907
-0,824
Foxp4
0,896
0,961
1,000
0,942
-0,966
0,982
-0,977
-0,953
Mapk1
-0,858
-0,901
-0,935
-0,930
0,960
-0,923
0,940
0,980
Mapk3
0,975
0,983
0,942
1,000
-0,930
0,934
-0,981
-0,930
Mapk8
-0,844
-0,930
-0,966
-0,930
1,000
-0,971
0,950
0,973
Nfkbib
0,952
0,989
0,959
0,967
-0,921
0,938
-0,980
-0,930
Nol3
0,864
0,954
0,982
0,934
-0,971
1,000
-0,957
-0,955
Nomo1
0,967
0,959
0,916
0,977
-0,891
0,917
-0,943
-0,883
Nosip
0,977
0,982
0,945
0,990
-0,922
0,936
-0,969
-0,918
Pdcd10
-0,944
-0,988
-0,977
-0,981
0,950
-0,957
1,000
0,936
Tnfaip8
-0,856
-0,913
-0,940
-0,928
0,959
-0,960
0,806
0,991
Tnfrsf25
0,898
0,937
0,925
0,953
-0,947
0,941
0,248
-0,983
Xiap
-0,864
-0,918
-0,953
-0,930
0,973
-0,955
0,936
1,000
Примечание. Жирным шрифтом выделены корреляции с уровнем статистической значимости p ≤ 0,001. Светло-серым
выделены отрицательные корреляции при r ≤ -0,950, темно-серым - положительные при r ≥ 0,950.
БИОХИМИЯ том 87 вып. 9 2022
ДИСФУНКЦИЯ ГЕНОВ ПРИ ХРОНИЧЕСКОМ СОЦИАЛЬНОМ СТРЕССЕ
1325
Рис. 3. Дендрограмма иерархической кластеризации ДЭГ в гипоталамусе депрессивных самцов. Темно-серым цветом
выделены гены со сниженной экспрессией, светло-серым цветом - гены с повышенной экспрессией
Рис. 4. Дендрограмма иерархической кластеризации ДЭГ в гипоталамусе агрессивных самцов. Темно-серым цветом
выделены гены со сниженной экспрессией, светло-серым цветом - гены с повышенной экспрессией
Согласно связям, представленным на ден-
вотных; ген Mapk3 - с геном Tnfrsf25 (у агрес-
дрограммах, среди ДЭГ, предполагаемых в ка-
сивных самцов) и с геном Nosip (у депрессивных
честве координирующих, существует несколько
животных); ген Pdcd10 - с Opa1 (у агрессивных
возможных взаимоотношений. Так, например,
самцов) и с геном Bnip3 (у депрессивных сам-
ген Akt1 кластеризуется с геном Bcl2l1 у агрес-
цов). В то же время ген Bag6 тесно взаимосвя-
сивных и депрессивных животных и с ге-
зан с геном Nfkbib у обеих групп животных; а
ном Alkbh7 - у депрессивных мышей; Foxp4
ген Mapk8 - с генами Mapk1, Tnfaip8, и Xiap (у
кластеризуется с геном Nosip у агрессивных
агрессивных самцов) и с генами Mapk1, Сyld
самцов и с геном Avpi1 - у депрессивных жи-
и Xiap - у депрессивных самцов.
БИОХИМИЯ том 87 вып. 9 2022
1
326
ГАЛЯМИНА и др.
Предварительный вывод из сопоставлений
данных (MGI Mouse Phenotypes related to Lung
кластеризации у двух групп экспериментальных
Cancer), относятся около 216 генов, среди ко-
животных: экспрессия генов может изменяться
торых были отобраны для анализа топ-гены с
в зависимости от нейрогеномного окружения,
неоднократно подтверждёнными результатами:
которое у агрессивных и депрессивных самцов
Akt1, Alk, Braf, Casp8, Cyp2a6, Egfr, Erbb2, Ercc6,
по многим показателям различается.
Faslg, Kras, Irf1, Pik3ca, Prkn и Slc22a18 для срав-
ДЭГ, вовлечённые в метаболические пути,
нительного анализа с ДЭГ, обнаруженными в
согласно базе данных KEGG. С использовани-
гипоталамусе мыши.
Среди ДЭГ в гипоталамусе депрессив-
kegg/pathway.html) были выделены метаболи-
ных животных были найдены сходные гены
ческие пути, в работе которых принимают уча-
(табл. 6 в Приложении) семейств Akt (Akt1,
стие ДЭГ, выявленные в нашем исследовании:
Akt2, Akt1s1), Alkb (Alkbh7), Casp (Casp2, Casp8,
2 гена кодируют сигнальные молекулы, 5 генов
Casp8ap2), Irf2bp (Irf2bpl, Irf2bp1, Irf5, Irf9),
являются транскрипционными факторами,
Ercc (Ercc1, Ercc2, Ercc4), а также гены Braf,
22 ДЭГ кодируют белки-ферменты. Самой мно-
Pik3ca и Erbb2i. У агрессивных животных была
гочисленной является группа генов (29 ДЭГ),
обнаружена аналогичная тенденция, однако
кодирующих регуляторные белки, вовлечён-
количество генов было меньшим: Akt1, Akt1s1,
ные в различные внутриклеточные процессы
Aqp4, Braf, Casp8ap2, Erbb2ip, Irf1, Irf2bpl, Irf2bp1
(табл. 5 в Приложении).
и Kras.
Все метаболические пути связаны с меха-
Кроме того, по сравнению с тканью здо-
низмами канцерогенеза и апоптоза либо с за-
ровых лёгких у пациентов с диагнозом рак
болеваниями, в патогенез которых входят эти
лёгких (Lung cancer) в лёгочной ткани были
процессы как в периферических органах, так
обнаружены в числе топ 10 ДЭГ - Adh1b, Ager,
и в головном мозге. В частности, пути, связан-
Aqp4, Cldn18, Clic5, Cyp4b1, Krt6a, Mmp12, Sftpc
ные с развитием паразитарных и инфекцион-
и Sprr1b. Дифференциальная экспрессия сход-
ных заболеваний, таких как токсоплазмоз, ле-
ных генов была обнаружена и у депрессивных
гионеллёз, туберкулёз, при которых отмечается
животных в гипоталамусе - Ager, Aqp4, Cyp4а14,
повышенная клеточная гибель в поражённых
семейств Cldn (Cldn5, Cldn12), Clic (Clic1, Clic5)
органах. Также обнаружены метаболические
и Mmp (Mmp15, Mmp17). У агрессивных жи-
пути заболеваний, чья основная патология вы-
вотных это были гены семейств Clic (Clic5,
зывается именно апоптозными процессами, как
Clic6), Cyp (Cyp4а15, Cyp46а1) и Mmp (Mmp16,
в случае амиотрофического латерального скле-
Mmp17), а также Adh5, Aqp4, Cldn9, Krtcap2 и
роза, неалкогольных повреждений печени или
Krt9 (табл. 6 в Приложении). Все эти данные
болезни Альцгеймера. Исследуемые гены во-
могут свидетельствовать о том, что в мозге, в
влечены также в функционирование сигнальных
частности в гипоталамусе, изменения экспрес-
путей, регулирующих апоптоз - p53, NF-kappa
сии генов под влиянием хронического социаль-
B, сфинголипидный сигнальный путь. Участие
ного стресса могут в той или иной степени со-
ДЭГ в процессах, связанных не только с апопто-
ответствовать изменениям в периферических
зом, наиболее часто выявляется для генов Aifm1,
тканях. Однако этот вывод нуждается в экспе-
Akt1, Akt2, Bax, Bcl2l1, Casp2, Casp8, Cyld, Clnd5,
риментальном подтверждении.
Hrk, Mapk1, Mapk3, Mapk8, Nos3, Nfkb1b, Pycard,
Pomc, Tnfrsf1a, Xiap и др. (табл. 5 в Приложении).
Наибольший уровень экспрессии (> 40-50
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
FPKM) показан для генов Akt (50,2), Alkph7
(47,4), Avp (439,9), Bag6 (53,9), Clip3 (153,5),
Ранее было показано, что хронический
Gapdh (320,6), Mapk1 (57,95), Mapk3 (69,8), Pdcd5
стресс различной этиологии может вызывать
(68,6), Pomc (61,6) (табл. 1 в Приложении).
апоптоз в культурах клеток гипоталамуса [37,
Сравнительный анализ генов, ассоциирован-
38]. Полученные нами данные также позволя-
ных с канцерогенезом и апоптозом в гипоталаму-
ют предположить, что хронический социаль-
се депрессивных животных, и состояния генов у
ный стресс может быть причиной изменения
больных с лёгочной карциномой (по литератур-
экспрессии генов, ассоциируемых с апоптозом
ным данным). С использованием баз данных по
и канцерогенезом в гипоталамусе у животных,
вступающих в конфликтные взаимодействия.
но изменённое состояние генов в разных тка-
При этом у самцов с негативным социальным
нях, а также непосредственно в лёгких у боль-
опытом, сопровождающимся развитием де-
ных с лёгочной карциномой. У людей к генам,
прессивноподобного состояния [1], показаны
ассоциируемых с раком лёгких, согласно базе
изменения экспрессии большего числа генов,
БИОХИМИЯ том 87 вып. 9 2022
ДИСФУНКЦИЯ ГЕНОВ ПРИ ХРОНИЧЕСКОМ СОЦИАЛЬНОМ СТРЕССЕ
1327
чем у агрессивных самцов с позитивным со-
экспрессии генов канцерогенеза и апопотоза
циальным опытом. Характерно, что подобная
при развитии иммунопатологии и взаимосвя-
тенденция была выявлена и для других отделов
занного с нею канцерогенеза. Так, ген Akt1 коди-
мозга и других групп генов, например, кодирую-
рует внутриклеточный фермент, RAC-альфа -
щих белки, участвующие в работе нейромедиа-
серин-треониновую протеинкиназу, для кото-
торных систем, митохондриальных, коллагено-
рой показано участие в процессах клеточного
вых, рибосомальных генов и др. [22-28, 39].
роста, пролиферации, а также в процессах ан-
Показано, что, за исключением генов
гиогенеза. Киназа AKT1 вовлечена в регуляцию
Tnfrsf22, Foxp2, Opa1 и Casp2, экспрессия кото-
пролиферации, роста и выживания клеток.
рых изменялась только под влиянием повторно-
Мутации гена Akt1 обнаружены в случаях рака
го опыта агрессии, у всех других генов экспрес-
молочной железы, рака толстой кишки, адено-
сия была изменена в гипоталамусе у обеих групп
карциномы лёгкого [44].
однонаправленно, не отличаясь по большин-
Ген Bag6 кодирует белки, которые положи-
ству ДЭГ. В то же время были обнаружены значи-
тельно регулируют апоптоз, взаимодействуя с
мые отличия в уровне экспрессии генов Alkbh7,
проапоптическим фактором AIFM1 и стабили-
Anxa1, Arc, Bax, Casp2, Casp8, Mapk3, Nosip, Pycard
зируя его. Контролируя устойчивую экспрес-
и Stk17b между экспериментальными группами,
сию рецептора IGF1R, он косвенно регулирует
что позволяет предположить их возможную роль
сигнальный путь рецептора инсулиноподобно-
в развитии более интенсивных процессов канце-
го фактора роста [45]. Для гена Foxp4 показана
рогенеза у депрессивных животных.
важная роль в развитии раковых заболеваний.
Однако необходимо обратить внимание
Он способствует клеточной пролиферации,
также на те гены, экспрессия которых не изме-
миграции, а также инвазии [46]. Также на роль
нялась у агрессивных животных, но изменялась
генов-«координаторов» претендуют два пред-
у депрессивных. Среди них присутствуют об-
ставителя семейства Mapk, кодирующие мито-
щепризнанные семейства генов, для которых
ген-активированные киназы Mapk3 и Mapk8,
неоднократно было подтверждено участие в
относящиеся к группе мультифункциональных
процессах канцерогенеза и/или апоптоза. Так,
внутриклеточных сигнальных путей, контро-
известно, что каспаза, белок, кодируемый ге-
лирующих транскрипцию генов, метаболизм,
ном Casp8, является инициатором запуска апоп-
пролиферацию и подвижность клеток, апоптоз
тоза по внутреннему механизму и некроптоза, в
и другие процессы. Хотя для гена Mapk3 более
то время как недостаток этого фермента приво-
характерно участие в других физиологических
дит к воспалительным процессам [40]. Проапоп-
процессах и в развитии различных патологиче-
тозная активность показана для белка, кодируе-
ских состояний, тем не менее известно, что для
мого геном Ccar1, являющегося стимулятором
многих форм рака отмечено изменение уровня
супрессора опухолей p53 [41]. Для белка синтазы
экспрессии этого гена [44]. Mapk8 традицион-
оксида азота, кодируемого геном Nos1, наоборот
но рассматривается в качестве гена, активация
характерна антиапоптозная активность [42]. Как
которого фактором некроза опухоли способна
об антиапоптозных агентах, говорят также и о
влиять на апоптоз [47]. В то же время известно,
белках, кодируемых генами Mcl1 и Bcl [43], хотя
что для белка, кодируемого этим геном, пока-
есть сведения, что на функционирование Mcl1
зан и антиапоптозный эффект, приводящий
может влиять альтернативный сплайсинг, пре-
даже к снижению эффективности противоопу-
вращая его то в антиапоптозный, то в проапоп-
холевой терапии [48]. Кроме того, белки, коди-
тозный агент. Возможно, именно белки, коди-
руемые этим геном, могут способствовать росту
руемые этими генами или какими-то из них,
опухоли [49].
оказывают, находясь в изменённом состоянии,
Ген Nol3 кодирует белок, который является
стимулирующее влияние на метастазирование
общепризнанным ингибитором апоптоза [50].
опухоли, показанное нами у депрессивных жи-
Ингибирование может происходить как за счёт
вотных в эксперименте [12-14].
блокировки каспаз - белков-эффекторов апоп-
Мы проанализировали ДЭГ Akt1, Bag6,
тоза, так и за счёт блокировки комплекса DISC
Foxp4, Mapk3, Mapk8, Nol3, Pdcd10 и Xiap, для
(death-inducing signaling complex), индуцирую-
которых было показано наибольшее количе-
щего апоптоз. Белки, кодируемые геном Pdcd10,
ство корреляций с экспрессией других ДЭГ со
выполняют большое количество внутрикле-
значением коэффициента корреляции r ≥ 0,950
точных функций, таких как образование меж-
и r ≤ -0,950, так как предположили, что они мо-
клеточных сцеплений цитоскелета, экзоцитоз
гут претендовать на роль генов, в наибольшей
и ангиогенез. Они участвуют также в регуля-
степени кодирующих белки, обеспечивающие
ции сигнальных путей [51]. Известно, что под
скоординированность процессов изменения
действием оксидативного стресса белки, коди-
БИОХИМИЯ том 87 вып. 9 2022
1
328
ГАЛЯМИНА и др.
руемые геном Pdcd10, взаимодействуют с се-
янии социального статуса на скорость роста
рин-треониновой киназой STK25, что приводит
пересаженных первичных опухолей или их есте-
к активации апоптоза [52]. В то же время извест-
ственное метастазирование в лёгких у мышей с
но, что этот ген может участвовать в усилении
негативным и позитивным социальным опы-
роста метастазов [53], что обычно связывают со
том [12-14, 59, 60].
снижением апоптоза. Показана двойная роль
Гипотеза, отвечающая на вопрос, но и тре-
этого гена в формировании нечувствительности
бующая доказательств. Неизбежно встаёт во-
к противораковой терапии [54].
прос: могут ли обнаруженные нами изменения
Антиапоптозный эффект белка, кодируемо-
экспрессии генов канцерогенеза и апоптоза в
го геном Xiap, установлен давно и подтвержда-
гипоталамусе у экспериментальных животных
ется новейшими исследованиями [55, 56]. Его
отражать некие изменения в периферических
антиапоптозные свойства используются для
тканях и органах и, в частности, в опухолях?
разработки более чувствительных методов про-
Скорее всего, это возможно, если предпо-
тивораковой терапии, способной вызывать
ложить скоординированность этих процессов
апоптоз только в опухолевых клетках [57]. Пола-
в мозге и периферических тканях (например,
гают, что снижение апоптоза является одной из
в лёгких) в том случае, если у генов изменяет-
причин усиления пролиферации клеток, и, сле-
ся экспрессия под влиянием одного и того же
довательно, ведёт к канцерогенезу [58]. Об этом
фактора, например кортикостероидов, других
же свидетельствуют наши данные по выявлению
гормонов или нейротрансмиттеров, выделя-
генов, у которых специфически была изменена
ющихся в кровь под влиянием какого-либо
экспрессия только у депрессивных животных.
воздействия, в нашем случае, хронического
Таким образом, мы можем предположить,
социального стресса. Это можно было бы под-
что однонаправленное изменение экспрессии
твердить, проведя одновременно исследование
изученных генов у животных с альтернативным
изменений экспрессии генов канцерогенеза и
социальным опытом в агонистических взаимо-
апопотоза в гипоталамусе и в ткани карциномы
действиях является следствием неспецифиче-
лёгких с увеличенным числом метастазов. Если
ской реакции на стресс, которому подвержены
так, то эти данные позволили бы найти пери-
обе группы экспериментальных животных с
ферические маркеры изменяющейся экспрес-
большей выраженностью изменений у депрес-
сии генов в мозге.
сивных животных. Изменение экспрессии уни-
С использованием литературных данных
кальных для депрессивных самцов генов явля-
мы провели сравнительный анализ между из-
ется результатом более жёсткого социального
менённой экспрессией генов канцерогене-
стресса, связанного не только с агонистиче-
за и апоптоза в гипоталамусе у депрессивных
скими столкновениями, но также с развитием
животных и состоянием топ-генов у больных
психоэмоционального расстройства. И в то же
с карциномой лёгких. Данные подробно из-
время неизменяющаяся экспрессия некоторых
ложены в разделе «Результаты исследования»
генов, отмеченная у агрессивных животных со
(табл. 6 в Приложении). Анализ показал, что
сниженным ростом метастазов, и специфиче-
по крайней мере гены Akt1, Braf, Casp8, Erbb2,
ски сниженная экспрессия только у депрессив-
Ercc6, Irf1, Kras и Pik3ca, изменения экспрессии
ных самцов может быть свидетельством важной
которых отмечены в гипоталамусе у депрес-
роли этих генов в стимуляции канцерогенеза.
сивных животных, участвуют в росте опухоли,
Иерархическая кластеризация ДЭГ в ка-
а экспрессия генов Ager, Clic5 и Aqp4 изменена
ждой группе агрессивных и депрессивных жи-
у больных с опухолью лёгких. Эти находки яв-
вотных позволяет выдвинуть предположение о
ляются косвенным доказательством того, что
зависимости функционирования гена от нейро-
в головном мозге, в частности в гипоталамусе,
химического окружения, определяемого разви-
изменённая экспрессия генов может в опреде-
тием той или иной патологии в нашем случае в
лённой степени соответствовать таким изме-
гипоталамусе, которые могут работать по-раз-
нениям в периферических тканях. Однако эта
ному в кооперации с разными генами. Тем не
гипотеза нуждается в экспериментальном под-
менее эти выводы необходимо подкрепить как
тверждении.
теоретически, так и экспериментально.
Кроме этого, очевидно и неоднократно по-
Полученные нейрогеномные данные, свиде-
казано, что на рост и метастазирование опухо-
тельствующие о возможном снижении апопто-
ли может влиять психоэмоциональный статус
за, как фактора, способствующего усиленному
индивида через модификацию его иммунной
канцерогенезу, могут, на наш взгляд, внести
реактивности. Имеются многочисленные дан-
вклад в понимание полученных ранее резуль-
ные, в том числе полученные во многих наших
татов о выраженном и воспроизводимом вли-
экспериментах, подтверждающие развитие
БИОХИМИЯ том 87 вып. 9 2022
ДИСФУНКЦИЯ ГЕНОВ ПРИ ХРОНИЧЕСКОМ СОЦИАЛЬНОМ СТРЕССЕ
1329
психогенного иммунодефицита у депрессивных
ЗАКЛЮЧЕНИЕ
животных под влиянием хронического социаль-
ного стресса, влияющего на рост опухолей [4, 6,
Хронический опыт агонистических взаимо-
7, 10, 61-65]. Таким образом, возможным объ-
действий вызывает в гипоталамусе изменение
яснением повышенного количества метастазов
экспрессии большого числа генов, кодирующих
в лёгких у депрессивных мышей в наших экс-
белки, ассоциируемые с механизмами канцеро-
периментах может быть снижение иммунитета.
генеза и апоптоза, как у животных со смешанным
В этом случае метастазирование опухоли будет
тревожно/депрессивным расстройством, сфор-
меньше у агрессивных животных, для которых
мированным под влиянием хронического соци-
предполагается развитие аутоиммунного забо-
ального стресса, так и у агрессивных животных
левания, сопровождающегося усилением им-
с повторным позитивным социальным опытом.
мунной реактивности [4, 66, 67]. Однако, мало-
В результате анализа были выделены гены (Akt1,
вероятно, что состояние иммунитета является
Bag6, Foxp4, Mapk3, Mapk8, Nol3, Pdcd10 и Xiap),
основным фактором, определяющим процессы
экспрессия которых максимально коррелирова-
метастазирования в этих экспериментах.
ла с экспрессией других генов, свидетельствуя о
Можно предположить также, что разная ин-
скоординированности нейрогеномных процес-
тенсивность метастазирования опухоли у агрес-
сов в этом отделе мозга. Эти гены представляют
сивных и депрессивных мышей может быть
интерес для дальнейшего более подробного из-
следствием различий их нейроэндокринного
учения с целью разработки методов фармаколо-
статуса, так как мозговая деятельность по-раз-
гической коррекции соматических нарушений,
ному изменяется под влиянием повторного по-
формирующихся под влиянием хронического
зитивного и негативного социального опыта в
социального стресса и поиска периферических
ежедневных агонистических взаимодействи-
маркеров нейрогеномных изменений.
ях [10, 35]. Нейротрансмиттеры могут влиять на
секрецию гормонов и таким образом приводить
к различным изменениям функций надпочеч-
Вклад авторов. Галямина А.Г., Смагин Д.А.,
ников и андрогенов у самцов мышей с проти-
Коваленко И.Л. - проведение экспериментов;
воположным типом социального поведения.
Редина О.Е., Бабенко В.Н. - обсуждение ре-
Некоторые нейроэндокринные факторы могут
зультатов исследования; Кудрявцева Н.Н., Га-
выступать в роли своеобразных факторов роста
лямина А.Г., Смагин Д.А. - написание текста;
и модифицировать развитие опухолей и мета-
Кудрявцева Н.Н. - концепция.
стазирование как путём прямого действия на
Финансирование. Работа выполнена при
опухолевые клетки, так и опосредованно через
финансовой поддержке Российского научного
влияние на сосудистое русло органов-мишеней
фонда (грант № 22-75-10095).
для метастазирования. Большое количество фи-
Благодарности. Авторы благодарны ЦКП
зиологических особенностей, таких как актив-
ность тканевых макрофагов и NK-клеток, про-
сква, Россия) за проведение технологической
ницаемость эндотелиальных капилляров, тонус
части эксперимента и первичный статисти-
и скорость васкуляризации и др., которые под-
ческий анализ. Авторы также благодарны
вергаются нейроэндокринной регуляции, могут
Н.А. Шевчуку за коррекцию английской вер-
влиять на колонизацию опухолевыми клетками
сии манускрипта (shevchuk-editing.com).
органов-мишеней и частоту метастазов [63, 68-
Конфликт интересов. Авторы заявляют об
70]. Имеется ряд работ, указывающих на то, что
отсутствии конфликта интересов.
усиленное метастазирование опухоли у депрес-
Соблюдение этических норм. Все экспери-
сивных животных купируется антагонистами
ментальные процедуры осуществляли в соот-
рецепторов некоторых нейромедиаторов [71-
ветствии с международными правилами про-
74]. Наши результаты по фармакологической
ведения экспериментов с животными (Directive
коррекции эмоционального статуса с помощью
2010/63/EU of the European Parliament and of
анксиолитика диазепама [8, 14, 60] или этано-
the Councilon the protection of animals used for
ла [59], которые снижали уровень тревожности
scientific purposes). Применяемые методики для
у депрессивных самцов мышей и при этом пре-
изучения поведения у мышей были одобрены
дотвращали развитие иммунных нарушений,
Научной комиссией № 9 Института цитологии
свидетельствуют о тесной взаимосвязи этих
и генетики СО РАН (Март, 24, 2010, N 613).
процессов. В то же время активация иммунной
Дополнительные материалы. Приложение к
системы с помощью хронического введения
статье опубликовано на сайте журнала «Биохи-
ронколейкина не влияла на этот процесс [75].
БИОХИМИЯ том 87 вып. 9 2022
1
330
ГАЛЯМИНА и др.
СПИСОК ЛИТЕРАТУРЫ
1.
Kudryavtseva, N. N., Bakshtanovskaya, I. V.,
12.
Каледин В. И., Кудрявцева Н. Н.
(1992)
Koryakina, L. A. (1991) Social model of depression in
Социальный конфликт и рост опухоли, Доклад.
mice of C57BL/6J strain, Pharmacol. Biochem. Behav.,
Ак. Наук, 234, 1117-1120.
38, 315-320, doi: 10.1016/0091-3057(91)90284-9.
13.
Каледин В. И., Тендитник М. В., Николин В. П.,
2.
Galyamina, A. G., Kovalenko, I. L., Smagin, D.
Попова Н. А., Кудрявцева Н. Н. (2006) Влияние
A., and Kudryavtseva, N. N. (2017). Interaction of
психоэмоционального состояния на рост и
depression and anxiety in the development of mixed
метастазирование опухоли Льюис у мышей, Докл.
anxiety/depression disorder. Experimental studies
Биол. Наук, 406, 57-59.
of the mechanisms of comorbidity, Neurosci. Behav.
14.
Kudryavtseva, N. N., Tenditnik, M. V., Nikolin, V. P.,
Physiol., 47, 699-713, doi: 10.1007/s11055-017-0458-3.
Popova, N. A., and Kaledin, V. I. (2007) The influ-
3.
Kudryavtseva, N. N. (2021) Development of mixed
ence of psychoemotional status on metastasis of Lew-
anxiety/depression-like state as a consequence
is lung carcinoma and hepatocarcinoma-29 in mice of
of chronic anxiety: review of experimental
C57BL/6J and CBA/Lac strains, Exp. Oncol., 29, 35-38.
data, Curr. Top. Behav. Neurosci.,
54,
125-152,
15.
Dubois, C., Vanden Abeele, F., and Prevarska-
doi: 10.1007/7854_2021_248.
ya, N. (2013) Targeting apoptosis by the remodel-
4.
Devoino, L. V., Alperina, E. L., Kudryavtseva, N. N.,
ling of calcium-transporting proteins in cancero-
and Popova, N. K. (1993) Immune responses in male
genesis, FEBS J.,
280,
5500-5510, doi:
10.1111/
mice with aggressive and submissive behavior patterns:
febs.12246.
strain differences, Brain. Behav. Immun., 7, 91-96,
16.
Kędzierska, H., and Piekiełko-Witkowska, A.
doi: 10.1006/brbi.1993.1009.
(2017) Splicing factors of SR and hnRNP families as
5.
Devoino, L., Idova, G., Alperina, E., and Cheido, M.
regulators of apoptosis in cancer, Cancer Lett., 396,
(1994) Brain neuromediator systems in the immune
53-65, doi: 10.1016/j.canlet.2017.03.013.
response control: pharmacological analysis of pre- and
17.
Hanahan, D., and Weinberg, R. A.
(2000)
postsynaptic mechanisms, Brain Res., 633, 267-274,
The hallmarks of cancer, Cell, 100, 57-70, doi: 10.1016/
doi: 10.1016/0006-8993(94)91548-2.
s0092-8674(00)81683-9.
6.
Попова Н. А., Ильницкая С. И., Колесникова Л. А.,
18.
Gross, A. L., Gallo, J. J., and Eaton, W. W. (2010)
Каледин, В. И., Кудрявцева, Н. Н. (1996) Влияние
Depression and cancer risk: 24 years of follow-up of
социального конфликта на некоторые параметры
the Baltimore Epidemiologic Catchment Area sample,
неспецифической резистентности у мышей, Росс.
Cancer Causes Control, 21, 191-199, doi: 10.1007/
Физиол. Журн. им. И.М. Сеченова, 82, 12-17.
s10552-009-9449-1.
7.
Тендитник М. В., Шурлыгина А. В., Мельникова
19.
Jia, Y., Li, F., Liu, Y. F., Zhao, J. P., Leng, M. M.,
Е. В., Кудрявцева Н. Н., Труфакин В. А. (2004)
et al. (2017) Depression and cancer risk: a systematic
Изменение субпопуляционного состава лимфо-
review and meta-analysis, Public Health, 149, 138-148,
цитов иммунокомпетентных органов мышей под
doi: 10.1016/j.puhe.2017.04.026.
влиянием хронического социального стресса, Росс.
20.
McEwen, B. S. (2016) Stress-induced remodeling of
Физиол. Журн. им. И.М. Сеченова, 90, 1522-1529.
hippocampal CA3 pyramidal neurons, Brain Res.,
8.
Тендитник М. В., Шурлыгина А. И., Мельникова
1645, 50-54, doi: 10.1016/j.brainres.2015.12.043.
Е. В., Пантелеева Н. Г., Смагин Д. А., и др. (2010)
21.
Patel, D., Anilkumar, S., Chattarji, S., and Buwalda,
Эффекты диазепама на субпопуляционный сос-
B. (2018) Repeated social stress leads to contrasting
тав лимфоцитов иммунокомпетентных органов
patterns of structural plasticity in the amygdala and
тревожных самцов мышей, Бюлл. СО РАМН,
hippocampus, Behav. Brain Res.,
347,
314-324,
30, 46-50.
doi: 10.1016/j.bbr.2018.03.034.
9.
Gryazeva, N. I. Shurlygina, A. V. Verbitskaya, L. V.
22.
Коваленко И. Л., Смагин Д. А., Галямина А. Г.,
Mel’nikova, E. V. Kudryavtseva, N. N., et al. (2001)
Орлов Ю. Л., Кудрявцева Н. Н. (2016) Изменение
Changes in various measures of immune status in mice
экспрессии дофаминергических генов в струк-
subject to chronic social conflict, Neurosci. Behav.
турах мозга самцов мышей под влиянием хрони-
Physiol., 31, 1, 75-81, doi: 10.1023/a:1026634532698.
ческого социального стресса: данные RNA-seq,
10.
Kudryavtseva, N. N., Shurlygina, A. V., Galyamina,
Мол. Биол., 50, 184-187, doi: 10.18699/VJ20.42-o.
A. G., Smagin, D. A., Popova, N. A., et al. (2019).
23.
Babenko, V. N., Smagin, D. A., Kovalenko, I. L., Ga-
Immunopathology of mixed anxiety/depression disor-
lyamina, A. G., and Kudryavtseva, N. N. (2020). Dif-
der: An experimental approach to studies of immuno-
ferentially expressed genes of the Slc6a family as mark-
deficiency state (review), Neurosci. Behav. Physiol., 49,
ers of altered brain neurotransmitter system function in
384-398, doi: 10.1007/s11055-019-00745-9.
pathological states in mice, Neurosci. Behav. Physiol.,
11.
Кудрявцева Н. Н., Шурлыгина А. В., Мельникова
50, 199-209, doi: 10.1007/s11055-019-00888-9.
Е. В., Тендитник М. В., Бондарь Н.П., и др. (2011)
24.
Kudryavtseva, N. N., Smagin, D. A., Kovalenko, I. L.,
Нарушение клеточного цикла в тимусе и селезенке
Galyamina, A. G., Vishnivetskaya, G. B., et al. (2017)
у самцов мышей под влиянием хронического
Serotonergic genes in the development of anxiety/de-
социального стресса: эффекты диазепама, Бюлл.
pression-like state and pathology of aggressive behav-
Экспер. Биол. Мед., 151, 391-394, doi: 10.1007/
ior in male mice: RNA-seq data, Mol. Biol., 51, 251-
s10517-011-1343-7.
262, doi: 10.7868/S0026898417020136.
БИОХИМИЯ том 87 вып. 9 2022
ДИСФУНКЦИЯ ГЕНОВ ПРИ ХРОНИЧЕСКОМ СОЦИАЛЬНОМ СТРЕССЕ
1331
25.
Smagin, D. A., Kovalenko, I. L., Galyamina, A. G.,
36.
Kudryavtseva, N. N., Smagin, D. A., Kovalenko, I.
Bragin, A. O., Orlov, Y. L., et al. (2016) Dysfunction
L., and Vishnivetskaya, G. B. (2014) Repeated positive
in ribosomal gene expression in the hypothalamus and
fighting experience in male inbred mice, Nat. Protoc.,
hippocampus following chronic social defeat stress
9, 2705-2717, doi: 10.1038/nprot.2014.156.
in male mice as revealed by RNA-Seq, Neural Plast.,
37.
Boyadjieva, N. I., and Sarkar, D. K. (2013) Microg-
2016, 3289187, doi: 10.1155/2016/3289187.
lia play a role in ethanol-induced oxidative stress and
26.
Smagin, D. A., Galyamina, A. G., Kovalenko,
apoptosis in developing hypothalamic neurons, Alco-
I. L., Babenko, V. N., and Kudryavtseva, N. N.
hol. Clin. Exp. Res., 37, 252-262, doi: 10.1111/j.1530-
(2019) Aberrant expression of collagen gene family
0277.2012.01889.x.
in the brain regions of male mice with behavioral
38.
Moraes, J. C., Coope, A., Morari, J., Cintra, D. E..,
psychopathologies induced by chronic agonistic
Roman, E. A., et al. (2009) High-fat diet induces
interactions, Biomed. Res. Int.,
2019,
7276389,
apoptosis of hypothalamic neurons, PLoS One, 4,
doi: 10.1155/2019/7276389.
e5045, doi: 10.1371/journal.pone.0005045.
27.
Redina, O., Babenko, V., Smagin, D., Kovalenko,
39.
Babenko, V. N., Smagin, D. A., Galyamina, A. G.,
I., Galyamina, A., et al. (2020) Gene expression
Kovalenko, I. L., and Kudryavtseva, N. N. (2018) Al-
changes in the ventral tegmental area of male mice
tered Slc25 family gene expression as markers of mi-
with alternative social behavior experience in chronic
tochondrial dysfunction in brain regions under exper-
agonistic interactions, Int. J. Mol. Sci., 21, 6599,
imental mixed anxiety/depression-like disorder, BMC
doi: 10.3390/ijms21186599.
Neurosci., 19, 79, doi: 10.1186/s12868-018-0480-6.
28.
Redina, O. E., Babenko, V. N., Smagin, D. A., Kova-
40.
Fritsch, M., Günther, S. D., Schwarzer, R., Albert, M.
lenko, I. L., Galyamina, A. G., et al. (2021) Correla-
C., Schorn, F., et al. (2019) Caspase-8 is the molecular
tion of expression changes between genes controlling
switch for apoptosis, necroptosis and pyroptosis, Na-
5-HT synthesis and genes Crh and Trh in the midbrain
ture, 575, 683-687, doi: 10.1038/s41586-019-1770-6.
raphe nuclei of chronically aggressive and defeat-
41.
Muthu, M., Cheriyan, V. T., and Rishi, A. K. (2015)
ed male mice, Genes (Basel), 12, 1811, doi: 10.3390/
CARP-1/CCAR1: a biphasic regulator of cancer
genes12111811.
cell growth and apoptosis, Oncotarget, 6, 6499-6510,
29.
Troubat, R., Barone, P., Leman, S., Desmidt, T.,
doi: 10.18632/oncotarget.3376.
Cressant, A., et al. (2021) Neuroinflammation and
42.
Wang, Q., Ye, S., Chen, X., Xu, P., Li, K., et al.
depression: A review, Eur. J. Neurosci., 53, 151-171,
(2019) Mitochondrial NOS1 suppresses apoptosis in
doi: 10.1111/ejn.14720.
colon cancer cells through increasing SIRT3 activ-
30.
Juruena, M. F., Bocharova, M., Agustini, B., and
ity, Biochem. Biophys. Res. Commun., 515, 517-523,
Young, A. H. (2018) Atypical depression and non-
doi: 10.1016/j.bbrc.2019.05.114.
atypical depression: Is HPA axis function a biomarker?
43.
Roberts, A. W., Wei, A. H., and Huang, D. C. S.
A systematic review, J. Affect. Disord., 233, 45-67,
(2021) BCL2 and MCL1 inhibitors for hematolog-
doi: 10.1016/j.jad.2017.09.052.
ic malignancies, Blood, 138, 1120-1136, doi: 10.1182/
31.
Carrera-Gonzalez, M. P., Ramirez-Exposito, M. J.,
blood.2020006785.
de Saavedra, J. M., Sanchez-Agesta, R., Mayas, M.
44.
AACR Project GENIE Consortium (2017) AACR
D., et al. (2011) Hypothalamus-pituitary-thyroid axis
Project GENIE: Powering Precision Medicine
disruption in rats with breast cancer is related to an al-
through an International Consortium, Cancer Discov.,
tered endogenous oxytocin/insulin-regulated amino-
7, 818-831, doi: 10.1158/2159-8290.CD-17-0151.
peptidase (IRAP) system, Tumour Biol., 32, 543-549,
45.
Osorio, F. G., Soria-Valles, C., Santiago-Fernández,
doi: 10.1007/s13277-010-0149-y.
O., Bernal, T., Mittelbrunn, M., et al. (2016) Loss
32.
Zuloaga, D. G., Carbone, D. L., Quihuis, A., Hiroi,
of the proteostasis factor AIRAPL causes myeloid
R., Chong, D. L., et al. (2012) Perinatal dexametha-
transformation by deregulating IGF-1 signaling, Nat.
sone-induced alterations in apoptosis within the hip-
Med., 22, 91-96, doi: 10.1038/nm.4013.
pocampus and paraventricular nucleus of the hypo-
46.
Zhang, G., and Zhang, G. (2019) Upregulation of
thalamus are influenced by age and sex, Neurosci. Res.,
FoxP4 in HCC promotes migration and invasion
90, 1403-1412, doi: 10.1002/jnr.23026.
through regulation of EMT, Oncol. Lett., 17, 3944-
33.
Buttenschon, H. N., Krogh, J., Nielsen, M. N., Kaer-
3951, doi: 10.3892/ol.2019.10049.
lev, L., Nordentoft, M., et al. (2017) Association anal-
47.
Hua, X., Chen, J., and Wu, L. (2019) Identification
yses of depression and genes in the hypothalamus-pi-
of candidate biomarkers associated with apopto-
tuitary-adrenal axis, Acta Neuropsychiatr., 29, 59-64,
sis in melanosis coli: GNG5, LPAR3, MAPK8, and
doi: 10.1017/neu.2016.26.
PSMC6, Biosci. Rep., 39, BSR20181369, doi: 10.1042/
34.
Kudryavtseva, N. N. (1991) A sensory contact mod-
BSR20181369.
el for the study of aggressive and submissive be-
48.
Xu, P., Zhang, G., Hou, S., and Sha, L. G. (2018)
havior in male mice, Aggress. Behav., 17, 285-291,
MAPK8 mediates resistance to temozolomide and
doi: 10.1002/1098-2337(1991)17:5<285::AID-AB2480
apoptosis of glioblastoma cells through MAPK signal-
170505>3.0.CO;2-P.
ing pathway, Biomed. Pharmacother., 106, 1419-1427,
35.
Kudryavtseva, N. N.
(2000) Agonistic behavior:
doi: 10.1016/j.biopha.2018.06.084.
A model, experimental studies, and perspectives,
49.
Chen, W., Zheng, G., Huang, J., Zhu, L., Li, W.,
Neurosci. Behav. Physiol., 30, 293-305, doi: 10.1007/
et al. (2021) CircMED13L_012 promotes lung ade-
BF02471782.
nocarcinoma progression by upregulation of MAPK8
БИОХИМИЯ том 87 вып. 9 2022
1
332
ГАЛЯМИНА и др.
mediated by miR-433-3p, Cancer Cell Int., 21, 111,
63.
Spiegel, D., and Sephton, S. E. (2001) Psychoneuro-
doi: 10.1186/s12935-021-01811-4.
immune and endocrine pathways in cancer: effects of
50.
Sohn, E. J., Shin, M. J., Eum, W. S., Kim, D. W., Yong,
stress and support, Semin. Clin. Neuropsychiatry, 6,
J. I., et al. (2016) Tat-NOL3 protects against hippo-
252-265, doi: 10.1053/scnp.2001.26995.
campal neuronal cell death induced by oxidative stress
64.
Vegas, O., Beitia, G., Sánchez-Martin, J. R.,
through the regulation of apoptotic pathways, Int. J.
Arregi, A., and Azpiroz, A. (2004) Behavioral and
Mol. Med., 38, 225-235, doi: 10.3892/ijmm.2016.2596.
neurochemical responses in mice bearing tumors
51.
Valentino, M., Dejana, E., and Malinverno, M. (2020)
submitted to social stress, Behav. Brain Res., 155, 125-
The multifaceted PDCD10/CCM3 gene, Genes Dis., 8,
134, doi: 10.1016/j.bbr.2004.04.006.
798-813, doi: 10.1016/j.gendis.2020.12.008.
65.
Azpiroz, A., Garmendia, L., Fano, E., and San-
52.
Zhang, H., Ma, X., Deng, X., Chen, Y., Mo, X., et al.
chez-Martin, J. R. (2003) Relations between aggres-
(2012) PDCD10 interacts with STK25 to accelerate
sive behavior, immune activity, and disease suscepti-
cell apoptosis under oxidative stress, Front. Biosci.
bility, Aggress. Viol. Behav., 8, 433-453.
(Landmark Ed), 17, 2295-2305, doi: 10.2741/4053.
66.
Шурлыгина А. В., Мельникова Е. В., Коваленко
53.
Dong, C., Fan, B., Ren, Z., Liu, B., and Wang, Y.
И. Л., Галямина А. Г., Грицык О. Б., и др. (2014, а)
(2021) CircSMARCA5 facilitates the progression
Изменения иммунного статуса у самцов мышей,
of prostate cancer through miR-432/PDCD10 axis,
вызванные повторным опытом агрессии, Росс.
Cancer Biother. Radiopharm., 36, 70-83, doi: 10.1089/
Физиол. Журн. им. И.М.Сеченова, 11, 1268-1279.
cbr.2019.3490.
67.
Alperina, E., Idova, G., Zhukova, E., Zhanaeva,
54.
Urfali-Mamatoglu, C., Kazan, H. H., and Gündüz,
S., and Kozhemyakina, R. (2019) Cytokine varia-
U. (2018) Dual function of programmed cell death 10
tions within brain structures in rats selected for dif-
(PDCD10) in drug resistance, Biomed. Pharmacother.,
ferences in aggression, Neurosci. Lett., 692, 193-198,
101, 129-136, doi: 10.1016/j.biopha.2018.02.020.
doi: 10.1016/j.neulet.2018.11.012.
55.
Liu, J., Chen, Z., Cui, Y., Wei, H., Zhu, Z., et al.
68.
Rowse, G. J., Weinberg, J., Bellward, G. D., and Emer-
(2020) Berberine promotes XIAP-mediated cells
man, J. T. (1992) Endocrine mediation of psychosocial
apoptosis by upregulation of miR-24-3p in acute
stressor effects on mouse mammary tumor growth, Can-
lymphoblastic leukemia, Aging (Albany NY), 12, 3298-
cer Lett., 65, 85-93, doi: 10.1016/0304-3835(92)90217-j.
3311, doi: 10.18632/aging.102813.
69.
Giraldi, T., Perissin, L., Zorzet, S., Rapozzi, V., and
56.
Yang, W.Z., Zhou, H., Yan, Y. (2018) XIAP underlies
Rodani, M. G. (1994) Metastasis and neuroendocrine
apoptosis resistance of renal cell carcinoma cells, Mol.
system in stressed mice, Int. J. Neurosci., 74, 265-278,
Med. Rep., 17, 125-130, doi: 10.3892/mmr.2017.7925.
doi: 10.3109/00207459408987245.
57.
Saraei, R., Soleimani, M., Movassaghpour Akbari,
70.
Palermo-Neto, J., de Oliveira Massoco, C., and Robe-
A. A, Farshdousti Hagh, M., Hassanzadeh, A., et al.
spierre de Souza, W. (2003) Effects of physical and
(2018) The role of XIAP in resistance to TNF-relat-
psychological stressors on behavior, macrophage activ-
ed apoptosis-inducing ligand (TRAIL) in leukemia,
ity, and Ehrlich tumor growth, Brain Behav. Immun.,
Biomed. Pharmacother., 107, 1010-1019, doi: 10.1016/
1, 43-54, doi: 10.1016/s0889-1591(02)00057-0.
j.biopha.2018.08.065.
71.
Perissin, L., Rapozzi, V., Zorzet, S., and Giraldi, T.
58.
Pfeffer, C. M., and Singh, A. T. K. (2018) Apoptosis:
(1996) Blockers of adrenergic neurons and receptors,
A target for anticancer therapy, Int. J. Mol. Sci., 19,
tumor progression and effects of rotational stress in
448, doi: 10.3390/ijms19020448.
mice, Anticancer Res., 16, 3409-3413.
59.
Ильницкая С. И., Николин В. П., Попова Н. А.,
72.
Ben-Eliyahu, S, Shakhar, G., Page, G. G.,
Августинович Д. Ф., Каледин В. И., и др. (2009)
Stefanski, V., and Shakhar, K. (2000) Suppression
Влияние этанола на процессы метастазирования
of NK cell activity and of resistance to metastasis by
аденокрациномы Льюис у самцов мышей с разным
stress: a role for adrenal catecholamines and beta-
эмоциональным состоянием, Росс. Физиол. Журн.
adrenoceptors, Neuroimmunomodulation, 8, 154-164,
им. И.М. Сеченова, 95, 74-78.
doi: 10.1159/000054276.
60.
Kaledin, V. I., Ilnitskaya, S. I., Nikolin, V. P., Popova,
73.
Freire-Garabal, M., Rey-Méndez, M., García-
N. A., Smagin, D. A., et al. (2009) Limiting effect
Vallejo, L. A., Balboa, J., Suárez, J. M., et al.
of diazepam on Lewis lung carcinoma metastasis in
(2004) Effects of nefazodone on the development
anxious male mice, Exp. Oncol., 31, 62-64.
of experimentally induced tumors in stressed
61.
Strange, K. S., Kerr, L. R., Andrews, H. N., Emer-
rodents, Psychopharmacology (Berl), 176, 233-238,
man, J. T., and Weinberg, J. (2000) Psychosocial
doi: 10.1007/s00213-004-1909-4.
stressors and mammary tumor growth: an animal
74.
Palm, D., Lang, K., Niggemann, B., Drell IV, T. L.,
model, Neurotoxicol. Teratol., 22, 89-102, doi: 10.1016/
Masur, K., et al. (2006) The norepinephrine-driven
s0892-0362(99)00049-5.
metastasis development of PC-3 human prostate cancer
62.
Wu, X., Liu, B. J., Ji, S., Wu, J. F., Xu, C. Q., et al.
cells in BALB/c nude mice is inhibited by β-blockers,
(2015) Social defeat stress promotes tumor growth
Int. J. Cancer, 118, 2744-2749, doi: 10.1002/ijc.21723.
and angiogenesis by upregulating vascular endotheli-
75.
Shurlygina, A. V., Galiamina, A. G., Mel’nikova, E. V.,
al growth factor/extracellular signal-regulated kinase/
Panteleeva, N. G., Tenditnik, M. V., et al. (2015) Ef-
matrix metalloproteinase signaling in a mouse mod-
fects of roncoleukin on measures of immunity and the
el of lung carcinoma, Mol. Med. Rep., 12, 1405-1412,
anxious-depressive state induced by chronic social stress
doi: 10.3892/mmr.2015.3559.
in male mice, Neurosci. Behav. Physiol., 45, 8, 902-909.
БИОХИМИЯ том 87 вып. 9 2022
ДИСФУНКЦИЯ ГЕНОВ ПРИ ХРОНИЧЕСКОМ СОЦИАЛЬНОМ СТРЕССЕ
1333
DYSFUNCTION OF THE GENES ASSOCIATED
WITH CARCINOGENESIS AND APOPTOSIS,
DEVELOPING IN THE HYPOTHALAMUS
UNDER CHRONIC SOCIAL DEFEAT STRESS IN MALE MICE
A. G. Galyamina1#, D. A. Smagin1#, I. L. Kovalenko1, O. E. Redina1,
V. N. Babenko1, N. N. Kudryavtseva1,2*
1 FRC Institute of Cytology and Genetics, Siberian Branch of Russian Academy of Sciences,
630090 Novosibirsk, Russia; e-mail: natnik@bionet.nsc.ru
2 Pavlov Institute of Physiology, Russian Academy of Sciences, 199034 Saint-Petersburg, Russia
Chronic social stress caused by daily agonistic interactions in male mice leads to the development of a mixed
anxiety/depression-like disorder, which is accompanied by the development of psychogenic immunodeficiency
and stimulation of oncological processes, accompanied by many neurogenomic changes in brain regions.
The aim of the study was to identify differentially expressed genes (DEGs) involved in the mechanisms of
carcinogenesis and apoptosis in the hypothalamus of male mice with depression-like symptoms and, for
comparison, in aggressive male mice with positive social experience. To obtain animals with a 20-day
alternative social experience, a model of chronic social conflict was used. The analysis of mouse hypothalamic
transcriptome data (RNA-Seq) revealed similar changes in many DEGs in aggressive and depressed animals
in relation to the control state, however, the number of differentially expressed genes was significantly less in
aggressive than in depressed males. It can be assumed that a unidirectional genes expression change associated
with the oncogenesis and apoptosis processes may be the result of a long-term social stress effect of different
severity caused by agonistic interactions. In addition, 26 genes with unchanged expression were found in
aggressive animals, which can provoke carcinogenesis or inhibit apoptosis. We suppose that the Akt1, Bag6,
Foxp4, Mapk3, Mapk8, Nol3, Pdcd10 and Xiap genes, the expression of which was most correlated with the
expression of other DEGs, makes it possible to put forward them as coordinating neurogenomic changes in the
hypothalamus. Further study of the role of these genes may be useful for the development of methods for the
pharmacological correction of psychosomatic pathologies.
Keywords: apoptosis, carcinogenesis, RNA-Seq, DEG, hypothalamus, chronic social stress
БИОХИМИЯ том 87 вып. 9 2022