ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 100, 3, 2021
УДК 595.728:591.185
ФУНКЦИИ ХОРДОТОНАЛЬНЫХ СЕНСИЛЛ КУЗНЕЧИКОВ
(ORTHOPTERA, TETTIGONIIDAE)
© 2021 г. Р. Д. Жантиев,* О. С. Корсуновская**
Кафедра энтомологии биологического факультета
Московского государственного университета им. М. В. Ломоносова
Ленинские горы, 1-12, Москва, 119234 Россия
*e-mail: zhantiev@mail.ru, **e-mail: korsuno@mail.ru
Поступила в редакцию 10.07.2021 г.
После доработки 28.08.2021 г.
Принята к публикации 28.08.2021 г.
С помощью электрофизиологических методов изучена слуховая и вибрационная чувстви-
тельность идентифицированных одиночных слуховых рецепторов кузнечиков. В промежуточ-
ном органе выявлены нейроны, реакция которых на виброускорение в значительном частотном
диапазоне не зависит от частоты стимула; наряду с ними имеются клетки, демонстрирующие
повышенную чувствительность к частотам 0.4-0.8 кГц (на смещение), и (или) 0.1-0.3, 1-1.2 кГц,
и 1.4-3 кГц при всех параметрах вибраций. Кроме того, у большинства изученных рецепторов
наблюдается зона повышенной чувствительности к высокочастотным вибрациям (1.5-2.5 кГц).
У сенсилл слухового гребня выявлена повышенная чувствительность в области 0.1-0.3, 0.4-0.8,
1.0-1.2 и 1.4-2.5 кГц. У одной и той же сенсиллы при восприятии разных параметров вибра-
ций оптимальные частоты могут находиться в разных частотных диапазонах. Такие различия
в чувствительности к виброускорению, виброскорости и смещению, а также разные оптималь-
ные частоты у рецепторов промежуточного органа и слухового гребня могут быть обусловлены
различиями в размерах, положении и деталях строения сенсилл, их собственными резонансами
и реакциями на резонансные колебания участка трахеи, на котором расположены виброрецепто-
ры. Таким образом, хордотональная сенсилла - это бифункциональный механорецептор, кото-
рый наряду со слуховой чувствительностью может сочетать функции как приемника смещения,
так и акселерометра за счет различий в механических свойствах клеток сенсиллы и окружающих
ее структур.
Ключевые слова: кузнечики, тимпанальный орган, бифункциональные слуховые рецепторы,
вибрационная чувствительность.
DOI: 10.31857/S0367144521030035
Восприятие акустических и вибрационных стимулов играет существенную роль
в жизни насекомых. Так, именно звуки и колебания субстрата сигнализируют о при-
ближении хищника. Кроме того, звуковые и вибрационные сигналы используются
многими насекомыми в брачном поведении, а хищниками - при поиске жертвы
(см., например, обзоры: Жантиев, 1981; Yack, 2016; Virant-Doberlet, 2019). Наиболее
развитой вибро-акустической коммуникацией обладают, наряду с цикадовыми (Hemi-
523
ptera, Auchenorrhyncha), прямокрылые насекомые (Orthoptera) и, в частности, кузнечи-
ковые (Tettigonioidea). Известно, что самка находит поющего на растении самца сна-
чала по звуку, а затем по вибрациям ветки, на которой он сидит (Latimer, Schatral,
1983). В некоторых случаях вибрационные сигналы, распространяющиеся по субстра-
ту, выполняют функцию призывных, например у Meconema thalassinum (De Geer,
1773) (Sismondo, 1980), или пре- и посткопуляционных, как у Siliquofera grandis
(Blanchard, 1853) (Korsunovskaya et al., 2020.) Это минимизирует риск нападения хищ-
ников, находящих жертву по издаваемому ею звуку.
Кузнечики воспринимают звук с помощью слуховых (тимпанальных) органов, рас-
положенных в проксимальном отделе передних голеней. Специализированными ви-
брочувствительными органами у насекомых, в том числе у кузнечиковых, являются
подколенные хордотональные органы (рис. 1, А). Помимо них вибрации могут вос-
принимать также рецепторы бедренного и тибио-тарзального хордотональных орга-
нов (см., например, обзор: Field, Matheson, 1998) и кампаниформные сенсиллы
(Kuehne, 1982). Однако виброчувствительность этих органов и отдельных рецепторов,
как правило, существенно ниже, чем у подколенных органов.
Ранее была предпринята попытка исследовать чувствительность к вибрациям у от-
дельных хордотональных рецепторов саранчи и кузнечика (Kuehne, 1982; Kalmring
et al., 1994, 1996), но авторы не смогли установить, в состав каких виброчувствитель-
ных органов они входят.
Наши предварительные результаты, полученные с помощью комбинированных
электрофизиологических и морфологических методов (Корсуновская, Жантиев, 2011),
показали, что часть рецепторов, отвечающих на вибрации, относится к слуховому ор-
гану и является, таким образом, бифункциональной.
В настоящей работе рассматривается чувствительность к звуку и вибрациям оди-
ночных рецепторов тимпанального (слухового) органа кузнечиков.
МАТЕРИАЛ И МЕТОДИКА
Эксперименты проводили на самцах и самках кузнечиков Tettigonia cantans (Fuessly, 1775).
В единичных опытах использовали Decticus verrucivorus (Linnaeus, 1758). Насекомых собирали
в Москве и ее окрестностях и содержали в лаборатории в кубических садках с размерами
30 × 30 × 30 см. В качестве корма использовали листья травянистых растений (одуванчика, кле-
вера, салата), кусочки фруктов, а также смесь овсяных хлопьев «Геркулес» и гаммаруса.
Электрофизиологическая установка для регистрации активности рецепторов тимпанального
органа состояла из изготовленного в лаборатории усилителя биопотенциалов, соединенного
с магнитофоном или через аналого-цифровой преобразователь Е-14440 (L-card, Россия) - с пер-
сональным компьютером. При необходимости физиологический сигнал фильтровали с помо-
щью низкочастотного фильтра, встроенного в усилитель, чтобы избежать влияния электрических
сетей переменного тока. Стимулирующая аппаратура была представлена аналоговым или циф-
ровым (программа SoundGen, DiSoft, Россия) звуковым генератором, широкополосным динами-
ческим громкоговорителем FT
17H Fostex (Япония) и вибростендом
11073 (Robotron
Messelektronik «Otto Schoen», ГДР). При использовании виртуального генератора установка
включала усилитель Pioneer А-10 (США), соединенный со звуковой картой (UR-12, Япония).
Нейронную активность и акустический стимул оцифровывали с помощью программ L-Graph
(L-card, Россия) или PowerGraph (DiSoft, Россия). Частота дискретизации составляла 10 кГц.
Стимуляцию осуществляли звуковыми или вибрационными посылками длительностью
524
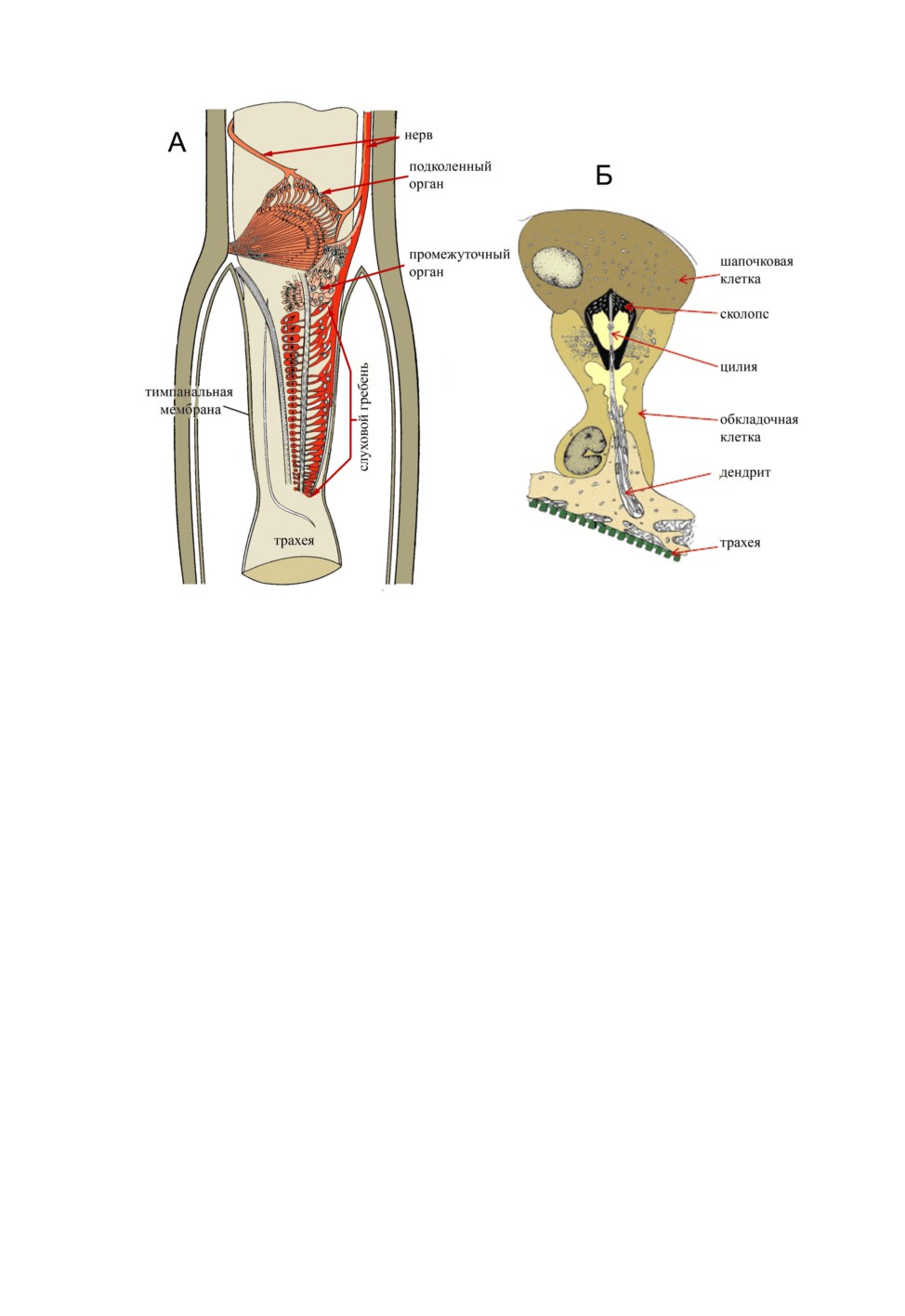
Рис. 1. Схема строения хордотональных органов голени кузнечика Decticus verrucivorus (L.)
(А) и сенсиллы слухового гребня (Б). По: Schwabe, 1906 (A); Жантиев, 1969 (Б).
40-80 мс со временем нарастания и падения амплитуды 10 мс. Частота предъявления раздражи-
теля составляла 1 с-1. Пороговой интенсивностью стимула считали такую, при которой рецептор
отвечал одним импульсом на один из трех последовательных стимулов. Препаратом для изуче-
ния реакций рецепторов на вибрационные и акустические раздражители была передняя нога
кузнечика, закрепленная так, чтобы бедро составляло с голенью угол в 90°, лапка касалась ви-
братора, а направление распространения вибрационной волны совпадало с продольной осью
голени. Для надежного контакта лапки с вибратором ее 1-й членик приклеивали маленькой
каплей смеси воска с канифолью; сочленение лапки с голенью оставалось подвижным, а шипы
и шпоры голени не касались вибратора. Угол между лапкой и голенью также составлял 90°.
В кутикуле на дорсальной поверхности голени проделывали небольшое отверстие, в которое
вводили стеклянный микроэлектрод с кончиком, заполненным 5%-ным водным раствором при-
жизненного люминесцентного красителя Lucifer Yellow CH, цилиндрическую часть микропи-
петки заполняли 1.5 М раствором LiCl. Регистрацию импульсной активности рецепторов
проводили внутриклеточно. Сопротивление кончика электрода составляло 10-40 мОм. По завер-
шении опыта препарат тимпанального органа помещали на предметный столик люминесцентно-
го микроскопа ML 40 Axioscop (ФРГ) и при необходимости фотографировали. Пороговые
значения уровня звукового давления и амплитуды вибраций измеряли в дБ. Для звука 0 дБ соот-
ветствовал 0.00002 H/м2, для вибраций - 0.00005 мм/c.
Эксперименты проводили в безэховой камере при температуре 22-26 °C. Для графического
представления и статистической обработки результатов использовали пакет программ Origin
6.1.6.
В работе содержатся результаты изучения 66 рецепторов.
525
РЕЗУЛЬТАТЫ
Слуховой орган кузнечиков состоит из двух отделов: промежуточного органа (pars
intermedia) и слухового гребня (crista acustica) (см. рис. 1, А). Сенсиллы первого из
них значительно меньше сенсилл слухового гребня, к которому они примыкают, и ле-
жат на трахее веерообразно, часть из них присоединена к кутикуле голени шапочковы-
ми клетками. Как ранее было показано (Жантиев, 1971; Жантиев, Корсуновская, 1978),
промежуточный орган воспринимает звуки в низкочастотном диапазоне от 1 до
12-14 кГц, причем чувствительность его рецепторов невелика: минимальные пороги
реакции на звуки оптимальных частот не опускались ниже 60 дБ.
Сенсиллы слухового гребня (рис. 1, Б) расположены на трахее, их шапочковые клет-
ки покрыты текториальной мембраной и не причленяются к кутикуле голени. Тела
рецепторных клеток расположены в небольшом углублении между трахеей голени и
тимпанальной мембраной, их аксоны образуют тимпанальный нерв, направляющийся
в бедро, а затем, через вертлуг и тазик, в первый грудной ганглий. Слуховой гребень
образован более чувствительными к звуку элементами. Для него характерна тонотопи-
ческая организация (Жантиев, Корсуновская, 1978; Oldfield, 1982, 1985; Stoelting,
Stumpner, 1998): проксимальные сенсиллы crista acustica наиболее чувствительны
к низкочастотным звукам, дистальные - к ультразвуку, а средние - к звукам промежу-
точных частот.
Различий в реакциях на стимулы между самцами и самками T. cantans нами обнару-
жено не было.
Частотные характеристики реакций сенсилл промежуточного органа
на звуковые и вибрационные стимулы
Изученные нами рецепторы реагировали на звук в диапазоне 1-12 (реже 14 кГц),
а их оптимальные частоты составляли 5-7 кГц.
Частотные характеристики ответов на вибрационные стимулы рецепторов промежу-
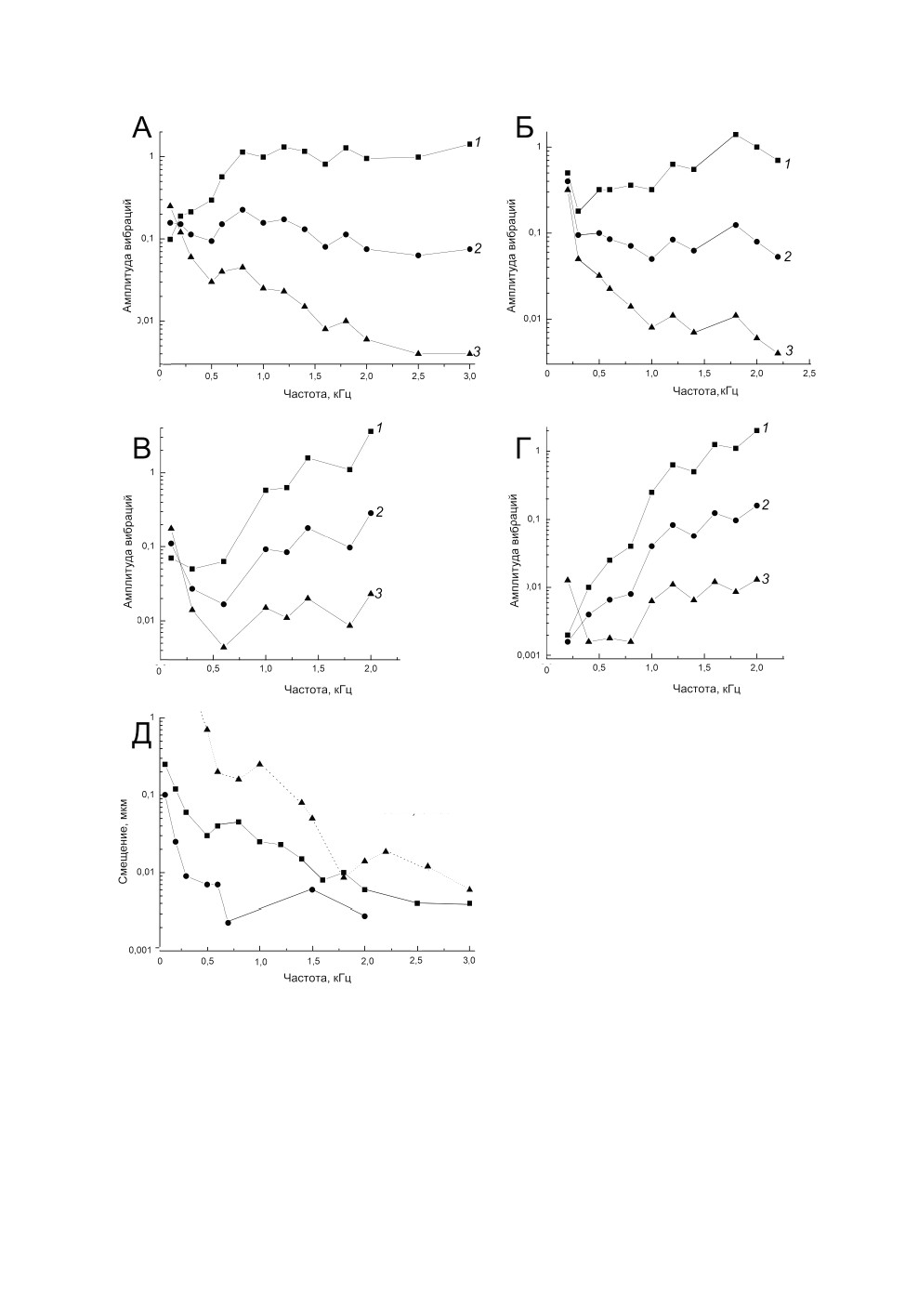
точного органа можно условно разделить на 3 группы (рис. 2). Первая характеризова-
лась тем, что реакция на ускорение (рис. 2, А, Б, 1) была максимальна на низких часто-
тах (в наших опытах на 0.1-0.2 кГц), а затем с ростом частоты стимула она постепенно
падала вплоть до 0.8 кГц, после чего либо сохранялась на относительно постоянном
уровне, либо продолжала снижаться. В то же время чувствительность этих рецепторов
к смещению (рис. 2, А, Б, 3) росла во всем исследованном диапазоне, а к скорости в
диапазоне 0.06-0.2 мм/с оставалась на примерно постоянном уровне (рис. 2, А, Б, 2).
Ко второй группе (рис. 2, В, Г) относятся частотно-пороговые кривые реакций, кото-
рые характеризуются минимальными значениями пороговых ускорений в низкоча-
стотном диапазоне. У таких кривых в полосе 0.2-0.8 кГц может быть выражен допол-
нительный минимум на 0.4-0.8 кГц на все параметры вибраций (рис. 2, В, 1-3) либо
только при реакциях на смещение (рис. 2, Г, 3).
Графики третьей группы демонстрируют повышение чувствительности рецепторов
ко всем параметрам вибраций с ростом их частоты. При этом на частотно-пороговых
кривых выделяется несколько минимумов: в области 0.6-0.8 или 1-1.2 кГц и
1.4-3 кГц. На рис. 2, Д приведены частотно-пороговые кривые реакций рецепторов на
смещение.
526
Рис. 2. Частотно-пороговые характеристики реакций на вибрационные стимулы рецепторов
промежуточного органа Tettigonia cantans (Fuessly).
A, Б - реакции на ускорение (1) (м/с2), скорость (2) (мм/с) и смещение (3) (мкм) рецепторов c308io (А)
и d269io (Б) группы 1; В, Г - то же рецепторов d209io (В) и c289io (Г) группы 2; Д - реакции
на смещение трех рецепторов группы 3.
527
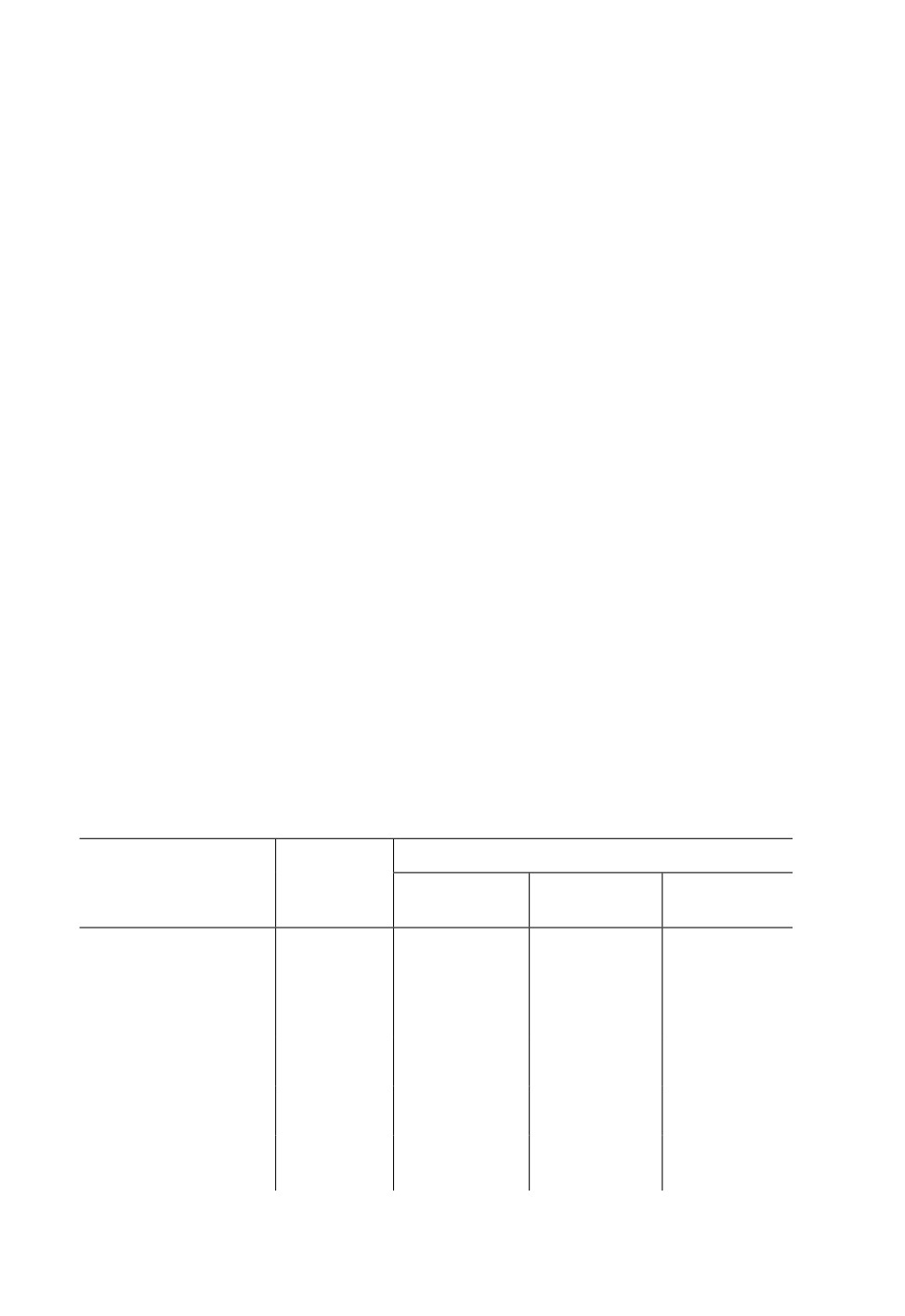
Минимальные пороговые значения ускорения, скорости и смещения, а также опти-
мальные частоты при восприятии вибраций рецепторами промежуточного органа от-
ражены в табл. 1.
Частотные характеристики сенсилл слухового гребня (рис. 3, А-В)
Максимальную чувствительность к звуку п р о к с и м а л ь н ы х р е ц е п т о р о в
слухового гребня регистрировали на частоте 8-10 кГц.
У большинства изученных проксимальных сенсилл на частотно-пороговых кривых
реакций на вибрации был выражен минимум в области 0.4-0.6 кГц. У двух рецепторов
максимум чувствительности регистрировался на 0.3 кГц (сенсилла № 9) и 1 кГц (сен-
силла № 5). Указанные оптимальные частоты отражались в виде минимумов на ча-
стотно-пороговых кривых при всех параметрах вибрационного стимула (скорости,
ускорения и смещения).
Как и у рецепторов промежуточного органа, помимо оптимума в диапазоне 0.1-
1 кГц наблюдалось повышение чувствительности к более высоким частотам. Так, на
2 кГц пороги по смещению у некоторых клеток составляли 0.0015-0.0030 мкм.
Регистрацию ответов на звуковые и вибрационные стимулы м е д и а л ь н о й ч а -
с ти слухового гребня регистрировали в зоне расположения его 10-14-й сенсилл. Эти
рецепторы были наиболее чувствительны к звукам частотой от 12-14 кГц (сенсиллы
№ 10-13).
Для клеток этой группы характерно относительно равномерное повышение чув-
ствительности к смещению по мере роста частоты стимула до 0.6-1 кГц (сенсилла
№ 11, рис. 3, Б, 1) и, нередко, выше - до 2 кГц (сенсилла № 14, рис. 3, Б, 2). На частот-
но-пороговых кривых реакций этих же сенсилл на скорость и/или ускорение имелись
дополнительные, значительно слабее выраженные минимумы в диапазоне 0.6-1.2 кГц.
Таблица 1. Оптимальные частоты и минимальные пороговые интенсивности вибраций,
воспринимаемых рецепторами тимпанального органа Tettigonia cantans (Fuess.). После значения
порога реакции в скобках приведена оптимальная частота рецептора (в кГц)
Слуховой гребень
Отделы тимпанального
Промежуточ-
органа
ный орган
Проксимальные
Медиальные
Дистальные
рецепторы
рецепторы
рецепторы
Зоны повышенной чув-
0.3;
0.2-0.3;
0.06-0.2;
0.1-0.3;
ствительности, кГц
0.4-0.8;
0.4-0.6;
0.5-0.8;
0.4-0.8;
1-1.2;
1-1.2;
2
1-1.2;
1.6 - >2
1.4-2.5
1.4-2
Минимальные пороги
0.03
(0.3)
0.005(0.4-0.6)
0.006
(0.8)
0.001
(0.8)
по ускорению, м/с2
Минимальные пороги
0.014 (1)
0.006
(0.6)
0.0015
(0.8)
0.008 (2)
по скорости, мм/с
Минимальные пороги
0.001
(1.8)
0.002
(0.6)
0.0002
0.0035
(1.6)
по смещению, мкм
(0.8 и 2)
528
Рис. 3. Частотно-пороговые характеристики реакций на вибрационные (слева) и звуковые
(справа) стимулы рецепторов проксимальной (А), средней (Б) и дистальной (В) частей
слухового гребня Decticus verrucivorus (L.) (A, 2) и Tettigonia cantans (Fuessly) (остальные
графики) и слуховой гребень Т. cantans с окрашенным 15-м рецептором (Г).
Одинаковыми цифрами обозначены реакции на звук и вибрации одного и того же рецептора.
Морфологически идентифицированные рецепторы: А - № 2 (1), № 4 (2), № 5 (4); Б - № 11 (1), № 12 (3),
№ 14 (2); В - № 15 (2), № 18 (5), № 20 (3).
529
Иногда медиальные сенсиллы обладали максимальной чувствительностью к вибраци-
ям частотой 0.6 либо 0.8-1 кГц как на смещение, так и на скорость или ускорение
(сенсиллы 10, 11). При этом форма их частотно-пороговых кривых приближалась
к V-образной (рис. 3, Б, 4).
Минимальные значения порогов реакции рецепторов данной группы приведены в
табл. 1.
Дис т а л ьна я ча с ть слухового гребня. Регистрацию ответов на акустические и
вибрационные стимулы проводили в дистальной трети crista acustica. Там располага-
ются рецепторы, более мелкие, чем лежащие проксимальнее, однако все еще образую-
щие ряд из одиночных сенсилл. Самые дистальные сенсиллы образуют двойной ряд,
тела их нейронов лежат очень компактно, а аксоны, входящие в тимпанальный нерв,
чрезвычайно коротки, что делает затруднительной регистрацию реакций одиночных
клеток. В связи с этим наши исследования ограничились сенсиллами, чьи оптималь-
ные частоты лежали в диапазоне от 20 до 40 кГц, что, согласно физиологическим и
морфологическим характеристикам, соответствует 15-30-й сенсиллам.
Чувствительность всех исследованных дистальных сенсилл слухового гребня к ви-
брациям характеризовалась следующими особенностями. Пики максимальной чув-
ствительности наблюдались в области как низких частот (до 1 кГц) (см., например,
рис. 3, В, 1, 5, 6), так и за пределами 1 кГц (рис. 3, В, 2-6). В первом случае оптималь-
ными частотами преимущественно были 0.6, 0.8 и 1 кГц, во втором - около 2 кГц,
у некоторых клеток наблюдался единственный или дополнительный максимум чув-
ствительности на частоте 1.2 или 1.4 кГц (см. табл. 1). Форма частотно-пороговых
кривых реакций на все параметры вибраций была сходной, однако степень выражен-
ности максимумов различалась. Минимальные пороги наиболее чувствительных ре-
цепторов приведены в табл. 1. У большинства сенсилл они по ускорению не превыша-
ли 0.9 м/с2 и лежали в диапазонах 0.008-0.07 мм/с по скорости и 0.004-0.02 мкм - по
смещению.
Амплитудные характеристики сенсилл тимпанального органа
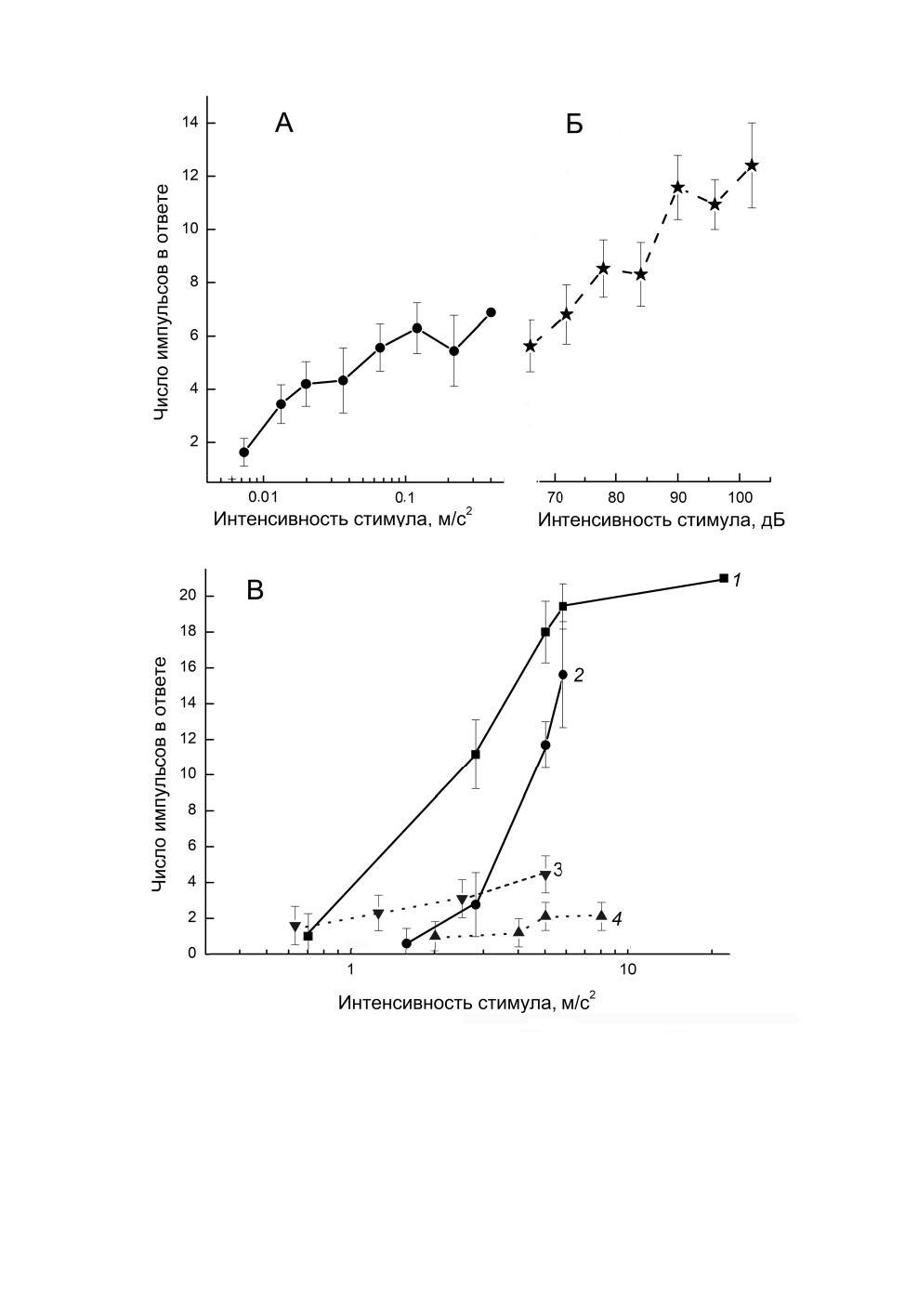
В результате исследования зависимости ответов рецепторов промежуточного органа
и слухового гребня от интенсивности стимула получены следующие данные. Ответы
одного и того же нейрона на звук и вибрации различались по максимальному числу
импульсов. Почти у всех изученных рецепторов тимпанального органа при стимуля-
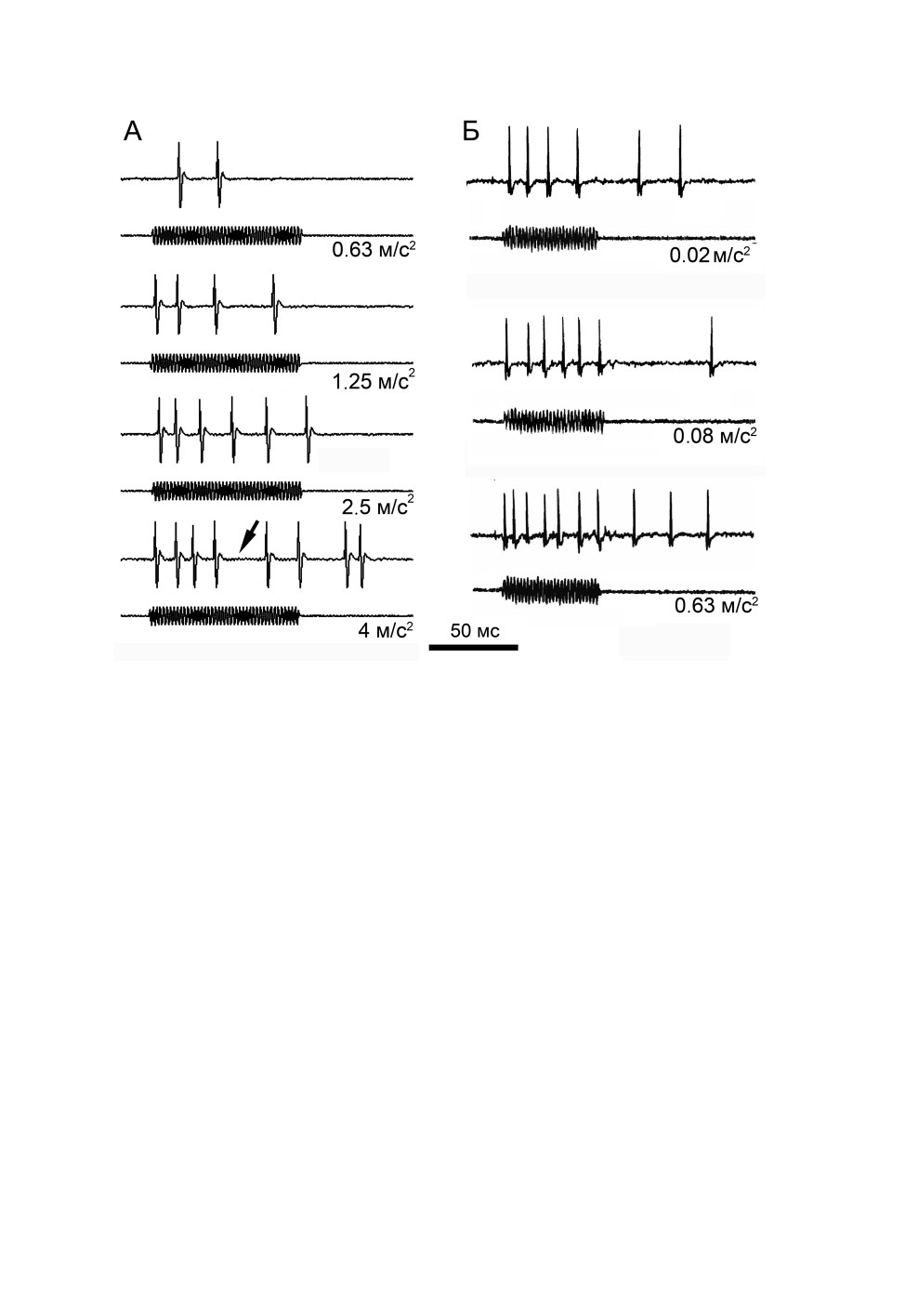
ции звуком оно было больше (рис. 4, А, Б). Реакции тонические, при высокой интен-
сивности как звукового, так и вибрационного стимулов развивалось последействие
(рис. 5). В некоторых опытах у рецепторов наблюдались паузы в ответном разряде
(рис. 5, А), которые были описаны нами ранее (Жантиев, Корсуновская, 1997). Насы-
щения в ответах на вибрации выявлено не было (рис. 4, Б, В). У ряда клеток импульса-
ция в ответ на вибрационные стимулы слабо менялась при повышении их амплитуды
(рис. 4, В, 3, 4).
ОБСУЖДЕНИЕ
К настоящему времени накопились данные о влиянии на реакции виброчувствитель-
ных органов ряда факторов (см. обзор: Strauß et al., 2019), в связи с этим большое зна-
чение имеют особенности подготовки препарата и методика экспериментов. В нашем
530
Рис. 4. Зависимость ответных реакций одиночных слуховых рецепторов Tettigonia cantans
(Fuessly) от интенсивности вибрационного и звукового стимулов.
А - рецептора № 5 слухового гребня на вибрации частотой 0.6 кГц, Б - того же рецептора на звук частотой
8 кГц; длительность стимула 55 мс; В - промежуточного органа: 1 и 2 - рецептора с298io
на вибрации частотой 1.6 кГц (1) и 0.5 кГц (2), длительность стимула 80 мс; 3 и 4 - рецептора b99io
на вибрации частотой 0.2 кГц (3) и 0.4 кГц (4), длительность стимула 40 мс.
Приведены средние значения и среднеквадратичные отклонения.
531
Рис. 5. Реакции на вибрации слуховых рецепторов Tettigonia cantans (Fuessly):
А - промежуточного органа, Б - рецептора слухового гребня.
Частота стимула 1.6 кГц (А) и 0.6 кГц (Б). Стрелкой отмечена пауза в ответе.
исследовании опыты проводили с отделенной от тела передней ногой кузнечиков, ко-
торая касалась вибратора так, что направление распространения вибрационной волны
совпадало с продольной осью голени, составлявшей с бедром угол 90о. Ранее было
установлено, что амплитуда механического ответа кутикулы голени зависит от направ-
ления распространения стимулирующих вибраций - вертикального или горизонталь-
ного, однако, частотная избирательность этого ответа не меняется.
Максимальную амплитуду механического ответа голени пещерного кузнечика
Troglophilus neglectus Krauss, 1879 регистрировали при вертикальном распростране-
нии вибраций, совпадающем с продольной осью голени. Не исключено, что на реак-
ции виброрецепторов Tettigonia cantans может оказывать влияние также угол между
бедром и голенью. Свидетельством этого служит максимальный механический ответ
ноги на вибрации у Troglophilus neglectus, когда данный угол составляет 45о (Stritih-
Peljhan, Strauß, 2018). Во время ходьбы насекомого угол между бедром и голенью по-
стоянно меняется. Отсутствие дополнительной нагрузки (в нашем случае - веса тела
исследуемого насекомого) также может влиять на реакции виброрецепторов. Извест-
но, что у сверчка нагрузка на свободно стоящую заднюю ногу несколько повышает
532
чувствительность к вибрациям частотой до 0.5 кГц и снижает ее в диапазоне 0.7-
2 кГц, но не меняет положения оптимума на частотно-пороговой кривой (Dambach,
1972). Судя по полученным нами данным, при работе с отделенной от тела кузнечика
ногой может незначительно измениться соотношение минимумов на частотно-порого-
вых кривых виброрецепторов, но частотная настройка сенсилл должна сохраниться.
Отсутствие тела насекомого в наших опытах может оказывать влияние на частотные
характеристики рецепторов при восприятии звука. Однако результаты экспериментов,
предшествовавших настоящей работе (наши неопубликованные данные), соответству-
ют выявленным ранее фактам (Чуканов, Жантиев, 1978), согласно которым акустиче-
ские трахейные мешки в переднегруди у Tettigonia cantans обеспечивают лишь повы-
шение чувствительности слухового органа преимущественно в ультразвуковой
области, не оказывая существенного влияния на его частотную настройку: оптималь-
ные частоты тимпанального органа как в отделенной от тела ноге, так и у интактного
насекомого лежат в диапазоне 12-20 кГц. При этом чувствительность высокочастот-
ных рецепторов к ультразвуку в отделенной от тела ноге остается на достаточно высо-
ком уровне (рис. 3, В, 1, 6).
Таким образом, угол между бедром и голенью, направление распространения вибра-
ций, положение ноги на субстрате могут оказывать влияние на виброчувствительность
рецепторов тимпанального органа, однако оно, по-видимому, незначительно и не ме-
няет частотных характеристик хордотональных сенсилл. Наиболее стабильны реак-
ции на вибрации при разных условиях стимуляции, вероятно, у субинтегументных
сенсилл слухового гребня и промежуточного органа, которые воспринимают вибра-
ции кутикулы, передающиеся опосредованно через гемолимфу ноги - по трахее под
сенсиллами и по текториальной мембране над ними. Кроме того, следует принять во
внимание, что насекомое во время восприятия вибрационных стимулов перемещается
по разным субстратам, поверхность которых расположена под разными углами к про-
дольной оси голени, а нога во время движения сгибается и разгибается, поэтому усло-
вия стимуляции, крепления к вибратору и положение препарата во время наших экс-
периментов полностью соответствуют естественным.
Ранее было отмечено, что на частотную настройку и чувствительность виброрецеп-
торов влияет положение ноги насекомого (Stritih-Peljhan, Strauss, 2018; Strauss et al.,
2019), однако наличие у кузнечиков рецепторов, чувствительность которых в широком
диапазоне не зависит от частоты (см. рис. 2, А, Б), позволяет движущемуся насекомо-
му получать достоверную информацию о положении источников разнообразных ви-
браций. Кроме того, наличие в тимпанальном органе рецепторов с разными оптималь-
ными частотами при восприятии скорости, ускорения и смещения позволяет
предположить, что вибрационная чувствительность кузнечика обеспечивает его реак-
цию на широкий спектр механических колебаний, распространяющихся по любым
поверхностям и в любых субстратах.
Сравнение частотно-пороговых характеристик реакций на вибрационные стимулы
рецепторов тимпанального органа показало, что оптимальные частоты этих нейронов
лежат преимущественно в диапазонах 0.1-1 кГц и 2-3 кГц. Повышенная чувствитель-
ность бифункциональных сенсилл обеспечивает восприятие как биотических и абио-
тических шумов (обычно они занимают низкочастотный диапазон до 0.2 кГц), так
и вибрационных компонентов конспецифических звуковых сигналов. У Tettigonia
cantans они расположены в диапазоне от 30 Гц до 5 кГц. Таким образом, тимпаналь-
533
ный орган кузнечика - это эквипотенциальный орган, который наряду со звуком мо-
жет воспринимать любые вибрации.
В промежуточном органе и слуховом гребне были выявлены элементы с разной чув-
ствительностью к параметрам вибраций: ускорению, скорости и смещению, а также
рецепторы с различными оптимальными частотами. Данные различия могут быть обу-
словлены факторами, действующими как по отдельности, так и совместно. Рецепторы
тимпанального органа располагаются на трахее, сверху их прикрывает текториальная
мембрана. Некоторые сенсиллы промежуточного органа являются интегументными,
прикрепленными к кутикуле голени. Большая часть рецепторов слухового гребня рас-
положена в ряд, и размеры рецепторов к его дистальному концу уменьшаются; самые
мелкие, дистальные сенсиллы лежат на мембране попарно. Угол наклона сколопсов
меняется на протяжении слухового гребня (Kalmring et al., 1993). Перечисленные осо-
бенности как строения, так и положения бифункциональных рецепторов слухового
органа кузнечика, несомненно, могут влиять на их функциональные свойства.
Проводя анализ чувствительности рецепторов к различным параметрам вибрацион-
ного стимула, на наш взгляд, полезно сравнить их с техническими устройствами
для измерения и анализа вибраций. Они представлены разными видами датчиков:
велосиметрами, акселерометрами и датчиками смещения. В зависимости от целей ис-
следования применяют разные устройства: для анализа низкочастотных вибраций
используют датчики смещения, высокочастотных - акселерометры, промежуточных
частот - велосиметры. Все эти устройства основаны на разных принципах действия.
Проводя аналогию с виброрецепторами насекомых, можно предположить, что кузне-
чики также получают информацию о вибрациях, основываясь на реакциях рецепторов
на разные параметры этих механических колебаний.
При этом механо-сенсорная передача осуществляется в результате функционирова-
ния механозависимых ионных каналов на мембране дендрита, испытывающей дефор-
мацию. Однако механическое воздействие на мембрану могут оказывать непосред-
ственно или опосредованно разные элементы. Хордотональная сенсилла представляет
собой комплекс из нескольких клеток, каждую из которых можно рассматривать как
механическую систему. При восприятии вибраций они подвергаются воздействию
вибрационной волны непосредственно (например, шапочковые клетки подколенного
органа) и (или) через контактирующие с гемолимфой шапочковые клетки субинтегу-
ментных сенсилл, обкладочные клетки и тела нейронов. Кроме того, эти клетки могут
обладать собственными резонансами и реагировать на резонансные колебания участка
трахеи, на котором расположена сенсилла. Поэтому хордотональная сенсилла может
сочетать в себе функции как приемника смещения, так и акселерометра за счет разли-
чий в механических свойствах между ее клетками и окружающими структурами. Кос-
венным подтверждением правомочности этой гипотезы могут служить частотно-поро-
говые кривые реакций некоторых рецепторов промежуточного органа на ускорение:
на них после роста пороговых ускорений с повышением частоты стимула наблюдают-
ся плато, а также зоны повышенной чувствительности у ряда рецепторов тимпаналь-
ного органа к определенным частотам.
Бифункциональные рецепторы промежуточного органа и слухового гребня облада-
ют высокой чувствительностью к механическим колебаниям в широком частотном ди-
апазоне. Повышенная чувствительность к высокочастотным вибрационным стимулам
позволяет насекомым воспринимать вибрационную компоненту коммуникационных
534
звуковых сигналов или конспецифические вибрационные сигналы, которые издают,
например, представители изученных нами видов прямокрылых (Benediktov et al.,
2020; Korsunovskaya et al., 2020). Абиотические шумы и вибрации, сопровождающие
локомоцию или иные проявления жизнедеятельности других животных, как правило,
лежат в низкочастотном диапазоне - 0.1-2 кГц. Они также вызывают реакции соответ-
ствующих рецепторов даже при чрезвычайно низких уровнях их амплитуды.
Ранее дифференцированные реакции на разные параметры вибраций были обнару-
жены у рецепторов бедренного хордотонального органа палочников и саранчи (см. об-
зоры: Field, Matheson, 1998; Eberl et al., 2016), а также слуховых органов саранчи и
кузнечика (Kuehne, 1982). Однако различия в функциях бедренного хордотонального
органа, основная функция которого - контроль движения ноги в коленном суставе, и
слухового тимпанального органа не позволяют проводить прямые аналогии, которые
помогли бы вскрыть причины этого явления. Результаты исследования Р. Кюне
(Kuehne, 1982), как упоминалось ранее, также не способствуют пониманию особенно-
стей восприятия вибраций рецепторами, так как принадлежность последних осталась
не выясненной. Следует также иметь в виду, что механизмы механо-сенсорной пере-
дачи в дендритах хордотональных рецепторов насекомых все еще далеки от понима-
ния, поэтому мы можем лишь указать на некоторые особенности этих сенсилл, воз-
можно, влияющие на данные процессы.
Наличие у насекомых бифункциональных механорецепторов - хордотональных сен-
силл слухового органа кузнечиков - существенно расширяет возможности анализа
биологически важных стимулов. Это повышает эффективность взаимодействия
конспецифических особей (например, во время брачного поведения) и вероятность
выживания при опасности нападения хищника. Наличие таких адаптаций, наряду
с другими особенностями морфо-функциональной организации, позволяет насекомым
быть одной из наиболее процветающих групп живых организмов на Земле.
БЛАГОДАРНОСТИ
Авторы выражают благодарность Л. С. Шестакову (Институт проблем передачи ин-
формации РАН) за помощь в калибровке вибро-акустической аппаратуры и М. В. Ива-
нову (Московский государственный технический университет им. Н. Э. Баумана) за
обсуждение результатов работы.
ФИНАНСИРОВАНИЕ
Работа выполнена по государственным темам
121032300064-0, ААА-
А-А19-119020590085-0 и при финансовой поддержке Российского фонда фундамен-
тальных исследований (грант № 19-04-00104а).
СПИСОК ЛИТЕРАТУРЫ
Жантиев Р. Д. 1969. Ультраструктурная организация механорецепторных сенсилл насекомых. Журнал общей
биологии 30 (2): 224-230.
Жантиев Р. Д. 1971. Частотные характеристики тимпанальных органов кузнечиков (Orthoptera, Tettigoniidae).
Зоологический журнал 50 (4): 507—514.
Жантиев Р. Д. 1981. Биоакустика насекомых. М.: Издательство МГУ, 256 с.
Жантиев Р. Д., Корсуновская О. С. 1978. Морфофункциональная организация тимпанальных органов кузне-
чика Tettigonia cantans (Orthoptera, Tettigoniidae). Зоологический журнал 57 (7): 1012-1016.
535
Жантиев Р. Д., Корсуновская О. С. 1997. Подавление импульсной активности рецепторов тимпанального ор-
гана кузнечиков (Orthoptera, Tettigoniidae). Сенсорные системы 11 (2): 118-127.
Корсуновская О. С., Жантиев Р. Д. 2011. Реакции рецепторов тимпанального органа кузнечиков на вибраци-
онные стимулы. В кн.: Материалы Международной научной конференции «Фундаментальные про-
блемы энтомологии в XXI веке». Санкт-Петербург,
16-20 мая
2011 г. СПб.: Издательство
Санкт-Петербургского университета, с. 76.
Чуканов В. С., Жантиев Р. Д. 1978. Влияние акустических трахей на функциональные характеристики
слуховых органов кузнечиков Tettigonia cantans Fuess. (Orthoptera, Tettigoniidae). Вестник
МГУ. Серия биология 4: 43-47.
Benediktov A. A., Korsunovskaya O. S., Polilov A. A., Zhantiev R. D. 2020. Unusual mechanism of emission of
vibratory signals in pygmy grasshoppers Tetrix tenuicornis (Sahlberg, 1891) (Orthoptera: Tetrigidae). Science
Dambach M. 1972. Der Vibrationssinn der Grillen. I. Schwellenmessungen an Beinen frei beweglicher Tiere.
Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology 79 (3):
Eberl D. F., Kamikouchi A., Albert J. T. 2016. Auditory transduction. In: G. Pollack, A. Mason, A. Popper, R. Fay
(eds). Insect Hearing. Springer Handbook of Auditory Research, vol. 55. Cham: Springer, p. 159-175.
Field L. H., Matheson T. 1998. Chordotonal organs of insects. In: P. D. Evans (ed.). Advances in Insect Physiology.
Kalmring K., Hoffmann E., Jahto M., Sickmann T., Grossbach M. 1996. The auditory-vibratory sensory system of
the bushcricket Polysarcus denticauda (Phaneropterinae, Tettigoniidae) II. Physiology of receptor cells.
Journal of Experimental Zoology
276
(5):
010x(19961201)276:5%3C315::aid-jez2%3E3.0.co;2-r
Kalmring K., Rössler W., Ebendt R., Ahi J., Lakes R. 1993. The auditory receptor organs in the forelegs of
bushcrickets: physiology, receptor cell arrangement, and morphology of the tympanal and intermediate
organs of three closely related species. Zoologische Jahrbücher. Abteilung für allgemeine Zoologie und
Physiologie der Tiere 97 (1): 75-94.
Kalmring K., Rössler W., Unrast C. 1994. Complex tibial organs in the fore-, mid- and hindlegs of the bushcricket
Gampsocleis gratiosa (Tettigoniidae): Comparison of physiology of the organs. Journal of Experimental
Korsunovskaya O., Berezin M., Heller K.-G., Tkacheva E., Kompantseva T., Zhantiev R. 2020. Biology, sounds and
vibratory signals of hooded katydids (Orthoptera: Tettigoniidae: Phyllophorinae). Zootaxa 4852: 309-322.
Kuehne R. 1982. Neurophysiology of the vibration sense in locusts and bushcrickets: response characteristics of
single receptor units. Journal of Insect Physiology
28
1910(82)90123-8
Latimer W., Schatral A. 1983. The acoustic behaviour of the bushcricket Tettigonia cantans. I. Behavioural responses
3
Oldfield B. P. 1982. Tonotopic organisation of auditory receptors in Tettigoniidae (Orthoptera: Ensifera). Journal of
Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology 147 (4): 461-469.
Oldfield B. P. 1985. The tuning of auditory receptors in bush crickets. Hearing Research 17 (1): 27-35. https://doi.
org/10.1016/0378-5955(85)90126-1
Schwabe J. 1906. Beiträge zur Morphologie und Histologie der tympanalen Sinnesapparate der Orthopteren.
Zoologica, Stuttgart 50: 1-154.
Sismondo E. 1980. Physical characteristics of the drumming of Meconema thalassinum. Journal of Insect Physiology
Stoelting H., Stumpner A. 1998. Tonotopical organization of auditory receptors of the bushcricket Pholidoptera
griseoaptera (De Geer 1773) (Tettigoniidae, Decticinae). Cell and Tissue Research 294 (2): 377-386. https://
doi.org/10.1007/s004410051187
Strauß J., Stritih-Peljhan N., Lakes-Harlan R. 2019. Determining vibroreceptor sensitivity in insects: the
influence of experimental parameters and recording techniques. In: P. Hill, R. Lakes-Harlan, V. Mazzoni,
P. Narins, M. Virant-Doberlet, A. Wessel (eds). Biotremology: Studying Vibrational Behavior. Animal
22293-2_11
Stritih-Peljhan N., Strauß J. 2018. The mechanical leg response to vibration stimuli in cave crickets and implications
for vibrosensory organ functions. Journal of Comparative Physiology A: Neuroethology, Sensory, Neural,
536
Virant-Doberlet M., Kuhelj A., Polajnar J., Šturm R. 2019. Predator-prey interactions and eavesdropping in
vibrational communication networks. Frontiers in Ecology and Evolution. Behavioral and Evolutionary
Ecology 7: article 203: 1-15. doi: 10.3389/fevo.2019.00203.
Yack J. 2016. Vibrational signaling. In: G. S. Pollack, A. C. Mason, A. N. Popper, R. R. Fay (eds). Insect Hearing.
FUNCTIONS OF CHORDOTONAL SENSILLA IN BUSHCRICKETS
(ORTHOPTERA, TETTIGONIIDAE)
R. D. Zhantiev, O. S. Korsunovskaya
Key words: bushcrickets, tympanal organ, bifunctional auditory receptors, vibrational
sensitivity.
SUMMARY
Using electrophysiological methods, the acoustic and vibrational sensitivity of identified single
auditory receptors in bushcrickets has been studied. In the intermediate organ, neurons were identified
whose response to acceleration in a significant frequency range does not depend on the frequency
of the stimulus; along with them, there are cells demonstrating increased sensitivity to frequencies
of 0.4-0.8 kHz (per displacement), and (or) 0.1-0.3; 1-1.2 kHz, and 1.4-3 kHz for all vibration
parameters. In addition, most of the studied receptors have a zone of increased sensitivity to high-
frequency vibrations (1.5-2.5 kHz). In sensilla of the crista acustica, increased sensitivity is recorded
in the range of 0.1-0.3, 0.4-0.8, 1-1.2, and 1.4-2.5 kHz. One and the same sensillum can have optimal
frequencies in different ranges for the perception of different vibration parameters. Such differences in
sensitivity to vibration acceleration, vibration velocity and displacement, as well as different optimal
frequencies of the receptors of the intermediate organ and the crista acustica, can be due to differences
in the size, position and details of the structure of the sensilla, their own resonances and reactions to
resonance vibrations of the trachea site, on which vibroreceptors lie. Thus, chordotonal sensillum is
a bifunctional mechanoreceptor, which, along with auditory sensitivity, can combine the functions of
both a displacement receiver and an accelerometer due to differences in the mechanical properties
between its cells and surrounding structures.
537