БИОХИМИЯ, 2023, том 88, вып. 4, с. 656 - 670
УДК 577.25;612.82;616-005
СРАВНИТЕЛЬНОЕ ИССЛЕДОВАНИЕ ЭКСПРЕССИИ ГЕНОВ
ГЛУТАМАТЕРГИЧЕСКОЙ И ГАМКЕРГИЧЕСКОЙ СИСТЕМ
В ГИППОКАМПЕ КРЫСЫ ПОСЛЕ ОККЛЮЗИИ СРЕДНЕЙ
МОЗГОВОЙ АРТЕРИИ И ЦЕНТРАЛЬНОГО ВВЕДЕНИЯ
ЛИПОПОЛИСАХАРИДА
© 2023 Т.С. Калинина1, Г.Т. Шишкина1*, Д.А. Ланшаков1, Е.В. Сухарева1,
М.В. Онуфриев2, Ю.В. Моисеева2, Н.В. Гуляева2, Н.Н. Дыгало1
1 ФИЦ Институт цитологии и генетики СО РАН,
600090 Новосибирск, Россия; электронная почта: gtshi@bionet.nsc.ru
2 ФГБУН Институт высшей нервной деятельности и нейрофизиологии РАН, 117485 Москва, Россия
Поступила в редакцию 23.11.2022
После доработки 14.03.2023
Принята к публикации 14.03.2023
Среди ответов на ранних стадиях инсульта ключевое значение для развития постишемических
функциональных последствий имеет активация нейродегенеративных и провоспалительных про-
цессов в гиппокампе. Однако остается не вполне ясным, какие именно гены вовлечены в эти про-
цессы. Целью данной работы было сравнительное исследование экспрессии генов, кодирующих
транспортеры и рецепторы глутамата и ГАМК, а также маркеров воспаления в гиппокампе через
сутки после двух видов ишемического воздействия (окклюзии средней мозговой артерии (MCAO)
по модели Коизуми - MCAO-MK и по модели Лонга - MCAO-ML), а также после прямой про-
воспалительной активации центральным введением бактериального липополисахарида (LPS).
Полученные результаты выявили различия и сходства между ответами на примененные в работе
воздействия. После MCAO-ML и введения LPS обнаружено большее число изменивших экспрес-
сию генов, связанных с активацией апоптоза и нейровоспаления, рецепцией глутамата, а также
маркеров ГАМКергической системы, чем после MCAO-MK. В свою очередь, MCAO-MK и LPS
характеризовались, по сравнению с MCAO-ML, изменением большего числа генов, вовлекаемых в
транспорт глутамата. Наиболее ярко проявившимися отличиями MCAO-ML от MCAO-MK и LPS
явились изменения в экспрессии генов кальмодулина и кальмодулин-зависимых киназ. Выявлен-
ные особенности ответов транскриптома гиппокампа на две модели ишемии в результате MCAO
и прямой провоспалительный стимул будут способствовать дальнейшему пониманию причин
и молекулярных механизмов, лежащих в основе разнообразия последствий инсультов как в модель-
ных исследованиях, так и в клинике.
КЛЮЧЕВЫЕ СЛОВА: гиппокамп, окклюзия средней мозговой артерии, липополисахарид, глутамат,
ГАМК, экспрессия генов.
DOI: 10.31857/S0320972523040097, EDN: ALGGNS
ВВЕДЕНИЕ
ное голодание. Однако при ишемии погибают
клетки и в «отдаленных» структурах мозга, та-
Острое нарушение кровоснабжения обла-
ких как гиппокамп, непосредственно не полу-
сти мозга, преимущественно неокортекса, в
чающие кровоснабжение по поврежденным
результате ишемии в бассейне средней мозго-
сосудам [1]. Гиппокамп играет ключевую роль
вой артерии в течение нескольких часов про-
в контроле психоэмоционального состояния,
воцирует гибель клеток и формирование очага
обучения и памяти [2]. С дегенерацией нейро-
инфаркта. Причиной этих событий считают
нов этой структуры связывают развитие пси-
нехватку питательных веществ и кислород- хопатологий, в том числе и постишемической
Принятые сокращения: LPS - липополисахарид; MCAO - окклюзия средней мозговой артерии; MCAO-MK - модель
МСАО Коизуми; MCAO-ML - модель МСАО Лонга.
* Адресат для корреспонденции.
656
ГЕНЫ ГЛУТАМАТ- И ГАМКЕРГИЧЕСКОЙ СИСТЕМ ПОСЛЕ MCAO И LPS
657
деменции [3]. Поэтому выяснение механизмов
трансмиттерных систем. Новые сведения в
повреждающего влияния ишемии на гиппо-
данной области могут быть получены путем
камп важно для дальнейшего поиска средств
сравнения данных, полученных на разных
ослабления негативных функциональных по-
моделях. Например, оказалось, что две наи-
следствий ишемического воздействия. Нару-
более часто используемые модели MCAO
шение кровотока по средней мозговой артерии
(«золотой стандарт» моделирования ише-
считают причиной значительной части всех
мического инсульта на грызунах [18]) значи-
ишемических патологий мозга [4], поэтому
тельно различаются по ряду функциональ-
окклюзию средней мозговой артерии (MCAO)
ных параметров, в том числе связанных с
используют для экспериментального модели-
эксайтотоксичностью глутамата и воспали-
рования ишемического инсульта на грызунах.
тельными процессами [19, 20]. С учетом этих
Среди предполагаемых путей провоциро-
результатов целью данной работы было про-
вания острой клеточной гибели после ишемии
ведение сравнительного исследования экс-
значительное внимание уделяется эксайтоток-
прессии генов, кодирующих транспортеры и
сическому действию глутамата [5, 6]. Однако
рецепторы глутамата и ГАМК, а также мар-
глутаматергическая система вовлекается во
керов воспаления в гиппокампе после двух
множество жизненно важных функций орга-
видов ишемического воздействия, модели
низма [7], в том числе и в положительную регу-
MCAO Коизуми (MCAO-MK) [21] и модели
ляцию возбуждающими стимулами дифферен-
Лонга (MCAO-ML) [22], а также после пря-
цировки нейронов [8]. Неоднозначное влияние
мой провоспалительной активации путем
глутамата на разные процессы, определяющие
центрального введения LPS. Недавно обнару-
жизнеспособность клеток, затрудняет выбор
жено, что модели ишемии по Коизуми и Лонга
способов воздействия на эту систему для те-
различаются по накоплению глюкокортикоид-
рапии вызванных ишемией повреждений, а
ных гормонов и провоспалительных цито-
также сроков их применения, например, для
кинов в гиппокампе в остром периоде после
лигандов глутаматергических рецепторов. Так,
ишемии [20], при этом различия сохраняются
накапливаются свидетельства, что увеличение
в течение месяцев [23]. Исследование особен-
экспрессии одних и тех же типов рецепторов
ностей провоспалительной активации, а также
глутамата в острой фазе инсульта может быть
нарушений экспрессии генов глутаматергиче-
усугубляющим постишемическое восстановле-
ской системы и оценка возможных нарушений
ние, в то время как их увеличение в более позд-
в других нейромедиаторных системах, прежде
ней фазе, напротив, благоприятным [9]. Оче-
всего, тормозной ГАМКергической, принци-
видно, что из-за неполного понимания роли
пиально важно для дальнейшего прояснения
отдельных компонентов глутаматергической
механизмов острого ишемического поражения
системы в ишемическом повреждении мозга
гиппокампа.
предпринимаемые попытки напрямую воздей-
ствовать на эту нейрохимическую систему с
целью предотвращения или замедления гибели
МАТЕРИАЛЫ И МЕТОДЫ
клеток не всегда приводили к желаемым эф-
фектам [10, 11].
Животные. Влияние центрального введе-
Ишемическое повреждение мозга сопро-
ния LPS исследовали на взрослых самцах крыс
вождается нейровоспалением
- активацией
линии Вистар, содержавшихся в стандартных
глиальных клеток, продукцией и секрецией
условиях вивария ИЦиГ СО РАН при темпера-
цитокинов в гиппокампе [12]. В исследованиях
туре 22-24°C, световом режиме 14С/10Т и сво-
патофизиологических механизмов нейродеге-
бодном доступе к воде и корму.
неративных заболеваний, индуцируемых в том
Влияние MCAO исследовали на взрослых
числе и ишемией, введение липополисахари-
самцах крыс линии Вистар, содержавшихся
да (LPS) получило широкое распространение
в стандартных условиях вивария Института
в качестве эффективного модельного подхода
высшей нервной деятельности и нейрофизио-
к оценке роли различных участников воспа-
логии РАН.
лительного процесса в этих заболеваниях [13].
Моделирование инсульта проводили по стан-
Полногеномные анализы транскриптомов гиппо-
дартным протоколам экспериментальной ише-
кампа через сутки после экспериментальной
мии (модель Коизуми
[20] и модель Лон-
ишемии [14, 15, 16] или центрального введе-
га [21]), а также прямой провоспалительной
ния LPS [16,
17] выявили изменение экс-
активации путем центрального введения LPS.
прессии множества генов, в том числе генов
Модель Коизуми (MCAO-MK). Под анесте-
регуляторов апоптоза, воспаления, нейро-
зией изофлураном выполняли разрез в области
9
БИОХИМИЯ том 88 вып. 4 2023
658
КАЛИНИНА и др.
шеи и, оттесняя мышечную ткань с левой сто-
ли таких дефицитов. По 5-балльной шкале все
роны, накладывали лигатуры на общую сон-
ложнооперированные животные имели балл 0,
ную артерию, а также на наружную и внут-
в то время как животные после MCAO в мо-
реннюю сонные артерии. Нейлоновую нить
делях Коизуми или Лонга демонстрировали
(3-0) с закругленным концом вводили через
тяжелый дефицит 4 балла во время операции
отверстие в месте бифуркации на наружную и
и 3 балла - через сутки после операции перед
внутреннюю ветви и продвигали по внутрен-
забоем. Через сутки после операции животные
ней сонной артерии до средней мозговой арте-
с MCAO независимо от модели демонстриро-
рии. Затем затягивали лигатуру на внутренней
вали неспособность высунуть язык, в то время
сонной артерии для фиксации нити. Окклю-
как у ложнооперированных крыс эта способ-
зию проводили в течение 60 мин, при этом
ность сохранялась на 92-100%. Следует отме-
общая, наружная и внутренняя сонные арте-
тить, что значения дефицитов в данной работе
рии оставались перевязанными, а температу-
совпадают с таковыми через сутки после ише-
ра тела животного поддерживалась на уровне
мии, представленными в предыдущих экспе-
37 ± 0,5 °С. Затем нить удаляли и затягивали
риментах [20].
лигатуру на внутренней сонной артерии. Кон-
Центральное введение LPS. Под анестезией
тролем являлись ложнооперированные живот-
изофлураном в правый стриатум вводили бакте-
ные, с которыми выполняли все манипуляции,
риальный эндотоксин липополисахарид (LPS)
кроме введения нити.
в дозе 30 мкг в 4 мкл физраствора по стереотак-
Модель Лонга (MCAO-ML). Под анесте-
сическим координатам: AP = + 0,5 мм, ML =
зией изофлураном через разрез на шее живот-
= + 3 мм, DV = -4,5 мм [16] в течение 5 мин.
ного лигировали общую сонную и наружную
Контрольным животным вводили в аналогич-
сонную артерии. После электрокоагуляции и
ных условиях 4 мкл физраствора. LPS вводили
диссекции фрагмента левой наружной сонной
в стриатум, поскольку он входит в число перво-
артерии через оставшуюся часть артерии вво-
очередных повреждаемых ишемическим воз-
дили филамент из нейлоновой нити до пере-
действием структур мозга. Например, у взрос-
сечения со средней мозговой артерией. Ок-
лых крыс
30-минутная MCAO по Коизуми
клюзия продолжалась в течение 60 мин, при
вызывала повреждение, ограниченное полоса-
этом температура тела крысы поддерживалась
тым телом, тогда как после этой MCAO в тече-
на уровне 37 ± 0,5 °С. Затем нить извлекали,
ние 2 ч наблюдались «инфаркты» как в стриа-
что восстанавливало кровоток по ипсилате-
туме, так и в коре [24].
ральной общей сонной артерии. Контролем
Секвенирование РНК. Через 24 ч после
являлись ложнооперированные животные, с
каждого из трех воздействий животных заби-
которыми выполняли все манипуляции, кроме
вали декапитацией, на холоду выделяли гиппо-
введения нити.
камп, который помещали в раствор RNAlater
В связи с необходимостью использова-
(«Life Technologies», США) и хранили при тем-
ния ткани мозга для анализа транскриптома в
пературе -70 °С до секвенирования, которое
качестве подтверждения формирования ише-
выполняли в ООО «Геноаналитика» (https://
мического очага у животных использовали
genoanalytica.ru, Москва). Тотальную РНК
оценку неврологических дефицитов, которая
выделяли из гиппокампа трех животных каж-
сопровождает формирование ишемического
дой экспериментальной и контрольной групп
очага [20]. Использовали тест, основанный на
при помощи реагента Trizol («Thermo Fisher
5-балльной поведенческой шкале, позволяю-
Scientific», США) в соответствии с рекомен-
щий оценить функциональное состояние кон-
дациями производителя. Качество и количе-
тралатеральной передней лапы крыс, наличие
ство выделенной тотальной РНК проверяли
поворотов и циркуляции в контралатеральную
при использовании набора RNA 6000 Nano Kit
сторону и общую подвижность животных, а
(«Agilent Technologies, США) на приборе
также тест на степень высовывания языка -
BioAnalyzer («Agilent Technologies»). Фракцию
по способности крысы вылизывать арахисо-
полиА из тотальной РНК получали при помо-
вое масло из стеклянного цилиндра с после-
щи магнитных шариков олигоТ Dynabeads®
дующим измерением расстояния от начала
mRNA Purification Kit («Thermo Fisher Scien-
цилиндра до уровня оставшегося масла [20].
tific»). Далее из полиА-РНК были приготов-
Все животные с MCAO, взятые в эксперимент,
лены библиотеки для массового параллель-
имели выраженные неврологические дефи-
ного секвенирования при помощи набора
циты, свидетельствующие о формировании
NEBNext® Ultra™ II RNA Library Prep («NEB»,
ишемического очага, в то время как ложно-
США), согласно инструкции к набору. Кон-
оперированные животные не демонстрирова-
центрацию библиотек определяли при помощи
БИОХИМИЯ том 88 вып. 4 2023
ГЕНЫ ГЛУТАМАТ- И ГАМКЕРГИЧЕСКОЙ СИСТЕМ ПОСЛЕ MCAO И LPS
659
набора Qubit dsDNA HS Assay Kit («Thermo
дом массового параллельного секвенирования
Fisher Scientific») на приборе Qubit 2.0 («Ther-
транскриптома, считалось достоверным при
mo Fisher Scientific»). Распределение длин
скорректированном p-значении с поправ-
фрагментов библиотеки проводили при по-
кой на множественность сравнений (adjusted
мощи набора Agilent High Sensitivity DNA Kit
p-value - padj) меньшем, чем 0,05. При вери-
(«Agilent Technologies»). Секвенирование про-
фикации уровня экспрессии методом ПЦР в
водили на приборе HiSeq1500
(«Illumina»,
реальном времени достоверность устанавлива-
США) с генерацией не менее 10 млн коротких
ли путем прямых парных сравнений с исполь-
чтений длиной 50 нуклеотидов. Риды вырав-
зованием t-критерия Стьюдента в программе
нивали на геном версии Rnor_6.0 при помощи
Statistica 6.0.
программы STAR, и подсчет дифференци-
ально экспрессирующихся генов проводили,
используя пакет DESeq2.0.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
ПЦР в реальном времени. Выявленные сек-
венированием изменения в экспрессии генов
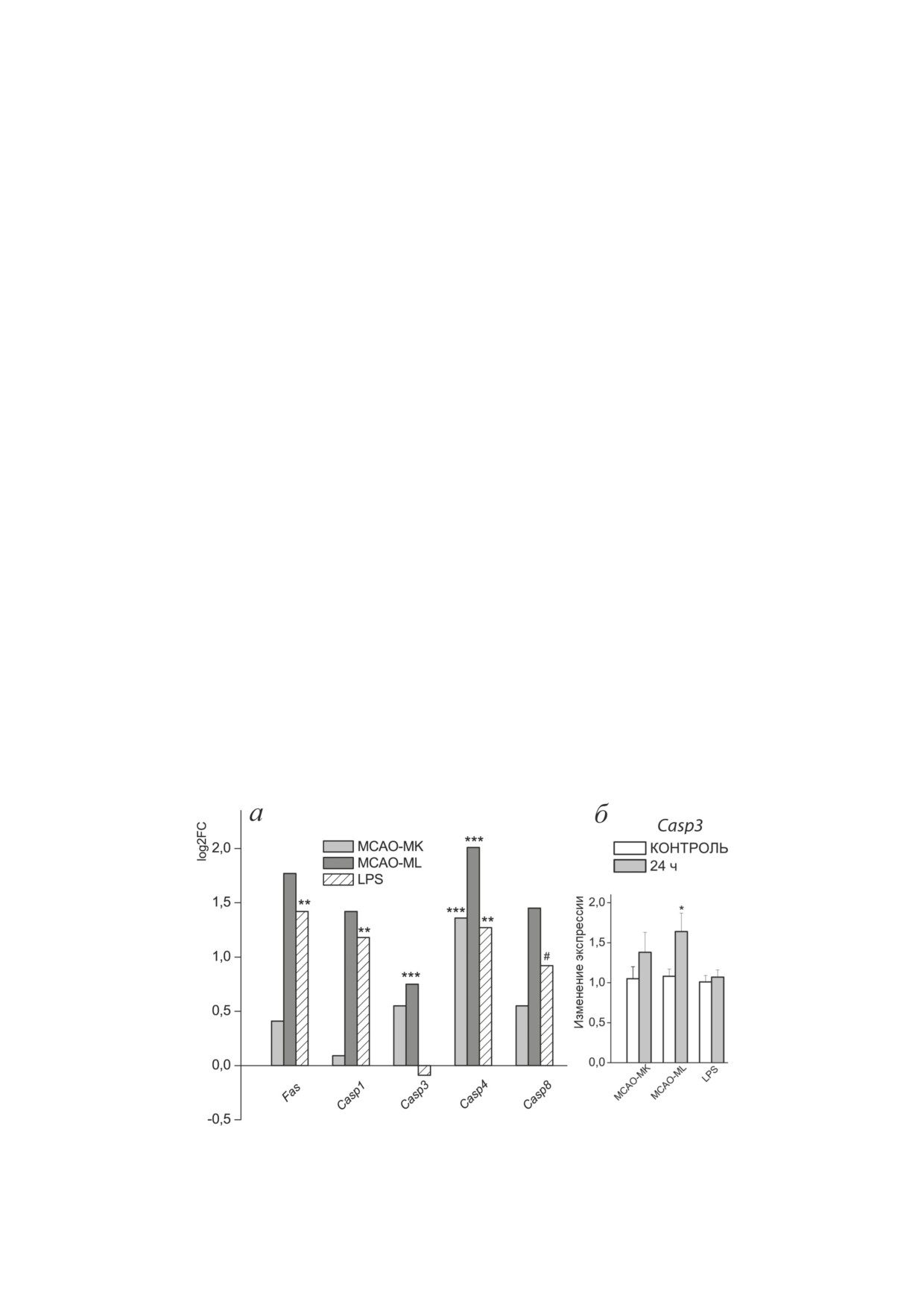
Гены, ассоциированные с гибелью клеток.
выборочно верифицировались методом ПЦР
Признаки гибели клеток в гиппокампе че-
в реальном времени на амплификаторе VIIA7
рез сутки после перенесенных воздействий
с применением технологии TaqMan («Thermo
обнаружены после MCAO-ML, на что, по
Fisher Scientific»). Выделение суммарной РНК,
результатам секвенирования, указывало до-
оценку ее качества и синтез кДНК проводи-
стоверное увеличение экспрессии гена ис-
ли по ранее описанным стандартным прото-
полнительной протеазы апоптоза каспазы-3
колам [25]. Уровни экспрессии генов: Casp3
(Сasp3) (рис. 1, а), подтвержденное методом
(Rn00563902_m1), IL1b (Rn00580432_m1), Slc1a2
ПЦР в реальном времени (p < 0,05; рис. 1, б).
(Rn00691548_m1), Gria2 (Rn00568514_m1), Glul
После всех воздействий
- MCAO-ML,
(Rn01483107_m1), Gad2 (Rn00561244_m1) опре-
MCAO-MK и LPS - была также увеличена
деляли в двух повторах в образцах РНК по-
экспрессия гена Сasp4. Кроме того, после LPS
сле секвенирования с добавлением до восьми
были достоверно увеличены экспрессии гена
образцов РНК из тех же экспериментальных
рецептора смерти Fas, Сasp1 и Сasp8 на уровне
групп. Все реакции проводили в соответствии
тенденции - p-value = 4,55E-03; padj = 0,0946.
с рекомендациями производителя, и уровень
Все эти гены были ассоциированы с выяв-
экспрессии генов рассчитывали по методу
ленными в результате анализа генной онто-
ddCt относительно экспрессии гена домашне-
логии биологическими процессами, такими
го хозяйства бета-актина (Rn00667869_m1).
как «апоптоз» («apoptotic process»), актива-
Статистическая обработка данных. Изме-
ция апоптоза нейронов («positive regulation of
нение экспрессии генов, определенное мето-
neuron apoptotic process»), активация процесса
Рис. 1. Изменение экспрессии генов, ассоциированных с гибелью клеток, после двух типов ишемического воздей-
ствия и центрального введения LPS. а - По результатам секвенирования; ** padj < 0,01; *** padj < 0,001; # padj < 0,1.
б - Верификация ПЦР в реальном времени; * p < 0,05
БИОХИМИЯ том 88 вып. 4 2023
9*
660
КАЛИНИНА и др.
апоптоза («positive regulation of apoptotic pro-
(рис. 2, б) верифицировал увеличение экс-
cess»), усиление гибели клеток («positive regula-
прессии гена интерлейкина после MCAO-ML
tion of cell death»).
и LPS, и, кроме того, показал достоверное
Следует, однако, отметить, что для оценки
повышение экспрессии гена Il1b и после
достоверности различий по результатам секве-
MCAO-MK, выявленное по результатам секве-
нирования был использован достаточно стро-
нирования только на уровне p-value = 0,0021
гий критерий - различия считались досто-
(padj > 1). К списку провоспалительных мар-
верными лишь при padj < 0,05. Такой строгий
керов был также добавлен ген Mmp9, экспрес-
отбор может, как отмечается большинством
сия которого была также значительно увеличе-
исследователей, приводить к потере генов,
на через сутки после MCAO-ML и LPS.
также реально изменивших экспрессию, но
Маркеры глутаматергической системы.
не на требуемом высоком уровне значимо-
На рис. 3 представлено изменение экспрессий
сти. Из рис. 1 видно, что такими генами для
генов-маркеров глутаматергической системы,
MCAO-ML могут быть гены Fas, Casp1 и Casp8,
транспортеров и рецепторов: на рис. 3, а -
которые значительно изменили экспрес-
по результатам секвенирования, на рис. 3, б -
сию по сравнению с соответствующими кон-
по результатам ПЦР-анализа.
трольными значениями, но только на уровне
Экспрессия генов ни одного из пяти из-
p-value < 0,05, в то время как значение padj не
вестных в настоящее время переносчиков
достигало достоверности.
глутамата через клеточную мембрану не из-
Гены, ассоциированные с нейровоспалением.
менилась в гиппокампе через сутки после
Через сутки после обоих ишемических воздей-
MCAO-MK или MCAO-ML. Прямая провос-
ствий в гиппокампе была достоверно увеличе-
палительная стимуляция центральным введе-
на экспрессия гена Gfap, кодирующего маркер-
нием LPS привела к достоверному увеличению
ный белок астроцитов (рис. 2, а). Экспрессия
экспрессии гена одного из таких переносчи-
Aif (гена маркерного белка микроглии Iba-1)
ков - Slc1a2 (рис. 3, а), этот эффект был под-
была увеличена через сутки только после пря-
твержден ПЦР-анализом (p <0,05; рис. 3, б).
мого провоспалительного воздействия - цен-
Ишемические воздействия не повлияли и
трального введения LPS. По результатам сек-
на экспрессию в гиппокампе генов везикуляр-
венирования, экспрессия ни одного из генов
ных транспортеров глутамата, в то время как
ключевых провоспалительных цитокинов не
после LPS было обнаружено достоверное сни-
была достоверно изменена после MCAO-MK,
жение экспрессии гена Slc17a6.
в то время как оба гена (Il1b и Il6) после
Экспрессии генов двух транспортеров
MCAO-ML и один ген (Il1b) после LPS до-
(Slc1a4 и Slc1a5) нейтральных аминокислот, в
стоверно увеличили экспрессию через сут-
их числе также глутамина и глутамата, были до-
ки после воздействия. Анализ экспрессии
стоверно увеличены после MCAO-MK и LPS.
мРНК Il1b методом ПЦР в реальном времени
После MCAO-ML достоверно была увеличена
Рис. 2. Изменение экспрессии генов, ассоциированных с нейровоспалением, после двух типов ишемического воздей-
ствия и центрального введения LPS. а - По результатам секвенирования; * padj < 0,05; ** padj < 0,01; *** padj < 0,001.
б - Верификация ПЦР в реальном времени; * p < 0,05
БИОХИМИЯ том 88 вып. 4 2023
ГЕНЫ ГЛУТАМАТ- И ГАМКЕРГИЧЕСКОЙ СИСТЕМ ПОСЛЕ MCAO И LPS
661
Рис. 3. Изменение экспрессии генов-маркеров глутаматергической системы после двух типов ишемического воздей-
ствия и центрального введения LPS. а - По результатам секвенирования; * padj < 0,05; ** padj < 0,01; *** padj < 0,001;
# padj < 0,1. б - Верификация ПЦР в реальном времени; * p < 0,05
экспрессия только Slc1a5. После MCAO-MK
нальные свойства рецептора. Метаботропные
экспрессия еще одного транспортера нейтраль-
рецепторы глутамата, каждый из которых ко-
ных аминокислот (Slc38a5) была снижена. Эти
дируется отдельным геном, относятся к супер-
результаты указывают на изменение обмена
семейству рецепторов, сопряженных с гуанин-
глутамина и глутамата в гиппокампе после как
связывающими белками - GPCR.
ишемических, так и прямых провоспалитель-
После MCAO-MK обнаружено изменение
ных воздействий. На такую возможность ука-
(снижение) на уровне тенденции (p-value =
зывает увеличение после MCAO-MK экспрес-
= 0,0023; padj = 0,0679) экспрессии гена только
сии гена Glul (катализирует синтез глутамина
одного рецептора - метаботропного Grm3.
из глутамата), подтвержденное ПЦР в реаль-
MCAO-ML привела через сутки к изме-
ном времени (p < 0,05).
нениям в экспрессии как ионотропных, так и
Рецепторы глутамата представлены у мле-
метаботропных рецепторов. Среди субъеди-
копитающих ионотропными и метаботропны-
ниц ионотропных AMPA-рецепторов досто-
ми типами. Ионотропные рецепторы (NMDA,
верное снижение по результатам секвенирова-
AMPA и каинатные) являются гетеротетра-
ния обнаружено для гена Gria4. Снижение, по
мерными трансмембранными каналами, про-
результатам секвенирования, только на уров-
пускающими в клетку при активации нейро-
не тенденции (p-value = 0,0272; padj = 0,1437)
трансмиттером положительно заряженные
экспрессии гена другой субъединицы AMPA-
ионы кальция или натрия. Каждая субъедини-
рецепторов - Gria2, показало достоверный эф-
ца кодируется отдельным геном, и субъединич-
фект (p < 0,05) при определении методом ПЦР
ный состав тетрамера определяет функцио-
в реальном времени. В отличие от снизивших
БИОХИМИЯ том 88 вып. 4 2023
662
КАЛИНИНА и др.
Рис. 4. Изменение экспрессии генов-маркеров ГАМКергической системы после двух типов ишемического воздей-
ствия и центрального введения LPS. а - По результатам секвенирования; * padj < 0,05; ** padj < 0,01; *** padj < 0,001;
# padj < 0,1. б - Верификация ПЦР в реальном времени; * p < 0,05
экспрессию генов, кодирующих субъединицы
Кальций/кальмодулин-зависимые протеин-
AMPA-рецепторов, экспрессия гена субъеди-
киназы. Важную роль в активности глутамат-
ницы Grin3b ионотропных NMDA-рецепторов
ергических и ГАМКергических рецепторов
была достоверно повышена. Помимо ионо-
играют Са2+/кальмодулин-зависимые киназы
тропных AMPA-рецепторов, после MCAO-ML
или СаМ-киназы. По результатам секвени-
обнаружено также достоверное снижение экс-
рования выявлено снижение экспрессии ге-
прессии гена метаботропного рецептора Grm6.
нов Camkk1, Camkk2, Calm1 и Camk4 - после
Через сутки после введения LPS, так же,
MCAO-ML (рис. 5). После MCAO-MK обна-
как и после MCAO-ML, была снижена, но
ружено снижение экспрессии только одного
только на уровне тенденции (p-value = 0,0022;
из этих генов - Camkk1, а после LPS не была
padj = 0,0531) экспрессия гена Gria4, кодирую-
изменена экспрессия ни одного из этих генов.
щего субъединицу AMPA-рецепторов. Также
только на уровне тенденции (p-value = 0,0033;
padj = 0,0734) после введения LPS была сни-
жена экспрессия гена Grik3, кодирующего
субъединицу каинатных рецепторов. После
введения LPS снижение экспрессии гена мета-
ботропного рецептора Grm2 достигало досто-
верного уровня.
Маркеры ГАМКергической нейротрансмис-
сии. Гены ГАМКергической системы, изме-
нившие экспрессии после воздействий, пред-
ставлены на рис. 4, а. После всех воздействий в
гиппокампе снизилась экспрессия гена Slc6a11:
на уровне тенденции (p-value = 0,0038; padj =
= 0,0932) после MCAO-MK и достоверно -
после MCAO-ML и LPS. Экспрессия гена дру-
гого транспортера ГАМК - Slc6a13, а также
гена фермента синтеза ГАМК Gad2 (p-value =
= 0,0041; padj = 0,0871) снизились только после
введения LPS. Верификация изменения Gad2,
подтвержденная ПЦР в реальном времени,
показала достоверность снижения после LPS
Рис. 5. Изменение экспрессии генов кальмодулина и
кальмодулин-зависимых протеинкиназ после двух ти-
(рис. 4, б). После MCAO-ML достоверно снизи-
пов ишемического воздействия и центрального введе-
лись экспрессии генов ГАМКергических рецеп-
ния LPS (по результатам секвенирования). * padj < 0,05;
торов - Gabra5 и Gabrd (рис. 4, а).
** padj < 0,01; # padj < 0,1
БИОХИМИЯ том 88 вып. 4 2023
ГЕНЫ ГЛУТАМАТ- И ГАМКЕРГИЧЕСКОЙ СИСТЕМ ПОСЛЕ MCAO И LPS
663
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
спективном исследовании нам удалось пока-
зать независимость развития постинсультных
Ранее мы предположили, что фокальные
кортизол-зависимых когнитивных и депрес-
ишемические поражения мозга могут вызы-
сивных расстройств от неврологических дефи-
вать вторичные, дистантные (расположенные
цитов (косвенно свидетельствующих о разме-
на расстоянии от первичного очага) поврежде-
рах ишемического очага), которые отражают
ния гиппокампа, реализуемые иными механиз-
развитие первичного ишемического очага в
мами. По-видимому, отложенные последствия
бассейне средней мозговой артерии [30].
фокального повреждения мозга, выражающие-
В данной работе уже через сутки после
ся клинически в когнитивных и эмоциональ-
MCAO-ML обнаружены признаки активации
ных расстройствах, независимо от размера и
апоптоза клеток, на что указывало увеличение
локализации первичного повреждения связаны
экспрессии гена исполнительной протеазы
с опосредованными глюкокортикоидами и ас-
апоптоза каспазы-3. Этот эффект согласу-
социированными с нейровоспалением измене-
ется с результатами предшествующих работ о
ниями в гиппокампе - структуре, селективно
появлении первых признаков гибели клеток
чувствительной к различным экстремальным
в гиппокампе в первые дни после ишемии/
воздействиям [26, 27]. Действительно, гиппо-
реперфузии [19, 31, 32]. Гибель нейрональных
камп, несмотря на отсутствие в нем зоны ин-
клеток может вызывать воспалительный про-
фаркта, относят к структурам мозга, наиболее
цесс, однако индуцированное воспаление, в
уязвимым в постишемический период [3].
свою очередь, может приводить к дальнейшей
МСАО-ML и МСАО-МК - 2 популяр-
гибели клеток [33]. Поэтому не удивительно,
ные и наиболее часто применяемые в пред-
что прямое воздействие провоспалительным
клинических исследованиях модели инсуль-
стимулом - бактериальным эндотоксином -
та. При этом метаанализ немногочисленных
приводило через сутки к увеличению в гип-
сравнительных исследований этих моделей на
покампе не только экспрессии гена Aif - гена
грызунах показывает, что данные о сходстве
маркерного белка активированной микро-
и различиях этих моделей противоречивы,
глии Iba-1, но и гена рецептора смерти Fas.
зависят от используемого вида (мыши, кры-
Нейровоспаление проявляется как акти-
сы), конкретного времени ишемии и наличия
вацией глиальных клеток, так и увеличением
или отсутствия реперфузии [28]. Это касается
продукции провоспалительных цитокинов.
данных об объеме инфаркта, смертности и
Наиболее быструю активацию после ишемии
неврологическом дефиците. В наших преды-
отмечают у астроцитов. Двухчасовая MCAO
дущих исследованиях на этих моделях на кры-
сопровождалась через сутки значительным
сах не удалось найти достоверные различия
увеличением числа активированных астроци-
в объеме инфаркта мозга, оцененном через
тов в гиппокампе крыс, в то время как замет-
72 ч после МСАО с использованием стан-
ной активации микроглиальных клеток в этот
дартного метода окраски митохондриальным
период не наблюдалось [34]. С этими резуль-
красителем
2,3,5-трифенил тетразолий хло-
татами полностью согласуются наши данные
ридом (ТТС) [20]. Не обнаружено также досто-
об увеличении в гиппокампе экспрессии гена
верных различий в развитии неврологических
маркерного белка астроцитов Gfap и отсутствие
дефицитов в первые 14 дней после МСАО и в
изменений в экспрессии гена Aif через 24 ч
выживаемости животных [20, 23]. Отсутствие
после обоих ишемических воздействий. Ише-
различий в неврологических дефицитах ме-
мия/реперфузия повышает проницаемость ге-
жду двумя моделями косвенно подтверждает
матоэнцефалического барьера, благодаря чему
и сходный объем инфаркта мозга, поскольку
к секреции провоспалительных цитокинов
между этими показателями существует пря-
активированными глиальными клетками под-
мая зависимость [29]. Несмотря на то что при
ключаются также мигрировавшие из перифе-
моделировании МСАО-ML и МСАО-МК на
рической крови лимфоциты [35]. Интересно,
крысах ишемический очаг ограничивается
что после MCAO по методу Лонга увеличению
областями неокортекса и стриатума [20], экс-
экспрессии гена матриксной металлопротеи-
периментально было показано, что имеются
назы-9 (MMP9), вовлекаемой в нарушение
достоверные различия связывания кортико-
проницаемости барьера
[36], сопутствовала
стерона в гиппокампе и фронтальной коре
более выраженная у этой модели провоспали-
(вне зоны первичного ишемического повреж-
тельная активация. По результатам секвени-
дения) не только в остром периоде [20], но и
рования, после MCAO-ML были достоверно
даже через 3 месяца после ишемии МСАО [23].
увеличены экспрессии генов двух ключевых
Важно отметить, что и в клиническом про-
провоспалительных цитокинов - Il1b и Il6,
БИОХИМИЯ том 88 вып. 4 2023
664
КАЛИНИНА и др.
в то время как достоверное увеличение экс-
мина и глутамата в гиппокампе после ишемии.
прессии лишь одного из них, Il1b, было выяв-
Увеличение экспрессии гена Slc1a5 согласует-
лено после MCAO-MK лишь ПЦР-анализом.
ся с обнаруженными нами ранее изменениями
Эти данные в значительной степени подтвер-
после LPS и инсульта по методу Коизуми [16]
ждают результаты работы Smith et al. [18],
и расширяет этот эффект ишемии и для мо-
которые свидетельствуют о достоверно бо-
дели Лонга. Предположение о связи повы-
лее выраженных провоспалительных про-
шенной экспрессии транспортеров нейтраль-
цессах у мышей с MCAO-ML в сравнении с
ных аминокислот с возможным изменением
MCAO-MK.
обмена глутамина и глутамата косвенно под-
Значительную роль в нейродегенеративных
тверждается увеличением экспрессии гена глу-
эффектах ишемии отводят глутаматергиче-
таминсинтетазы Glul после MCAO-MK, харак-
ской эксайтотоксичности, которую связывают
теризующейся изменением экспрессии обоих
с накоплением глутамата в экстраклеточном
транспортеров. Прямая провоспалительная
пространстве и гиперактивацией глутаматных
стимуляция центральным введением LPS при-
рецепторов [5, 37, 38], логически обусловив-
вела к достоверному увеличению экспрессии
ших представление, что ослабление этих собы-
гена переносчика глутамата через клеточную
тий является благоприятным для смягчения
мембрану Slc1a2 и снижению экспрессии гена
клеточной гибели [39]. Действительно, экспе-
везикулярного транспортера Slc17a6. Экспрес-
риментальная индукция ишемии с помощью
сии генов транспортеров этих типов не были
широко используемого метода, в том числе и
изменены через сутки после обеих ишемий.
в нашей работе - окклюзии средней мозговой
Действие глутамата осуществляется через
артерии - буквально с первых минут вызывает
ионотропные и метаботропные глутаматные
быстрое увеличение уровня внеклеточного глу-
рецепторы, экспрессии генов которых изменя-
тамата в гиппокампе [40, 41]. Это острое увели-
лись через сутки после примененных нами воз-
чение уровня внеклеточного глутамата в струк-
действий. Более выраженные изменения на-
туре, однако, также быстро нормализуется,
блюдались после MCAO-ML и LPS, которые,
что, очевидно, обеспечивается работой специ-
как показано выше, приводили к большему
фических переносчиков нейромедиатора через
провоспалительному ответу, чем MCAO-MK,
клеточную мембрану. В настоящее время опи-
что, учитывая локализацию глутаматных ре-
сано 5 типов таких переносчиков [42], которые
цепторов на глиальных клетках [44], являются
переносят высвободившийся глутамат в астро-
дополнительным свидетельством взаимодей-
циты, где он под действием глутаматсинтазы
ствия ответов глутаматергической и провос-
превращается в глутамин. Далее белки-пере-
палительной систем. После MCAO-ML и LPS,
носчики глутамина на плазматической мем-
но не после MCAO-MK в гиппокампе наблю-
бране астроцитов и нейронов опосредуют пе-
далось снижение экспрессии гена Gria4, коди-
ренос глутамина от астроцитов к нейронам, где
рующего субъединицу AMPA-рецепторов. Эти
под действием глутаминазы глутамин превра-
результаты согласуются со снижением экспрес-
щается в глутамат, который с помощью вези-
сии белка AMPA-рецепторов в гиппокампе
кулярных транспортеров глутамата загружается
крыс, обнаруженным через сутки после перма-
в синаптические везикулы для дальнейшего
нентной окклюзии средней мозговой артерии,
использования. Полагают, что более половины
обычной или отягощенной диабетическим
глутамата образуется в результате такого глу-
состоянием [19]. В этой работе было также
тамато-глутаминового цикла между нейрона-
обнаружено различие в ответах глутаматных
ми и астроцитами [43]. Несмотря на быструю
рецепторов на тип ишемического воздей-
нормализацию острого повышения уровня
ствия - изменений в экспрессии белка AMPA-
экстраклеточного глутамата, выяснилось, что
рецепторов в гиппокампе крыс не было выяв-
ишемия мозга может вызывать длительные
лено через 24 ч после транзиторной ишемии.
изменения в регуляторах глутаматергической
После MCAO-ML обнаружено увеличение
нейротрансмиссии, что, в свою очередь, может
экспрессии гена Grin3b, кодирующего субъ-
обусловливать отсроченные изменения как в
единицу NMDA-рецепторов, изменения кото-
активности нейротрансмиссии, так и в связан-
рого не обнаружено ни после MCAO-MK, ни
ных с нею функциях.
после LPS. Обнаруженное увеличение может от-
Примененные нами ишемические воздей-
ражать либо развитие у животных посттравма-
ствия приводили через сутки к повышению в
тического стрессового расстройства [45], либо,
гиппокампе экспрессии генов транспортеров
напротив, более раннюю после MCAO-ML
нейтральных аминокислот, Slc1a4 и Slc1a5, что
активацию восстанавливающих процессов.
может указывать на изменение обмена глута-
Относительно второй возможности следует
БИОХИМИЯ том 88 вып. 4 2023
ГЕНЫ ГЛУТАМАТ- И ГАМКЕРГИЧЕСКОЙ СИСТЕМ ПОСЛЕ MCAO И LPS
665
отметить, что эксайтотоксическую гибель ней-
после MCAO-ML были также снижены экс-
ронов после ишемического повреждения свя-
прессии генов ГАМКергических рецепто-
зывают, прежде всего, с гиперактивацией
ров - Gabra5 и Gabrd.
NMDA-рецепторов, однако клинические испы-
Хотя проверка в качестве нейрозащитных
тания антагонистов этих рецепторов не только
агентов препаратов, нацеленных на NMDA-
не улучшали, но даже усугубляли повреждение
рецепторы, потерпела в клинических испыта-
головного мозга [10]. В отличие от антагони-
ниях неудачу, имеются отдельные свидетель-
стов, агонисты в острый период после ин-
ства смягчения вызываемых ими негативных
сульта через модулирующий «сайт глицина»
побочных эффектов через влияние на сиг-
NMDA оказывались более «полезными» для
нальный каскад ниже стимуляции рецепто-
восстановления
[11]. мРНК Grin3b широко
ров [52]. Перспективным может быть также
распространена в мозге взрослых крыс [46], а
путь воздействия на активность ГАМКергиче-
комбинация субъединиц NR1/NR3B (Grin1/
ских рецепторов через сигнальные пути рецеп-
Grin3b) может представлять собой тип воз-
торов и регуляторы синаптической пластич-
буждающего глицинового рецептора [47]. Од-
ности, к которым принадлежат, в частности,
нако прояснение роли субъединицы Grin3b
белковые продукты генов Camkk1 и Camkk2,
NMDA-рецепторов в постишемический пери-
изменивших экспрессию после MCAO-ML.
од требует специальных исследований.
В недавней работе было показано, что восста-
Все три примененных в работе воздей-
новление сниженной ишемией экспрессии ре-
ствия приводили к снижению экспрессии генов
цепторов ГАМК путем блокады механизма их
метаботропных рецепторов, но если после
взаимодействия с CaMKII, фосфорилируемой
MCAO-MK это был ген Grm3, то после MCAO-
Camkk1 и Camkk2, ингибировало эксайтоток-
ML - Grm6, а после LPS - Grm2. Grm3 и Grm2
сическую гибель нейронов [53], что, по мне-
относятся ко второй группе метаботропных
нию авторов, может значительно расширить
рецепторов, в то время как Grm6 - к треть-
фармакологические возможности торможе-
ей. Стимуляция метаботропных рецепторов
ния прогрессирующей гибели нейронов после
третьей группы, экспрессирующихся на мик-
ишемического инсульта.
роглиальных клетках, трансформирует мик-
роглию в нейропротективный фенотип, в то
время как стимуляция рецепторов второй
ЗАКЛЮЧЕНИЕ
группы, особенно mGluR2 (Grm2) - в нейро-
токсический фенотип, проявляющийся выс-
В целом, полученные результаты пока-
вобождением Fas-лиганда и запуском апоп-
зывают как различие, так и сходство между
тоза через активацию каспазы-3 [45]. После
ответами на примененные в работе воздей-
MCAO мыши-нокауты по Grm2 демонстри-
ствия (таблица). Животные после MCAO-ML
ровали меньший объем поражения и ускоре-
демонстрировали большее число изменивших
ние поведенческого восстановления [48]. Раз-
экспрессию генов, связанных с активацией
личные эффекты MCAO-MK и MCAO-ML на
апоптоза и нейровоспаления, рецепцией глу-
экспрессию компонентов глутаматергической
тамата, а также маркерами ГАМКергической
системы могут быть ассоциированы с раз-
системы, чем MCAO-MK, и были ближе в
личной аккумуляцией кортикостерона в гип-
этом плане к ответам, индуцированным цен-
покампе в этих моделях [20, 23], потому что
тральным введением LPS. В свою очередь, по
глюкокортикоидные гормоны являются регу-
сравнению с MCAO-ML, MCAO-MK и LPS
ляторами практически всех компонентов этой
характеризовались изменением большего
системы [49], а также контролируют развитие
числа генов, вовлекаемых в транспорт глута-
гиперглутаматергической трансмиссии [50], ре-
мата. Наиболее ярко проявившимся отличием
зультатом которой является развитие эксайто-
MCAO-ML от MCAO-MK и введения LPS яви-
токсичности.
лись обнаруженные после этого типа ишеми-
Помимо усиления глутаматергического
ческого воздействия изменения в экспрессии
сигнала, нарушение угнетающего влияния
генов кальмодулина и кальмодулин-зависимых
ГАМК на возбудимость нейронов также вно-
киназ. Выявленные особенности ответов тран-
сит вклад в ухудшение состояния после ин-
скриптома гиппокампа на две модели MCAO и
сульта [51]. Примененные в работе воздействия
прямой провоспалительный стимул будут спо-
повлияли на экспрессию маркеров ГАМК-
собствовать дальнейшему пониманию причин
ергической системы. Общим ответом на все
и молекулярных механизмов, лежащих в осно-
воздействия было снижение экспрессии гена
ве разнообразия последствий инсультов, как в
транспортера ГАМК Slc6a11. Кроме того,
модельных исследованиях, так и в клинике.
БИОХИМИЯ том 88 вып. 4 2023
666
КАЛИНИНА и др.
Изменение экспрессии генов в гиппокампе после окклюзии средней мозговой артерии или центрального введения
липополисахарида по скорректированному p-значению (padj)
Процесс
Ген
MCAO-MK
MCAO-ML
LPS
Fas
≈
≈
↑**
Casp1
≈
≈
↑**
Casp3
≈
↑***
≈
Casp4
↑***
↑***
↑**
Casp8
≈
≈
↑#
Gfap
↑***
↑***
↑**
Aif (IBA1)
≈
≈
↑**
Il1b
≈
↑*
↑**
Il6
≈
↑***
≈
MMP9
≈
↑***
↑**
Slc1a2
≈
≈
↑**
Slc17a6
≈
≈
↓**
Slc1a4
↑***
≈
↑**
Slc1a5
↑**
↑***
↑**
Slc38a5
↓**
≈
≈
Glul
↑#
≈
≈
Grin3b
≈
↑***
≈
Gria2
≈
↓#
≈
Gria4
≈
↓*
↓#
Grik3
≈
≈
↓#
Grm2
≈
≈
↓**
Grm3
↓#
≈
≈
Grm6
≈
↓*
≈
Gad2
≈
≈
↓#
Gabra5
≈
↓*
≈
Gabrd
≈
↓*
≈
Slc6a11
↓#
↓***
↓**
Slc6a13
≈
≈
↓*
Calm1
≈
↓*
≈
Camk4
≈
↓#
≈
Camkk1
≈
↓*
≈
Camkk2
↓*
↓**
≈
Примечание. Обозначения: ↑↓ - достоверное увеличение или снижение; ≈ - отсутствие достоверных изменений;
* padj < 0,05; ** padj < 0,01; *** padj < 0,001; # padj < 0,1; серым цветом выделены гены, экспрессия которых верифициро-
вана методом ПЦР в реальном времени.
БИОХИМИЯ том 88 вып. 4 2023
ГЕНЫ ГЛУТАМАТ- И ГАМКЕРГИЧЕСКОЙ СИСТЕМ ПОСЛЕ MCAO И LPS
667
Ограничения работы. Основной задачей
писание текста; Н.В. Гуляева, Т.С. Калинина
данной работы было исследование раннего
и Н.Н. Дыгало - редактирование текста статьи.
транскрипционного ответа гиппокампа на
Финансирование. Работа выполнена при
окклюзию средней мозговой артерии при ис-
финансовой поддержке Российского научного
пользовании двух моделей МСАО; для этого
фонда (грант № 20-64-47013), а также направ-
производили выделение гиппокампа через 24 ч
ленного исключительно на содержание и уход
после ишемии или ложной операции. Дизайн
экспериментальных животных бюджетного
эксперимента не позволял провести прямую
проекта № FWNR-2022-0023.
оценку ишемического очага у эксперимен-
Благодарности. Благодарим ООО
«Гено-
тальных животных и межгрупповое сравнение,
аналитика» (Москва, Россия) за секвениро-
однако результаты физиологического тестиро-
вание образцов и первичный биоинформати-
вания, представленные в работе, позволяют,
ческий анализ. Благодарим В.Н. Бабенко за
хотя и косвенно, но достаточно адекватно оце-
первичный биоинформатический анализ дан-
нить наличие очагов по развитию неврологи-
ных после введения липополисахарида.
ческого дефицита. Данные использованных
Конфликт интересов. Авторы заявляют об
методов оценки неврологического дефицита,
отсутствии конфликта интересов.
как было показано ранее, коррелируют с объе-
Соблюдение этических норм. Все проводи-
мом ишемического очага [29].
мые исследования выполнялись в соответствии
с Международными стандартами по работе с
Вклад авторов. Н.В. Гуляева и Г.Т. Шиш-
лабораторными животными, согласно Дирек-
кина - концепция и руководство работой;
тиве 86/609/EEC и рекомендациям Комиссии
Т.С. Калинина, Д.А. Ланшаков, Е.В. Сухарева,
по биоэтике ФИЦ Институт цитологии и гене-
М.В. Онуфриев и Ю.В. Моисеева - проведение
тики СО РАН (Протокол № 115 от 20.12.2021 г.),
экспериментов; Н.В. Гуляева, Г.Т. Шишкина,
а также по протоколам, утвержденным Комис-
Т.С. Калинина, Н.Н. Дыгало - обсуждение ре-
сией по биоэтике Института высшей нервной
зультатов исследования; Г.Т. Шишкина - на-
деятельности и нейрофизиологии РАН.
СПИСОК ЛИТЕРАТУРЫ
1. Rudolph, M., Schmeer, C. W., Günther, M.,
6. Luo, Y., Ma, H., Zhou, J. J., Li, L., Chen, S. R.,
Woitke, F., Kathner-Schaffert, C., Karapetow, L.,
Zhang, J., Chen, L., and Pan, H. L. (2018) Focal ce-
Lindner, J., Lehmann, T., Jirikowski, G., Witte, O. W.,
rebral ischemia and reperfusion induce brain injury
Redecker, C., and Keiner, S.
(2021) Microglia-
through α2δ-1-bound NMDA receptors, Stroke, 49,
mediated phagocytosis of apoptotic nuclei is impaired
2464-2472, doi: 10.1161/STROKEAHA.118.022330.
in the adult murine hippocampus after stroke, Glia,
7. Magi, S., Piccirillo, S., and Amoroso, S.
(2019)
69, 2006-2022, doi: 10.1002/glia.24009.
The dual face of glutamate: from a neurotoxin to a
2. Rolls, E. T. (1996) A theory of hippocampal function
potential survival factor-metabolic implications in
in memory, Hippocampus, 6, 601-620, doi: 10.1002/
health and disease, Cell. Mol. Life Sci., 76, 1473-1488,
(SICI)1098-1063(1996)6:6<601::AID-HIPO5>
doi: 10.1007/s00018-018-3002-x.
3.0.CO;2-J.
8. Deisseroth, K., Singla, S., Toda, H., Monje, M.,
3. Gulyaeva, N. V., Onufriev, M. V., and Moiseeva, Y. V.
Palmer, T. D., and Malenka, R. C. (2004) Excitation-
(2021) Ischemic stroke, glucocorticoids, and remote
neurogenesis coupling in adult neural stem/progenitor
hippocampal damage: a translational outlook and
cells, Neuron, 42, 535-552, doi: 10.1016/s0896-6273
implications for modeling, Front. Neurosci.,
15,
(04)00266-1.
781964, doi: 10.3389/fnins.2021.781964.
9. Hu, J., Li, C., Hua, Y., Liu, P., Gao, B., Wang, Y., and
4. Robinson, R. G., and Jorge, R. E. (2016) Post-stroke
Bai, Y. (2020) Constraint-induced movement therapy
depression: a review, Am. J. Psychiatry, 173, 221-231,
improves functional recovery after ischemic stroke
doi: 10.1176/appi.ajp.2015.15030363.
and its impacts on synaptic plasticity in sensorimotor
5. Globus, M. Y., Busto, R., Martinez, E., Valdes, I.,
cortex and hippocampus, Brain Res. Bull., 160, 8-23,
Dietrich, W. D., and Ginsberg, M. D. (1991) Compar-
doi: 10.1016/j.brainresbull.2020.04.006.
ative effect of transient global ischemia on extracellular
10. Ikonomidou, C., and Turski, L.
(2002) Why
levels of glutamate, glycine, and gamma-aminobutyr-
did NMDA receptor antagonists fail clinical tri-
ic acid in vulnerable and nonvulnerable brain regions
als for stroke and traumatic brain injury? Lan-
in the rat, J. Neurochem., 57, 470-478, doi: 10.1111/
cet Neurol.,
1,
383-386, doi:
10.1016/s1474-4422
j.1471-4159.1991.tb03775.x.
(02)00164-3.
БИОХИМИЯ том 88 вып. 4 2023
668
КАЛИНИНА и др.
11.
Biegon, A., Liraz-Zaltsman, S., and Shohami, E.
in the hippocampus and frontal cortex, Int. J. Mol.
(2018) Stimulation of N-methyl-D-aspartate receptors
Sci., 22, 13544, doi: 10.3390/ijms222413544.
by exogenous and endogenous ligands improves
21.
Koizumi, J.Y., Nakazawa, T., and Ooneda, G. (1986)
outcome of brain injury, Curr. Opin. Neurol., 31, 687-
Experimental studies of ischemic cerebral edema. I.
692, doi: 10.1097/WCO.0000000000000612.
A new experimental model of cerebral embolism in
12.
Shishkina, G. T., Kalinina, T. S., Gulyaeva, N. V.,
rats in which recirculation in the ischemic area can
Lanshakov, D. A., and Dygalo, N. N. (2021) Changes
be introduced, Jpn. J. Stroke, 8, 1-8, doi: 10.3995/
in gene expression and neuroinflammation in the
jstroke.8.1.
hippocampus after focal brain ischemia: involvement
22.
Longa, E. Z., Weinstein, P. R., Carlson, S., and
in the long-term cognitive and mental disorders,
Cummins, R. (1989) Reversible middle cerebral artery
Biochemistry (Moscow), 86, 657-666, doi: 10.1134/
occlusion without craniectomy in rats, Stroke, 20, 84-
S0006297921060043.
91, doi: 10.1161/01.str.20.1.84.
13.
Batista, C. R. A., Gomes, G. F., Candelario-Jalil, E.,
23.
Onufriev, M. V., Stepanichev, M. Y., Moiseeva, Y. V.,
Fiebich, B. L., and de Oliveira, A. C. P. (2019) Lipo-
Zhanina, M. Y., Nedogreeva, O. A., Kostryukov,
polysaccharide-induced neuroinflammation as a
P. A., Lazareva, N. A., and Gulyaeva, N. V. (2022)
bridge to understand neurodegeneration, Int. J. Mol.
A comparative study of two models of intraluminal
Sci., 20, 2293, doi: 10.3390/ijms20092293.
filament middle cerebral artery occlusion in rats:
14.
Chung, J. Y., Yi, J. W., Kim, S. M., Lim, Y. J., Chung,
long lasting accumulation of corticosterone and
J. H., and Jo, D. J. (2011) Changes in gene expression
interleukins in the hippocampus and frontal cortex in
in the rat hippocampus after focal cerebral ischemia,
Koizumi model, Biomedicines, 10, 3119, doi: 10.3390/
J. Korean Neurosurg. Soc., 50, 173-178, doi: 10.3340/
biomedicines10123119.
jkns.2011.50.3.173.
24.
Arvidsson, A., Kokaia, Z., and Lindvall, O. (2001)
15.
Wang, C., Liu, M., Pan, Y., Bai, B., and Chen, J.
N-methyl-D-aspartate receptor-mediated increase of
(2017) Global gene expression profile of cerebral
neurogenesis in adult rat dentate gyrus following stroke,
ischemia-reperfusion injury in rat MCAO mod-
Eur. J. Neurosci., 14, 10-18, doi: 10.1046/j.0953-
el, Oncotarget,
8,
74607-74622, doi:
10.18632/
816x.2001.01611.x.
oncotarget.20253.
25.
Dygalo, N. N., Bannova, A. V., Kalinina, T. S., and
16.
Shishkina, G. T., Gulyaeva, N. V., Lanshakov, D. A.,
Shishkina, G. T. (2004) Clonidine increases caspase-3
Kalinina, T. S., Onufriev, M. V., Moiseeva, Y. V.,
mRNA level and DNA fragmentation in the devel-
Sukhareva, E. V., and Babenko, V. N. (2021) Identify-
oping rat brainstem, Dev. Brain Res., 152, 225-231,
ing the involvement of pro-inflammatory signal in hip-
doi: 10.1016/j.devbrainres.2004.06.018.
pocampal gene expression changes after experimental
26.
Gulyaeva, N. V. (2019) Biochemical mechanisms and
ischemia: transcriptome-wide analysis, Biomedicines,
translational relevance of hippocampal vulnerability to
9, 1840, doi: 10.3390/biomedicines9121840.
distant focal brain injury: the price of stress response,
17.
Bonow, R. H., Aïd, S., Zhang, Y., Becker, K. G.,
Biochemistry (Moscow), 84, 1306-1328, doi: 10.1134/
and Bosetti, F. (2009) The brain expression of genes
S0006297919110087.
involved in inflammatory response, the ribosome, and
27.
Gulyaeva, N. V. (2019) Functional neurochemistry
learning and memory is altered by centrally injected
of the ventral and dorsal hippocampus: stress, de-
lipopolysaccharide in mice, Pharmacogenomics J., 9,
pression, dementia and remote hippocampal dam-
116-126, doi: 10.1038/tpj.2008.15.
age, Neurochem. Res., 44, 1306-1322, doi: 10.1007/
18.
Smith, H. K., Russell, J. M., Granger, D. N., and
s11064-018-2662-0.
Gavins, F. N. (2015) Critical differences between two
28.
Li, Y., Tan, L., Yang, C., He, L., Deng, B., Huang, X.,
classical surgical approaches for middle cerebral artery
Liu, S., Liu, L., Wang, J., and Guo, J.
(2022)
occlusion-induced stroke in mice, J. Neurosci. Meth-
Comparison of middle cerebral artery occlusion
ods, 249, 99-105, doi: 10.1016/j.jneumeth.2015.04.008.
models conducted by Koizumi and Longa methods:
19.
Shah, F. A., Li, T., Kury, L. T. A., Zeb, A., Khatoon, S.,
a systematic review and meta-analysis of rodent
Liu, G., Yang, X., Liu, F., Yao, H., Khan, A.-U.,
data
[Preprint], Research Square, doi:
10.21203/
Koh, P. O., Jiang, Y., and Li, S. (2019) Pathological
rs.3.rs-2398116/v1.
comparisons of the hippocampal changes in the tran-
29.
Gulyaeva, N., Thompson, C., Shinohara, N.,
sient and permanent middle cerebral artery occlusion
Lazareva, N., Onufriev, M., Stepanichev, M.,
rat models, Front. Neurol., 10, 1178, doi: 10.3389/
Moiseeva, Y., Fliss, H., and Hakim, A. M. (2003)
fneur.2019.01178.
Tongue protrusion: a simple test for neurological
20.
Onufriev, M. V., Moiseeva, Y. V., Zhanina, M. Y.,
recovery in rats following focal cerebral ischemia,
Lazareva, N. A., and Gulyaeva, N. V. (2021) A com-
J. Neurosci. Methods, 125, 183-193, doi: 10.1016/
parative study of Koizumi and Longa methods of in-
s0165-0270(03)00056-6.
traluminal filament middle cerebral artery occlusion in
30.
Zhanina, M. Y., Druzhkova, T. A., Yakovlev, A. A.,
rats: early corticosterone and inflammatory response
Vladimirova, E. E., Freiman, S. V., Eremina, N. N.,
БИОХИМИЯ том 88 вып. 4 2023
ГЕНЫ ГЛУТАМАТ- И ГАМКЕРГИЧЕСКОЙ СИСТЕМ ПОСЛЕ MCAO И LPS
669
Guekht, A. B., and Gulyaeva, N. V. (2022) Devel-
and Pera, J. (2017) Ceftriaxone- and N-acetylcysteine-
opment of post-stroke cognitive and depressive dis-
induced brain tolerance to ischemia: influence on
turbances: associations with neurohumoral indices,
glutamate levels in focal cerebral ischemia, PLoS One,
Curr. Issues Mol. Biol., 44, 6290-6305, doi: 10.3390/
12, e0186243, doi: 10.1371/journal.pone.0186243.
cimb44120429.
42.
Magi, S., Piccirillo, S., Amoroso, S., and Lariccia, V.
31.
States, B. A., Honkaniemi, J., Weinstein, P. R., and
(2019) Excitatory amino acid transporters (EAATs):
Sharp, F. R. (1996) DNA fragmentation and HSP70
glutamate transport and beyond, Int. J. Mol. Sci., 20,
protein induction in hippocampus and cortex occurs in
5674, doi: 10.3390/ijms20225674.
separate neurons following permanent middle cerebral
43.
Jiang, T., Jiao, J., Shang, J., Bi, L., Wang, H., Zhang, C.,
artery occlusions, J. Cereb. Blood Flow Metab., 16,
Wu, H., Cui, Y., Wang, P., and Liu, X.
(2022)
1165-1175, doi: 10.1097/00004647-199611000-00011.
The differences of metabolites in different parts of
32.
Uchida, H., Fujita, Y., Matsueda, M., Umeda, M.,
the brain induced by Shuxuetong Injection against
Matsuda, S., Kato, H., Kasahara, J., Araki, T. (2010)
cerebral ischemia-reperfusion and its corresponding
Damage to neurons and oligodendrocytes in the hip-
mechanism, Evid. Based Complement. Alternat. Med.,
pocampal CA1 sector after transient focal ischemia in
2022, 9465095, doi: 10.1155/2022/9465095.
rats, Cell. Mol. Neurobiol., 30, 1125-1134, doi: 10.1007/
44.
Pocock, J. M., and Kettenmann, H. (2007) Neuro-
s10571-010-9545-5.
transmitter receptors on microglia, Trends Neurosci.,
33.
Ransohoff, R. M. (2016) How neuroinflammation
30, 527-535, doi: 10.1016/j.tins.2007.07.007.
contributes to neurodegeneration, Science, 353, 777-
45.
Lori, A., Schultebraucks, K., Galatzer-Levy, I.,
783, doi: 10.1126/science.aag2590.
Daskalakis, N. P., Katrinli, S., Smith, A. K., Myers,
34.
Xu, A. L., Zheng, G. Y., Ye, H. Y., Chen, X. D.,
A. J., Richholt, R., Huentelman, M., Guffanti, G.,
and Jiang, Q. (2020) Characterization of astrocytes
Wuchty, S., Gould, F., Harvey, P. D., Nemeroff, C. B.,
and microglial cells in the hippocampal CA1 region
Jovanovic, T., Gerasimov, E. S., Maples-Keller, J. L.,
after transient focal cerebral ischemia in rats treated
Stevens, J. S., Michopoulos, V., Rothbaum, B. O.,
with Ilexonin A, Neural Regen. Res., 15, 78-85,
Wingo, A. P., and Ressler, K. J. (2021) Transcriptome-
doi: 10.4103/1673-5374.264465.
wide association study of post-trauma symptom
35.
Rosenberg, G. A. (2009) Matrix metalloproteinases
trajectories identified GRIN3B as a potential biomarker
and their multiple roles in neurodegenerative
for PTSD development, Neuropsychopharmacology,
diseases, Lancet Neurol., 8, 205-216, doi: 10.1016/
46, 1811-1820, doi: 10.1038/s41386-021-01073-8.
S1474-4422(09)70016-X.
46.
Andersson, O., Stenqvist, A., Attersand, A., and
36.
Hannocks, M. J., Zhang, X., Gerwien, H.,
von Euler, G. (2001) Nucleotide sequence, genomic
Chashchina, A., Burmeister, M., Korpos, E., Song, J.,
organization, and chromosomal localization of genes
and Sorokin, L. (2019) The gelatinases, MMP-2
encoding the human NMDA receptor subunits NR3A
and MMP-9, as fine tuners of neuroinflammatory
and NR3B, Genomics, 78, 178-184, doi: 10.1006/
processes, Matrix Biol., 75-76, 102-113, doi: 10.1016/
geno.2001.6666.
j.matbio.2017.11.007.
47.
Chatterton, J. E., Awobuluyi, M., Premkumar, L. S.,
37.
Liu, Y., Wong, T. P., Aarts, M., Rooyakkers, A.,
Takahashi, H., Talantova, M., Shin, Y., Cui, J., Tu, S.,
Liu, L., Lai, T. W., Wu, D. C., Lu, J., Tymianski, M.,
Sevarino, K. A., Nakanishi, N., Tong, G., Lipton, S. A.,
Craig, A. M., and Wang, Y. T. (2007) NMDA receptor
and Zhang, D. (2002) Excitatory glycine receptors
subunits have differential roles in mediating excitotoxic
containing the NR3 family of NMDA receptor sub-
neuronal death both in vitro and in vivo, J. Neurosci., 27,
units, Nature, 415, 793-798, doi: 10.1038/nature715.
2846-2857, doi: 10.1523/JNEUROSCI.0116-07.2007.
48.
Mastroiacovo, F., Moyanova, S., Cannella, M., Ga-
38.
Szydlowska, K., and Tymianski, M. (2010) Calcium,
glione, A., Verhaeghe, R., Bozza, G., Madonna, M.,
ischemia and excitotoxicity, Cell Calcium, 47, 122-129,
Motolese, M., Traficante, A., Riozzi, B., Bruno, V.,
doi: 10.1016/j.ceca.2010.01.003.
Battaglia, G., Lodge, D., and Nicoletti, F.
(2017)
39.
Kalia, L. V., Kalia, S. K., and Salter, M. W. (2008)
Genetic deletion of mGlu2 metabotropic glutamate
NMDA receptors in clinical neurology: excitatory
receptors improves the short-term outcome of ce-
times ahead, Lancet Neurol., 7, 742-755, doi: 10.1016/
rebral transient focal ischemia, Mol. Brain, 10, 39,
S1474-4422(08)70165-0.
doi: 10.1186/s13041-017-0319-6.
40.
Yang, Y., Li, Q., Miyashita, H., Yang, T., and
49.
Gulyaeva, N. V. (2021) Glucocorticoid regulation
Shuaib, A. (2001) Different dynamic patterns of ex-
of the glutamatergic synapse: mechanisms of stress-
tracellular glutamate release in rat hippocampus after
dependent neuroplasticity, J. Evol. Biochem. Physiol.,
permanent or 30-min transient cerebral ischemia and
57, 564-576, doi: 10.1134/S0022093021030091.
histological correlation, Neuropathology, 21, 181-187,
50.
Gulyaeva, N. V. (2022) Neuroendocrine control of
doi: 10.1046/j.1440-1789.2001.00397.x.
hyperglutamatergic states in brain pathologies: the
41.
Krzyżanowska, W., Pomierny, B., Bystrowska, B.,
effects of glucocorticoids, J. Evol. Biochem. Physiol.,
Pomierny-Chamioło, L., Filip, M., Budziszewska, B.,
58, 1425-1438, doi: 10.1134/S0022093022050131.
БИОХИМИЯ том 88 вып. 4 2023
670
КАЛИНИНА и др.
51. Neumann, S., Boothman-Burrell, L., Gowing, E. K.,
cellular events in excitotoxic neuronal death as poten-
Jacobsen, T. A., Ahring, P. K., Young, S. L., Sandager-
tial neuroprotective strategies, Pharmacol. Ther., 160,
Nielsen, K., and Clarkson, A. N. (2019) The delta-
159-179, doi: 10.1016/j.pharmthera.2016.02.009.
subunit selective GABA a receptor modulator, DS2,
53. Balakrishnan, K., Hleihil, M., Bhat, M. A., Ganley,
improves stroke recovery via an anti-inflammatory
R. P., Vaas, M., Klohs, J., Zeilhofer, H. U., and
mechanism, Front. Neurosci., 13, 1133, doi: 10.3389/
Benke, D. (2022) Targeting the interaction of GABAB
fnins.2019.01133.
receptors with CaMKII with an interfering peptide
52. Hoque, A., Hossain, M. I., Ameen, S. S., Ang, C. S.,
restores receptor expression after cerebral ischemia
Williamson, N., Ng, D. C. H., Chueh, A. C., Roul-
and inhibits progressive neuronal death in mouse brain
ston, C., and Cheng, H.-C. (2016) A beacon of hope
cells and slices, Brain Pathol., 33, e13099, doi: 10.1111/
in stroke therapy-Blockade of pathologically activated
bpa.13099.
COMPARATIVE INVESTIGATION OF GLUTAMATE
AND GABA GENE EXPRESSION IN THE HIPPOCAMPUS
AFTER FOCAL BRAIN ISCHEMIA
AND CENTRAL LPS ADMINISTRATION
T. S. Kalinina1, G. T. Shishkina1*, D. A. Lanshakov1, E. V. Sukhareva1, M. V. Onufriev2,
Y. V. Moiseeva2, N. V. Gulyaeva2, and N. N. Dygalo1
1 Institute of Cytology and Genetics, Siberian Branch of Russian Academy of Sciences,
630090 Novosibirsk, Russia; e-mail: gtshi@bionet.nsc.ru
2 Institute of Higher Nervous Activity and Neurophysiology of Russian Academy of Sciences,
117485 Moscow, Russia
Among the responses in the early stages of stroke, activation of neurodegenerative and proinflammatory
processes in the hippocampus is of key importance for the development of negative post-ischemic function-
al consequences. However, it remains unclear which genes are involved in these processes. The aim of this
work was a comparative study of the expression of genes encoding glutamate and GABA transporters and
receptors, as well as inflammation markers in the hippocampus one day after two types of ischemic exposure
(according to Koizumi - MCAO-MK, and Longa - MCAO-ML), as well as after direct pro-inflamma-
tory activation by central administration of lipopolysaccharide (LPS). The results obtained revealed both
differences and similarities between the responses to the impacts applied in the work. A greater number
of genes that changed the expression associated with the activation of apoptosis and neuroinflammation,
glutamate reception, and markers of the GABAergic system were found after MCAO-ML and LPS, than
after MCAO-MK. In turn, MCAO-MK and LPS were characterized, in comparison with MCAO-ML,
by changes in a larger number of genes involved in glutamate transport. The most pronounced difference
between MCAO-ML and MCAO-MK and LPS was changes in the expression of genes for calmodulin and
calmodulin-dependent kinases. The revealed features of the responses of the hippocampal transcriptome
to two types of ischemia and a pro-inflammatory stimulus will contribute to further understanding of the
causes of the diversity of stroke consequences, both in model studies and in the clinic.
Keywords: hippocampus, middle cerebral artery occlusion, lipopolysaccharide, glutamate, GABA, genes expression
БИОХИМИЯ том 88 вып. 4 2023