ЭНТОМОЛОГИЧЕСКОЕ ОБОЗРЕНИЕ, 100, 4, 2021
УДК 595.7-153.11
ОСОБЕННОСТИ РАЗВИТИЯ ОСЕННИХ ПОКОЛЕНИЙ
И ОТКЛАДКИ ЯИЦ У METOPOLOPHIUM DIRHODUM WALK.
(HEMIPTERA, STERNORRHYNCHA: APHIDIDAE)
© 2021 г. А. Б. Верещагина,* Е. С. Гандрабур**
Всероссийский научно-исследовательский институт защиты растений
Шоссе Подбельского, 3, С.-Петербург, Пушкин, 196608 Россия
*e-mail: aphidabver@gmail.com, **e-mail: helenagandrabur@gmail.com
Поступила в редакцию 19.07.2021 г.
После доработки 4.08.2021 г.
Принята к публикации 4.08.2021 г.
В течение трех лет изучалось развитие осенних поколений популяции тли Metopolophium
dirhodum Walk. в окрестностях С.-Петербурга. Отмечена способность этого вида развиваться на
Rosa rugosa Thunb., R. canina L. и R. glauca Pourr. Впервые установлено, что наряду с ремиграци-
ей M. dirhodum на шиповники развитие и репродукция овипар могут происходить одновременно
и на вторичном хозяине (пшенице). Место откладки яиц тлями на шиповниках различается в
зависимости от видовых особенностей растений: на R. rugosa и R. canina яйца откладывают-
ся преимущественно около шипов или на шипах (96.3 % и не менее 70.5 % соответственно).
На бесшипном R. glauca - только на листьях. Опушенные и железистые листья R. rugosa имаго
овипар не заселяли. При ремиграции с пшеницы овипары предпочитали заселять и откладывать
яйца на R. glauca, а не на R. rugosa (F = 12.9, p = 0.002 и F = 12.3, p = 0.003 соответственно),
однако весной из яиц на листьях R. glauca основательницы не появились, а на шипах R. canina
выжило около 80 % яиц. Отмечена способность тлей питаться на опавших листьях и молодых
стеблях шиповника до наступления устойчивых заморозков (˗5…˗9 °C). Погодные изменения
осенью, в том числе понижение среднесуточной температуры на 5.1°, вызвали увеличение про-
должительности преимагинального развития самцов (F = 20.9, p = 0.00002) и растянутость их
лёта. Определены некоторые параметры развития гинопар. Выявленные особенности осенних
поколений M. dirhodum могут быть использованы при планировании мер контроля численности
данного вредителя и в селекции шиповника.
Ключевые слова: тли, ремигранты, овипары, яйца, шиповник, шипы, листья, распределение,
вторичный хозяин, питание.
DOI: 10.31857/S036714452104002X
Тли - широко известные вредители зерновых культур. Всего среди них отмечено
10 вредящих злакам видов (Бокина, 2009), из которых черемухово-злаковая тля
[Rhopalosiphum padi (L.)], большая злаковая тля (Sitobion avenae F.) и розанно-злаковая
тля (Metopolophium dirhodum Walk.) имеют наибольшее экономическое значение для
многих стран и территорий, в том числе и для Северо-Запада России. Metopolophium
dirhodum реже доминирует на злаковых посевах, чем Rh. padi или S. avenae (Берим,
708
2008), и поэтому меньше привлекает внимание специалистов, из-за чего многие де-
тали развития этого вида остаются неясными. Однако в Центральной Европе, Се-
верной и Южной Америке M. dirhodum относится к массовым вредителям (Honěk et
al., 2018). Крупные вспышки численности этого вида в Англии (1979 г.) и Шотландии
(1982 г.) выявили большой потенциал его вредоносности на зерновых культурах
(Howard, Dixon, 1992). Питание M. dirhodum на флаговом и предфлаговом листьях, где
формируются ассимиляты, питающие колос, развитие вторичной инфекции на ее экс-
крементах и перенос вирусов вызывают существенное снижение продуктивности
растений (van Emden, Harrington, 2007). Если на юге ареала M. dirhodum живет аноло-
циклически (без обоеполого размножения) только на вторичных хозяевах - представи-
телях порядка Poales, то в более северной части - голоциклически (с обоеполым раз-
множением) и сезонной сменой первичных (зимних, древесных) и вторичных (летних,
травянистых) хозяев, т. е. двудомно. В качестве первичных хозяев указываются раз-
личные виды рода Rosa L. (Alford, 2012) и наиболее часто упоминаются R. rugosa и
R. canina (Lubiarz, Cichocka, 2014). На шиповнике тли зимуют в фазе яйца. Весной из
яиц выходят основательницы, способные питаться только на первичном хозяине (Ма-
монтова, 1973; Williams, Dixon, 2007). Потомки основательниц в мае - начале июня
мигрируют на сельскохозяйственные и дикорастущие злаки, а осенью ремигранты (ги-
нопары и самцы) возвращаются на шиповник. Гинопары партеногенетически воспро-
изводят овипар, которые после спаривания откладывают яйца (Honěk, Martinkova,
2004; Krzyźanowski, 2017). В Польше отмечено анолоциклическое развитие тли в те-
плицах только на розах (Kmieć, 2006). Значимость повреждения шиповников опреде-
ляется их широким использованием в озеленении, как лекарственных и пищевых
растений. В России широко ведется селекция садового шиповника, в том числе с по-
вышенной устойчивостью к болезням и вредителям (Ильин, 2017). Наиболее значи-
мыми для культивирования и селекции считаются R. rugosa, R. canina и R. glauca, осо-
бенно бесшипная форма (Евтухова, 2016). Особенности развития осенних поколений
и вредоносность тлей на шиповниках изучены еще недостаточно.
Целью работы было выявление особенностей развития, ремиграции и откладки яиц
у M. dirhodum на первичных хозяевах в условиях северо-запада европейской части
России.
МАТЕРИАЛ И МЕТОДИКА
Основной объект исследования - тля M. dirhodum с гетероцийным голоциклическим циклом
развития, обитающая в окр. С.-Петербурга. Модельные опыты выполняли в течение 3 лет в про-
ветриваемых вегетационных помещениях без отопления. В качестве растений-хозяев использо-
вали изолированные побеги или трехлетние саженцы R. rugosa, R. canina и R. glauca.
Rosa rugosa (роза морщинистая) - один из самых распространенных видов шиповников в го-
родах и пригородах. Ветви растений опушены и густо покрыты разнотипными шипами; листья
крупные, темно-зеленые, морщинистые, с короткостебельчатыми желёзками и густым (особенно
по жилкам) опушением с адаксиальной стороны. Вегетация начинается в первой декаде апреля.
Характеризуется продолжительным цветением, зимостойкостью и засухоустойчивостью.
Листопад заканчивается во второй половине октября - ноябре (Евтухова, 2016; Плантариум…,
2007-2021).
709
Rosa glauca (= R. rubrifolia Vill., роза сизая, краснолистная) также широко используется в озе-
ленении. Характеризуется красновато-коричневыми, практически бесшипными ветвями; листья
красновато-зеленые, снизу по главной жилке шиповатые, сложные - состоят из 5-7 листочков.
Относится к видам с ранним началом вегетации. Цветение в июне, обычно продолжается две
недели. Листопад заканчивается немного раньше, чем у R. rugosa. Rosa glauca зимостоек, не
дает корневых отпрысков (Евтухова, 2016; Gardenia.RU, 2004-2021).
Rosa canina (роза собачья) - кустарник с обильными шипами на многочисленных корневых
отпрысках и более редкими - на главных побегах. Листья гладкие или иногда на адаксиальной
стороне по жилкам с редкими железками. Начало вегетации приходится на первые числа апреля.
Цветет в мае-июне. Листопад заканчивается в начале ноября или раньше, как у R. rugosa (Евту-
хова, 2016; Плантариум…, 2007-2021).
Саженцы шиповников были приобретены в ООО «Павловский питомник» и содержались
в пластиковых сосудах диаметром 34 см. Вторичным хозяином служила яровая мягкая пшеница
Triticum aestivum L. сорт Ленинградская 6. Пшеницу высевали в сосуды диаметром 14 см по
10 семян и в фазе кущения заселяли тлями. Тлей воспитывали под изоляторами из прозрачной
ткани в течение всего сезона.
В первый год исследований (2018) была поставлена задача изучения миграции и откладки яиц
осенними поколениями тлей на изолированные побеги R. rugosa и R. glauca.
В опыте использовали 9 клонов M. dirhodum, собранных на посевах пшеницы в Пушкинском
р-не С.-Петербурга и Гатчинском р-не Ленинградской обл. в первой декаде июля. Каждый клон
содержали в отдельном сосуде под изолятором.
Через каждые 2-3 недели в отдельных помещениях насекомых пересаживали на свежие рас-
тения пшеницы и вновь закрывали изоляторами. Перед началом осенней миграции в конце сен-
тября в каждом сосуде обитало около 200 тлей. Второго октября в сосуды с пшеницей ставили по
одному однолетнему побегу R. rugosa и R. glauca длиной 20-21 см на расстоянии 12 см от центра
сосуда. Учеты количества крылатых ремигрантов, имаго овипар и яиц на шиповнике проводили
5, 10 и 17 октября. Во второй учет были поставлены свежие побеги, а тли с неопавших листьев
были пересажены на свежие. Как черенки R. rugosa, так и опавшие листья R. glauca с яйцами
были оставлены в сосудах. В последнем учете подсчитаны также крылатые тли, имаго овипар и
яйца на пшенице и тли в верхней части садков. Бескрылых партеногенетических самок и ли-
чинок не учитывали. После учетов листья R. glauca с яйцами тлей были вынесены на улицу и
помещены у основания кустов шиповника. Весной 2019 г. проверено наличие личинок основа-
тельниц.
Во второй год исследований (2019) в отличие от предыдущего года было изучено поведение
осенних поколений и откладка яиц M. dirhodum на трехлетних саженцах (n = 3) R. canina. Для
этого в период массовой откладки яиц (15 октября) к каждому из саженцев в пластиковых со-
судах диаметром 34 см помещали по 2 сосуда (диаметр 9 см) с пшеницей, в массе заселенной
тлями, и накрывали общим садком. Растения шиповника имели шипы лишь в нижней части
стеблей. Основной учет отложенных яиц был проведен 22 ноября, далее наблюдения проводи-
лись до полной гибели насекомых 16 января. Регистрировали локализацию и численность яиц.
Яйца были оставлены на зимовку, а весной подсчитано количество появившихся личинок осно-
вательниц. 30 личинок были пересажены на всходы пшеницы, и было подсчитано количество
погибших особей.
Были установлены также некоторые параметры развития ремигрантов, определяющие чис-
ленность зимующих яиц. Для этого 18 сентября в пяти изолированных сосудах с пшеницей были
оставлены только личинки тлей. Через сутки в верхнюю часть садков мигрировали особи, пере-
линявшие на имаго. Садки с крылатыми тлями были надеты на сосуды с тремя побегами ши-
повника R. glauca в каждом. Через 6 дней был проведен учет количества гинопар и родившихся
овипар, самцы были удалены, а гинопары (103 особи) пересажены на свежие побеги шиповника.
710
Дальнейшие учеты и пересадки гинопар производили по мере увядания побегов шиповника.
Отмечали продолжительность жизни имаго и реализованную плодовитость гинопар.
Для определения срока преимагинального развития самцов 17 сентября и 25 октября от три-
дцати бескрылых самок с вторичного хозяина изолировали однодневное потомство в пяти со-
судах с пшеницей. Сразу после линьки потомства на имаго 107 особей было вскрыто. На осно-
вании наличия эмбрионов были подсчитаны гинопары и самцы. Потенциальная плодовитость
гинопар определена путем учета эмбрионов с пигментированными глазами.
В третий год исследований (2020) отмечена локализация яиц M. dirhodum при заселении са-
женцев R. canina только ремигрантами. Для этого в период массового лёта гинопар (18 сентября)
на саженцы шиповника (n = 6) были надеты садки с большим количеством ремигрантов, скопив-
шихся в результате миграции с пшеницы. Учет отложенных яиц проведен 28 октября в период
окончания листопада. Наблюдения за жизнеспособностью овипар, продолжавших жить на коре
побегов и опавших листьях, проводили до 1 декабря.
В последние два года исследований регистрировали сроки появления гинопар, самцов и яиц,
а также сроки листопада R. canina. Данные по температуре воздуха (World-Weather.ru, 2021),
использованные при последующем анализе результатов исследований, приведены в табл. 1.
Экспериментальные результаты были обработаны с помощью дисперсионного анализа
ANOVA в программе Statistica. Для статистической обработки данных был использован одно-
факторный дисперсионный анализ с применением теста Фишера. Значимость различий между
средними значениями определяли с помощью критерия Стьюдента (t-тест).
РЕЗУЛЬТАТЫ
На северо-западе европейской части России на посевах зерновых культур
M. dirhodum встречается ежегодно, однако в течение 5 лет в окр. С.-Петербурга обна-
ружить этот вид на шиповнике нам не удалось.
В 2018 г. были начаты исследования пригодности видов шиповника для развития
тлей. Для этого в течение лета поддерживали развитие 9 клонов M. dirhodum на пше-
нице сорта Ленинградская 6. К середине сентября среди потомков каждого из клонов
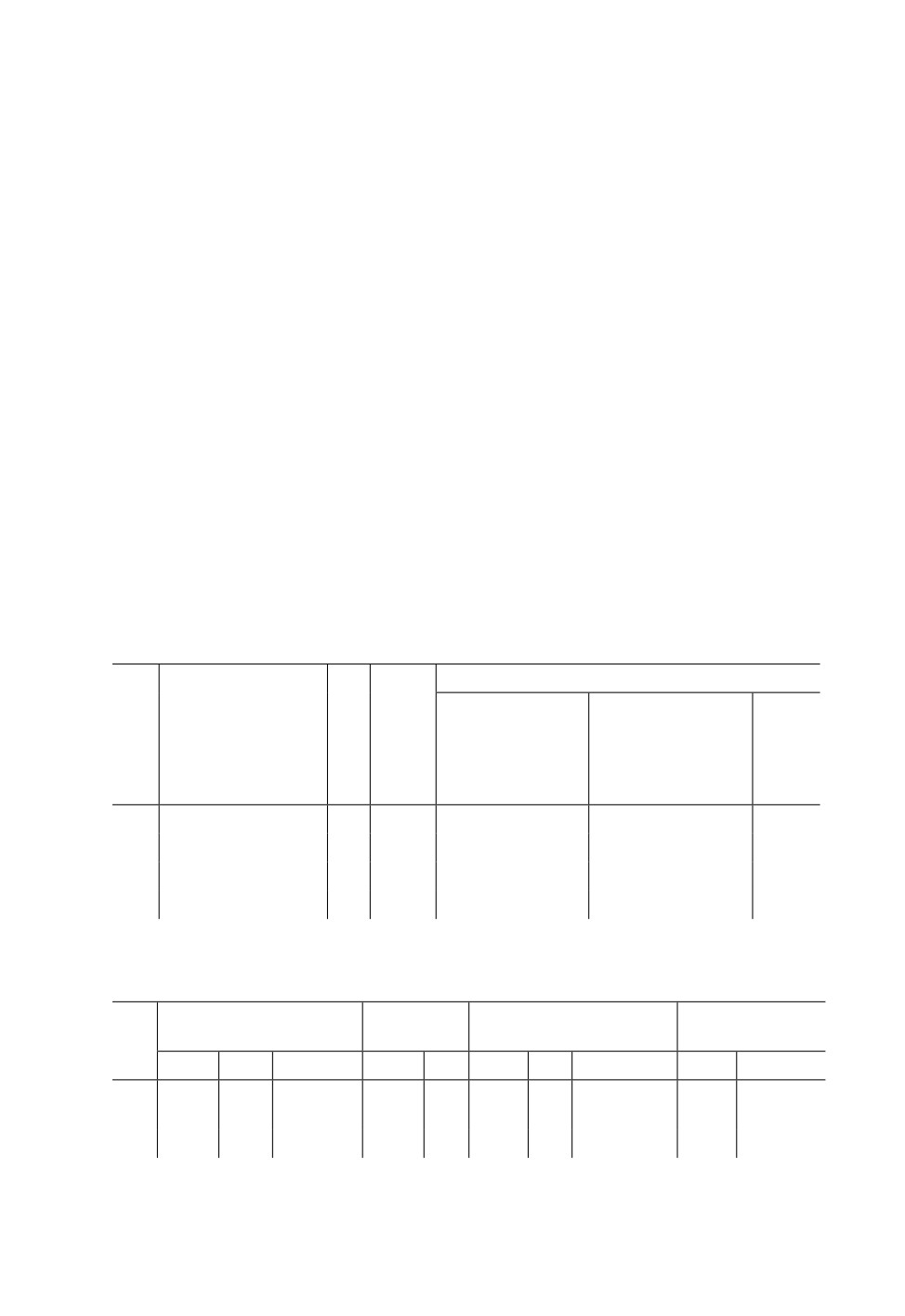
Таблица 1. Температура воздуха (°С) на территории С.-Петербурга (г. Пушкин) в период
проведения исследований осенних поколений Metopolophium dirhodum Walk. (World-
Weather.ru, 2021)
Среднесуточная
(декада)
Максимальная
Минимальная
Средняя
Год
Месяц
(дни)
(дни)
за месяц
1-я
2-я
3-я
2019
сентябрь
16.6
11.0
7.0
25 (2, 10)
0 (26)
11.5
октябрь
4.7
7.6
5.7
13 (25)
˗3; ˗4 (6, 30, 31)
6.0
ноябрь
0.5
5.7
˗1.3
10 (14)
˗5; ˗9 (23, 24)
1.5
декабрь
1.6
0.9
1.6
6 (6, 7)
˗6 (29)
1.5
2020
январь
1.7
3.1
˗0.1
8 (16)
˗4 (27, 28)
1.5
сентябрь
15.0
12.1
14.7
23 (3, 24)
7 (18, 20)
14.0
октябрь
13.1
7.8
7.4
18 (2, 3, 5)
0 (21)
9.4
ноябрь
5.9
2.9
1.8
11 (7)
˗3 (11)
3.5
711
на пшенице неожиданно были обнаружены не только гинопары и самцы, но и большое
количество овипар и яиц, которые были отложены на листья. Когда в сосуды с пше-
ницей поместили побеги шиповников R. rugosa и R. glauca, ремигранты и овипары
начали переходить на них со злака. Изменения их численности при перемещениях от-
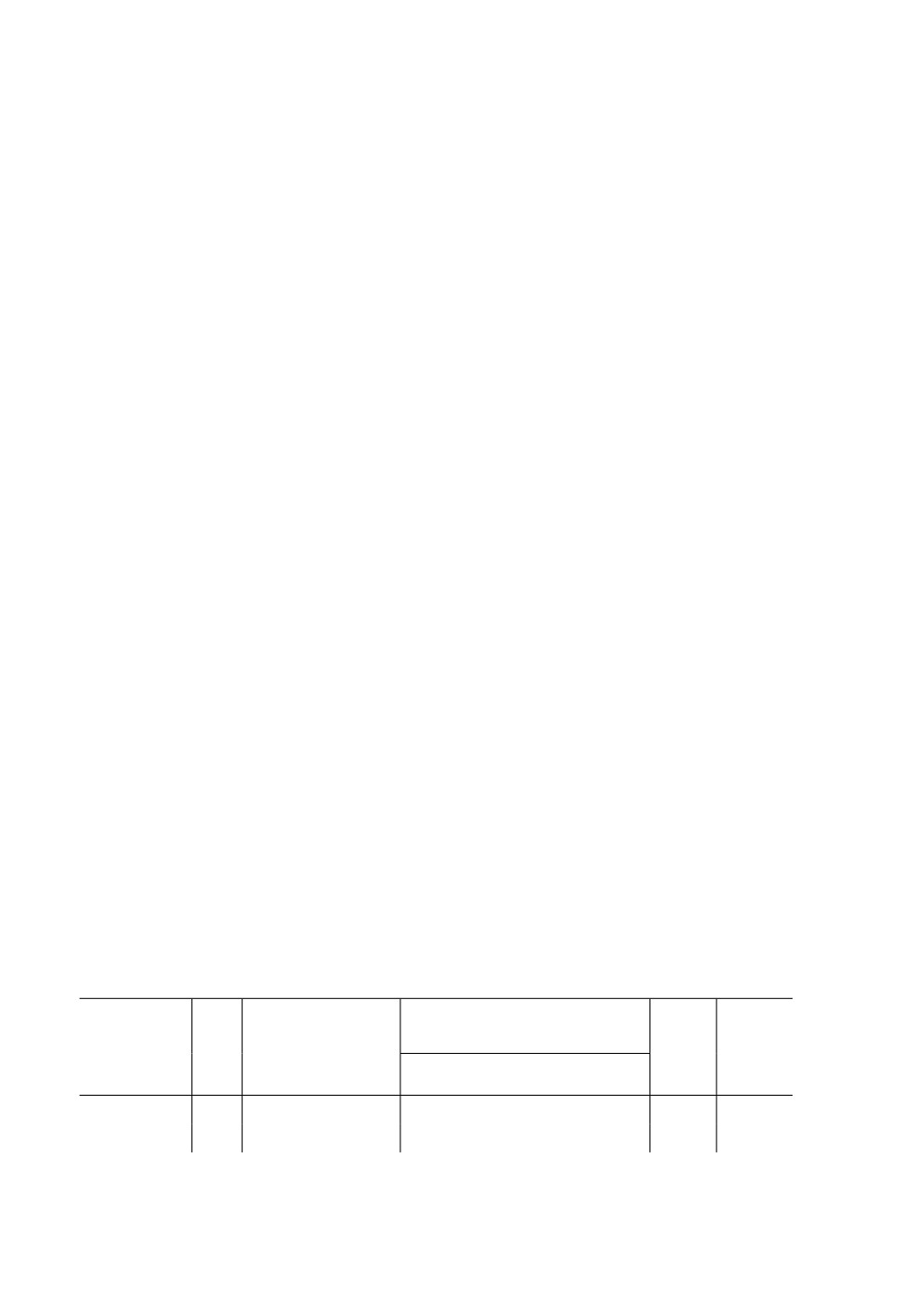
мечали в течение 12 дней до окончания откладки яиц (табл. 2, 3).
В процессе опыта ремигранты совершали полеты различной длительности, ча-
стично погибали; их количество на растениях не увеличивалось, но всегда было выше
на R. glauca, чем на R. rugosa (табл. 2). В период первого учета эти различия оказались
несущественными, поскольку в отличие от быстро расселившихся овипар многие ре-
мигранты не завершили облигатного полета и не осели. Ко второму учету избиратель-
ность ремигрантов при выборе растения была доказана (F = 3.80, p = 0.069). В резуль-
тате проведения двух первых учетов отмечено высоко достоверное влияние вида
шиповника на число переходящих на него с пшеницы овипар. Второй и третий учеты
показали также сильную зависимость числа отложенных овипарами яиц от вида ши-
повника (табл. 2). В обоих случаях - и для заселения, и при откладке яиц - тли предпо-
читали R. glauca. В конце опыта (17 октября) на пшенице оставалось 10 крылатых
особей (4.9 %), 17 имаго овипар (6.5 %) и 42 яйца (7.8 %). В верхней части садков
Таблица 2. Влияние вида шиповника на количество ремигрантов, овипар и яиц Metopolophium
dirhodum Walk. при миграции с пшеницы сорта Ленинградская 6
Число особей разных форм на побег
Дата
Вид шиповника
Ремигранты
Овипары
Яйца
X ± SE
X ± SE
X ± SE
5.X.2018
Rosa rugosa Thunb.
6.7 ± 1.26
4.0 ± 1.12
0.33 ± 0.23
R. glauca Pourr.
9.0 ± 2.54
14.4 ± 2.69
1.44 ± 0.78
10.X.2018
R. rugosa
3.22 ± 1.10
7.44 ± 1.86
2.78 ± 1.54
R. glauca
7.44 ± 1.86
21.0 ± 4.98
14.55 ± 2.98
17.X.2018
R. rugosa
6.0 ± 1.87
9.99 ± 2.39
20.89 ± 3.69
R. glauca
8.11 ± 2.74
17.22 ± 2.80
34.22 ± 4.58
Таблица 3. Результаты однофакторного дисперсионного анализа влияния вида шиповника на
количество ремигрантов, овипар и яиц Metopolophium dirhodum Walk. при миграции с пшеницы
сорта Ленинградская 6 (на основании данных табл. 2)
Ремигранты
Овипары
Яйца
Дата
n
F
p
F
p
F
p
5.X.2018
9
0.68
0.423
12.871
0.002
1.843
0.193
10.X.2018
9
3.80
0.069
7.41
0.015
12.303
0.0029
17.X.2018
9
0.404
0.534
3.965
0.064
5.1396
0.0376
П р и м е ч а н и е. n - число сосудов с пшеницей и побегами шиповника (см. раздел «Материал
и методика»), F - критерий Фишера, p - уровень значимости F.
712
скопилось 65 крылатых особей (32.1 %), которые не заселили ни одно из растений.
Распределение тлей и яиц на видах шиповника оказалось также различным (табл. 4).
Если на бесшипных побегах R. glauca все особи и яйца были найдены только на ли-
стьях и в период листопада опадали вместе с ними, то на побегах R. rugosa на листьях
яиц не было, а овипары откладывали яйца преимущественно на шипы или рядом
с ними.
В 2018 г. на пшеницу, находящуюся в сосудах рядом с шиповником, тлями было
отложено 42 яйца, в 2019 г. - 80 яиц, а в 2020 г. 76 яиц было обнаружено на всходах
злака Poa sp., случайно занесенного в сосуды с саженцами шиповника, где не было
пшеницы.
Весной 2019 г. на дикорастущей R. glauca яйца M. dirhodum зимовали в опаде под
кустами, но вышедших из яиц основательниц не обнаружено.
В 2019 г. был проведен модельный опыт (15 октября - 16 января), в ходе которого
исследовали поведение тлей в период миграции с пшеницы на молодые растения
R. canina, имевшие шипы лишь в нижней части стебля. К началу опыта уже наблюда-
лись массовый лёт гинопар и самцов, а также откладка яиц на пшенице (табл. 5).
В первые сутки переместившиеся с пшеницы на саженцы шиповника гинопары
(более 200), многие самцы и овипары различного возраста (более 300) скопились на
листьях, но уже на следующий день имаго овипар начали в массе перемещаться
Таблица 4. Число и доля (%) яиц Metopolophium dirhodum Walk., отложенных на разные
участки трехлетних саженцев и изолированных побегов разных видов шиповника
Доля яиц на разных участках
саженцы или
поросль саженцев
Число
верхние части
Год
Вид шиповника
n
или побеги (на
яиц
побегов (почки,
листья
нижней части у
ответвления,
шипов)
стебель)
2018
Rosa rugosa Thunb.
9*
188
96.3
3.7
0
R. glauca Pourr.
9*
308
0
0
100.0
2019
R. canina L.
3
845
96.4
3.6
0
2020
То же
6
4693
70.5
2.4
27.1
П р и м е ч а н и е. n - число саженцев или изолированных побегов (помечено звездочкой) шиповника.
Таблица 5. Фенология развития Metopolophium dirhodum Walk. в окрестностях С.-Петербурга
Листопад Rosa
Лёт гинопар
Лёт самцов
Откладка яиц
Год
canina
начало
пик
окончание
начало
пик
начало
пик
окончание
начало
окончание
2019
11.IX
20.X
25.X
30.IX
14.X
11.X
23.X
13-30.XI
14.X
11.XI
(16.I)*
2020
30.VIII
15.IX
7.X
7.IX
29.X
25.IX
15.X
26.XI (1.XII)
8.X
30.X
Примечание. *В скобках указаны даты гибели последних овипар.
713
кшипам в нижней части стеблей, где откладывали яйца и оставались до конца жизни.
Некоторые самцы следовали за овипарами. В период листопада (14 октября - 11 но-
ября; см. табл. 4) овипары продолжали питаться на опавших листьях до начала тре-
тьей декады ноября. К этому времени (23-24 ноября) начались заморозки до -5…-9 °C
(см. табл. 1) и листья полностью утратили пищевую ценность для тлей. К 24 ноября у
шипов саженцев размещались 52 овипары и 25 ремигрантов, 40 овипар оставались на
разных частях стебля, постепенно перемещаясь по нему вниз. В начале декабря реми-
грантов уже не было. Последние живые овипары (5 особей) были обнаружены 16 ян-
варя (спустя 53 дня после заморозков 24 ноября). Можно предполагать, что в конце
вегетации тли питались на коре молодых побегов шиповника. Ко времени последнего
учета на шипах и рядом с ними в нижней части стебля тлями было отложено 814 яиц,
на стеблях, где не было шипов - 28, у почек - 3. На пшенице оставалось 80 яиц, часть
из которых была отложена до начала опыта (см. табл. 4). На опавших листьях R. canina
яйца не были обнаружены. За период наблюдений температура не опускалась ниже
˗9 °C (см. табл. 1). Весной следующего, 2020 г. жизнеспособными оказались 80 % яиц.
Личинки отродились с 29 марта по 10 апреля и выживали исключительно на пер-
вичном хозяине, как это свойственно гетероцийным видам тлей (Мамонтова, 1973;
Williams, Dixon, 2007).
При среднесуточной температуре воздуха менее 10 °C (см. табл. 1) имагинальный
период гинопар, родившихся от бескрылых партеногенетических самок 17 сентября
(см. «Материал и методика»), в среднем продолжался более месяца (32.2 ± 1.56 дня) -
до третьей декады ноября, когда среднесуточные температуры стали уже отрицатель-
ными (см. табл. 1). При этом реализованная плодовитость одной гинопары в среднем
составляла 11.96 ± 1.31 овипары и оказалась несколько выше, чем потенциальная
(10.61 ± 0.58 эмбриона), но существенных различий между реализованной и потенци-
альной плодовитостями не выявлено согласно критерию Стьюдента (t = 1.03 < t 0.05 =
1.98). Среди 36 особей, родившихся 25 октября, не было обнаружено гинопар.
С приближением зимы погодные изменения, в том числе понижение среднесуточной
температуры в период между опытами 17 сентября - 15 октября (t = 6.8 °C) и 25 ок-
тября - 06 декабря (t =1.7 °C) на 5.1 °C, вызвали задержку преимагинального развития
самцов M. dirhodum в среднем на 4.4 дня (F = 20.949, p = 0.00002) (табл. 6).
В 2020 г. очень большое количество ремигрантов было помещено в сосуды с сажен-
цами R. canina до начала листопада и откладки яиц (см. табл. 4). Уже через сутки на
Таблица
6. Влияние среднесуточной температуры воздуха на продолжительность
преимагинального развития самцов Metopolophium dirhodum Walk. различных дат отрождения
(однофакторный дисперсионный анализ)
Среднесуточная
Продолжительность развития,
Дата
температура в
дни
n
F
p
отрождения
период развития
самцов
X ± SE
17.IX.2019
70
6.8 ℃
36.96 ± 0.405
20.949
0.00002
25.X.2019
14
1.7 ℃
41.36 ± 0.700
Примечание. Обозначения как в табл. 3.
714
листьях шиповника появилось множество личинок овипар, которые питались на ли-
стьях и по мере созревания мигрировали к шипам для откладки яиц. Осень в 2020 г.
была более теплой и благоприятной для развития овипар и откладки яиц, чем в 2019 г.
(см. табл. 1). Первые яйца у почек и в местах ответвлений боковых побегов обнару-
жены 15 октября, когда около шипов было уже множество яиц. К концу листопада
(28 октября) (см. табл. 4) около шипов наблюдалось гораздо больше яиц, чем в преды-
дущие годы. В отличие от 2019 г., около четверти яиц было отложено на листьях са-
женцев R. canina и опало вместе с ними, 113 яиц было отложено на стебле и у почек,
76 яиц оказалось в одном из сосудов на сорном злаке Poa sp., который было решено не
удалять (см. табл. 3). В 2020 г., хотя осень в целом была более теплой, полный ли-
стопад закончился раньше (30 октября) (см. табл. 1), чем в 2019 г., возможно, из-за
более холодной второй декады ноября. К 10 ноября на опавших листьях тлей уже не
было, овипары частично мигрировали на молодые стебли; 26 ноября на шиповнике
находились 10 живых овипар, которые погибли 1 декабря. Таким образом, как и в пре-
дыдущем году, овипары обнаружили способность более 20 дней оставаться живыми,
когда питание могло происходить только на коре молодых побегов шиповника.
ОБСУЖДЕНИЕ
Запас зимующих яиц гетероцийных видов тлей, в том числе M. dirhodum, их локали-
зация, количество и выживаемость во многом определяют развитие популяций вреди-
телей не только на первичных, но и на вторичных хозяевах. Не случайно прогнозы
численности вредящих видов тлей делают на основании учетов количества зимующих
яиц (Берим, 2014; Ежегодный справочник агронома, 2018). Считается, что M. dirhodum,
как и многие другие виды тлей, откладывает яйца преимущественно рядом с почками,
иногда между шипами, на шипах или коре побегов (Lubiarz, Cichocka, 2014), однако
чаще указываются лишь места питания основательниц (верхушки побегов шиповника,
обратная сторона листьев) (Kmieć, 2006; Сауткин, Буга, 2007; Krzyźanowski, 2017),
а процесс откладки яиц не рассматривается (Сауткин, Буга, 2011).
Массовое развитие овипар и откладку яиц на вторичном хозяине у M. dirhodum мы
наблюдали впервые, хотя и у других видов злаковых тлей, в частности у Rhopalosiphum
padi, отдельные овипары иногда могут достичь репродуктивного возраста и даже от-
ложить единичные яйца на пшенице (Верещагина, Гандрабур, 2016; Peng et al., 2017).
Такие различия между видами можно рассматривать в зависимости от мотивирован-
ности ремигрантов на смену хозяина и полет. В случае отсутствия побегов первичного
хозяина ремигранты Rh. padi, за редким исключением, скапливались на садке, покидая
пшеницу без воспроизводства (мотивация на смену хозяина и полет); ремигранты
M. dirhodum, наоборот, часто оставались на пшенице и завершали жизненный цикл
откладкой яиц. При наличии близко расположенных побегов первичного хозяина по-
ведение ремигрантов обоих видов тлей оказалось сходным. Часть из них быстро пере-
мещалась на побеги первичного хозяина, другая часть, мотивированная на миграцию
на дальние расстояния (облигатные мигранты), скапливалась по мере окрыления
особей в верхней части садка. Количество ремигрантов, мотивированных на дальний
полет, у Rh. padi в среднем по 9 клонам составило 43.3 %, остальные сразу перелетали
на побеги черемухи (Верещагина, Гандрабур, 2016). У M. dirhodum к концу опыта
(2018 г.) 32.1 % от общего количества ремигрантов еще не заселили растения, воз-
можно, поэтому различия в выборе R. rugosa или R. glauca в нашем опыте не сразу
715
были очевидными. Сокращение продолжительности лёта при ремиграции в случае
близкого расположения первичного хозяина встречается и у других видов тлей, на-
пример, у Phorodon humuli (Schrank) (Hardie, Campbell, 1998). Овипары M. dirhodum
явно предпочитали листья R. glauca, где и откладывали яйца. На R. rugosa имаго
овипар оказалось меньше, чем на R. glauca, и они располагались только на стебле, а на
листьях присутствовало лишь небольшое количество овипар в личиночной стадии.
Этим определялось и достоверно большее число яиц на R. glauca. Возможно, причины
различий в степени заселения тлями были такие особенности листьев R. rugosa, как
глубокая морщинистость, довольно густая опушенность и обилие железок на адакси-
альной стороне (Плантариум…, 2007-2021), что затрудняет тлям питание на этих
листьях. Независимо от того, были ли овипары отрождены гинопарами непосред-
ственно на шиповнике или перешли на него с пшеницы, а также в какой форме (изоли-
рованные побеги или саженцы) этот шиповник был предложен им в садках, овипары
для откладки яиц, как правило, мигрировали к шипам на R. rugosa и R. canina либо
только на листья у бесшипного R. glauca. Такие различия в поведении овипар отме-
чены впервые. Лишь при массовом размножении тлей яйца были найдены на листьях
или у почек R. canina и даже на злаке, ранее не заселенном летними морфами. Эти
особенности овипар позволяют выявить и контролировать места резервации зимую-
щего поколения M. dirhodum на шипах в нижней части молодых побегов шиповника.
Способность M. dirhodum откладывать яйца на листья шиповника, а также на вто-
ричного хозяина снижает численность популяции тлей весной: на листьях R. glauca
яйца, вероятно, не выжили, так как основательниц весной не было обнаружено, а на
травах основательницы, даже если и появятся весной, погибнут из-за их узкой пи-
щевой специализации на первичном хозяине. Таким образом, бесшипные формы ши-
повника могут быть перспективными для селекции устойчивых к тлям форм
шиповника, так как отложенные на листья яйца в течение зимы погибают, как это про-
изошло на R. glauca. Способность гетероцийных видов тлей завершать жизненный
цикл на однолетних растениях служит примером трофической приемлемости для тлей
очень отдаленно родственных зимних и летних хозяев, позволяющих осенним поколе-
ниям по крайней мере некоторых видов (M. dirhodum) успешно питаться на тех и
других.
Опубликованы сведения о положительной корреляции длительности периода осен-
него размножения M. dirhodum на озимой пшенице с численностью тли весной. Ука-
зывается, что пороговая температура для развития тлей определяется пороговой тем-
пературой для развития озимой пшеницы и составляет 0-5 °C (Brabec et al., 2014).
Некоторые авторы отмечали продолжительность осенней ремиграции и репродукции
овипар M. dirhodum после окончания листопада до наступления устойчивых замо-
розков (Сауткин, Буга, 2007, 2011; Lubiarz, Cichocka, 2014), однако пороговые темпе-
ратуры для данного вида тли и продолжительность ее жизни после листопада не ука-
заны. В целом для шести видов тлей, питающихся на шиповнике в Польше, в качестве
пороговой температуры для откладки яиц указывается ˗5 °C (Lubiarz, Cichocka, 2014).
Нами было показано длительное выживание (более 20 дней) овипар на молодых по-
бегах шиповника и на опавших листьях с последующей миграцией к шипам в нижние
части стеблей, что значительно удлиняет период откладки яиц после листопада. Даже
при отрицательных температурах воздуха ˗3…˗9 °C в 2019 г. (22-27 ноября) некоторые
овипары оставались живыми, если в последующие дни температура была положи-
тельной и не опускалась ниже ˗5 °C ночью (см. табл. 1), поэтому удаление опавших
716
листьев с овипарами будет способствовать снижению численности яиц на шиповнике.
С уменьшением среднедекадных температур ниже +10 °C продолжительность имаги-
нального периода гинопар составляла более месяца. Однако дополнительное созре-
вание эмбрионов в этот период не было статистически доказано, поскольку суще-
ственные различия между потенциальной и реализованной плодовитостью гинопар не
выявлены. Осеннее понижение температур сопровождалось значительным замедле-
нием преимагинального развития самцов, и способность спариваться некоторые из
них сохраняли до декабря, как было отмечено в 2019 г. Размножение тлей поздней
осенью происходит практически при отсутствии энтомофагов и способствует повы-
шению численности весенних поколений.
ЗАКЛЮЧЕНИЕ
В результате исследований было установлено, что M. dirhodum в окрестностях
С.-Петербурга может осенью успешно развиваться на Rosa rugosa, R. canina и
R. glauca. Впервые была показана способность особей осенних поколений этого вида
тлей в массе развиваться и откладывать яйца на вторичном хозяине (пшенице), что
свидетельствует о широте пищевых адаптаций данного вида. Однако при близком раз-
мещении в садках первичного и вторичного хозяев ремигранты и овипары большей
частью переселялись на первичного хозяина. Распределение тлей и яиц на растении
зависело от морфологических характеристик вида шиповника - на R. rugosa и
R. canina овипары преимущественно перемещались к шипам и там откладывали яйца
(96.3 % и более 70.5 % соответственно). Впервые отмечено, что на бесшипной форме
шиповника (R. glauca) овипары и яйца располагались исключительно на листьях
и оставались в опаде под растениями. На опушенных листьях R. rugosa яиц тлей
не найдено. Установлено также, что овипары предпочитали заселять и откладывать
яйца на R. glauca, а не на R. rugosa (F = 12.9, p = 0.002 и F = 12.3, p = 0.003 со-
ответственно). На листьях R. canina яйца тлей появлялись лишь после массового ско-
пления их на шипах. Культивирование бесшипных сортов шиповника, гибель яиц
M. dirhodum на опавших листьях и травах снижают численность весенней популяции
тлей.
Отмечены некоторые параметры развития осенних морф M. dirhodum. Продолжи-
тельность жизни имаго гинопар при среднедекадных температурах ниже +10 °C в
среднем составляла 32.2 ± 1.56 дня, при этом реализованная плодовитость суще-
ственно не превышала потенциальную. Понижение температуры воздуха поздней
осенью существенно тормозило преимагинальное развитие самцов (F
=
20.9,
p = 0.00002), но жизнеспособность наряду с гинопарами они сохраняли даже после
окончания листопада до 20 дней благодаря возможности питания на опавших листьях.
Более того, выживание некоторых овипар до наступления устойчивых отрицательных
температур (-5…-9 °C) предполагает их питание у шипов молодых побегов и после
миграции с опавших листьев.
Выявленные новые особенности развития и откладки яиц M. dirhodum на первичном
хозяине позволяют объяснить пути формирования численности этой тли на предста-
вителях рода Rosa и могут быть использованы для совершенствования контроля дан-
ного вредителя.
717
СПИСОК ЛИТЕРАТУРЫ
Берим М. Н. Metopolophium dirhodum Walk. - Розанно-злаковая тля. В: Афонин А. Н.; Грин С. Л.; Дзюбенко
Н. И.; Фролов А. Н. (ред.). Агроэкологический атлас России и сопредельных стран: экономически
значимые растения, их вредители, болезни и сорные растения. [Интернет-версия 2.0]. 2008. [http://
Берим М. Н. 2014.Тли на зерновых культурах. Сельскохозяйственные вести 1: 16-18.
Бокина И. Г. 2009. Злаковые тли и их энтомофаги в лесостепи Западной Сибири. Новосибирск: Сибирский
НИИ земледелия и химизации сельского хозяйства, 182 с.
Верещагина А. Б., Гандрабур Е. С. 2016. Изменчивость параметров развития клонов черемухово-злаковой
тли Rhopalosiphum padi (L.) (Homoptera, Aphididae) в течение жизненного цикла как генотипическая
адаптация данного вида. Энтомологическое обозрение 95 (4): 729-747.
Евтухова М. В. 2016. Изучение эколого-биологической изменчивости представителей р. Rosa L. для селек-
ции в условиях юго-запада ЦЧЗ. Автореферат диссертации на соискание ученой степени кандидата
биологических наук. Рамонь: Белгородский государственный национальный исследовательский уни-
верситет, 25 с.
Ежегодный справочник агронома. 2018. СПб.: Россельхозцентр, филиал по Ленинградской области, 168 с.
Ильин В. С. 2017. Шиповник, клюква и другие редкие культуры сада. Челябинск: Южно-Уральский НИИ
садоводства и картофелеводства, 318 с.
Мамонтова В. А. 1973. Подотряд тли - Aphididae. В кн.: В. П. Васильев (ред.). Вредители сельскохозяйствен-
ных культур и лесных насаждений. Т. 1. Киев: Урожай, с. 214-305.
Плантариум: открытый онлайн атлас-определитель растений и лишайников России и сопредельных стран.
Сауткин Ф. В., Буга С. В. 2007. Дендрофильные и дендрогербофильные тли - вредители цветочно-декора-
тивных растений, интродуцированных в Беларусь. В кн.: Теоретические и прикладные аспекты
интродукции растений как перспективного направления развития науки и народного хозяйства: Мате-
риалы международной научной конференции, посвященной 75-летию со дня образования Централь-
ного ботанического сада НАН Беларуси. Т. 2. Минск: Эдит ВВ, с. 230-232.
Сауткин Ф. В., Буга С. В. 2011. Особенности биологии и экологии розанно-злаковой тли (Metopolophium
dirhodum (Walk.)) на розах (Rosa spp.) в условиях г. Минска. В кн.: Структура биологического разно-
образия центрального региона Белорусской гряды (на примере модельных групп беспозвоночных и
позвоночных животных. Минск: Белорусский государственный университет, с. 69-73. Депонирована
в Белорусском институте системного анализа. 09.09.2011. №Д201140
Alford D. V. 2012. Pests of Ornamental Trees, Shrubs and Flowers: A Colour Handbook, Second Edition. CRC
Press, 480 p.
Brabec M., Honěk A., Pekár S., Martinkova Z. 2014. Population dynamics of aphids on cereals: digging in the time-
series data to reveal population regulation caused by temperature. PLoS ONE 9: e106228. https://doi:
10.1371/journal.pone.0162139
Emden H. F. van, Harrington R. 2007. Aphids as Crop Pests. Wallingford, United Kingdom: CABI, 717 p. http://
dx.doi.org/10.1079/9780851998190.0000
щения 25.12.2020
Hardie J., Campbell C. A. M. 1998. The flight behaviour of spring and autumn forms of the damson-hop aphid,
Phorodon humuli, in the laboratory. In: J. M. Nieto Nafría, A. F. G. Dixon (eds). Aphids in Natural and
Managed Ecosystems. León, Spain: Universidad de León, p. 205-212.
Honěk A., Martinkova Z. 2004. Host plant age and population development of a cereal aphid, Metopolophium
dirhodum (Hemiptera: Aphididae). Bulletin of Entomological Research 94 (1): 19-26. doi: 10.1079/
BER2003280
Honěk A., Martinkova Z., Saska P., Dixon A. F. G. 2018. Aphids (Homoptera: Aphididae) on winter wheat: predicting
maximum abundance of Metopolophium dirhodum. Journal of Economic Entomology 20 (10): 1-9.
Howard М. Т., Dixon A. F. G. 1992. The effect of plant phenology on the induction of alatae and the development
of populations of Metopolophium dirhodum (Walker), the rose-grain aphid, on winter wheat. Annals of
Applied Biology 120 (2): 203-213. doi: 10.1111/j.1744-7348.1992.tb03418.x
Kmieć K. 2006. Dynamics of number of Metopolophium dirhodum (Walk.) (Hemiptera, Aphididae) occurring on
shrubs of roses in Lublin. Acta Scientiarum Polonorum, Hortorum Cultus 5 (2): 11-21.
Krzyźanowski R. 2017. Dynamics of serious pest of Metopolophium dirhodum (Walk.) (Hemiptera: Aphididae) on
718
Lubiarz M., Cichocka E. 2014. The process of aphid egg-laying and the little known role of the Coccinellidae in
aphid egg destruction in Poland - preliminary results. Journal of Plant Protection Research 54 (3): 242-249.
Peng X., Qiao X., Chen M. 2017. Responses of holocyclic and anholocyclic Rhopalosiphum padi populations to
low-temperature and short-photoperiod induction. Ecology and Evolution 7 (4): 1030-1042. https://
doi:10.1002/ece3.2720
Williams I. S., Dixon A. F. G. 2007. Chapter 3. Life cycles and polymorphism. In: H. F. van Emden, R. Harrington
(eds). Aphids as Crop Pests. Wallingford, United Kingdom: CABI, p.
org/10.1079/9780851998190.0000
CHARACTERISTICS OF THE DEVELOPMENT OF AUTUMN GENERATIONS
AND OVIPOSITION IN METOPOLOPHIUM DIRHODUM WALK.
(HEMIPTERA, STERNORRHYNCHA: APHIDIDAE)
A. B. Vereschagina, E. S. Gandrabur
Key words: aphids, remigrants, oviparae, eggs, dog-rose, spines, leaves, distribution,
secondary host plant, feeding.
SUMMARY
Modelling experiments to investigate autumn generations of the Metopolophium dirhodum Walk.
population in St. Petersburg vicinity have been carried out during three years. Successful completing
development of the aphid on Rosa rugosa Thunb., R. canina L., and R. glauca Pourr. was observed.
It was shown for the first time, that the oviparae can proceed the development and reproduction on
the dog-roses and secondary host plants such as wheat. The location of the eggs on the dog-roses
depends on the morphological features of the plant species: mainly close to the spines or on the latter
on R. rugosa and R. canina (96.3% and not less than 70.5%, respectively), and only on the leaves of the
spineless R. glauca. The adult oviparae did not colonize the pubescent leaves of R. rugosa. Oviparae
remigrating from wheat preferred for oviposition R. glauca over R. rugosa (F = 12.9, p = 0.002 and
F = 12.3, p = 0.003, respectively), but the foundatrices did not hatch from the eggs laid on R. glauca
leaves, whereas about 80% of the eggs on the spines of R. canina had overwintered. The ability of the
aphids’ feeding on the fallen leaves and on young stems of dog-roses until constant frosts (-5…-9 ºС)
beginning was found. A decrease in the average daily air temperature by 5.1º caused a slow-down of the
preimaginal development of the males (F = 20.9, p = 0.00002) and the extension of their flight period.
Some parameters of the gynoparae development have been established. The features of the autumn
M. dirhodum generations may be useful for planning control of the pest population densities and in the
dog-roses selection.
719