БИОХИМИЯ, 2023, том 88, вып. 7, с. 1237 - 1253
УДК 577.19
ИССЛЕДОВАНИЕ ПРОТИВООПУХОЛЕВОГО ПОТЕНЦИАЛА
СОЛОКСОЛОН ТРИПТАМИДА В ОТНОШЕНИИ
МУЛЬТИФОРМНОЙ ГЛИОБЛАСТОМЫ С ПОМОЩЬЮ
in silico, in vitro И in vivo ПОДХОДОВ
© 2023 А.В. Марков*, К.В. Одаренко, А.В. Сенькова, А.А. Ильина, М.А. Зенкова
Институт химической биологии и фундаментальной медицины СО РАН,
630090 Новосибирск, Россия; электронная почта: andmrkv@gmail.com
Поступила в редакцию 02.02.2023
После доработки 13.03.2023
Принята к публикации 03.04.2023
Мультиформная глиобластома (МГБ) представляет собой крайне агрессивное онкологическое
заболевание, характеризующееся неконтролируемым диффузным ростом, развитием резистент-
ности к радио- и химиотерапии и высокой вероятностью рецидивов, что определяет низкую вы-
живаемость пациентов с МГБ. Учитывая большое количество сигнальных путей, задействованных
в регуляции патогенеза МГБ, разработка новых антиглиобластомных препаратов на основе муль-
титаргетных природных метаболитов является перспективным направлением исследований. В на-
стоящей работе изучен противоопухолевый потенциал полусинтетического тритерпеноида солок-
солон триптамида (СТА) в отношении клеток глиобластомы человека U87. Было установлено, что
СТА эффективно блокирует пролиферацию клеток U87 как в 2D-, так и в 3D-культуре in vitro,
усиливает адгезивные свойства опухолевых клеток и проявляет цитотоксическое действие синер-
гично с темозоломидом. С помощью подходов компьютерной биологии установлено, что выражен-
ная антиглиобластомная активность СТА может быть обусловлена его прямым взаимодействием с
EGFR, ERBB2 и AKT1, играющими важную роль в регуляции злокачественности МГБ. Помимо
прямого действия на клетки U87, СТА оказывал нормализующее действие на опухолевое микро-
окружение на мышиной гетеротопической ксенографтной модели глиобластомы U87, подавляя
развитие сети незрелых тонкостенных кровеносных сосудов и продукцию эластина в опухолевой
ткани. Полученные данные позволяют предложить СТА в качестве перспективного антиглиобла-
стомного кандидата.
КЛЮЧЕВЫЕ СЛОВА: мультиформная глиобластома, цианоеноновый фармакофор, пентациклические
тритерпеноиды, сфероиды, ангиогенез, синергичность, молекулярный докинг, генные сети, опухолевое
микроокружение.
DOI: 10.31857/S0320972523070126, EDN: FYGZSR
ВВЕДЕНИЕ
продолжительности жизни пациентов с МГБ
составляет лишь 15-18 месяцев [2]. Данный
Согласно данным Всемирной организации
факт объясняется высокой злокачественно-
здравоохранения, мультиформная глиобласто-
стью клеток глиобластомы, их способностью
ма (МГБ) представляет собой одно из наибо-
к диффузному инфильтративному росту в
лее агрессивных неопластических заболеваний
окружающие ткани и развитием устойчивости
головного мозга, распространенность кото-
клеток опухоли к радио- и химиотерапии, что
рого составляет примерно 1 случай на 100 000
определяет склонность МГБ к рецидивам [1].
населения в год [1]. Несмотря на получение
Низкая эффективность лечения данного онко-
медицинской помощи в полном объеме, вклю-
логического заболевания также связана с крайне
чающем максимальную резекцию опухоли, лу-
ограниченным арсеналом лекарственных пре-
чевое воздействие на ее ложе и химиотерапию
паратов, одобренных для терапии МГБ, вклю-
темозоломидом (ТМЗ), медианное значение чающим ТМЗ (алкилирующий агент на основе
Принятые сокращения: МГБ - мультиформная глиобластома; СТА - солоксолон триптамид; ТМЗ - темозоломид;
ALDH - альдегиддегидрогеназа; TGF-β - трансформирующий ростовой фактор бета.
* Адресат для корреспонденции.
1237
1238
МАРКОВ и др.
имидазолтетразина, препарат первой линии) и
их адгезионные свойства, проанализирована
бевацизумаб (гуманизированное рекомбинант-
синергичность цитотоксического действия СТА
ное антитело к эндотелиальному фактору роста
с ТМЗ в отношении клеток глиобластомы U87
сосудов (VEGF), препарат второй линии), ис-
и влияние СТА на ряд характеристик опухо-
пользование которых ограничивается развити-
левого микроокружения в эксперименте с глио-
ем у клеток МГБ лекарственной устойчивости
бластомой U87 in vivo. Кроме этого, с помо-
и низкой эффективностью их проникновения
щью подходов компьютерной биологии уста-
в опухолевую ткань соответственно [3]. Таким
новлен ряд потенциальных белковых мише-
образом, поиск новых лекарственных агентов,
ней СТА, определяющих его выраженные анти-
способных эффективно проникать через гема-
глиобластомные свойства. Полученные данные
тоэнцефалический барьер и блокировать про-
позволяют предложить СТА в качестве перспек-
лиферацию и инвазионные свойства клеток
тивного лекарственного кандидата, оказываю-
МГБ является критически важным направле-
щего комплексный модулирующий эффект на
нием современной медицинской химии.
ключевые характеристики, определяющие вы-
Отличительной особенностью МГБ явля-
сокую злокачественность МГБ.
ется ее выраженная гетерогенность, т.е. суще-
ствование в рамках одной опухоли различных
популяций опухолевых клеток, отличающихся
МАТЕРИАЛЫ И МЕТОДЫ
друг от друга профилем ключевых сигналь-
ных путей и, как результат, характеризующих-
Исследуемое соединение. Солоксолон трип-
ся разным функциональным состоянием [2].
тамид (СТА) был синтезирован в лаборатории
Данная особенность лежит в основе низкой
фармакологически активных веществ Ново-
эффективности современной химиотерапии
сибирского института органической химии
МГБ [4] и свидетельствует о перспективно-
им. Н.Н. Ворожцова СО РАН с.н.с., к.х.н. Са-
сти использования мультитаргетной страте-
ломатиной О.В., согласно опубликованной ра-
гии для создания новых антиглиобластомных
нее методике [7], и передан для проведения
препаратов [5]. Одним из важных источников
биологических испытаний в виде кристалли-
мультитаргетных фармакологически активных
ческого порошка. СТА был растворен в ди-
соединений являются природные метаболиты,
метилсульфоксиде (ДМСО) в концентрации
активно исследуемые в настоящее время в ка-
10 мМ и хранился при -20 °С до проведения
честве агентов, блокирующих пролиферацию
экспериментов.
и инвазивность клеток МГБ не только в кле-
Культивирование клеток МГБ человека U87.
точных экспериментах, но и на ортотопиче-
Клетки глиобластомы человека U87, получен-
ских моделях глиобластомы in vivo [6].
ные из Американской коллекции клеточных
Ранее нами в сотрудничестве с исследо-
культур
(«ATCC», США), культивировали
вателями из лаборатории фармакологически
в питательной среде Игла в модификации
активных веществ Новосибирского институ-
Дульбекко (DMEM; «Sigma Aldrich», США),
та органической химии им. Н.Н. Ворожцова
содержащей
10% эмбриональной телячьей
СО РАН было разработано полусинтетическое
сыворотки (FBS; «Gibco», США) и раствор
производное глицирретовой кислоты солоксо-
антибиотиковантимикотиков («MP Biomedicals»,
лон триптамид (СТА), обладающий комплекс-
Франция), включающий пенициллин, стреп-
ным противоопухолевым эффектом в отно-
томицин и амфотерицин в конечных концен-
шении клеток МГБ in vitro, включающим ин-
трациях 100 ед./мл, 0,1 мг/мл и 0,25 мкг/мл со-
дукцию митохондриального стресса с после-
ответственно. Культуру клеток поддерживали
дующим запуском апоптоза, блокировку клоно-
в инкубаторе с увлажненной атмосферой при
генного потенциала, клеточной подвижности
температуре 37 °C с 5%-ным содержанием CO2
и способности клеток МГБ к васкулогенной
(далее - стандартные условия).
мимикрии [7]. Кроме этого, на гетеротопиче-
Оценка цитотоксичности СТА на 2D-культу-
ской ксенографтной мышиной модели было
ре клеток U87 (МТТ-тест). Клетки U87 выса-
установлено, что СТА способен эффективно
живали в
96-луночные планшеты
(«TPP»,
проникать через гематоэнцефалический барь-
Швейцария) в концентрации 104 клеток на
ер и достоверно подавлять рост глиобласто-
лунку в среде DMEM, содержащей 10% FBS
мы человека U87 in vivo [7]. Целью настоя-
(DMEM/10% FBS), и инкубировали в стан-
щего исследования является расширение зна-
дартных условиях в течение 24 ч. Затем пи-
ний об антиглиобластомном потенциале СТА.
тательную среду заменяли на бессыворо-
В данной работе изучен эффект СТА на рост
точную с добавлением СТА (100-800 нМ), и
клеток МГБ человека U87 в 3D-культуре и
инкубировали клетки дополнительные 24 ч,
БИОХИМИЯ том 88 вып. 7 2023
АНТИГЛИОБЛАСТОМНЫЙ ПОТЕНЦИАЛ СТА
1239
после чего добавляли в каждую лунку раствор
групп фотографировали 4 поля при 200× уве-
соли тетразола (MTT) до конечной концен-
личении микроскопа с использованием встро-
трации 0,5 мг/мл. Спустя 2 ч инкубации среду
енной CMOS-камеры. Для каждой экспе-
удаляли, добавляли 100 мкл ДМСО для раство-
риментальной группы определяли площадь
рения кристаллов формазана и измеряли по-
не менее 10 сфероидо-подобных структур с
глощение растворов при длине волны 570 нм и
помощью программы ImageJ и нормировали
референсной длине волны 620 нм на многока-
данный показатель на среднюю площадь сфе-
нальном спектрофотометре Multiscan RC plate
роидов в контрольной группе (инкубация с
reader («Thermo LabSystems», Финляндия).
TGF-β, но без СТА).
Исследование эффекта СТА на рост сфе-
Исследование эффекта СТА на адгезивные
роидов клеток U87. Оценку воздействия СТА
свойства клеток U87 (трипсиновый тест). Клет-
на рост сфероидов U87 проводили с помощью
ки U87 высаживали в 96-луночные планшеты
двух экспериментальных методик: путем куль-
в концентрации 104 клеток на лунку в среде
тивирования клеток на планшетах, дно кото-
DMEM/10% FBS и культивировали в течение
рых было покрыто 1%-ной агарозой (1) либо
ночи в стандартных условиях. Затем заменяли
гидрофильным, неиногенным, нейтрально за-
питательную среду на среду без сыворотки,
ряженным гидрогелем («Corning», США); да-
содержащую СТА в концентрации 25-100 нМ,
лее - планшеты со сверхнизким сцеплением
и инкубировали 24 ч в стандартных условиях.
дна (2). На первом этапе клетки U87 в среде
По окончании инкубации к клеткам добавля-
DMEM/10% FBS помещали в покрытые агаро-
ли 30 мкл TrypLETM («Gibco»), разбавленного
зой 48-луночные планшеты («TPP») и 6-луноч-
фосфатно-солевым буфером (PBS) (1 : 10), по-
ные планшеты («Servicebio», Китай) со сверх-
мещали планшет в CO2-инкубатор на 3 мин,
низким сцеплением дна в концентрации 2 × 103
после чего дважды промывали клетки средой
и 5 × 105 клеток на лунку соответственно. После
DMEM/10% FBS. Количество клеток, остав-
инкубации в течение 7 и 4 дней в стандартных
шихся прикрепленными ко дну лунки после
условиях для методик (1) и (2) соответственно
отмывки, оценивали с помощью МТТ-теста.
к получившимся сфероидам добавляли СТА
Исследование эффекта СТА на адгезивность
в среде DMEM без добавления сыворотки до
клеток U87 к коллагеновой подложке. Клетки
конечных концентраций 0,1-2 мкМ. Фото-
U87 высаживали в 6-луночные планшеты в
графии сфероидов делали с помощью инвер-
концентрации 2,8 × 105 клеток на лунку в среде
тированного микроскопа ZEISS Primo Vert со
DMEM/10% FBS и инкубировали в течение
встроенной камерой ZEISS AxioCam ERc5s
ночи в стандартных условиях. Затем среду уби-
(«Carl Zeiss Microscopy GmbH», Германия)
рали, к клеткам добавляли СТА (25-100 нМ)
сразу после добавления растворов СТА и затем
в среде DMEM без добавления сыворотки и
каждые 2-3 суток до 28 суток инкубации (1)
дополнительно культивировали 24 ч в стан-
либо однократно через 2 суток после добавле-
дартных условиях. По окончании инкубации
ния тритерпеноида (2). Для каждой экспери-
клетки открепляли от поверхности планше-
ментальной группы определяли площадь 4 (1)
тов с помощью TrypLETM, ресуспендировали
или 250 (2) сфероидов с помощью программы
в среде DMEM/10% FBS и высаживали в кон-
ImageJ («NIH», США) и нормировали данный
центрации 105 клеток на лунку в 96-лучночные
показатель на среднюю площадь сфероидов
планшеты, дно которых было предварительно
в контроле (клетки U87 без добавления СТА).
покрыто коллагеном, выделенным из хвоста
Исследование эффекта СТА на TGF-β-сти-
крысы («Cell Applications, Inc.», США). Через
мулированное формирование сфероидо-подоб-
1 ч инкубации в стандартных условиях лунки
ных структур на монослое клеток U87. Клет-
планшетов дважды промывали PBS и оценива-
ки U87 в среде DMEM/10% FBS высажива-
ли с помощью MTT-теста количество клеток,
ли в 96-луночные планшеты в концентрации
прикрепившихся к коллагеновой подложке.
5 × 103 клеток на лунку и культивировали в те-
Исследование сочетанного действия СТА и
чение ночи в стандартных условиях. Затем
ТМЗ на клетках U87. Клетки U87 высажива-
заменяли питательную среду на среду без
ли в 96-луночные планшеты в концентрации
сыворотки, содержащую TGF-β1
(50 нг/мл)
104 клеток на лунку в среде DMEM/10% FBS
(«Prospec», Израиль) и СТА (200, 400 мкМ).
и инкубировали ночь в стандартных условиях.
Через 48 ч после индукции формирование сфе-
Затем среду в лунках заменяли на DMEM без
роидо-подобных структур регистрировали с по-
сыворотки, добавляли СТА и/или темозоло-
мощью инвертированного микроскопа EVOS XL
мид до конечных концентраций 0,1-1 и/или
Core («Thermo Fisher Scientific», США). Для
100-500 мкМ соответственно и культивировали
каждой экспериментальной и контрольной
клетки в течение 72 ч в стандартных условиях.
БИОХИМИЯ том 88 вып. 7 2023
1240
МАРКОВ и др.
По окончании инкубации количество живых
На следующем этапе с помощью молеку-
клеток U87 оценивали с помощью MTT-те-
лярного докинга проводили моделирование
ста. Анализ типа сочетанного действия соеди-
взаимодействия СТА с наиболее связанными
нений проводили с помощью веб-ресурса
с генной сетью глиобластомы (т.е. характе-
ризующимися наименьшим значением ран-
Анализ активности альдегиддегидрогеназы
га) мишенями. Кристаллические структуры
в клетках U87. Клетки U87 в среде DMEM/10%
EGFR (Protein Data Bank (PDB) ID: 8A2D),
FBS высаживали в 6-луночные планшеты в кон-
AKT1 (PDB ID: 3O96), FYN (PDB ID: 2DQ7)
центрации 2,8 × 105 клеток на лунку и инкуби-
и ERBB2 (PDB ID: 3RCD) были взяты из базы
ровали в течение ночи в стандартных усло-
виях. Затем среду убирали, и к клеткам добав-
них удаляли молекулы воды и кокристалли-
ляли СТА в концентрациях 0,5 и 1 мкМ в среде
зованные лиганды (известные ингибиторы)
DMEM без сыворотки и вновь инкубировали
с помощью Discovery Studio Visualizer v21.1.0
24 ч в стандартных условиях. После этого ак-
и добавляли полярные водороды и частичные
тивность альдегиддегидрогеназы (ALDH) опре-
атомные заряды по методу Гастейгера с помо-
деляли с помощью коммерческого набора
щью AutoDock Tools 1.5.7. 2D-Структуры СТА
AldeRed® ALDH Detection Assay
(«Sigma-
и известных ингибиторов переводили в трех-
Aldrich Inc.», Германия), согласно протоколу
мерную форму в MarvinSketch v22.1, опти-
производителя. Коротко, клетки снимали с
мизировали геометрию молекул с помощью
поверхности планшета с помощью TrypLETM,
Avogadro v1.2.0 (силовое поле MMFF94) и
ресуспендировали в 1 мл буфера AldeRed Assay
устанавливали способность связей внутри
Buffer и добавляли в каждую пробирку 5 мкл
лигандов к свободному вращению с помо-
флуоресцентного красителя AldeRed, а в кон-
щью AutoDock Tools
1.5.7. Моделирование
трольные пробирки дополнительно добавляли
взаимодействий
«лиганд-белок» проводили
5 мкл ингибитора ALDH диэтиламинобенз-
в программе AutoDockVina [13] с использо-
альдегида (DEAB). После инкубации в течение
ванием следующих параметров: EGFR
-
45 мин в стандартных условиях клетки соби-
size_x = 22, size_y = 14, size_z = 16, center_x =
рали с помощью центрифугирования, ресус-
= -1,379, center_y = -9,243, center_z = 18,98;
пендировали их в 200 мкл буфера, помещали
AKT1 - size_x = 14, size_y = 18, size_z = 18,
пробирки в лед и проводили анализ образ-
center_x = 8,507, center_y = -7,504, center_z =
цов на проточном цитофлуориметрe NovoCyte
= 10,15; FYN - x = 14, size_y = 14, size_z = 14,
(«ACEA Biosciences, Inc», США) с помощью
center_x = -15,594, center_y = 17,532, center_z =
программного обеспечения NovoExpress Soft-
= -12,787; ERBB2 - x = 16, size_y = 16, size_z =
ware («ACEA Biosciences, Inc»). Для каждого
= 14, center_x = 13,147, center_y = 29,114, center_z =
образца проводили анализ 10 000 событий.
= 29,114. Докинг-комплексы с прочными взаи-
Идентификация потенциальных белковых ми-
модействиями между лигандом и белком
шеней СТА с помощью in silico подходов. На пер-
(ΔG ≤ 7,0 ккал/моль) визуализировали с по-
вом этапе анализа определяли возможные пер-
мощью Discovery Studio Visualizer v21.1.0 и
вичные белковые мишени СТА путем анализа
LigPlot+ v.2.2.7 в 3D- и 2D-форматах соответ-
его структуры хемоинформатическими платфор-
ственно и отбирали структуры, стабилизиро-
мами Polypharmacology Browser 2.0 [9] и Swiss-
ванные наибольшим количеством водородных
TargetPrediction
[10], после чего добавляли
связей.
их по отдельности к списку генов, ассоции-
Потенциальные белковые мишени СТА,
рованных, согласно базе данных DisGeNET,
отобранные с помощью молекулярного до-
с патогенезом МГБ C1621958 (3197 генов), и
кинга, наносили на иерархическое дерево ки-
строили для дополненных списков генные
нома человека с помощью платформы KinMap
ассоциативные сети с использованием базы
данных STRING (the Search Tool for Retrieval
которого отмечали киназы, ассоциирован-
of Interacting Genes/Proteins) [11]. Полученные
ные с патогенезом МГБ (согласно базе дан-
сети визуализировали в программной среде
Cytoscape 3.9.1 и анализировали с помощью
opentargets.org/)).
плагина cytoHubba [12]: для каждой первич-
Лабораторные животные. В работе исполь-
ной мишени СТА вычисляли «Degree», «MCC»
зовали 6-8-недельных бестимусных мышей-
и «Betweenness» (характеристики центральности
самок линии Nude развода Российского на-
гена в сети), ранжировали списки мишеней по
ционального центра генетических ресурсов
данным показателям и вычисляли «общий ранг»
лабораторных животных на базе SPF-вива-
как среднее от ранга мишени по трем показателям.
рия Института цитологии и генетики (ИЦиГ)
БИОХИМИЯ том 88 вып. 7 2023
АНТИГЛИОБЛАСТОМНЫЙ ПОТЕНЦИАЛ СТА
1241
СО РАН (Новосибирск, Россия). Животных со-
где Pструктура - количество точек, которые при-
держали в SPF-виварии ИЦиГ СО РАН по 6 осо-
ходятся на структуру, а Pтест - общее количе-
бей в клетке в соответствии с нормами содер-
ство тестовых точек, в данном случае - 100.
жания SPF-животных. Мыши имели свобод-
Численную плотность оцениваемой гисто-
ный доступ к еде и воде. Клетки, подстилка,
логической структуры определяли путем под-
корм и вода подвергались автоклавированию.
счета количества структур в пределах тестового
Гетеротопическая ксенографтная модель
поля, в данном случае - 3,2 × 106 мкм2. Всего
МГБ. Для формирования ксенографтной моде-
оценивали по 5 полей зрения в каждом об-
ли с первичным опухолевым узлом суспензию
разце для каждой гистологической структуры;
клеток глиобластомы U87 (2 × 107 клеток/мл)
таким образом, всего было проанализировано
вводили подкожно в левый бок мышам линии
по 30 полей зрения в каждой эксперименталь-
Nude в 0,1 мл 0,9%-ного раствора натрия хло-
ной группе.
рида c 0,05 мл матригеля (Matrigel® Matrix;
Статистический анализ полученных данных
«Corning», США). На 5-й день после трансплан-
проводили с помощью программного пакета
тации животных распределяли на группы -
GraphPad Prism 8.0.1. Проверку нормальности
по 6 мышей в каждой: (1) мыши, получавшие
распределения количественных показателей
внутрибрюшинные (в.б.) инъекции 10%-ного
проводили с использованием критерия Ша-
Tween-80 (контроль) и
(2) мыши, получав-
пиро-Уилка. Для оценки достоверности раз-
шие в.б. инъекции соединения СТА в 10%-ном
личий между группами с нормальным и не-
Tween-80 в дозе 20 мг/кг. Все в.б. инъекции
нормальным распределениями применяли
проводили в объеме 200 мкл 3 раза в неделю.
непарный критерий Стьюдента и непарамет-
Всего было произведено 7 инъекций. Мышей
рический U-критерий Манна-Уитни соответ-
выводили из эксперимента на 21-й день роста
ственно. Различия считались достоверными
опухоли путем дислокации шейных позвонков
при уровне значимости p < 0,05.
под изофлюрановой анестезией. Производили
забор опухолевых узлов для последующего гис-
тологического исследования.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Гистология. Опухолевые узлы фиксирова-
ли в 10%-ном нейтральном формалине («Bio-
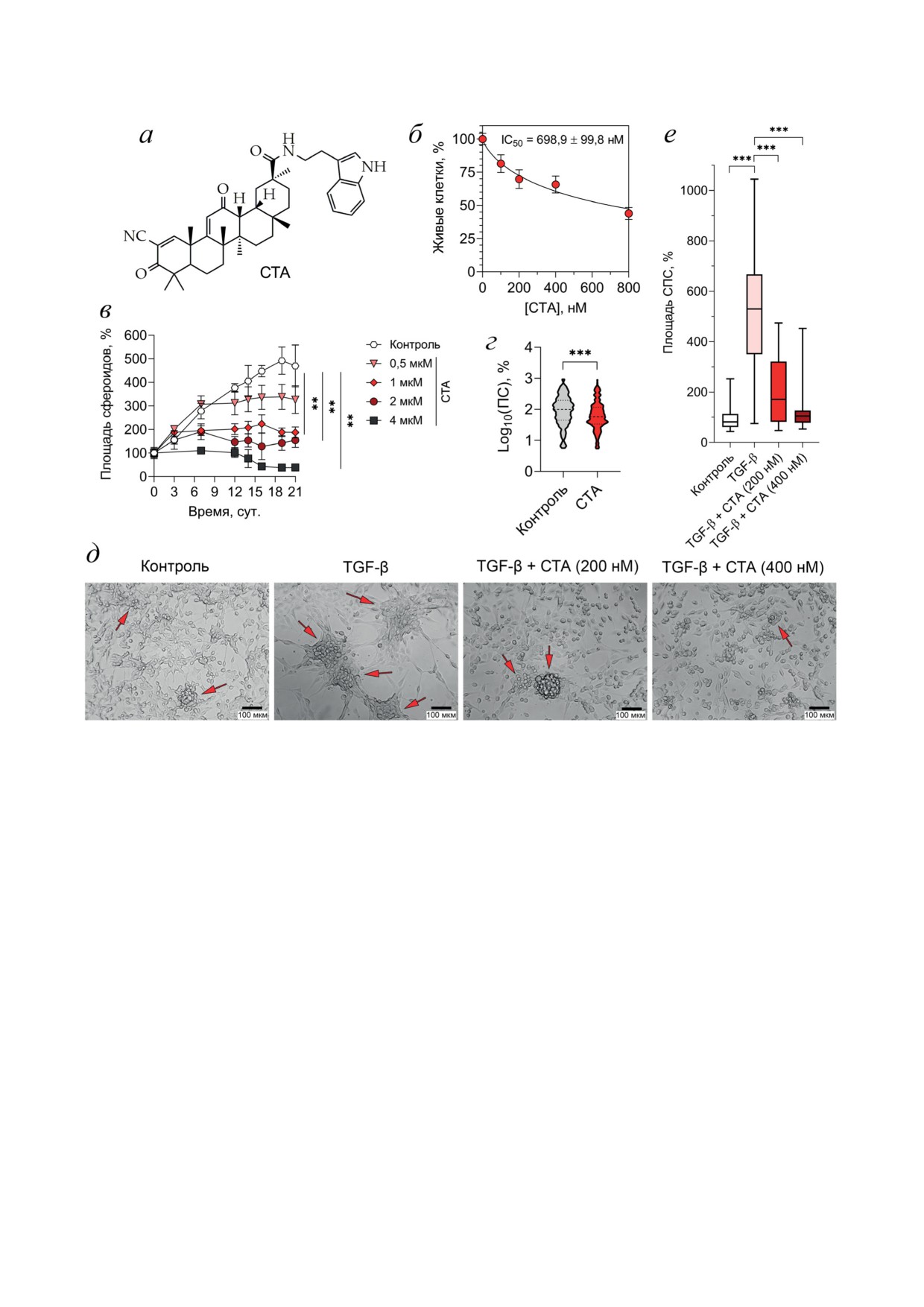
СТА эффективно подавляет рост сфероидов
Vitrum», Россия), затем производили обез-
клеток U87. На первом этапе исследования
воживание в спиртах возрастающей концен-
была оценена цитотоксичность СТА (рис. 1, а)
трации, просветляли ксилолом и заключали в
в отношении клеток МГБ человека U87 in vitro.
парафин HISTOMIX («BioVitrum»). Парафи-
Исследуемый тритерпеноид эффективно по-
новые срезы толщиной до 5 мкм окрашивали
давлял жизнеспособность клеток U87 в моно-
гематоксилином и эозином. Локализацию и
слое в низких субмикромолярных концен-
количество стромального компонента опухоли
трациях: его значение IC
24 ч) составило около
(эластиновые волокна) определяли с помо-
0,7 мкМ (рис. 1, б). Учитывая, что монослой
щью окрашивания срезов по Ван Гизону [14].
клеток (2D-культура) не в полной мере моде-
Гистологические срезы анализировали на мик-
лирует опухолевый рост in vivo, при котором
роскопе Axiostar plus, оснащенном камерой
опухолевые клетки в зависимости от поло-
Axiocam MRc5 («Zeiss», Германия).
жения в опухолевом узле имеют различный
Морфометрическое исследование опухоле-
доступ к питательным веществам и кисло-
вых узлов. Морфометрический анализ гисто-
роду, противоопухолевый потенциал СТА был
логических срезов опухолевых узлов проводи-
далее проанализирован на сфероидах МГБ
ли с использованием закрытой тестовой си-
(3D-культура). Для этого к сфероидам клеток
стемы из 100 точек площадью 3,2 × 106 мкм2
U87, сформированным в культуральных план-
при 200× увеличении микроскопа, наклады-
шетах, покрытых 1%-ной агарозой, добавляли
вая сетку случайным образом. При проведе-
СТА (0,5-4 мкМ) и оценивали их относитель-
нии морфометрического исследования опу-
ную площадь каждые 2-4 дня в течение 21 су-
холевых узлов подсчитывали объемные плот-
ток. Как видно из рис. 1, в, в ходе эксперимен-
ности стромального компонента опухолевой
та площадь сфероидов в контроле (без СТА)
ткани и численную плотность тонкостенных
линейно росла, тогда как СТА дозозависимо
кровеносных сосудов в ткани опухоли. Объ-
тормозил рост сфероидов, причем эти раз-
емную плотность оцениваемой гистологиче-
личия в площади сфероидов становятся замет-
ской структуры подсчитывали по формуле:
ными, начиная с 7 дня инкубации. К концу
эксперимента наблюдалось значительное сни-
Vv = (Pструктура/Pтест) × 100%,
жение площади сфероидов в 2,5, 3 и 12,1 раза
БИОХИМИЯ том 88 вып. 7 2023
1242
МАРКОВ и др.
Рис. 1. Влияние СТА на рост клеток глиобластомы человека. а - Структурная формула СТА. б - Цитотоксичность
СТА в отношении клеток U87. Клетки инкубировали с указанными концентрациями СТА в течение 24 ч, и затем оце-
нивали уровень живых клеток с помощью МТТ-теста. в - Влияние СТА на рост сфероидов U87 в планшетах, дно ко-
торых покрыто агарозой, и планшетах со сверхнизким сцеплением дна (г). К сфероидам U87 добавляли СТА в указан-
ных концентрациях и оценивали площадь сфероидов в указанных временных точках (в) или через 48 ч инкубации (г)
с помощью фазово-контрастной микроскопии. д и е - Влияние СТА на формирование сфероидо-подобных структур
в монослое клеток U87 под действием TGF-β. Клетки инкубировали с TGF-β1 (50 нг/мл) и СТА (200, 400 мкМ)
в течение 48 ч, и оценивали площадь сфероидов с помощью фазово-контрастной микроскопии. Фотографии сделаны
при 200× увеличении (д), стрелками показаны сфероидо-подобные структуры. Площади сфероидов (в, г) и сфероидо-
подобных структур (е) были подсчитаны с помощью программы ImageJ
по сравнению с контролем для концентра-
вая их стволовые свойства, инвазивность и
ций СТА 1, 2 и 4 мкМ соответственно (рис. 1, в).
метастазирование [15]. Учитывая, что TGF-β
Наблюдаемый супрессивный эффект СТА в
является также известным стимулятором
отношении роста клеток МГБ в 3D-культуре
сфероидного роста клеток глиобластом [16],
был также подтвержден в независимом экс-
на следующем этапе работы был исследован
перименте со сфероидами клеток U87, сфор-
эффект СТА на формирование сфероидо-по-
мированными на культуральных планшетах
добных структур в монослое TGF-β-стиму-
со сверхнизким сцеплением дна. Из графика,
лированных клеток U87. Мы показали, что
представленного на рис. 1, г, видно, что инку-
3-дневный интактный монослой клеток U87 ха-
бация сфероидов в присутствии 2 мкМ СТА в
рактеризовался небольшим количеством мно-
течение 48 ч приводила к достоверному сниже-
гослойных структур, размер которых в 5,3 раза
нию их площади.
увеличивался при стимуляции клеток TGF-β
Известно, что трансформирующий росто-
по сравнению с контролем, причем данный
вой фактор бета (TGF-β) играет важную роль
эффект не был связан с усилением пролифера-
в регуляции злокачественности МГБ, усили-
ции клеток (рис. 1, д и е). Инкубация TGF-β-
БИОХИМИЯ том 88 вып. 7 2023
АНТИГЛИОБЛАСТОМНЫЙ ПОТЕНЦИАЛ СТА
1243
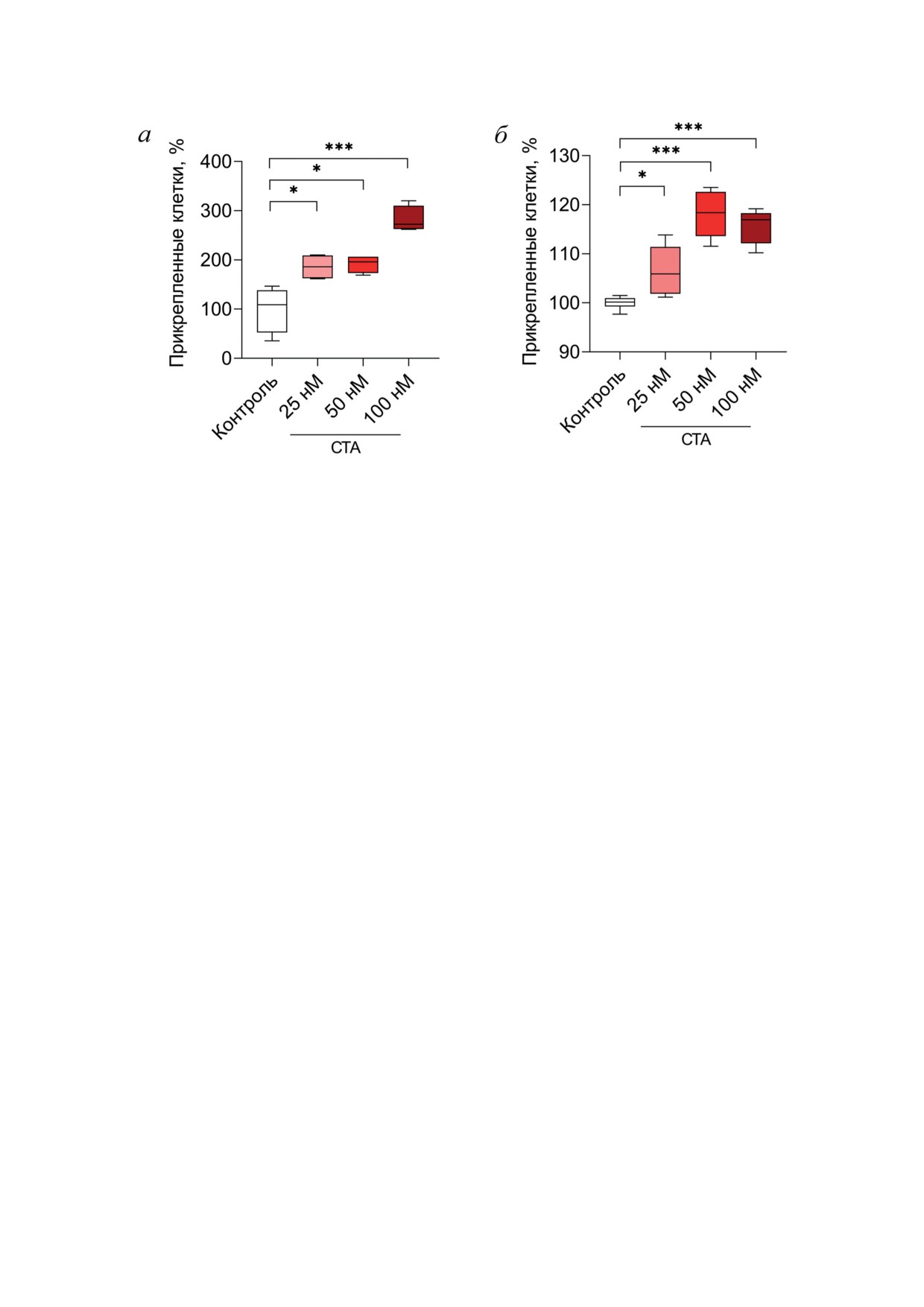
Рис. 2. Влияние СТА на адгезивные свойства клеток глиобластомы человека. а - Воздействие СТА на адгезивность
клеток U87, оцененную методом трипсинизации. Клетки инкубировали с СТА в указанных концентрациях в течение
24 ч, затем обрабатывали TrypLETM, отмывали средой DMEM и определяли число клеток, оставшихся прикреплен-
ными ко дну планшета с помощью МТТ-теста. б - Эффект СТА на адгезию клеток U87 к коллагеновой подложке.
Клетки индуцировали СТА в указанных концентрациях в течение 24 ч, затем высаживали на 1 ч в лунки 96-луночного
планшета, дно которого было покрыто коллагеном из хвоста крыс, после чего отмывали PBS и оценивали количество
клеток, прикрепившихся к коллагеновой подложке, с помощью MTT-теста. Данные представлены как среднее ± стан-
дартное отклонение четырех (а) и шести (б) независимых экспериментов
стимулированных клеток в присутствии СТА
стоверно усиливает адгезивность клеток U87,
в концентрации 200 нМ достоверно в 2,6 раза
причем с увеличением концентрации СТА в
снижала размер сфероидо-подобных форми-
среде увеличивается и количество неоткрепив-
рований по сравнению с TGF-β-обработан-
шихся клеток. Количество клеток, оставшихся
ным контролем, а при дальнейшем повыше-
прикрепленными ко дну планшета после дей-
нии концентрации СТА до 400 нМ площадь
ствия TrypLETM, в экспериментальных груп-
сфероидо-подобных структур в TGF-β-стиму-
пах превышало данный показатель в контроле
лированных клетках U87 снижалась до значе-
в 1,9-2,8 раза.
ний интактного контроля (рис. 1, д и е).
Дополнительно был изучен эффект СТА в
СТА усиливает адгезивные свойства кле-
отношении адгезии клеток U87 к коллагено-
ток U87. Установив наличие у СТА выражен-
вой подложке, имитирующей in vitro внекле-
ных антипролиферативных свойств в отноше-
точный матрикс. Клетки U87 инкубировали
нии клеток U87 (рис. 1), далее мы проанали-
в присутствии 25-100 нМ СТА в течение 24 ч,
зировали, обладает ли исследуемый тритерпе-
после чего их снимали с планшета с помо-
ноид противоопухолевым потенциалом в не-
щью TrypLETM и переносили на 96-луночный
токсичных концентрациях, сконцентрировав
планшет, дно лунок которого было покрыто
внимание на таком свойстве клеток U87, как
коллагеном из хвоста крыс. Клетки инкубиро-
адгезивность, ослабление которой ассоцииро-
вали на коллагеновых подложках в течение 1 ч,
вано с высоким метастатическим потенциа-
отмывали от неприкрепившихся клеток и оце-
лом МГБ [17].
нивали количество прикрепившихся клеток
Для оценки влияния СТА на адгезионную
с помощью МТТ-теста. Как и в случае с пре-
активность клеток монослой клеток U87 инку-
дыдущим экспериментом, СТА достоверно
бировали в отсутствие (контроль) или в при-
усиливал адгезионную активность клеток U87,
сутствии СТА в концентрациях 25-100 нМ в
при этом наибольшее усиление адгезии на-
течение 24 ч, после чего обрабатывали клетки
блюдалось при концентрации СТА в среде 50
разбавленным раствором рекомбинантного
и 100 нМ (рис. 2, б).
трипсина TrypLETM в течение 3 мин, отмывали
СТА действует синергически в комбинации
открепившиеся клетки и оценивали количе-
с ТМЗ в отношении жизнеспособности кле-
ство клеток, прикрепленных ко дну планшета,
ток U87. Как было отмечено выше, препа-
с помощью МТТ-теста. Как видно из рис. 2, а,
ратом первой линии при терапии МГБ яв-
СТА во всех используемых концентрациях до-
ляется ТМЗ, который представляет собой
БИОХИМИЯ том 88 вып. 7 2023
1244
МАРКОВ и др.
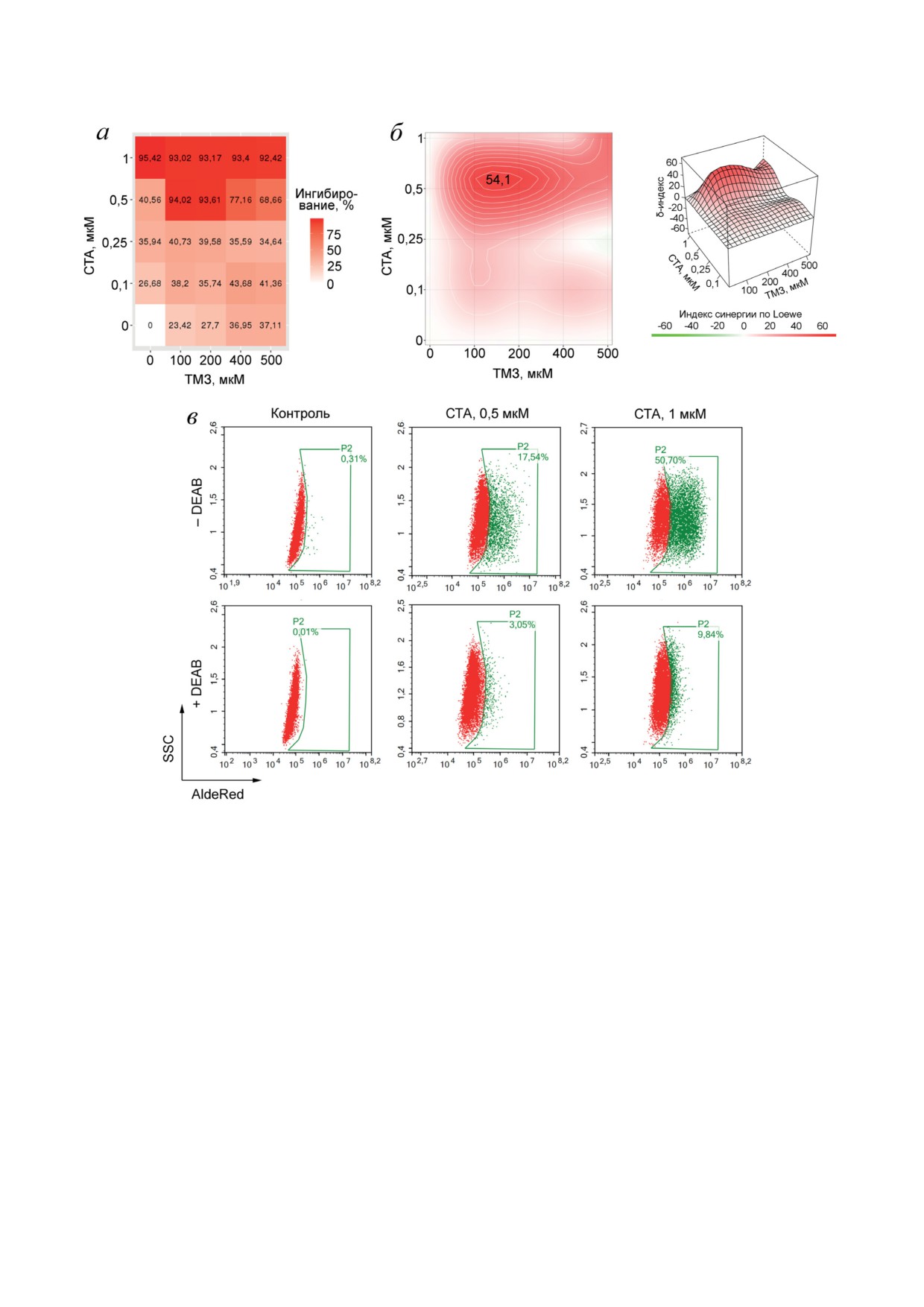
Рис. 3. Синергическое действие СТА и ТМЗ на клетки глиобластомы U87. а - Сочетанное действие СТА и ТМЗ
в отношении жизнеспособности клеток U87. Клетки U87 культивировали с СТА и/или ТМЗ в указанных концентра-
циях в течение 72 ч, после чего количество живых клеток определяли с помощью МТТ-теста. б - Профиль совмест-
ного цитотоксического действия СТА и ТМЗ, построенный с помощью веб-ресурса SynergyFinder. в - Влияние СТА
на активность ALDH в клетках U87. Клетки U87 инкубировали с СТА в указанных концентрациях в течение 24 ч
и оценивали активность ALDH с помощью набора AldeRed® ALDH Detection Assay и проточной цитометрии. DEAB -
диэтиламинобензальдегид
ДНК-метилирующий агент, способствующий
500 мкМ) в течение 72 ч, после чего опреде-
образованию в цепях ДНК O6-метилгуанина,
ляли количество жизнеспособных клеток с по-
последующему появлению в них двуцепочеч-
мощью МТТ-теста и анализировали характер
ных разрывов в результате репарации нека-
сочетанного действия исследуемых препаратов
нонических пар нуклеотидов и, как результат,
с помощью платформы SynergyFinder 2.0 [8].
гибели клеток МГБ [18]. Учитывая критиче-
Как видно из тепловой карты, представлен-
скую важность данного алкилирующего агента
ной на рис. 3, а, ТМЗ обладал слабым уров-
в схемах лечения МГБ, важным аспектом раз-
нем цитотоксичности в отношении клеток
работки новых антиглиобластомных лекар-
U87 (IC
72 ч) > 500 мкМ), в то время как СТА в
ственных кандидатов является оценка их со-
концентрации 1 мкМ приводил практически
72 ч)
четанного действия с ТМЗ. Для этого клетки
к полной гибели опухолевых клеток IC
=
U87 инкубировали в присутствии различных
= 0,4 ± 0,1 мкМ). При совместном примене-
концентраций СТА (0,1-1 мкМ) и ТМЗ (100-
нии ТМЗ и СТА действовали синергически
БИОХИМИЯ том 88 вып. 7 2023
АНТИГЛИОБЛАСТОМНЫЙ ПОТЕНЦИАЛ СТА
1245
при использовании СТА в пределах значе-
потенциальные мишени тритерпеноида по
ния его IC50: наибольший δ-индекс синергии
уровню взаимодействия с МГБ-ассоциирован-
составлял 54,1 при сочетании 0,5 мкМ СТА
ным регуломом. Проведенный анализ показал,
и 100 мкМ ТМЗ, вызывая гибель 94% клеток
что наибольшей взаимосвязью с патогенезом
глиобластомы U87 через 72 ч (рис. 3, б). Инте-
МГБ среди проанализированных белков обла-
ресно, что дальнейшее повышение концен-
дали рецептор эпидермального ростового фак-
траций исследуемых соединений приводило к
тора (EGFR), серин-треониновая протеинки-
снижению δ-индекса (рис. 3, б), что может сви-
наза AKT1, тирозиновая протеинкиназа FYN
детельствовать о запуске в клетках U87 ком-
и рецепторная тирозиновая протеинкиназа
пенсационных механизмов в ответ на нараста-
ERBB2 (известная также как HER2) (рис. 4, а).
ющий ксенобиотический стресс. Для проверки
Дальнейшее молекулярное моделирование под-
данной гипотезы был исследован эффект СТА
твердило способность СТА взаимодействовать
на уровень активности альдегиддегидрогена-
с активными центрами EGFR, AKT1 и ERBB2
зы, играющей важную роль в детоксикации ан-
в сайтах связывания их известных ингибито-
тинеопластических препаратов и подавлении
ров с низкими значениями свободной энер-
вызванного ими окислительного стресса [19].
гии Гиббса (ΔG = -9,2, -7,1 и -7,0 ккал/моль
Клетки U87 инкубировали в присутствии СТА
соответственно) (рис. 4. б). FYN был исклю-
в концентрациях 0,5 и 1 мкМ в течение 24 ч,
чен из дальнейшего анализа, так как получен-
после чего проводили их окраску с помощью
ный докинг-комплекс СТА с данным белком
AldeRed, являющегося флуоресцирующим суб-
характеризовался энергией связывания ΔG =
стратом ALDH, и анализировали клетки с
= -5,3 ккал/моль, значительно превышающей
помощью проточной цитометрии. Как видно
пороговую величину (ΔG ≤ -7,0 ккал/моль)
из цитограмм, представленных на рис. 3, в,
(рис. 4, б). Подробный анализ полученных струк-
в контроле отсутствовали клетки с высоким
тур показал, что молекула СТА формирует свя-
уровнем активности ALDH (клетки ALDHhigh;
зи с аминокислотными остатками, играющи-
0,3%), в то время как СТА в концентрациях
ми важную роль в функционировании EGFR,
0,5 и 1 мкМ вызывал увеличение доли клеток
AKT1 и ERBB2, включая водородные связи
ALDHhigh до 17,5% и 50,7% соответственно.
с Arg858 (EGFR), Glu85 и Cys296 (AKT1),
Внесение в культуральную среду селективного
Lys753, Asn850 и Asp863 (ERBB2), причем вы-
ингибитора ALDH диэтиламинобензальдегида
явленная сеть взаимодействий СТА оказалась
значительно снижало наблюдаемый эффект
схожей с сетью взаимодействия известных ин-
тритерпеноида, свидетельствуя о том, что вы-
гибиторов анализируемых белков (рис. 4, в).
явленное усиление флуоресцентного сигнала
Последующее нанесение белковых мишеней
клеток в экспериментальных группах связано
СТА, верифицированных молекулярным до-
именно с активностью ALDH (рис. 3, в).
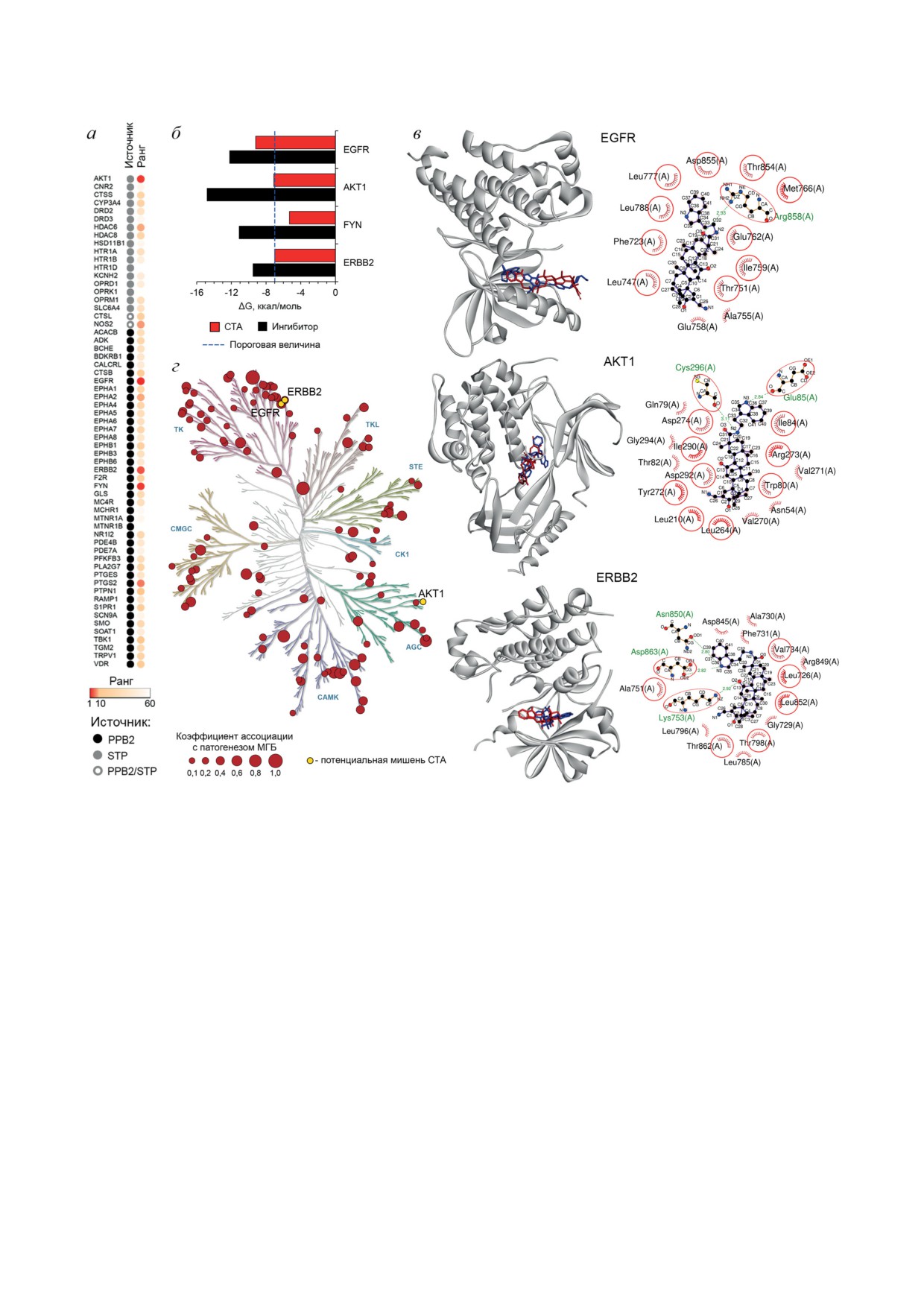
кингом, на филогенетическое дерево, содер-
EGFR, ERBB2 и AKT1 являются потенци-
жащее МГБ-ассоциированный кином челове-
альными белковыми мишенями СТА, опреде-
ка (реконструированный на основе базы дан-
ляющими его выраженную антиглиобластомную
ных Open Targets Platform), независимо под-
активность. Известно, что пентациклические
твердило связь EGFR и ERBB2 с патогенезом
тритерпеноиды, как природные, так и полу-
МГБ (рис. 4, г). Интересно, что AKT1, несмот-
синтетические, характеризуются широким
ря на ее высокий уровень интеграции в МГБ-
спектром молекулярных мишеней внутри кле-
специфичный регулом (рис. 4, а), оказалась
ток [20]. Для того чтобы определить потенци-
не ассоциированной с патогенезом глиобла-
альные белковые мишени СТА, ассоцииро-
стомы (рис. 4, г), что, возможно, объясняется
ванные с патогенезом МГБ, был использован
сложностями в заполнении базы данных Open
комплекс методов компьютерной биологии.
Targets Platform информацией по AKT1, так
На первом этапе на основе анализа структуры
как в большинстве опубликованных работ,
СТА с использованием хемоинформатических
посвященных связи AKT c МГБ, использова-
платформ Polypharmacology Browser
2.0
[9]
лись ингибиторы пан-АКТ, обладающие сход-
и SwissTargetPrediction [10] мы идентифици-
ной эффективностью действия на различные
ровали ряд его возможных белковых мише-
изоформы AKT [21]. Несмотря на это, AKT1,
ней (рис. 4, а). Параллельно с этим был рекон-
по-видимому, играет важную роль в прогрессии
струирован МГБ-ассоциированный регулом
МГБ - подавление ее экспрессии с помощью
на основе ключевых генов, связанных с МГБ,
малых интерферирующих РНК и антисмыс-
по данным DisGeNET, в который далее вне-
ловых олигонуклеотидов достоверно снижало
дряли выявленные белки-мишени СТА, ана-
пролиферацию клеток МГБ в клеточных и мы-
лизировали топологию сетей и ранжировали
шиных экспериментах соответственно [22, 23].
БИОХИМИЯ том 88 вып. 7 2023
1246
МАРКОВ и др.
Рис. 4. Определение возможных белковых мишеней СТА, ассоциированных с МГБ. а - Анализ интегрированности
потенциальных мишеней СТА в регулом МГБ. Первичные мишени СТА были предсказаны с помощью веб-ресурсов
Polypharmacology Browser (PPB2) и SwissTargetPrediction (SWT), их взаимодействие с МГБ-ассоциированным регуло-
мом было воссоздано на основе информации из баз данных DisGeNET и STRING и проанализировано с помощью
плагина cytoHubba в программной среде Cytoscape. Ранг мишени определен на основе трех характеристик централь-
ности гена в генной регуляторной сети (Degree, MCC и Betweenness), и его величина обратно пропорциональна зна-
чимости мишени в регуляции МГБ. б - Энергии связывания СТА и известных ингибиторов с AKT1 и EGFR, опре-
деленные с помощью молекулярного докинга. в - Трехмерное и двухмерное изображения докинг-комплексов EGFR
и AKT1 с СТА. Красными окружностями обозначены аминокислотные остатки, являющиеся общими для докинг-
комплексов СТА и известных ингибиторов EGFR и AKT1. Зелеными пунктирными линиями обозначены водородные
связи. г - Филогенетическое дерево кинома человека, реконструированное с помощью платформы KinMap. Взаимо-
связь киназ с патогенезом МГБ восстановлена с использованием базы данных Open Targets Platform [24]
Таким образом, проведенный in silico ана-
на гетеротопической ксенографтной модели
лиз выявил потенциальную способность СТА
глиобластомы U87 нами было показано, что
взаимодействовать с активными центрами ряда
7-кратная внутрибрюшинная инъекция СТА в
киназ, связанных с патогенезом МГБ, включая
дозе 20 мг/кг эффективно снижала рост пери-
EGFR, ERBB2 и AKT1. Полученные данные
ферических опухолевых узлов, подавляя про-
требуют дальнейшего экспериментального под-
лиферативный потенциал опухолевых клеток
тверждения.
и модулируя опухолевое микроокружение [7].
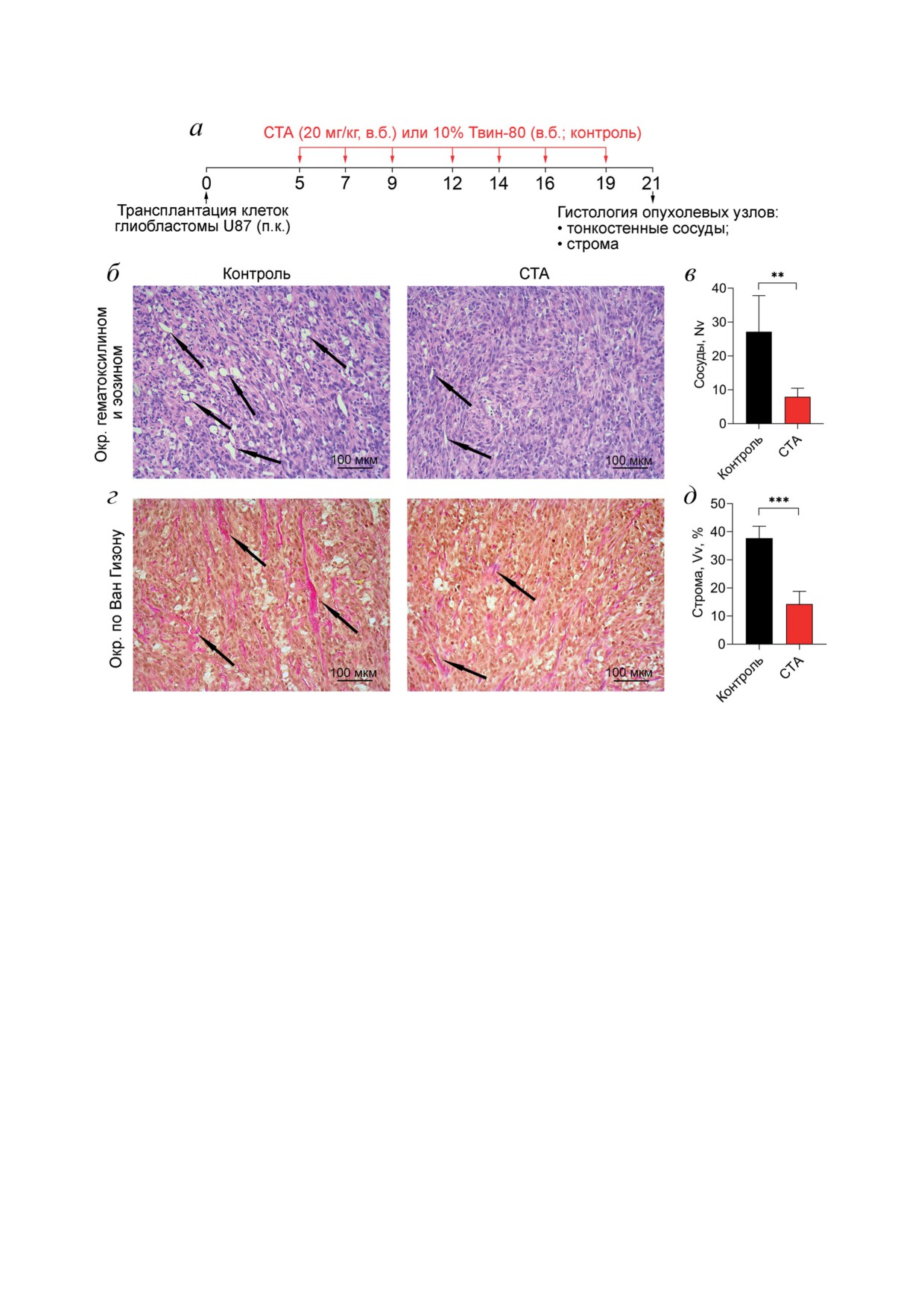
СТА блокирует развитие сети незрелых тон-
Данный эффект в сочетании с установлен-
костенных сосудов и снижает количество соеди-
ной нами ранее способностью СТА блокиро-
нительнотканных волокон в ткани МГБ. Ранее
вать васкулогенную мимикрию клеток U87
БИОХИМИЯ том 88 вып. 7 2023
АНТИГЛИОБЛАСТОМНЫЙ ПОТЕНЦИАЛ СТА
1247
Рис. 5. Эффект СТА на содержание тонкостенных кровеносных сосудов и эластических волокон в опухолевой ткани
на ксенографтной модели глиобластомы U87. а - Схема эксперимента. Мышам линии Nude подкожно транспланти-
ровали клетки глиобластоны U87. На 5-й день роста опухоли мышам вводили соединение СТА или Tween-80. Всего
было проведено 7 инъекций. На 21-й день роста опухоли мышей выводили из эксперимента. б - Гистологическое иссле-
дование опухолевых узлов. Окрашивание гематоксилином и эозином. Увеличение ×200. Черными стрелками указаны
тонкостенные кровеносные сосуды. в - Количественная оценка содержания тонкостенных кровеносных сосудов в
ткани глиобластомы U87. Nv - численная плотность структуры. г - Анализ соединительнотканных структур в опухо-
левых узлах. Окраска по Ван Гизону. Увеличение ×200. д - Количественная оценка содержания эластических волокон
в ткани глиобластомы U87. Vv - объемная плотность структуры (%)
in vitro [7] побудили нас к проведению деталь-
жит большое количество незрелых тонкостен-
ного исследования эффекта СТА на интенсив-
ных кровеносных сосудов, пронизывающих
ность ангиогенеза в ткани МГБ на вышеука-
опухоль (рис. 5. б), численная плотность кото-
занной мышиной модели.
рых достоверно в 3,4 раза снижалась при тера-
Учитывая тесную морфологическую связь
пии СТА по сравнению с контролем (рис. 5, в).
сосудистого и стромального компонентов опу-
Наблюдаемый эффект полностью согласуется
холи, нами была проанализирована представ-
с показанной нами ранее способностью СТА
ленность не только тонкостенных кровенос-
блокировать процесс формирования тубуляр-
ных сосудов, но также соединительнотканных
ных структур клетками U87 in vitro (васкуло-
волокон в опухолевой ткани мышей с транс-
генная мимикрия) [7]. Кроме того, гистологи-
плантированной подкожно глиобластомой U87,
ческое исследование выявило обширную сеть
получавших в.б. инъекции СТА в дозе 20 мг/кг
соединительнотканных волокон в опухолевой
или 10%-ный Tween-80, который использова-
ткани, представленность которой значительно
ли в качестве контроля. Схема эксперимента
обеднялась при введении исследуемого три-
представлена на рис. 5, а.
терпеноида (рис. 5, г): объемная плотность ок-
При гистологическом исследовании было
рашенных по Ван Гизону волокон в экспери-
выявлено, что ткань глиобластомы U87 содер- ментальной группе оказалась в 2,7 раза ниже
БИОХИМИЯ том 88 вып. 7 2023
1248
МАРКОВ и др.
по сравнению с контролем (рис. 5, д). Эти ре-
поэтому модулирование данной характери-
зультаты полностью согласуются с выявленной
стики опухолевых клеток можно рассматри-
ранее способностью СТА снижать содержание
вать в качестве перспективного направления
коллагеновых волокон в ткани глиобласто-
в противоопухолевой терапии [35]. Показан-
мы U87 на мышиной модели [7].
ная в настоящей работе способность СТА уве-
личивать межклеточную адгезию клеток U87
в трипсиновом тесте (рис. 2, а), с одной сто-
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
роны, согласуется со схожим эффектом азиа-
тиковой кислоты в отношении адгезивности
Учитывая высокий уровень злокачествен-
клеток рака легких человека А549 [36], однако,
ности МГБ, определяемой не только усилен-
с другой стороны, противоречит описанному в
ной клеточной пролиферацией, но также вы-
работе Tsai et al. [30] ингибирующему эффекту
раженной подвижностью опухолевых клеток,
RTA-404 (аналог СТА) в отношении адгези-
их инвазивностью и предрасположенностью
онных свойств клеток U87. Данное несоответ-
к развитию лекарственной устойчивости [2],
ствие можно объяснить значительными отли-
одним из многообещающих направлений в
чиями в используемых концентрациях три-
области создания новых классов антиглиобла-
терпеноидов: в нашей работе эффект СТА
стомных препаратов является поиск и синтез
был изучен в нетоксичном диапазоне кон-
соединений, способных оказывать не только
центраций (25-100 нМ) (рис. 2, а), тогда как
прямое токсическое воздействие на клетки
в работе Tsai et al. [30] влияние RTA-404 на
МГБ, но также блокировать их прометаста-
адгезивность клеток U87 исследовалось в кон-
тический фенотип [25]. В настоящей работе
центрации 500 нМ, вызывающей гибель при-
были продолжены исследования антиглио-
мерно 50% клеток, т.е. наблюдаемое снижение
бластомного потенциала СТА, относящегося
адгезивности клеток МГБ, стимулированных
к группе полусинтетических пентацикличе-
RTA-404, с высокой долей вероятности могло
ских тритерпеноидов, несущих в кольце А
быть вызвано их апоптозом. Помимо меж-
цианоеноновый фармакофорный заместитель
клеточной адгезии, СТА усиливал адгезив-
и обладающих выраженным комплексным
ность клеток U87 к коллагену, моделирую-
фармакологическим эффектом в отношении
щему внеклеточный матрикс (рис. 2, б), что
опухолевой прогрессии, воспаления и повре-
также свидетельствует в пользу его выражен-
ждения тканей [20, 26, 27]. В ходе проведен-
ного антиглиобластомного потенциала: ранее
ного исследования было установлено, что
Ramaswamy et al. [37] показали подобный эф-
СТА эффективно ингибирует рост клеток U87
фект в отношении адгезивности клеток U87 к
в 3D-культуре (рис. 1, в-е), причем данный
желатину при действии U0126, селективного
эффект был показан для цианоенон-содер-
ингибитора ERK1/2, обладающего выражен-
жащих тритерпеноидов впервые - в предыду-
ными антиглиобластомными свойствами.
щих работах антиглиобластомный потенциал
Наличие синергизма между цитотоксиче-
соединений данного ряда был оценен лишь
скими эффектами СТА и ТМЗ на модели кле-
на монослойных моделях [28-30], и только в
ток U87 (рис. 3, б), а также показанная нами
работах Zhou et al. [31], So et al. [32] и Murad
ранее способность СТА эффективно накап-
et al. [33] был установлен блокирующий эф-
ливаться в тканях головного мозга мышей [7]
фект CDDO-Me и CDDO-Im на рост сферои-
говорят о потенциальной возможности вве-
дов клеток рака молочной железы и простаты
дения исследуемого тритерпеноида в совре-
человека. Выявленная способность СТА инги-
менные схемы химиотерапии МГБ, однако
бировать рост клеток U87 в сфероидах хорошо
вызванное им усиление активности ALDH в
согласуется с его супрессивным эффектом на
клетках U87 (рис. 3, в) свидетельствует о необ-
рост опухолевых узлов на мышиной гетеро-
ходимости тщательного подбора оптималь-
топической ксенографтной модели глиобла-
ных дозировок и схем введения СТА. Актива-
стомы U87, показанным нами ранее [7].
ция ALDH в опухолевых клетках связана с их
Помимо подавления сфероидного роста,
цитопротекторным ответом на ксенобиотики,
наблюдаемый эффект СТА в отношении повы-
электрофильный и окислительный стресс [38],
шения адгезивности клеток U87 (рис. 2) также
являясь также ключевым маркером опухоле-
подтверждает его выраженный антиглиобла-
вых стволовых клеток (ОСК) [39]. Учитывая
стомный потенциал. Известно, что слабый уро-
наличие в структуре СТА электрофильного
вень межклеточной адгезии опухолевых кле-
акцептора Михаэля (цианоеноновая группа),
ток связан с их мезенхимальным фенотипом и
его стимулирующий эффект на продукцию
играет важную роль в метастазировании [34],
активных форм кислорода в клетках U87,
БИОХИМИЯ том 88 вып. 7 2023
АНТИГЛИОБЛАСТОМНЫЙ ПОТЕНЦИАЛ СТА
1249
показанный нами ранее [7], и способность
пеноидов эффективно ингибировать сигналь-
ингибировать рост сфероидов клеток U87
ные пути EGFR, ERBB2 и AKT1 в различ-
(рис. 1, в-е), являющихся важным источни-
ных опухолевых клетках [20, 45], в том числе
ком ОСК [40], мы предполагаем, что наблю-
в клетках глиобластомы [30, 46]. Более того,
даемая активация ALDH под действием СТА
Liu et al. [47] экспериментально доказали пря-
(рис. 3, в) все же является следствием компен-
мую блокировку киназной активности AKT1
сационных механизмов в клетках МГБ в ответ
аналогом СТА CDDO-Me, Kim et al. [48] вы-
на ксенобиотический стресс, а не маркером уси-
явили способность CDDO-Me напрямую взаи-
ления их стволовых свойств, что требует даль-
модействовать с ERBB2, а в нашей недавней
нейшего экспериментального подтверждения.
работе было установлено, что CDDO-Im спо-
Проведенные гистологические исследо-
собен взаимодействовать с активным цен-
вания срезов опухолевой ткани глиобласто-
тром EGFR с энергией связывания ниже энер-
мы U87 показали, что СТА способен воздей-
гии связывания известного ингибитора EGFR
ствовать не только напрямую на опухолевые
эрлотиниба
[49]. Учитывая ключевую роль
клетки, но и на опухолевое микроокружение,
EGFR, ERBB2 и AKT1 в регуляции прогрессии
подавляя формирование в ней сети незрелых
МГБ [50-52], в том числе в приобретении клет-
тонкостенных кровеносных сосудов (рис. 5, в),
ками МГБ мезенхимального фенотипа и ство-
хаотичная разветвленная структура и высокая
ловых свойств [53, 54], выявленная с помощью
проницаемость которых формирует в опухо-
in silico подходов способность СТА формировать
левой ткани очаги гипоксии, способствующие
комплексы с данными белками требует даль-
появлению высокозлокачественных клеток
нейшего экспериментального подтверждения.
МГБ мезенхимального фенотипа, которые и
определяют диффузный рост глиобластом [41].
Наблюдаемый супрессорный эффект СТА в от-
ЗАКЛЮЧЕНИЕ
ношении тонкостенных кровеносных сосудов,
стенки которых представлены не только эндо-
В результате проведенного исследования
телиоцитами, но и опухолевыми клетками [42],
было установлено, что СТА обладает выра-
согласуется с его блокирующим воздействием
женным антиглиобластомным потенциалом,
на формирование тубулярных структур из кле-
опосредуемым как его прямым модулирующим
ток U87 (васкулогенная мимикрия) in vitro, по-
воздействием на клетки МГБ, включая бло-
казанный нами ранее [7]. Полученные данные
кировку сфероидного роста, усиление адге-
в сочетании с выявленной ранее способностью
зивности и синергичность цитотоксического
СТА усиливать экспрессию маркеров зрелых
эффекта с ТМЗ, так и нормализацией ряда
сосудов α-SMA и CD31 в ткани глиобласто-
параметров опухолевого микроокружения. По-
мы U87 на ксенографтной модели in vivo [7]
лученные данные свидетельствуют о необходи-
свидетельствуют о нормализации патологи-
мости дальнейшего изучения его эффектов на
ческого ангиогенеза в опухолевой ткани, что
приобретение клетками МГБ мезенхимального
может способствовать сокращению объемов
фенотипа и стволовых свойств и позволяют
гипоксических зон в ткани МГБ и, как резуль-
предложить СТА в качестве перспективного
тат, усилению ее чувствительности к химио-
мультитаргетного антиглиобластомного канди-
и радиотерапии [43]. Наряду с влиянием на
дата.
рост тонкостенных кровеносных сосудов, СТА
также снижал содержание в опухолевом мик-
Вклад авторов. А.В. Марков
- концеп-
роокружении соединительнотканных волокон
ция и руководство работой; К.В. Одаренко,
(рис. 5, г и д), высокая представленность кото-
А.В. Сенькова, А.А. Ильина
- проведение
рых формирует барьер для химиопрепаратов,
экспериментов; А.В. Марков, А.В. Сенько-
питательных веществ, кислорода и иммунных
ва - обсуждение результатов исследования;
клеток (иммунные ловушки), который сни-
А.В. Марков, К.В. Одаренко, А.В. Сенькова -
жает чувствительность клеток МГБ к химио-,
написание текста; М.А. Зенкова - редакти-
радио- и иммунотерапии [44].
рование текста статьи и общее руководство
Согласно данным компьютерного моде-
исследованиями.
лирования, наблюдаемые антиглиобластомные
Финансирование. Работа выполнена при
эффекты СТА могут быть основаны на его
финансовой поддержке Российского науч-
прямом ингибирующем воздействии на EGFR,
ного фонда (грант
ERBB2 и AKT1 (рис. 4, б и в). Накопленные к
rscf.ru/project/19-74-30011/) и базового бюд-
настоящему моменту данные подтверждают
жетного финансирования ИХБФМ СО РАН
способность цианоенон-содержащих тритер-
№ 121031300044-5.
14
БИОХИМИЯ том 88 вып. 7 2023
1250
МАРКОВ и др.
Благодарности. Авторы выражают осо-
ветствии с нормами обращения с животными,
бую признательность с.н.с., к.х.н. Саломати-
изложенными в Директиве Европейского Пар-
ной О.В. (НИОХ СО РАН) за предоставленный
ламента и Совета ЕС 2010/63/EU от 22 сентяб-
СТА и ведущему инженеру Владимировой А.В.
ря 2010 г. и «Правилам проведения работ с
(ИХБФМ СО РАН) за помощь в поддержании
использованием экспериментальных живот-
клеточных культур.
ных» (приказ Минздрава СССР № 755 от 12 ав-
Конфликт интересов. Авторы заявляют об
густа 1977 г.). Все эксперименты на животных
отсутствии конфликта интересов.
были одобрены межинститутской Комиссией
Соблюдение этических норм. Все экспери-
по биоэтике ИЦиГ СО РАН (протокол № 56
менты на мышиной модели проведены в соот- от 10.08.2019 г.).
СПИСОК ЛИТЕРАТУРЫ
1.
Sanati, M., Binabaj, M. M., Ahmadi, S. S.,
biological activities in vitro and in vivo, Pharmaceuti-
Aminyavari, S., Javid, H., Mollazadeh, H., Bibak, B.,
cals, 15, 603, doi: 10.3390/ph15050603.
Mohtashami, E., Jamialahmadi, T., Afshari, A. R., and
8.
Ianevski, A., Giri, A. K., and Aittokallio, T. (2020)
Sahebkar, A. (2022) Recent advances in glioblastoma
SynergyFinder 2.0: visual analytics of multi-drug com-
multiforme therapy: a focus on autophagy regulation,
bination synergies, Nucleic Acids Res., 48, W488-W493,
Biomed. Pharmacother., 155, 113740, doi: 10.1016/
doi: 10.1093/nar/gkaa216.
j.biopha.2022.113740.
9.
Awale, M. and Reymond, J. L. (2019) Polyphar-
2.
Torrisi, F., Alberghina, C., D’Aprile, S., Pavone,
macology browser PPB2: target prediction com-
A. M., Longhitano, L., Giallongo, S., Tibullo, D.,
bining nearest neighbors with machine learning,
Di Rosa, M., Zappalà, A., Cammarata, F. P., Russo, G.,
J. Chem. Inf. Model., 59, 10-17, doi: 10.1021/acs.jcim.
Ippolito, M., Cuttone, G., Li Volti, G., Vicario, N.,
8b00524.
and Parenti, R. (2022) The hallmarks of glioblastoma:
10.
Daina, A., Michielin, O., and Zoete, V.
(2019)
heterogeneity, intercellular crosstalk and molecular
SwissTargetPrediction: updated data and new features
signature of invasiveness and progression, Biomedicines,
for efficient prediction of protein targets of small
10, 806, doi: 10.3390/biomedicines10040806.
molecules, Nucleic Acids Res.,
47, W357-W364,
3.
Cha, G. D., Kang, T., Baik, S., Kim, D., Choi, S. H.,
doi: 10.1093/nar/gkz382.
Hyeon, T., and Kim, D.-H. (2020) Advances in drug
11.
Szklarczyk, D., Franceschini, A., Wyder, S., Fors-
delivery technology for the treatment of glioblastoma
lund, K., Heller, D., Huerta-Cepas, J., Simonovic, M.,
multiforme, J. Control. Release,
328,
350-367,
Roth, A., Santos, A., Tsafou, K. P., Kuhn, M.,
doi: 10.1016/j.jconrel.2020.09.002.
Bork, P., Jensen, L. J., and Von Mering, C. (2015)
4.
Qazi, M. A., Vora, P., Venugopal, C., Sidhu, S. S.,
STRING v10: protein-protein interaction networks,
Moffat, J., Swanton, C., and Singh, S. K. (2017)
integrated over the tree of life, Nucleic Acids Res., 43,
Intratumoral heterogeneity: pathways to treatment
D447-D452, doi: 10.1093/nar/gku1003.
resistance and relapse in human glioblastoma, Ann.
12.
Chin, C.-H., Chen, S.-H., Wu, H.-H., Ho, C.-W.,
Oncol., 28, 1448-1456, doi: 10.1093/annonc/mdx169.
Ko, M.-T., and Lin, C.-Y. (2014) cytoHubba: iden-
5.
Sestito, S., Runfola, M., Tonelli, M., Chiellini, G.,
tifying hub objects and sub-networks from complex
and Rapposelli, S. (2018) New multitarget approach-
interactome, BMC Syst. Biol., 8, S11, doi: 10.1186/
es in the war against glioblastoma: a mini-per-
1752-0509-8-S4-S11.
spective, Front. Pharmacol., 9, 874, doi: 10.3389/
13.
Trott, O., and Olson, A. J. (2010) AutoDock Vina:
fphar.2018.00874.
improving the speed and accuracy of docking with
6.
Behl, T., Sharma, A., Sharma, L., Sehgal, A., Singh, S.,
a new scoring function, efficient optimization, and
Sharma, N., Zengin, G., Bungau, S., Toma, M. M.,
multithreading, J. Comput. Chem.,
31,
455-461,
Gitea, D., Babes, E. E., Judea Pusta, C. T., and
doi: 10.1002/jcc.21334.
Bumbu, A. G. (2021) Current perspective on the
14.
Suhovskih, A. V., Kazanskaya, G. M., Volkov, A. M.,
natural compounds and drug delivery techniques in
Tsidulko, A. Y., Aidagulova, S. V., and Grigorieva, E. V.
glioblastoma multiforme, Cancers (Basel), 13, 2765,
(2019) Suitability of RNALater solution as a tissue-
doi: 10.3390/cancers13112765.
preserving reagent for immunohistochemical analysis,
7.
Markov, A. V., Ilyina, A. A., Salomatina, O. V.,
Histochem. Cell Biol., 152, 239-247, doi: 10.1007/
Sen’kova, A. V., Okhina, A. A., Rogachev, A. D.,
s00418-019-01799-z.
Salakhutdinov, N. F., and Zenkova, M. A. (2022)
15.
Joseph, J. V., Balasubramaniyan, V., Walenkamp, A.,
Novel soloxolone amides as potent anti-glioblastoma
and Kruyt, F. A. E. (2013) TGF-β as a therapeutic
candidates: design, synthesis, in silico analysis and
target in high grade gliomas - promises and challenges,
БИОХИМИЯ том 88 вып. 7 2023
АНТИГЛИОБЛАСТОМНЫЙ ПОТЕНЦИАЛ СТА
1251
Biochem. Pharmacol.,
85,
478-485, doi:
10.1016/
prioritisation, Nucleic Acids Res., 49, D1302-D1310,
j.bcp.2012.11.005.
doi: 10.1093/nar/gkaa1027.
16.
Peñuelas, S., Anido, J., Prieto-Sánchez, R. M.,
25.
Bazan, N. G., Reid, M. M., Flores, V. A. C., Gallo,
Folch, G., Barba, I., Cuartas, I., García-Dorado, D.,
J. E., Lewis, W., and Belayev, L. (2021) Multi-
Poca, M. A., Sahuquillo, J., Baselga, J., and Seoane, J.
prong control of glioblastoma multiforme invasive-
(2009) TGF-β increases glioma-initiating cell self-
ness: blockade of pro-inflammatory signaling, anti-
renewal through the induction of LIF in human
angiogenesis, and homeostasis restoration, Cancer
glioblastoma, Cancer Cell, 15, 315-327, doi: 10.1016/
Metastasis Rev., 40, 643-647, doi: 10.1007/s10555-
j.ccr.2009.02.011.
021-09987-x.
17.
Beri, P., Popravko, A., Yeoman, B., Kumar, A.,
26.
Liby, K. T., and Sporn, M. B. (2012) Synthetic ole-
Chen, K., Hodzic, E., Chiang, A., Banisadr, A.,
anane triterpenoids: multifunctional drugs with
Placone, J. K., Carter, H., Fraley, S. I., Katira, P.,
a broad range of applications for prevention and
and Engler, A. J. (2020) Cell adhesiveness serves
treatment of chronic disease, Pharmacol. Rev., 64,
as a biophysical marker for metastatic potential,
972-1003, doi: 10.1124/pr.111.004846.
Cancer Res., 80, 901-911, doi: 10.1158/0008-5472.
27.
Sen’kova, A. V., Savin, I. A., Odarenko, K. V.,
CAN-19-1794.
Salomatina, O. V., Salakhutdinov, N. F., Zenkova,
18.
Beltzig, L., Schwarzenbach, C., Leukel, P., Frauen-
M. A., and Markov, A. V. (2023) Protective effect of
knecht, K. B. M., Sommer, C., Tancredi, A., Hegi,
soloxolone derivatives in carrageenan- and LPS-driv-
M. E., Christmann, M., and Kaina, B. (2022) Se-
en acute inflammation: Pharmacological profiling
nescence is the main trait induced by temozolo-
and their effects on key inflammation-related process-
mide in glioblastoma cells, Cancers (Basel), 14, 2233,
es, Biomed. Pharmacother., 159, 114231, doi: 10.1016/
doi: 10.3390/cancers14092233.
j.biopha.2023.114231.
19.
Poturnajova, M., Kozovska, Z., and Matuskova, M.
28.
Tsai, T.-H., Lieu, A.-S., Huang, T.-Y., Kwan, A.-L.,
(2021) Aldehyde dehydrogenase 1A1 and 1A3 iso-
Lin, C.-L., and Hsu, Y.-C. (2021) Induction of mitosis
forms - mechanism of activation and regulation
delay and apoptosis by CDDO-TFEA in glioblastoma
in cancer, Cell. Signal., 87, 110120, doi: 10.1016/
multiforme, Front. Pharmacol.,
12,
756228, doi:
j.cellsig.2021.110120.
10.3389/fphar.2021.756228.
20.
Markov, A. V., Zenkova, M. A., and Logashenko, E. B.
29.
Tsai, T. H., Hsu, Y. C., Lieu, A. S., Huang, T. Y.,
(2017) Modulation of Tumour-related signaling path-
Kwan, A. L., and Lin, C. L. (2021) RTA404, an
ways by natural pentacyclic triterpenoids and their
activator of Nrf2, activates the checkpoint kinases
semisynthetic derivatives, Curr. Med. Chem., 24, 1277-
and induces apoptosis through intrinsic apoptotic
1320, doi: 10.2174/0929867324666170112115313.
pathway in malignant glioma, J. Clin. Med., 10,
21.
McDowell, K. A., Riggins, G. J., and Gallia, G. L.
4805, doi: 10.3390/jcm10214805.
(2011) Targeting the AKT pathway in glioblasto-
30.
Tsai, T.-H., Lieu, A.-S., Wang, Y.-W., Yang, S.-F.,
ma, Curr. Pharm. Des., 17, 2411-2420, doi: 10.2174/
Hsu, Y.-C., and Lin, C.-L. (2021) Therapeutic poten-
138161211797249224.
tial of RTA 404 in human brain malignant glioma cell
22.
Mure, H., Matsuzaki, K., Kitazato, K. T., Mizo-
lines via cell cycle arrest via p21/AKT signaling, Biomed
buchi, Y., Kuwayama, K., Kageji, T., and Nagahiro, S.
Res. Int., 2021, 5552226, doi: 10.1155/2021/5552226.
(2010) Akt2 and Akt3 play a pivotal role in malignant
31.
Zhou, L., Wang, Z., Yu, S., Xiong, Y., Fan, J., Lyu, Y.,
gliomas, Neuro. Oncol., 12, 221-232, doi: 10.1093/
Su, Z., Song, J., Liu, S., Sun, Q., and Lu, D.
neuonc/nop026.
(2020) CDDO-Me elicits anti-breast cancer activity
23.
Yoon, H., Kim, D. J., Ahn, E. H., Gellert, G. C., Shay,
by targeting LRP6 and FZD7 receptor complex,
J. W., Ahn, C.-H., and Lee, Y. B. (2009) Antitumor
J. Pharmacol. Exp. Ther., 373, 149-159, doi: 10.1124/
activity of a novel antisense oligonucleotide against
jpet.119.263434.
Akt1, J. Cell. Biochem., 108, 832-838, doi: 10.1002/
32.
So, J. Y., Lin, J. J., Wahler, J., Liby, K. T., Sporn,
jcb.22311.
M. B., and Suh, N. (2014) A synthetic triterpenoid
24.
Ochoa, D., Hercules, A., Carmona, M., Suveges, D.,
CDDO-Im inhibits tumorsphere formation by regu-
Gonzalez-Uriarte, A., Malangone, C., Miranda, A.,
lating stem cell signaling pathways in triple-negative
Fumis, L., Carvalho-Silva, D., Spitzer, M., Baker, J.,
breast cancer, PLoS One, 9, e107616, doi: 10.1371/
Ferrer, J., Raies, A., Razuvayevskaya, O., Faulcon-
journal.pone.0107616.
bridge, A., Petsalaki, E., Mutowo, P., Machlitt-
33.
Murad, H. Y., Chandra, P. K., Kelly, C. A., Khurana, N.,
Northen, S., Peat, G., McAuley, E., Ong, C. K.,
Yu, H., Bortz, E. P., Hong, S. N., Mondal, D.,
Mountjoy, E., Ghoussaini, M., Pierleoni, A., Papa, E.,
and Khismatullin, D. B. (2022) Pre-exposure to
Pignatelli, M., Koscielny, G., Karim, M., Schwartzen-
stress-inducing agents increase the anticancer effi-
truber, J., Hulcoop, D. G., Dunham, I., and
cacy of focused ultrasound against aggressive pros-
McDonagh, E. M. (2021) Open Targets Platform:
tate cancer cells, Antioxidants, 11, 341, doi: 10.3390/
supporting systematic drug-target identification and
antiox11020341.
БИОХИМИЯ том 88 вып. 7 2023
14*
1252
МАРКОВ и др.
34.
Le Bras, G. F., Taubenslag, K. J., and Andl, C. D.
M. C., and Andreeff, M. (2006) Synthetic triterpenoid
(2012) The regulation of cell-cell adhesion during
2-cyano-3,12-dioxooleana-1,9-dien-28-oic acid in-
epithelial-mesenchymal transition, motility and tumor
duces growth arrest in HER2-overexpressing breast
progression, Cell Adh. Migr., 6, 365-373, doi: 10.4161/
cancer cells, Mol. Cancer Ther., 5, 317-328, doi:
cam.21326.
10.1158/1535-7163.MCT-05-0350.
35.
Janiszewska, M., Primi, M. C., and Izard, T. (2020)
46.
Gao, X., Deeb, D., Jiang, H., Liu, Y., Dulchavsky,
Cell adhesion in cancer: beyond the migration of single
S. A., and Gautam, S. C. (2007) Synthetic triter-
cells, J. Biol. Chem., 295, 2495-2505, doi: 10.1074/
penoids inhibit growth and induce apoptosis in human
jbc.REV119.007759.
glioblastoma and neuroblastoma cells through inhi-
36.
Cui, Q., Ren, J., Zhou, Q., Yang, Q., and Li, B. (2019)
bition of prosurvival Akt, NF-κB and Notch1 signal-
Effect of asiatic acid on epithelial-mesenchymal tran-
ing, J. Neurooncol., 84, 147-157, doi: 10.1007/s11060-
sition of human alveolar epithelium A549 cells induced
007-9364-9.
by TGF-β1, Oncol. Lett., 17, 4285-4292, doi: 10.3892/
47.
Liu, Y., Gao, X., Deeb, D., and Gautam, S. C.
ol.2019.10140.
(2012) Oleanane triterpenoid CDDO-Me inhibits
37.
Ramaswamy, P., Nanjaiah, N. D., and Borkotokey, M.
Akt activity without affecting PDK1 kinase or PP2A
(2019) Role of MEK-ERK signaling mediated adhe-
phosphatase activity in cancer cells, Biochem. Bio-
sion of glioma cells to extracellular matrix: possible
phys. Res. Commun., 417, 570-575, doi: 10.1016/
implication on migration and proliferation, Ann. Neu-
j.bbrc.2011.12.007.
rosci., 26, 52-56, doi: 10.5214/ans.0972.7531.260203.
48.
Kim, E.-H. H., Deng, C., Sporn, M. B., Royce, D. B.,
38.
Singh, S., Brocker, C., Koppaka, V., Chen, Y.,
Risingsong, R., Williams, C. R., and Liby, K. T. (2012)
Jackson, B. C., Matsumoto, A., Thompson, D. C.,
CDDO-methyl ester delays breast cancer develop-
and Vasiliou, V. (2013) Aldehyde dehydrogenases in
ment in Brca1-mutated mice, Cancer Prev. Res., 5,
cellular responses to oxidative/electrophilicstress,
89-97, doi: 10.1158/1940-6207.CAPR-11-0359.
Free Radic. Biol. Med., 56, 89-101, doi: 10.1016/
49.
Markov, A. V., Odarenko, K. V., Ilyina, A. A., and
j.freeradbiomed.2012.11.010.
Zenkova, M. A. (2022) Uncovering the anti-angio-
39.
Rodriguez, S. M., Staicu, G.-A., Sevastre, A.-S.,
genic effect of semisynthetic triterpenoid CDDO-Im
Baloi, C., Ciubotaru, V., Dricu, A., and Tataranu,
on HUVECs by an integrated network pharmacol-
L. G. (2022) Glioblastoma stem cells - useful tools
ogy approach, Comput. Biol. Med.,
141,
105034,
in the battle against cancer, Int. J. Mol. Sci., 23, 4602,
doi: 10.1016/J.COMPBIOMED.2021.105034.
doi: 10.3390/ijms23094602.
50.
Westphal, M., Maire, C. L., and Lamszus, K. (2017)
40.
Wang, X., Zhou, W., Li, X., Ren, J., Ji, G., Du, J.,
EGFR as a target for glioblastoma treatment: an
Tian, W., Liu, Q., and Hao, A. (2020) Graphene ox-
unfulfilled promise, CNS Drugs,
31,
723-735,
ide suppresses the growth and malignancy of glio-
doi: 10.1007/s40263-017-0456-6.
blastoma stem cell-like spheroids via epigenetic
51.
Majewska, E., and Szeliga, M. (2017) AKT/GSK3β
mechanisms, J. Transl. Med., 18, 200, doi: 10.1186/
signaling in glioblastoma, Neurochem. Res.,
42,
s12967-020-02359-z.
918-924, doi: 10.1007/s11064-016-2044-4.
41.
Angara, K., Borin, T. F., and Arbab, A. S. (2017)
52.
Vitanza, N. A., Johnson, A. J., Wilson, A. L.,
Vascular mimicry: a novel neovascularization mech-
Brown, C., Yokoyama, J. K., Künkele, A., Chang,
anism driving anti-angiogenic therapy (AAT) resis-
C. A., Rawlings-Rhea, S., Huang, W., Seidel, K.,
tance in glioblastoma, Transl. Oncol., 10, 650-660,
Albert, C. M., Pinto, N., Gust, J., Finn, L. S.,
doi: 10.1016/j.tranon.2017.04.007.
Ojemann, J. G., Wright, J., Orentas, R. J., Baldwin, M.,
42.
Viallard, C., and Larrivée, B. (2017) Tumor angiogen-
Gardner, R. A., Jensen, M. C., and Park, J. R. (2021)
esis and vascular normalization: alternative therapeu-
Locoregional infusion of HER2-specific CAR T cells
tic targets, Angiogenesis, 20, 409-426, doi: 10.1007/
in children and young adults with recurrent or refrac-
s10456-017-9562-9.
tory CNS tumors: an interim analysis, Nat. Med., 27,
43.
Zheng, R., Li, F., Li, F., and Gong, A. (2021) Target-
1544-1552, doi: 10.1038/s41591-021-01404-8.
ing tumor vascularization: promising strategies for vas-
53.
Takashima, Y., Kawaguchi, A., and Yamanaka, R.
cular normalization, J. Cancer Res. Clin. Oncol., 147,
(2019) Promising prognosis marker candidates on
2489-2505, doi: 10.1007/s00432-021-03701-8.
the status of epithelial-mesenchymal transition
44.
Mohiuddin, E. and Wakimoto, H. (2021) Extracellu-
and glioma stem cells in glioblastoma, Cells, 8, 1312,
lar matrix in glioblastoma: opportunities for emerg-
doi: 10.3390/cells8111312.
ing therapeutic approaches, Am. J. Cancer Res.,
54.
Barzegar Behrooz, A., Talaie, Z., Jusheghani, F.,
11, 3742-3754.
Łos, M. J., Klonisch, T., and Ghavami, S. (2022)
45.
Konopleva, M., Zhang, W., Shi, Y. X., McQueen, T.,
Wnt and PI3K/Akt/mTOR survival pathways as
Tsao, T., Abdelrahim, M., Munsell, M. F., Johan-
therapeutic targets in glioblastoma, Int. J. Mol. Sci.,
sen, M., Yu, D., Madden, T., Safe, S. H., Hung,
23, 1353, doi: 10.3390/ijms23031353.
БИОХИМИЯ том 88 вып. 7 2023
АНТИГЛИОБЛАСТОМНЫЙ ПОТЕНЦИАЛ СТА
1253
EVALUATION OF THE ANTITUMOR POTENTIAL OF SOLOXOLONE
TRYPTAMIDE AGAINST GLIOBLASTOMA MULTIFORME
USING in silico, in vitro AND in vivo APPROACHES
A. V. Markov*, K. V. Odarenko, A. V. Sen’kova, A. A. Ilyina, and M. A. Zenkova
Institute of Chemical Biology and Fundamental Medicine, Siberian Branch of the Russian Academy of Sciences,
630090 Novosibirsk, Russia; e-mail: andmrkv@gmail.com
Glioblastoma multiforme (GBM) is a highly aggressive neoplasia characterized by uncontrollable diffu-
sive growth, development of resistance to chemo- and radiotherapy and a high rate of recurrence that leads
to low survival of patients with GBM. Given the number of signaling pathways regulating GBM patho-
genesis, the development of novel anti-glioblastoma compounds based on multitarget natural metabolites
is a promising direction for the study. In the current work, the antitumor potential of semisynthetic triter-
penoid soloxolone tryptamide (STA) against human glioblastoma U87 cells was explored. It was found that
STA effectively blocked the growth of U87 cells in 2D and 3D culture systems, enhanced the adhesive-
ness of tumor cells and displayed synergistic cytotoxicity with temozolomide. Performed in silico analysis
revealed that the pronounced anti-glioblastoma activity of STA can be explained by its direct interaction
with EGFR, ERBB2 and AKT1, which play an important role in the regulation of GBM malignancy. Along
with observed direct modulatory action of STA on U87 cells, explored compound had a normalizing effect
on the tumor microenvironment in murine heterotopic U87 xenograft model, suppressing the development
of immature blood vessels and the production of elastin in tumor tissue. Taken together, obtained results
clearly demonstrate that STA can be considered a novel promising antitumor candidate for GMB treatment.
Keywords: glioblastoma multiforme, cyanoenone pharmacophore, pentacyclic triterpenoids, spheroids, angiogenesis,
synergistic effect, molecular docking, gene network, tumor microenvironment
БИОХИМИЯ том 88 вып. 7 2023