БИОХИМИЯ, 2023, том 88, вып. 8, с. 1302 - 1312
УДК 57.044
АГОНИСТ КОНСТИТУТИВНОГО АНДРОСТАНОВОГО РЕЦЕПТОРА
ИНИЦИИРУЕТ МЕТАБОЛИЧЕСКУЮ АКТИВНОСТЬ,
НЕОБХОДИМУЮ ДЛЯ ПРОЛИФЕРАЦИИ ГЕПАТОЦИТОВ
© 2023 М.Э. Мазин1,2, А.М. Перевалова1, А.А. Ярушкин2, Ю.А. Пустыльняк1,
А.Д. Рогачев1, Е.А. Прокопьева1,2, Л.Ф. Гуляева1,2, В.О. Пустыльняк1,2*
1 Новосибирский национальный исследовательский государственный университет,
630090 Новосибирск, Россия; электронная почта: pustylnyak@post.nsu.ru
2 Федеральный исследовательский центр фундаментальной и трансляционной медицины,
630117 Новосибирск, Россия
Поступила в редакцию 10.04.2023
После доработки 22.05.2023
Принята к публикации 24.05.2023
Активация конститутивного андростанового рецептора (CAR, NR1I3) химическими соединениями
вызывает гиперплазию печени у грызунов. Чаще всего для изучения химически индуцированной
гиперплазии печени и пролиферации гепатоцитов in vivo используется 1,4-бис[2-(3,5-дихлорпири-
дилокси)] бензол (TCPOBOP), агонист мышиного CAR. TCPOBOP является мощным химическим
митогеном, который вызывает гиперплазию печени мышей. В последние годы накоплено много
данных о транскрипционных изменениях, характеризующих TCPOBOP-индуцированную проли-
ферацию гепатоцитов. Однако данных о метаболических потребностях гепатоцитов, делящихся
при воздействии ксенобиотика, немного. В настоящем исследовании мы использовали техноло-
гию высокоэффективной жидкостной хроматографии и масс-спектрометрии в сочетании со ста-
тистическим анализом для описания изменения профиля метаболитов малых биомолекул, чтобы
выявить ключевые метаболические изменения в печени самцов мышей после введения TCPOBOP.
Анализ метаболитов в печени мышей позволил обнаружить биохимические процессы, активация
которых осуществляется при воздействии химического митогена TCPOBOP. Так, сравнение мета-
боломных профилей показало, что воздействие ТСРОВОР приводит к изменению биохимических
процессов, которые имеют отношение к метаболизму нуклеотидов, аминокислот и энергетических
субстратов. Наши результаты позволяют сделать вывод о том, что агонист CAR инициирует внут-
риклеточную программу, которая способствует глобальной скоординированной метаболической
активности, необходимой для пролиферации гепатоцитов.
КЛЮЧЕВЫЕ СЛОВА: конститутивный андростановый рецептор, TCPOBOP, печень, гепатоцит, метабо-
ломика.
DOI: 10.31857/S032097252308002X, EDN: IHUBDC
ВВЕДЕНИЕ
широким набором функций в гепатоцитах [2].
Так, например, активация CAR значительно
Конститутивный андростановый рецеп-
усиливает пролиферацию гепатоцитов с по-
тор (CAR, NR1I3) является членом суперсе-
следующей гиперплазией печени мышей [3].
мейства ядерных рецепторов. Первоначально
Более того, показано, что активация CAR
CAR был охарактеризован как ксеносенсор,
химическими соединениями вызывает не
который в ответ на воздействие ксенобиотиков
только гиперплазию печени у грызунов, но
индуцирует их метаболизм и элиминацию [1].
и может значительно улучшить регенерацию
Маркерными генами-мишенями для CAR яв-
печени грызунов после экстремальных резек-
ляются гены, кодирующие цитохром Р450
ций (более 80% паренхимы), тем самым пред-
подсемейства 2В (CYP2B). Впоследствии было
отвращая возникновение печеночной недос-
показано, что CAR обладает гораздо более таточности из-за дефицита ткани органа [4].
Принятые сокращения: CAR - конститутивный андростановый рецептор; CYP2B - ген, кодирующий цитохром Р450
подсемейства 2В; TCPOBOP - 1,4-бис[2-(3,5-дихлорпиридилокси)] бензол.
* Адресат для корреспонденции.
1302
АГОНИСТ CAR ИЗМЕНЯЕТ МЕТАБОЛОМ В ПЕЧЕНИ
1303
Поэтому расширение научных данных, опи-
бирск, Россия). Животные прошли пятиднев-
сывающих молекулярные и биохимические
ный период акклиматизации со свободным
процессы, сопряженные с активацией проли-
доступом к пище и воде. Животным одно-
ферации гепатоцитов при участии CAR, может
кратно внутрибрюшинно вводили TCPOBOP
позволить сделать выводы о возможности кли-
(3 мг/кг массы тела), растворенный в пище-
нического использования агонистов этого ре-
вом рафинированном подсолнечном масле.
цептора для профилактики печеночной недос-
Контрольные животные получали равный
таточности при резекции или трансплантации
объем (100 мкл) подсолнечного масла. Мышей
органа.
декапитировали через
72 ч после введения
Большинство исследований in vivo, посвя-
TCPOBOP (n = 12) или подсолнечного масла
щенных CAR-опосредованной пролиферации
(n = 12). При заборе органов осуществляли
гепатоцитов, были выполнены с использова-
взвешивание печени, и с учетом массы жи-
нием 1,4-бис[2-(3,5-дихлорпиридилокси)] бен-
вотного вычисляли индекс ее относительной
зола (TCPOBOP), агониста мышиного CAR.
массы («масса печени/масса тела»). Значение
TCPOBOP является мощным химическим ми-
индекса относительной массы печени одного
тогеном у мышей, который вызывает быструю
из контрольных животных принималось за 1.
гиперплазию печени [3]. В последние годы на-
Гистологическое исследование тканей пече-
коплено много данных о транскрипционных
ни. Образцы печени фиксировали в 10%-ном
изменениях, характеризующих TCPOBOP-ин-
растворе нейтрального формалина, затем
дуцированную пролиферацию гепатоцитов [5,
обезвоживали по стандартной методике [8]
6]. Однако данных о метаболических измене-
и заливали в парафин. Парафиновые срезы
ниях, происходящих в гепатоцитах, делящих-
(4-5 мкм) получали с помощью ротацион-
ся при воздействии ксенобиотиков, немного.
ного микротома HM 340E («CarlZeiss», Гер-
Печень является одним из наиболее метабо-
мания). Срезы, полученные из ткани печени
лически активных органов в организме и иг-
мышей, окрашивали гематоксилином и эози-
рает важную роль в регулировании различных
ном (H&E). Исследование гистологических
метаболических процессов. Метаболическое
препаратов проводили на микроскопе Axio-
ремоделирование клеток необходимо для кле-
ImagerA1 («CarlZeiss»), используя программное
точного деления, и гепатоциты оснащены гиб-
обеспечение AxioVision (rel. 4.12) («CarlZeiss»).
ким метаболическим механизмом, способным
Определение маркера пролиферации. Мар-
быстро адаптироваться к изменениям во время
кер пролиферации Ki67 в образцах печени
пролиферации [7]. Ранее уже было показано,
определяли с использованием иммуногистохи-
что воздействие TCPOBOP модулирует метабо-
мического метода и кроличьего поликлональ-
лизм глюкозы и липидов в печени грызунов [6].
ного антитела против Ki67 («Abcam», Велико-
Однако глобальные эффекты воздействия
британия). Парафиновые срезы толщиной
TCPOBOP на профиль низкомолекулярных ме-
3-4 мкм подвергали депарафинизации и ре-
таболитов в печени охарактеризованы не были.
гидратации по стандартной методике [9]. Сре-
Определение путей метаболической адап-
зы прогревали на водяной бане в 10 мМ цит-
тации, которые сопряжены с пролиферацией
ратном буфере (pH 6,0) в течение 20 мин для
гепатоцитов после воздействия агониста CAR,
демаскировки антигенов. Затем, после одно-
является основной целью настоящего иссле-
кратной промывки в дистиллированной воде
дования. Технология высокоэффективной жид-
и 10 мМ фосфатно-солевом буфере (pH 7,4),
костной хроматографии (ВЭЖХ) и масс-спек-
блокировали эндогенную пероксидазу в тече-
трометрии (МС) использовалась для опреде-
ние 5 мин в 3%-ной охлажденной перекиси
ления профиля малых биомолекул (сахаров,
водорода. Время экспозиции с первичными
аминокислот и нуклеотидов) для выявления
антителами против Ki67 составляло 60 мин
ключевых метаболических изменений в пе-
при комнатной температуре. Затем срезы ин-
чени мышей в ответ на введение химического
кубировали со стрептавидин-пероксидазным
митогена TCPOBOP.
комплексом, в качестве субстрата применяли
диаминобензидин. Дополнительно ядра до-
крашивали гематоксилином.
МАТЕРИАЛЫ И МЕТОДЫ
Выделение РНК, синтез кДНК и ПЦР в ре-
жиме реального времени. Выделение суммарной
Животные, использованные в исследовании.
РНК, обратную транскрипцию РНК и ПЦР
Самцы мышей (C57BL, масса 25-30 г) были
в режиме реального времени для определе-
предоставлены Научно-исследовательским ин-
ния уровней мРНК проводили согласно ранее
ститутом нейронаук и медицины (Новоси-
опубликованному протоколу [10]. Выделение
БИОХИМИЯ том 88 вып. 8 2023
1
304
МАЗИН и др.
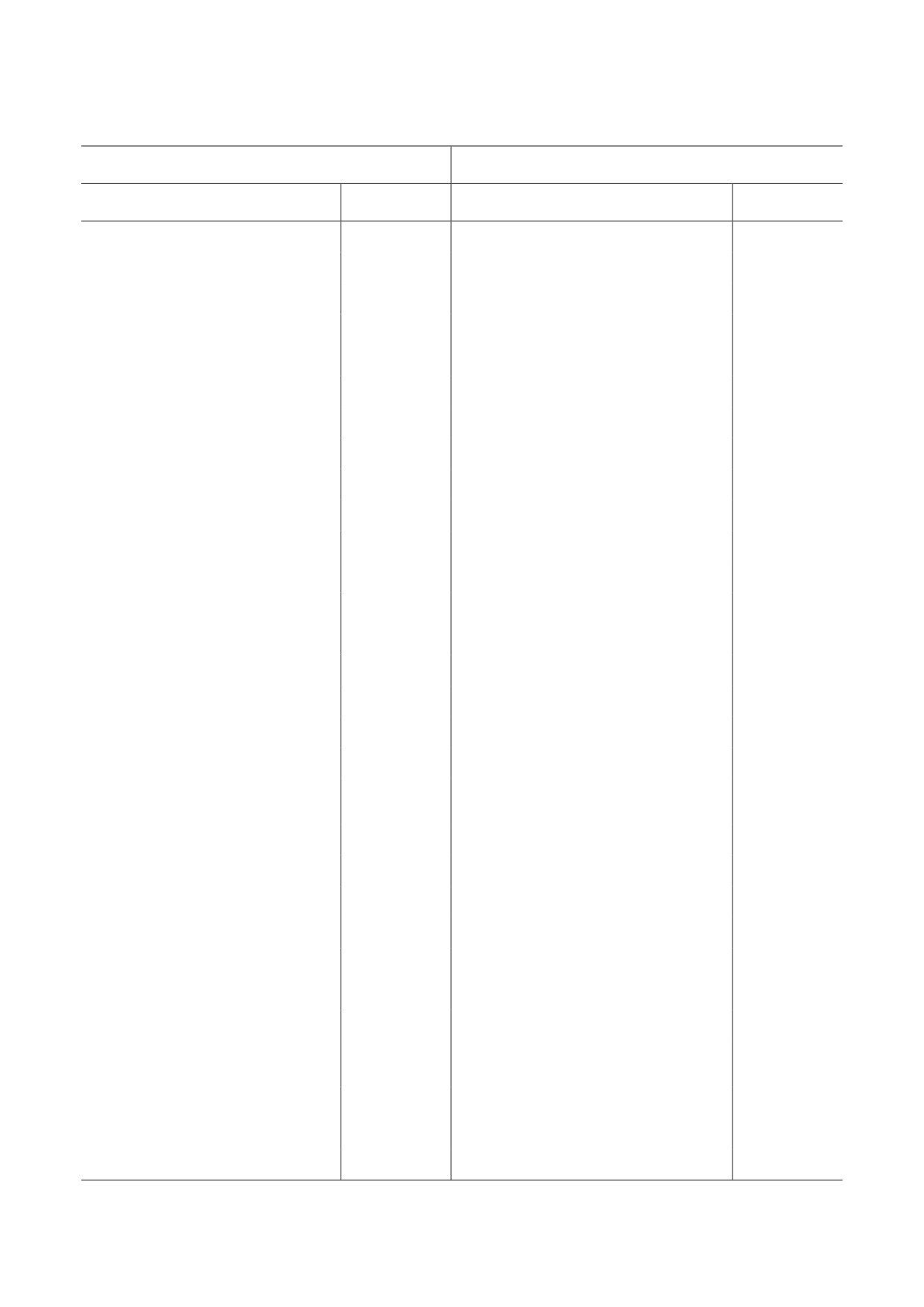
Таблица 1. Последовательности олигонуклеотидных праймеров, использующихся при ПЦР в режиме реального
времени для исследования относительного уровня мРНК
Ген
Прямой праймер (5′→3′)
Обратный праймер (5′→3′)
Cyp2b10
CCCAGTGTTCCACGAGACTT
GGTGCCGACAAAGAAGAGAG
Cyclind1
TGAAGGAGACCATTCCCTTG
CCACTTGAGCTTGTTCACCA
cMyc
ACGAGCACAAGCTCACCTCT
TCCAGCTCCTCCTCGAGTTA
E2f1
TCTGTACCACACAGCTGCAA
GCACAGGAAAACATCAATGG
Foxm1
TCCAAGGCAAAGACAGGAGA
GCTCCTCAACCTTAACCCGA
Mrpl46
GGGAGCAGGCATTCCTACAG
GGTCCGGTCATTTTTTTTGTCA
суммарной РНК из образцов печени мышей
10 мин. Супернатант переносили в новую поли-
проводили с использованием реагента TRIzol
пропиленовую пробирку и сушили в центри-
(«Invitrogen», США), согласно рекомендаци-
фуге-концентраторе SpeedVac («Thermo Fisher
ям производителя. Обратную транскрипцию
Scientific», США). Полученный остаток разво-
осуществляли с помощью набора RT-M-MuLV-
дили в 100 мкл реконституционного раствора
RH kit («Биолабмикс», Россия) в соответствии
вода/метанол (80/20) и подвергали метаболом-
с протоколом производителя. Для определе-
ному анализу с использованием ВЭЖХ-МС/МС.
ния уровня экспрессии исследуемых генов
Анализ ВЭЖХ-МС/МС. Хроматографичес-
проводили ПЦР в режиме реального вре-
кое разделение и анализ метаболитов проводи-
мени с использованием BioMaster HS-qPCR
ли по ранее опубликованному протоколу [12].
SYBR Blue (2×) («Биолабмикс») на амплифи-
Пробы анализировали на хроматографе
каторе CFX96™ («Bio-Rad Laboratories», США).
Shimadzu LC-20AD Prominence
(«Shimadzu
Для ПЦР использовали специфические оли-
Corporation», Япония), оснащенном автодо-
гонуклеотиды (табл.
1). Подбор специфич-
затором SIL-20AC («Shimadzu Corporation»).
ных праймеров осуществляли с использова-
Хроматографическое разделение проводили
нием онлайн-сервиса Primer-BLAST (https://
на колонке Prontosil 120-5-Amino (2,1 × 75 мм)
(«Эконова», Россия). Сбор данных проводи-
cgi?GROUP_TARGET=on). Оптимальная кон-
ли на масс-спектрометре API 6500 QTRAP
центрация всех пар праймеров в реакционной
(«AB SCIEX», США) c использованием про-
смеси составила 300 нМ. ПЦР проводили в
граммного обеспечения Analyst
1.6.2
(«AB
следующих условиях: предварительный про-
SCIEX»). Масс-спектрометрическую детекцию
грев при 95 °С - 3 мин; после этого следовали
проводили в режиме MRM в положительной и
40 основных циклов: денатурация при 95 °С -
отрицательной ионизации. Полученные хро-
15 с, отжиг при 59 °С - 20 с, элонгация при
матограммы обрабатывались в программе
72 °С - 20 с, сбор данных по флюоресценции
MultiQuantTM 2.1 Software («AB SCIEX») с
при 80 °С - 10 с. Для контроля специфичности
экспортированием данных в таблицы Microsoft
ПЦР использовали кривые плавления. Уровни
Excel. Последующий мультивариантный ана-
экспрессии мРНК рассчитывали на основе эф-
лиз и анализ биохимических путей проводи-
фективности ПЦР и Ct. Нормирование про-
лись с использованием онлайн-сервиса Metabo-
водили по гену Mrpl46. Уровень экспрессии
гена в печени одного из контрольных живот-
биохимических путей был проведен на основе
ных принимался за 1.
данных о метаболитах, нанесенных на карту в
Подготовка проб для метаболомного иссле-
соответствии с идентификаторами Киотской
дования. Все образцы печени обрабатывали
энциклопедии генов и геномов (KEGG).
одновременно в соответствии с протоколом,
Статистическая обработка результатов. Ста-
описанным ранее Yuan et al. [11]. Образцы пе-
тистический анализ проводился с использо-
чени (20-30 мг) гомогенизировали в 500 мкл
ванием t-критерия Стьюдента. Расчеты про-
80%-ного (v/v) охлажденного метанола для
изводились с использование программного
ВЭЖХ и инкубировали в течение ночи при
обеспечения GraphPad Prism 5.0 («GraphPad
-80 °C для осаждения белка. Затем образцы
Software Inc.», США). Значение р < 0,05 счита-
центрифугировали при 4 °С и 17 000 g в течение
лось статистически значимым.
БИОХИМИЯ том 88 вып. 8 2023
АГОНИСТ CAR ИЗМЕНЯЕТ МЕТАБОЛОМ В ПЕЧЕНИ
1305
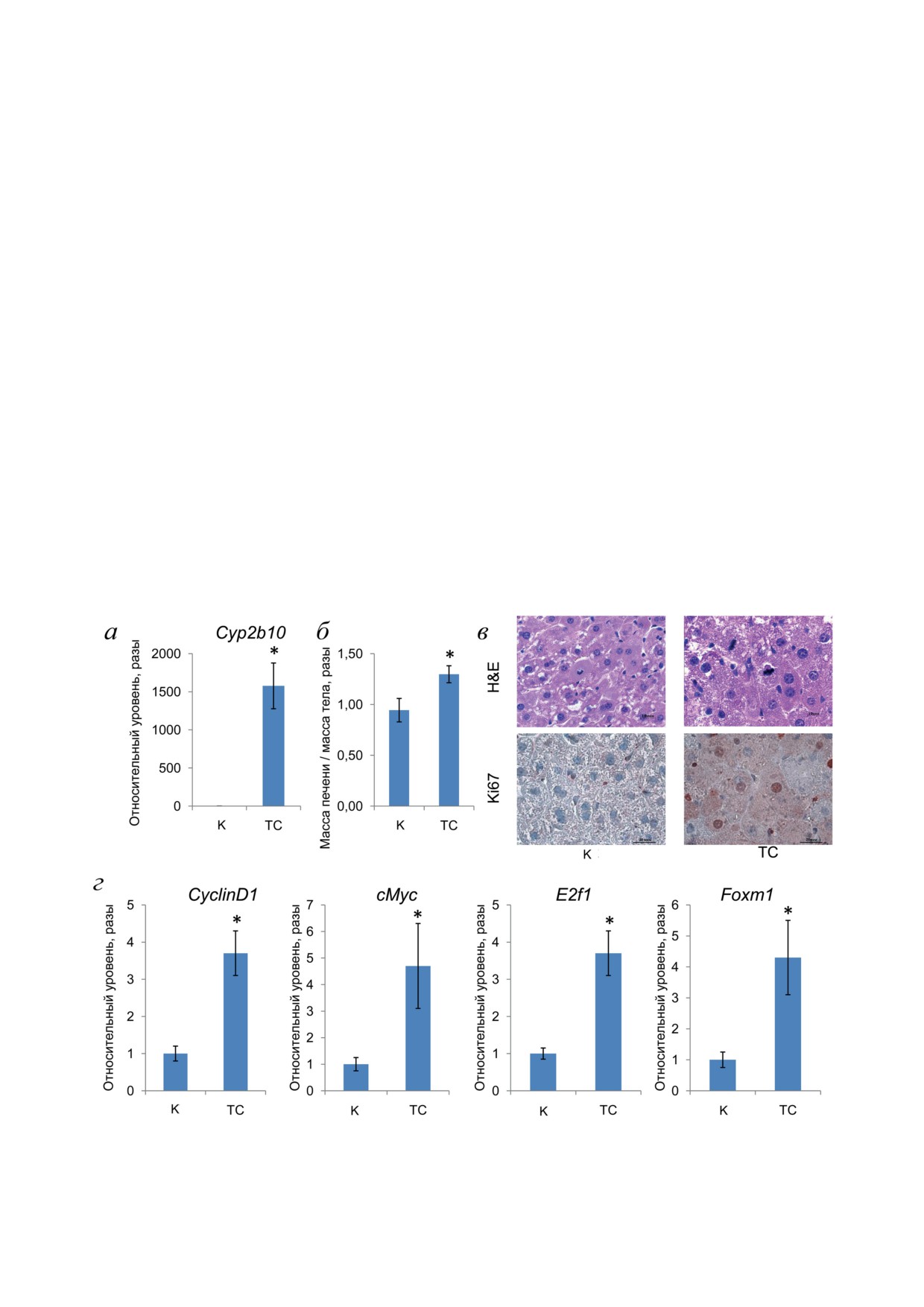
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
ночной дольки вокруг собирательных вен. Об-
разцы из контрольной группы животных были
В первую очередь была подтверждена
без явных Ki67-положительных ядер, тогда как
TCPOBOP-опосредованная активация CAR
в образцах, полученных от ТСРОВОР-индуци-
в печени мышей путем изучения экспрессии
рованных мышей, выявлялось до 40% Ki67-по-
гена Cyp2b10, ключевого биомаркера для изме-
ложительных ядер гепатоцитов от общего чис-
рения активации CAR. Как и ожидалось, экс-
ла гепатоцитов в 5 случайно выбранных полях.
прессия гена Cyp2b10 в печени мышей увеличи-
Кроме того, воздействие TCPOBOP достовер-
лась после воздействия TCPOBOP (рис. 1, а).
но увеличивало экспрессию генов, кодирую-
Далее были проведены эксперименты по
щих регуляторы клеточного цикла, Cyclind1,
оценке гиперплазии печени при воздействии хи-
cMyc, E2f1 и Foxm1, в печени мышей по срав-
мического митогена. Увеличение экспрессии
нению с контрольными животными (рис. 1, г).
гена Cyp2b10 после воздействия ТСРОВОР со-
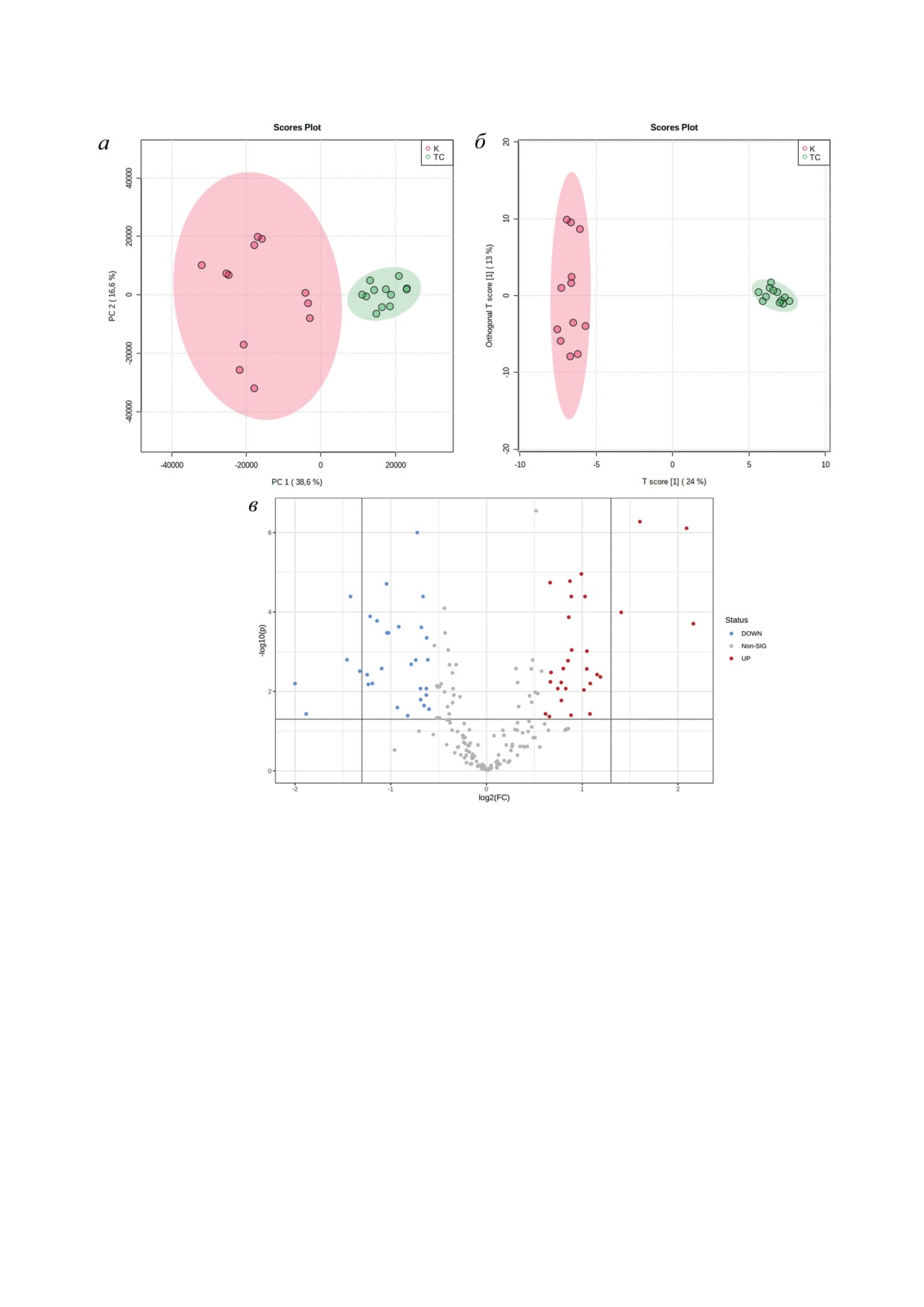
С помощью ВЭЖХ-МС/МС анализа об-
провождалось достоверным увеличением ин-
разцов печени были изучены различия в мета-
декса относительной массы печени, который
боломном профиле контрольных и ТСРОВОР-
определяли как масса печени по отношению
индуцированных мышей. Мультивариантный
к массе тела животного (рис. 1, б). Гистологи-
анализ полученных результатов методом анали-
ческий анализ печени мышей и иммуноокра-
за главных компонент (PCA) продемонстри-
шивание срезов печени с использованием ан-
ровал наличие различий в метаболомном
тител против Ki67 показали, что воздействие
профиле печени, вызванных воздействием
TCPOBOP усиливало митотические процессы
TCPOBOP. Проведенный анализ не показал
в гепатоцитах (рис. 1, в). Так, на срезах, полу-
перекрытия контрольной группы и группы,
ченных от ТСРОВОР-индуцированных мышей,
получавшей TCPOBOP, что указывает на чет-
наблюдались фигуры митоза, особенно среди
кую кластеризацию двух групп образцов пече-
гепатоцитов, расположенных в центре пече-
ни (рис. 2, а).
Рис. 1. Влияние TCPOBOP на мишень CAR - ген Cyp2b10 (а), размер печени (б), гистопатологию и иммуноокраши-
вание Ki67 (в) и гены, кодирующие регуляторы клеточного цикла (г). * Достоверное отличие от контрольных живот-
ных (р < 0,05). Мышей обрабатывали 1,4-бис [2-(3,5-дихлорпиридилокси)] бензолом (TC) или только подсолнечным
маслом (К)
БИОХИМИЯ том 88 вып. 8 2023
1306
МАЗИН и др.
Рис. 2. Изменение метаболомного профиля в печени мышей после воздействия TCPOBOP (TC) по сравнению с кон-
трольными животными (К). a - Мультивариантный анализ методом анализа главных компонент; б - ортогональный
дискриминантный анализ методом наименьших квадратов образцов печени на основе всех метаболитов. Каждый
символ представляет один образец одной мыши. Образцы одной группы показаны одним цветом. Цветное поле пред-
ставляет 95%-ный доверительный интервал группы с кружками одного цвета. в - Вулканическая диаграмма показы-
вает метаболиты, уровень которых достоверно изменяется при воздействии TCPOBOP. Метаболиты, уровень которых
достоверно снижается, обозначены синим. Метаболиты, уровень которых достоверно повышен, обозначены красным
Дополнительно был проведен ортогональ-
30 метаболитов уровень снижался, а у 28 - по-
ный дискриминантный анализ методом наи-
вышался (рис. 2, в; табл. 2).
меньших квадратов (OPLS-DA). Данный ана-
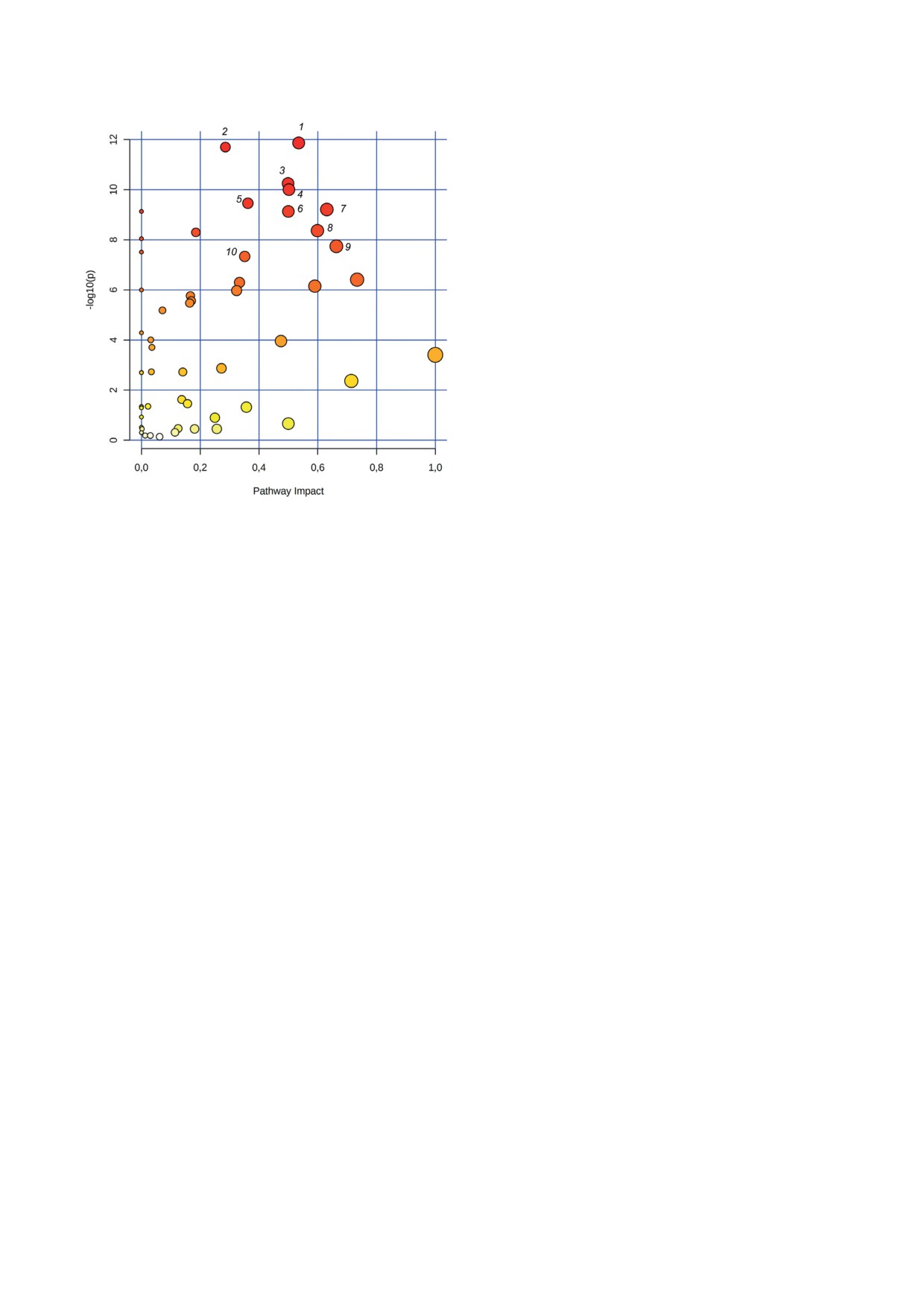
В заключении был проведен анализ биохи-
лиз позволил лучше выделить метаболические
мических путей, на которые повлиял агонист
различия между двумя группами образцов
мышиного CAR. Результаты анализа проде-
печени мышей (рис. 2, б). Данные результаты
монстрировали перепрограммирование мета-
свидетельствуют о том, что контрольные мыши
болической сети после воздействия TCPOBOP.
и мыши, получавшие TCPOBOP, имеют раз-
Как показано на рис. 3, TCPOBOP-индуциро-
ные метаболомные профили печени. Сравне-
ванная пролиферация гепатоцитов сопрово-
ние уровней метаболитов между группами жи-
ждается изменениями в метаболизме нуклео-
вотных показало, что воздействие TCPOBOP
тидов (как пиримидиновых, так и пуриновых),
приводит к достоверному изменению уровней
метаболизме аминокислот (тирозина, глици-
58 метаболитов в печени мышей, из которых у
на, серина и треонина, глутамина и глутамата,
БИОХИМИЯ том 88 вып. 8 2023
АГОНИСТ CAR ИЗМЕНЯЕТ МЕТАБОЛОМ В ПЕЧЕНИ
1307
Таблица 2. Метаболиты, уровень которых достоверно изменяется в печени мышей после воздействия TCPOBOP
TCPOBOP-опосредованное снижение уровня
TCPOBOP-опосредованное увеличение уровня
метаболит
количество раз
метаболит
количество раз
ксантиловая кислота
0,25000
2-аминооктановая кислота
1,5766
гиппуровая кислота
0,27108
никотинамид мононуклеотид
1,5854
глицерофосфохолин
0,36442
1-метилгистидин
1,5886
акадезин
0,37370
N-ацетилорнитин
1,5961
бетаин
0,39950
гуанозинмонофосфат
1,6758
лактоза
0,42146
пантотеновая кислота
1,7183
аргининосукцинат
0,42503
изобутирилглицин
1,7203
цитруллин
0,43072
дезоксиинозин
1,7443
3-метилгистидин
0,43746
уридин-5′-дифосфат
1,7756
аскорбиновая кислота
0,45287
3-фосфоглицериновая кислота
1,8045
ксантозин
0,46816
пировиноградная кислота
1,8142
гипоксантин
0,48508
2-изопропилмалат
1,8303
инозин
0,48643
уреидосукцинат
1,8442
1-метилникотинамид
0,49199
цитидин
1,8486
оротидиловая кислота
0,52455
триптофанол
1,8527
аденозин
0,52988
глутамат
1,9878
3-гидроксиантраниловая кислота
0,56541
D-глицеральдегид-3-фосфат
2,0244
ксантин
0,57976
фосфоенолпируват
2,0415
5-аминоимидазол-4-карбоксамид
гипотаурин
0,59937
2,0675
рибонуклеотид
L-серин
0,60573
дезоксиуридинмонофосфат (dUMP)
2,0697
N-ацетил-L-аланин
0,62012
спермидин
2,1151
дофамин
0,62094
L-кинуренин
2,1205
метанефрин
0,62424
уреидопропионат
2,2277
имидазолуксусная кислота
0,63156
аллантоиновая кислота
2,2815
5′-метилтиоаденозин
0,63666
глутатиондисульфид
2,6520
фосфорилхолин
0,64681
глюконовая кислота
3,0366
дезокситимидинмонофосфат
2-арахидоноилглицерин
0,64719
4,2578
(dTMP)
гуанин
0,64855
D-седогептулозо-7-фосфат
4,4718
O-ацетилсерин
0,65384
цистин
0,65976
БИОХИМИЯ том 88 вып. 8 2023
1308
МАЗИН и др.
точного цикла (Cyclind1, cMyc, E2f1 и Foxm1),
что коррелировало с увеличением массы пече-
ни животных. Известно, что некоторые регу-
ляторы клеточного цикла, такие как Cyclin D1
и E2F1, принимают участие в регуляции экс-
прессии генов, кодирующих метаболические
ферменты, и тем самым контролируют метабо-
лизм [14]. Эти ферменты участвуют в разнооб-
разных биохимических путях синтеза метабо-
лических предшественников для роста клеток.
Пролиферация гепатоцитов требует уве-
личения внутриклеточной биомассы [7]. Соот-
ветственно, производство белков, липидов и
нуклеиновых кислот необходимо для успешно-
го деления гепатоцитов. Более того, производ-
ство биомассы гепатоцитов должно быть ско-
ординировано с событиями клеточного цикла,
чтобы получить две здоровые дочерние клет-
ки. Таким образом, характеристика принци-
пов, лежащих в основе этих путей, может
дать важную информацию для понимания
Рис. 3. Метаболические пути, активность которых изме-
няется после воздействия TCPOBOP в печени мышей.
роста и пролиферации гепатоцитов. В настоя-
Размер (диаметр круга) и цвет (оттенок) каждого круга
щем исследовании был проведен комплекс-
основан на значимости воздействия ТСРОВОР на мета-
ный метаболомный анализ с использованием
болический путь. Анализ путей проводился с использо-
ВЭЖХ-МС/МС, который позволил выявить
ванием онлайн-сервиса MetaboAnalyst. 1 - Пиримиди-
новый обмен; 2 - метаболизм тирозина; 3 - пуриновый
58 метаболитов, уровень которых изменяется
обмен; 4 - метаболизм глицина, серина и треонина;
при TCPOBOP-индуцированной гиперплазии
5 - метаболизм пирувата; 6 - метаболизм D-глютами-
печени у мышей. Анализ путей этих дифферен-
на и D-глутамата; 7 - метаболизм аргинина и пролина;
циально затронутых метаболитов в печени мы-
8 - биосинтез аргинина; 9 - метаболизм цистеина и ме-
тионина; 10 - пентозофосфатный путь
шей продемонстрировал значительное, опо-
средованное TCPOBOP, обогащение ряда био-
химических процессов. Так, в работе было
аргинина и пролина, цистеина и метионина) и
продемонстрировано, что большинство изме-
биосинтезе аргинина. Кроме того, показано,
ненных под действием химического митогена
что пути метаболизма энергетических субстра-
метаболитов вовлечены в пути биосинтеза
тов (метаболизм пентозофосфата) участвуют в
и метаболизма нуклеотидов и аминокислот.
метаболической адаптации печени мышей при
В совокупности эти данные указывают на ме-
воздействии TCPOBOP.
таболические предпочтения, лежащие в основе
высокой способности к пролиферации гепато-
цитов после введения TCPOBOP.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Глюкоза является основным субстратом
для пролиферирующих клеток, обеспечивая
Хорошо известно, что TCPOBOP-опосре-
клетки как ATP в качестве источника энергии,
дованная активация CAR вызывает гиперпла-
так и мономерными блоками для макромоле-
зию печени мышей [3]. CAR-опосредованная
кулярного синтеза [15]. В связи с этим гли-
гиперплазия печени связана с изменением экс-
колиз считают биохимической платформой,
прессии большого набора генов, кодирующих
поддерживающей пролиферацию клеток [16].
регуляторы пролиферации гепатоцитов [13].
В настоящем исследовании обнаружено, что
В настоящем исследовании доза и схема вве-
TCPOBOP-индуцированная пролиферация ге-
дения ТСРОВОР полностью соответствовали
патоцитов сопровождается увеличением со-
ранее опубликованным данным, в которых
держания метаболитов, которые образуются
впервые было показано, что ТСРОВОР-опо-
за счет гликолитического разрушения глюко-
средованная активация CAR приводит к ги-
зы, таких как глицеральдегид-3-фосфат, фос-
перплазии печени мышей [3]. Наши резуль-
фоенолпируват и пировиноградная кислота.
таты подтвердили, что активация CAR при
Это может свидетельствовать об активации
воздействии ТСРОВОР сопровождается уве-
гликолиза при TCPOBOP-индуцированной про-
личением экспрессии генов-регуляторов кле-
лиферации гепатоцитов мышей.
БИОХИМИЯ том 88 вып. 8 2023
АГОНИСТ CAR ИЗМЕНЯЕТ МЕТАБОЛОМ В ПЕЧЕНИ
1309
Накопление гликолитических промежу-
нокислоты служат субстратами для энергети-
точных соединений может приводить к их
ческого метаболизма. По-видимому, наблю-
перераспределению в другие биохимические
даемое снижение уровня серина, аланина,
пути, ответвляющиеся от гликолиза. Одним
метионина и тирозина связано с тем, что гли-
из таких путей является пентозофосфатный
когенные аминокислоты выступают субстра-
путь (PPP), который играет важную роль в
тами для биосинтеза углеводов и нуклеотидов.
биосинтезе нуклеотидов. РРР использует про-
Метионин также является основным ис-
межуточные продукты гликолиза в реакциях,
точником метильных групп, превращаясь в
производящих рибозо-5-фосфат для синтеза
универсальный донор метильных групп S-аде-
нуклеиновых кислот и NADPH для анаболиз-
нозилметионин (SAM). SAM обеспечивает
ма [17]. Наши данные показали, что воздей-
метильную группу для метилирования ДНК и
ствие TCPOBOP на печень мышей значительно
гистонов. После переноса метильной груп-
увеличивало уровень седогептулозо-7-фосфа-
пы SAM превращается в S-аденозилгомоци-
та, который связывает гликолиз, пентозофос-
стеин (SAH). Наши результаты показали опо-
фатный путь и биосинтез нуклеотидов [18].
средованное TCPOBOP снижение в печени
Более того, наши результаты показали, что при
мышей уровня метионина, а также увеличение
воздействии ТСРОВОР происходит измене-
уровня SAH. Этот факт может указывать на то,
ние уровней метаболитов, связанных с мета-
что TCPOBOP может вызывать метилирование
болизмом пиримидинов (UDP, dUMP, dTMP
ДНК и гистонов, что может способствовать
и цитидин) и пуринов (ксантозин, ксантин,
регуляции транскрипции. Предыдущие иссле-
гипоксантин, инозин, GMP и IMP). Эти мета-
дования показали, что активация CAR связана
болиты связаны с биосинтезом нуклеотидов.
с эпигенетическими изменениями, такими как
По-видимому, TCPOBOP-индуцированные пути
метилирование ДНК и гистонов, что нарушает
метаболизма, связанные с синтезом пирими-
регуляцию экспрессии генов в печени [24, 25].
дина и пурина, поддерживают пролиферацию
гепатоцитов за счет предоставления нуклеоти-
дов, которые являются необходимыми пред-
ЗАКЛЮЧЕНИЕ
шественниками для биосинтеза РНК и ДНК.
Ранее было показано, что высокая регене-
Ранее с использованием технологии
ративная способность органов сочетается с
RNA-seq установлено, что активация CAR
высоким уровнем метаболизма пиримиди-
под действием TCPOBOP вызывает измене-
нов, а пиримидиновые нуклеозиды могут спо-
ние экспрессии более чем 2000 генов в печени
собствовать регенерации различных тканей
мышей [26]. В настоящей работе показано,
in vivo [19].
что при воздействии TCPOBOP наблюдается
Наше исследование показало, что после
скоординированный контроль метаболизма и
воздействия TCPOBOP также наблюдаются
пролиферации гепатоцитов. По всей видимо-
изменения в биосинтезе и метаболизме ряда
сти, активация CAR инициирует внутрикле-
аминокислот в печени мышей. Так, в печени
точную программу, которая способствует гло-
мышей, получавших TCPOBOP, наблюдалось
бальной скоординированной метаболической
увеличение метаболитов, участвующих в био-
активности, необходимой для пролиферации
синтезе аргинина. Аргинин является предше-
гепатоцитов. Эти данные могут свидетельство-
ственником не только для синтеза белков, но
вать о том, что CAR-опосредованные сигналь-
и полиаминов. В этом исследовании воздей-
ные каскады образуют сеть сигнальных цепей,
ствие TCPOBOP повышало уровень двух поли-
ведущих к сложной регуляции пролифера-
аминов, путресцина и спермидина, в печени
ции гепатоцитов не только на транскриптом-
мышей. И путресцин, и спермидин являются
ном, но и на метаболомном уровне. Требуются
хорошо известными прорегенеративными ме-
дальнейшие углубленные исследования для
таболитами [20], способными усиливать реге-
выявления более нисходящих селективных
нерацию печени после частичной гепатэкто-
метаболических путей, которые участвуют в
мии [21-23].
метаболическом гомеостазе во время проли-
Аминокислоты также могут выступать в
ферации гепатоцитов после TCPOBOP-опо-
качестве субстратов для разнообразных био-
средованной активации CAR. Тем не менее
химических путей. Результаты, полученные в
наши метаболомные данные могут дать но-
этом исследовании, показали снижение глико-
вое представление о биологических механиз-
генных аминокислот, таких как серин, аланин,
мах, которые происходят во время TCPOBOP-
метионин и тирозин, при воздействии химиче-
индуцированной пролиферации гепатоцитов
ского митогена TCPOBOP. Гликогенные ами-
у мышей.
БИОХИМИЯ том 88 вып. 8 2023
1310
МАЗИН и др.
Вклад авторов. В.О. Пустыльняк, Л.Ф. Гу-
Минобрнауки России (соглашение № 075-15-
ляева - концепция и руководство работой;
2021-691).
М.Э. Мазин, А.М. Перевалова, А.А. Ярушкин,
Конфликт интересов. Авторы заявляют об
Ю.А. Пустыльняк, А.Д. Рогачев, Е.А. Прокопь-
отсутствии конфликта интересов.
ева - проведение экспериментов и статисти-
Соблюдение этических норм. Содержание
ческая обработка результатов; М.Э. Мазин,
животных и все эксперименты с животными
А.А. Ярушкин, В.О. Пустыльняк, Л.Ф. Гуляе-
проводились в соответствии с позицией по
ва - обсуждение результатов исследования;
этике использования животных в исследова-
М.Э. Мазин
- написание текста; В.О. Пу-
ниях, выполняемых при поддержке Россий-
стыльняк -редактирование текста статьи.
ского научного фонда, а также в соответствии
Финансирование. Работа выполнена при
с Директивой 2010/63/ЕС Европейского пар-
поддержке Российского научного фонда
ламента и Совета Европейского Союза от
(грант № 18-15-00021).
22 сентября 2010 г. и одобрены Комиссией по
Благодарности. Работа выполнена с исполь-
биоэтике Федерального исследовательского
зованием оборудования ЦКП «Протеомный
центра фундаментальной и трансляционной
анализ», поддержанного финансированием
медицины (протокол № 23-17).
СПИСОК ЛИТЕРАТУРЫ
1.
Yan, J., and Xie, W. (2016) A brief history of the
7.
Solhi, R., Lotfinia, M., Gramignoli, R., Najimi, M.,
discovery of PXR and CAR as xenobiotic receptors,
and Vosough, M. (2021) Metabolic hallmarks of liver
Acta Pharm. Sin. B, 6, 450-452, doi: 10.1016/j.apsb.
regeneration, Trends Endocrinol. Metab., 32, 731-745,
2016.06.011.
doi: 10.1016/j.tem.2021.06.002.
2.
Cai, X., Young, G. M., and Xie, W. (2021) The xe-
8.
Cardiff, R. D., Miller, C. H., and Munn, R. J. (2014)
nobiotic receptors PXR and CAR in liver phys-
Manual hematoxylin and eosin staining of mouse
iology, an update, Biochim. Biophys. Acta Mol.
tissue sections, Cold Spring Harb. Protoc.,
2014,
Basis Dis.,
1867,
166101, doi:
10.1016/j.bbadis.
655-658, doi: 10.1101/pdb.prot073411.
2021.166101.
9.
Graefe, C., Eichhorn, L., Wurst, P., Kleiner, J.,
3.
Blanco-Bose, W. E., Murphy, M. J., Ehninger, A.,
Heine, A., Panetas, I., Abdulla, Z., Hoeft, A., Frede, S.,
Offner, S., Dubey, C., Huang, W., Moore, D. D.,
Kurts, C., Endl, E., and Weisheit, C. K. (2019) Opti-
and Trumpp, A. (2008) C-Myc and its target FoxM1
mized Ki-67 staining in murine cells: a tool to deter-
are critical downstream effectors of constitutive
mine cell proliferation, Mol. Biol. Rep., 46, 4631-4643,
androstane receptor (CAR) mediated direct liver
doi: 10.1007/s11033-019-04851-2.
hyperplasia, Hepatology, 48, 1302-1311, doi: 10.1002/
10.
Mazin, M. E., Yarushkin, A. A., Pustylnyak, Y. A.,
hep.22475.
Prokopyeva, E. A., and Pustylnyak, V. O. (2022) Pro-
4.
Tschuor, C., Kachaylo, E., Limani, P., Raptis, D. A.,
motion of NR1I3-mediated liver growth is accompa-
Linecker, M., Tian, Y., Herrmann, U., Grabliaus-
nied by STAT3 activation, Mol. Biol. Rep., 49, 4089-
kaite, K., Weber, A., Columbano, A., Graf, R.,
4093, doi: 10.1007/s11033-022-07340-1.
Humar, B., and Clavien, P. A. (2016) Constitutive
11.
Yuan, M., Breitkopf, S. B., Yang, X., and Asara, J. M.
androstane receptor (Car)-driven regeneration protects
(2012) A positive/negative ion-switching, targeted
liver from failure following tissue loss, J. Hepatol., 65,
mass spectrometry-based metabolomics platform for
66-74, doi: 10.1016/j.jhep.2016.02.040.
bodily fluids, cells, and fresh and fixed tissue, Nat.
5.
Lodato, N. J., Melia, T., Rampersaud, A., and
Protoc., 7, 872-881, doi: 10.1038/nprot.2012.024.
Waxman, D. J. (2017) Sex-differential responses of
12.
Rogachev, A. D., Alemasov, N. A., Ivanisenko, V. A.,
tumor promotion-associated genes and dysregulation
Ivanisenko, N. V., Gaisler, E. V., Oleshko, O. S.,
of novel long noncoding RNAs in constitutive
Cheresiz, S. V., Mishinov, S. V., Stupak, V. V., and
androstane receptor-activated mouse liver, Toxicol.
Pokrovsky, A. G. (2021) Correlation of metabolic pro-
Sci., 159, 25-41, doi: 10.1093/toxsci/kfx114.
files of plasma and cerebrospinal fluid of high-grade
6.
Skoda, J., Dohnalova, K., Chalupsky, K., Stahl, A.,
glioma patients, Metabolites, 11, 133, doi: 10.3390/
Templin, M., Maixnerova, J., Micuda, S., Grøntved, L.,
metabo11030133.
Braeuning, A., and Pavek, P. (2022) Off-target lip-
13.
Kazantseva, Y. A., Pustylnyak, Y. A., and Pustylnyak,
id metabolism disruption by the mouse constitu-
V. O. (2016) Role of nuclear constitutive androstane
tive androstane receptor ligand TCPOBOP in hu-
receptor in regulation of hepatocyte proliferation and
manized mice, Biochem. Pharmacol., 197, 114905,
hepatocarcinogenesis, Biochemistry (Moscow),
81,
doi: 10.1016/j.bcp.2021.114905.
338-347, doi: 10.1134/S0006297916040040.
БИОХИМИЯ том 88 вып. 8 2023
АГОНИСТ CAR ИЗМЕНЯЕТ МЕТАБОЛОМ В ПЕЧЕНИ
1311
14.
Huber, K., Mestres-Arenas, A., Fajas, L., and Leal-
polyamines, spermidine and spermine, causes a
Esteban, L. C. (2021) The multifaceted role of cell cycle
total arrest in translation and growth in mammalian
regulators in the coordination of growth and metabo-
cells, Proc. Natl. Acad. Sci. USA, 110, 2169-2174,
lism, FEBS J., 288, 3813-3833, doi: 10.1111/febs.15586.
doi: 10.1073/pnas.1219002110.
15.
Locasale, J. W., and Cantley, L. C. (2011) Metabolic
22.
Alhonen, L., Räsänen, T.L., Sinervirta, R., Parkkinen,
flux and the regulation of mammalian cell growth, Cell
J. J., Korhonen, V. P., Pietilä, M., and Jänne, J.
Metab., 14, 443-451, doi: 10.1016/j.cmet.2011.07.014.
(2002) Polyamines are required for the initiation of
16.
Lunt, S. Y., and Vander Heiden, M. G. (2011) Aerobic
rat liver regeneration, Biochem. J., 362, 149-153,
glycolysis: meeting the metabolic requirements of cell
doi: 10.1042/0264-6021:3620149.
proliferation, Annu. Rev. Cell Dev. Biol., 27, 441-464,
23.
Chattopadhyay, M. K., Park, M. H., and Tabor, H.
doi: 10.1146/annurev-cellbio-092910-154237.
(2008) Hypusine modification for growth is the major
17.
Ge, T., Yang, J., Zhou, S., Wang, Y., Li, Y., and
function of spermidine in Saccharomyces cerevisiae
Tong, X. (2020) The role of the pentose phosphate
polyamine auxotrophs grown in limiting spermidine,
pathway in diabetes and cancer, Front Endocrinol.
Proc. Natl. Acad. Sci. USA,
105,
6554-6559,
(Lausanne), 11, 365, doi: 10.3389/fendo.2020.00365.
doi: 10.1073/pnas.0710970105.
18.
Jin, L., and Zhou, Y. (2019) Crucial role of the pentose
24.
Lempiäinen, H., Müller, A., Brasa, S., Teo, S. S.,
phosphate pathway in malignant tumors, Oncol. Lett.,
Roloff, T. C., Morawiec, L., Zamurovic, N., Vicart, A.,
17, 4213-4221, doi: 10.3892/ol.2019.10112.
Funhoff, E., Couttet, P., Schübeler, D., Grenet, O.,
19.
Liu, Z., Li, W., Geng, L., Sun, L., Wang, Q., Yu, Y.,
Marlowe, J., Moggs, J., and Terranova, R. (2011)
Yan, P., Liang, C., Ren, J., Song, M., Zhao, Q., Lei, J.,
Phenobarbital mediates an epigenetic switch at
Cai, Y., Li, J., Yan, K., Wu, Z., Chu, Q., Li, J.,
the constitutive androstane receptor (CAR) tar-
Wang, S., Li, C., Han, J. J., Hernandez-Benitez, R.,
get gene Cyp2b10 in the liver of B6C3F1 mice,
Shyh-Chang, N., Belmonte, J. C. I., Zhang, W., Qu, J.,
PLoS One,
6, e18216, doi:
10.1371/journal.pone.
and Liu, G. H. (2022) Cross-species metabolom-
0018216.
ic analysis identifies uridine as a potent regeneration
25.
Rampersaud, A., Lodato, N.J., Shin, A., and Wax-
promoting factor, Cell Discov., 8, 6, doi: 10.1038/
man, D. J. (2019) Widespread epigenetic changes to
s41421-021-00361-3.
the enhancer landscape of mouse liver induced by a
20.
Doi, J., Fujimoto, Y., Teratani, T., Kasahara, N.,
specific xenobiotic agonist ligand of the nuclear re-
Maeda, M., Tsuruyama, T., Iida, T., Yagi, S., and
ceptor CAR, Toxicol. Sci., 171, 315-338, doi: 10.1093/
Uemoto, S. (2019) Bolus administration of polyamines
toxsci/kfz148.
boosts effects on hepatic ischemia-reperfusion injury
26.
Cui, J. Y., and Klaassen, C. D. (2016) RNA-Seq
and regeneration in rats, Eur. Surg. Res., 60, 63-73,
reveals common and unique PXR- and CAR-target
doi: 10.1159/000497434.
gene signatures in the mouse liver transcriptome,
21.
Mandal, S., Mandal, A., Johansson, H. E., Orjalo,
Biochim. Biophys. Acta, 1859, 1198-1217, doi: 10.1016/
A. V., and Park, M. H. (2013) Depletion of cellular
j.bbagrm.2016.04.010.
CONSTITUTIVE ANDROSTANE RECEPTOR AGONIST
INITIATES METABOLIC ACTIVITY
REQUIRED FOR HEPATOCITE PROLIFERATION
M. E. Mazin1,2, A. M. Perevalova1, A. A. Yarushkin2, Y. A. Pustylnyak1,
A. D. Rogachev1, E. A. Prokopyeva1,2, L. F. Gulyaeva1,2, and V. O. Pustylnyak1,2*
1 Novosibirsk State University, 630090 Novosibirsk, Russia; e-mail: pustylnyak@post.nsu.ru
2 Federal Research Center of Fundamental and Translational Medicine,
630117 Novosibirsk, Russia
Constitutive androstane receptor (CAR, NR1I3) activation by chemical compounds evokes liver hyperplasia
in rodent. 1,4-Bis[2-(3,5-dichloropyridyloxy)] benzene (TCPOBOP), a mouse CAR agonist, is most com-
monly used to study chemically induced liver hyperplasia and hepatocyte proliferation in vivo. TCPOBOP
is potent murine liver chemical mitogen, which induces rapid direct liver hyperplasia independent of liver
injury. In recent years, a lot of data has been accumulated on the transcription program that characterizes
TCPOBOP-induced hepatocyte proliferation. However, there are scarce data about metabolic requirements
of hepatocytes dividing upon treatment with xenobiotics. In present study, we employed liquid chromatog-
raphy - mass spectrometry technology combined with statistical analysis to develop a metabolite profile of
БИОХИМИЯ том 88 вып. 8 2023
1312
МАЗИН и др.
small biomolecules, to identify key metabolic changes in male mouse liver tissue after TCPOBOP adminis-
tration. Analysis of biochemical pathways of the differentially affected metabolites in mouse livers demon-
strated significant TCPOBOP-mediated enrichment of several processes including those relevant to nucle-
otide metabolism, amino acid metabolism, and energy substrate metabolism. Our findings provide evidence
to support the conclusion that CAR agonist, TCPOBOP, initiates an intracellular program that promotes
the global coordinated metabolic activities required for hepatocyte proliferation. Our metabolic data may
provide novel insight into the biological mechanisms that occur during TCPOBOP-induced hepatocyte
proliferation in mice.
Keywords: constitutive androstane receptor, TCPOBOP, liver, hepatocyte, metabolomics
БИОХИМИЯ том 88 вып. 8 2023