ПАРАЗИТОЛОГИЯ, 2020, том 54, № 6, с. 470-490.
УДК 576.895
СТРОЕНИЕ ОРГАНА ГАЛЛЕРА И СИСТЕМАТИКА
ИКСОДОВЫХ КЛЕЩЕЙ ПОДСЕМЕЙСТВА IXODINAE
(СЕМЕЙСТВО IXODIDAE)
© 2020 г. С. А. Леонович⁕
Зоологический институт РАН,
Университетская наб. 1, Санкт-Петербург, 199034 Россия
⁕ e-mail: leonssa@mail.ru
Поступила в редакцию.10.09.2020 г.
После доработки 20.09.2020 г.
Принята к публикации 25.09.2020 г.
На основании собственных и литературных данных проведен анализ строения органа Галлера,
главного рецепторного органа иксодовых клещей, изученного методами растровой электронной
микроскопии, у 43 видов клещей рода Ixodes Latreille, 1795, единственного рода в составе под-
семейства Ixodinae. Описано строение органа Галлера у представителей 13 подродов: Ixodes
s. str. (14 видов), Pholeoixodes Schulze, 1942 (3 вида), Ixodiopsis Filippova, 1957 (4 вида),
Ceratixodes Neumann, 1902 (1 вид), Scaphixodes Schulze 1941 (2 вида), Trichotoixodes Reznik,
1961 (2 вида), Partipalpiger Hoogstraal, Clifford, Saito and Keirans 1973 (1 вид), Pomerantzevella
Feider, 1965 (1 вид), Eschatocephalus Frauenfeld, 1851 (2 вида), Afrixodes Morel, 1966 (1 вид),
Sternalixodes Schulze, 1938 (8 видов), Multidentatus Clifford, Sonenshine, Keirans and Kohls, 1973
(3 вида), Endopalpiger Schulze (1935) (1 вид). Выделены таксономические признаки (число и
топография сенсилл в разных отделах органа, наличие и форма капсулярного отверстия, форма
углубления сенсилл передней группы и др.). Показано, что строение органа Галлера специфич-
но для отдельных подродов, в то время как видовые различия (в пределах каждого подрода)
практически не выражены. Проведено сравнение полученных данных с существующими пред-
ставлениями о таксономии рода Ixodes.
Ключевые слова: орган Галлера, растровая электронная микроскопия, Ixodinae, Ixodes,
систематика
DOI: 10.31857/S1234567806060024
Орган Галлера, впервые описанный естествоиспытателем Галлером в 1881 г. (Haller,
1881), - основной чувствительный орган иксодовых клещей, переносчиков опасных
470
трансмиссивных заболеваний. У клещей, как известно, нет антенн - придатков голов-
ной капсулы, несущих основную массу сенсорных образований, как это наблюдается
у насекомых, поэтому роль антенн насекомых у клещей выполняют передние ноги.
Клеши перемещаются на трех парах ног, а передняя пара ног служит им своеобразны-
ми аналогами антенн насекомых. Именно поэтому на передних ногах, а точнее, на их
дистальных сегментах, лапках, располагаются основные органы чувств, позволяющие
обнаруживать источник пищи или полового партнера. У гамазовых клещей - это тар-
зальный рецепторный орган (Leonovich, Stanyukovich, 2011; Leonovich, Dimov, 2012),
а у иксодовых клещей - это орган Галлера. Кроме того, у большинства клещей от-
сутствуют глаза (Леонович, 2005)
Орган Галлера - чрезвычайно сложно устроенный орган чувств, в составе которого
обнаружены сенсиллы (элементарные чувствительные образования членистоногих),
реагирующие на запах, градиенты температуры, влажности и на некоторые другие
стимулы (Леонович, 2005).
В строении органа Галлера есть черты, определяющие его функциональные осо-
бенности. И одновременно есть морфологические признаки, никак не связанные
с функциональными особенностями. Примеры - пористый волосок передней груп-
пы сенсилл (обонятельный рецептор) может располагаться по-разному среди осталь-
ных сенсилл передней группы, но при этом он остается сенсиллой, расположенной
в переднем углублении, и, вместе с таким же волоском (сенсиллой), расположенным
на симметричной передней лапке, способен определять направление источника обоня-
тельного стимула. Учитывая мельчайшие размеры органа Галлера, расположение этой
сенсиллы относительно остальных сенсилл (а оно может значительно различаться ) на
физиологическую и особенно поведенческую роль такой сенсиллы никак не влияет.
Далее, основное функциональное значение отверстия капсулы - обеспечить доступ
обонятельных (летучих) молекул к обонятельным сенсиллам капсулы. При этом форма
отверстия, т.е. вытянуто оно вдоль или поперек продольной оси членика, округлое оно
или овальное, или изрезанное - никакого функционального значения не имеет. То есть,
подобные морфологические признаки, на которые эволюционный пресс не оказывает
давления, могут быть использованы в качестве таксономических признаков (см. ниже).
Следует отметить, что ни в одной из таксономических работ данные по строению
органа Галлера никак не использовались (и не используются до сих пор). Вместе
с тем игнорировать данные по такому важному органу вряд ли правомочно.
Анализ современных работ, которые посвящены таксономии иксодовых клещей
и в которых в том числе использованы молекулярные данные, показывает, что на
основании этих данных делаются весьма скоропалительные выводы, никак не соот-
ветствующие морфологическому анализу. В связи с этим автору представляется важ-
ным рассмотреть строение органа Галлера, основного сенсорного органа иксодовых
471
клещей, не в связи с его функциональными особенностями, а в связи с таксономией
этой группы.
Данное первое сообщение посвящено рассмотрению особенностей строения органа
Галлера у клещей подсемейства Ixodinae, представленного в мировой фауне един-
ственным родом Ixodes.
МАТЕРИАЛ И МЕТОДИКА
Всего в данной работе было проанализировано строение органа Галлера у следующих видов
иксодовых клещей рода Ixodes: (подродовая принадлежность вида приведена по: Филиппова
(1977) и Clifford et al. (1973).
Подрод Ixodes s. str.
I. persulcatus Schulze, 1930
I. redicorzevi Olenev, 1927
I. ricinus (Linnaeus, 1758)
I. pavlovskyi Pomerantzev, 1946
Подрод Pholeoixodes Schulze, 1942
I. arboricola Schulze & Schlottke, 1930
I. lividus Koch, 1844
Подрод Ixodiopsis Filippova, 1957
I. angustis Neumann, 1899
I. granulatus Supino, 1827
Подрод Ceratixodes Neumann, 1902
I. uriae White, 1852
Подрод Scaphixodes Schulze, 1941
I. caledonicus Nuttall, 1910
I. signatus Birula, 1895
Подрод Trichotoixodes Reznik, 1961
I. frontalis Panzer, 1795
Подрод Partipalpiger Hoogstraal, Clifford, Saito & Keirans 1973
I. ovatus Neumann, 1899
Подрод Pomerantzevella Feider, 1965
I. simplex Neumann, 1906
Подрод Eschatocephalus Frauenfeld, 1851
I. vespertilionis Koch, 1844
Подрод Afrixodes Morel, 1966
I. rubicundus Neumann, 1904
Кроме того, были использованы данные, приведенные в работах других авторов, как по-
священных изучению органа Галлера непосредственно, так и в работах, не посвященных не-
посредственно органу Галлера, но содержащих электронные фотографии органа, полученные
при помощи растрового электронного микроскопа.
472
Подрод Ixodes s. str.
I. acutitarsus Karsch, 1880 (Ronghang, Roy, 2014)
I. asanumai (Hayashi, Hasegawa, 1983)
I. boliviensis Neumann, 1904 (Homsher, Sonenshine, 1975)
I. dentatus Marx, 1899 (Homsher, Sonenshine, 1975)
I. granulatus Supino, 1897 (Homsher, Sonenshine, 1975)
I. laguri Olenev, 1929 (Honzаkovа et al., 1975)
I. loricatus Neumann 1899 (Homsher, Sonenshine, 1975)
I. luciae Senevet, 1940 (Homsher, Sonenshine, 1975)
I. muris Bishopp et Smith, 1937 (Homsher, Sonenshine, 1975)
I. scapularis Say, 1821 (Homsher, Sonenshine, 1975)
Подрод Pholeoixodes Schulze, 1942
I. texanus Banks, 1909 (Homsher, Sonenshine, 1975)
Подрод Ixodiopsis Filippova, 1957
I. cookie Packard, 1869 (Homsher, Sonenshine, 1975)
I. marxi Banks, 1908 (Homsher, Sonenshine, 1975)
Подрод Trichotoixodes Reznik, 1961
I. brunneus Koch, 1844 (Homsher, Sonenshine, 1977)
Подрод Sternalixodes Schulze, 1938
I. anatis Chilton, 1904 (Homsher et al., 1988)
I. confusus Roberts, 1960 (Homsher et al., 1988)
I. cordifer Neumann, 1908 (Homsher et al., 1988)
I. cornuatus Roberts, 1960 (Homsher et al., 1988)
I. dendrolagi Wilson, 1967 (Homsher et al., 1988)
I. hirsti Hassall, 1931 (Homsher et al., 1988)
I. holocyclus Neumann, 1899 (Homsher et al., 1988)
I. trichosuri Roberts, 1960 (Sternalixodes) (Homsher et al., 1988)
Подрод Multidentatus Clifford, Sonenshine, Keirans & Kohls, 1973
I. paranaensis Barros-Battesti, Arzua, Pichorim & Keirans, 2003 (Barros-Battesti et al., 2003)
I. percavatus Neumann, 1906 (Homsher, Sonenshine, 1979; Homsher et al., 1988)
I. auritulus Neumann, 1904 (Barros-Battesti et al., 2003)
Подрод Endopalpiger Schulze (1935)
I. woyliei Ash et al., 2017 (Ash et al., 2017)
Подрод Eschatocephalus Frauenfeld, 1851
I. ariadnae Hornok et al., 2014 (Hornok et al., 2015)
Часть исследованных видов, непосредственно собранных автором, была изучена в растровом
электронном микроскопе (РЭМ) по десяти и более экземпляров каждого пола и фазы развития
(I. ricinus, I. persulcatus, I. redicorzevi, I. rubicundus, I. lividus). Сборы проводили в разных районах
России и других стран. Остальные изученные автором виды были взяты из коллекции иксодовых
клещей ЗИН РАН и изучены в РЭМ по двум-трем экземплярам каждой фазы развития и пола.
473
Для исследования в растровом электронном микроскопе клещи были зафиксированы в 70 %
растворе этилового спирта, очищены в ультразвуковой ванне, а затем высушены методом критиче-
ской точки в установке Hitachi Critical Point Dryer (HCP-2) c использованием жидкой углекислоты
в качестве рабочего агента. Высушенные препараты клещей наклеивали на столики-подложки
при помощи двусторонне-липкой ленты, напыляли платиной в установку EIKO-5 (Япония), и
исследовали в растровом электронном микроскопе Hitachi S570.
Результаты и обсуждение
У всех исследованных видов клещей орган Галлера состоит из нескольких мор-
фологически обособленных частей: капсулы, передней группы сенсилл, сенсилл
дистального бугорка и посткапсулярных сенсилл (рис. 1). Капсула - отдел орга-
на Галлера, в состав которого входят от 5 до 7 пористых волосков обонятельных
сенсилл. Сенсилла - элементарный орган чувств членистоногих, включающий
в себя кутикулярный отдел, представленный волоском (в таксономической литера-
туре называемый также сетой, щетинкой, хетой и т. п.), иннервируемым одним или
несколькими дендритами биполярных сенсорных нейронов и определяющим модаль-
ность (функцию) сенсиллы. Так, в обонятельных сенсиллах полый тонкостенный во-
лосок (сета, хета) пронизан многочисленными мелкими порами, а в полости волоска
обнаруживаются многочисленные ветвления рецепторных ресничек (Леонович, 1985;
2005). Контактные хеморецепторные сенсиллы (органы вкуса) характеризуются полым
волоском, в полости которого проходят неветвящиеся дендриты, а на вершине волоска
находятся одна крупная пора или несколько более мелких пор (однако в сравнении
с порами обонятельных сенсилл они значительно более крупные. Более подробно
о связи ультраструктуры сенсилл клещей с их функцией можно узнать из моногра-
фии Леоновича (2005). Капсула представляет собой углубление, часто погруженное
под покровы, и сообщается со средой посредством капсулярного отверстия. У кле-
щей подсемейства Ixodinae капсула может быть открытой, у представителей второго
подсемейства (Amblyomminae или Metastriata) она всегда закрытая. Обонятельные
сенсиллы капсулы реагируют на летучие запахи хозяина, в частности на углекислый
газ и аммиак (Леонович, 2005).
Перед капсулой у всех иксодовых клещей расположен набор из 4 типов сенсилл
(пористый, тонкий, конический и двуслойный волоски сенсилл соответствующих ти-
пов) (рис. 1). Пористый волосок принадлежит обонятельной сенсилле, бороздчатые
волоски содержат хемо-терморецепторы, тонкий волосок - гигрорецептор, функция
конического волоска до конца не выяснена (рис. 1А). Количество сенсилл разных
типов может варьировать, но их набор (из 4 типов) остается неизменным (Леонович,
2005). У клещей подсемейства Ixodinae эти сенсиллы располагаются на дне обшир-
ного углубления дистальнее капсулы - плоского формирования, слегка погруженного
474
под поверхность кутикулы тарзального членика и часто отграниченного кутикуляр-
ным валиком (рис. 1). У клещей подсемейства Amblyomminae (Prostriata) обширное
углубление, в котором располагаются сенсиллы передней группы, отсутствует и все
сенсиллы передней группы сконцентрированы на небольшом участке, отграниченном
общим кутикулярным валиком (см. вторую часть настоящего сообщения).
Для каждого вида клещей характер взаимного расположения сенсилл передней
группы остается строго постоянным. В специальных исследованиях, посвященных
регенерации органа Галлера у клеща Ixodes ricinus, путем суперимпозиции (последо-
вательного наложения изображений, полученных в одном ракурсе и при одинаковом
увеличении растрового электронного микроскопа), было показано, что расположение
сенсилл передней группы варьирует в определенных пределах, но эти пределы никак
не меняют характер относительного взаиморасположения сенсилл (рис. 2).
Те же результаты были получены на африканском виде I. rubicundus, причем вы-
яснилось, что видоспецифичное взаиморасположение сенсилл передней группы ха-
рактерно для всех фаз развития (личинок, нимф и имаго) (рис. 3)
Отметим, что у личинок характер расположения сенсилл остается типичным для
вида, при этом в передней группе отсутствует один из тонких волосков, а число кап-
сулярных сенсилл также уменьшено. Дистальнее переднего углубления находится
дистальный бугорок [distal knoll, по терминологии Леоновича (Leonovich, 2004)], вы-
раженный только у представителей подсемейства Ixodinae (рис. 1). На этом бугорке
располагаются одна или две (у большинства Ixodinae) пар сенсилл. Одна из сенсилл
пары всегда представлена пористым обонятельным волоском, а вторая - редуциро-
ванной вкусовой сенсиллой (рис. 1). Пористый волосок дистального бугорка - один
из важнейших рецепторов органа Галлера у клещей подсемейства Ixodinae. У клещей
рода Ixodes это самая большая сенсилла в составе органа Галлера (если средний размер
сенсилл передней группы составляет примерно 10-20 мкм, а размер сенсилл капсулы
колеблется в пределах 20-50 мкм (у разных видов), то размер дистальной пористой
обонятельной сенсиллы переднего бугорка составляет 150-200 мкм).
У клеща Ixodes ricinus эта сенсилла содержит фенольный и лактоновый рецепторы,
позволяющие определять приближение потенциального хозяина с больших расстояний
(Leonovich, 2004).
Проксимальнее капсулы располагается группа посткапсулярных волосков (сенсилл)
(рис. 1). Латеральные сенсиллы представлены вкусовыми рецепторами (так же как и
латеральные волоски по бокам сенсилл дистального бугорка), центральные сенсиллы
этой группы относятся к двухслойным сенсиллам (комбинированным хемо-, термо-,
гигрорецепторам). Функция этих сенсилл остается неясной.
Как будет показано в дальнейшем, строение углубления для сенсилл передней груп-
пы, форма капсулярного отверстия (или его отсутствие в случае открытой капсулы),
475
набор и расположение сенсилл передней группы, количество обонятельных сенсилл
капсулы - являются специфичными для каждого подрода, в то время как располо-
жение (а в некоторых случаях и набор) посткапсулярных сенсилл видоспецифичны.
Так как в задачи автора не входит обсуждение видовых различий, а только различий
подродовых, в дальнейшем особенности топографии и строения посткапсулярных
сенсилл приведены не будут.
Результаты исследований автора и анализ литературы продемонстрировали, что
строение органа Галлера является специфичным для представителей подродовых груп-
пировок рода Ixodes. Приведем эти данные (рис. 4-7).
Подрод Ixodes s. str. (рис. 4A-4F, 6А-6D)
Всесветно распространенный подрод, характеризующийся широчайшим спектром
хозяев, биотопов и паразито-хозяинных связей. Включает как немобионтов (пастбищ-
ных паразитов), так и нидобионтов (гнездо-норовых паразитов) (Леонович, 2019).
Самый крупный по количеству видов подрод (свыше 60 видов) (Филиппова, 1977).
Дистальный бугорок (anterior knoll) крупный, хорошо выражен, на нем располага-
ются две пары сенсилл (пористый волосок обонятельной сенсиллы и короткий гладкий
волосок контактного хеморецептора) (Леонович, 1977, 1985) (рис. 1).
Углубление для сенсилл передней группы (anterior trough) обширное, довольно
глубокое, трапециевидное, окружено кутикулярным валиком (рис. 4А, 4С, 4E, 4F).
В передней группе один пористый, один тонкий, один конический и два бороздчатых
волоска (Леонович, 1976) (терминология см. Леонович, 2005) (рис. 1A; 4С, 4D). По-
ристый волосок передней группы сенсилл занимает дистально-латеральное положение
(рис. 4). Пористый волосок передней группы примерно в полтора раза длиннее осталь-
ных (рис. 4C, 4D).
Капсула закрытая, сообщается со средой довольно крупным капсулярным отвер-
стием округлой формы, у некоторых видов отверстие может быть проксимально из-
резано (рис. 4A, 4E, 4F). В капсуле располагаются пористые волоски 7 обонятельных
сенсилл (Леонович, 1977), обычно один волосок далеко высовывается из отверстия
наружу (рис. 4).
Посткапсулярных волосков 6: имеются центральная группа из четырех бороздчатых
волосков двуслойных хемо-терморецепторов (Леонович, 2005) и пара латеральных
вкусовых сенсилл (рис. 4).
У личинки дистальный бугорок несет не две, как у взрослых клещей, а одну пару
сенсилл, отсутствует также один из бороздчатых волосков передней группы сенсилл
(рис. 4В).
У изученных представителей подрода строение органа Галлера может довольно
сильно варьировать, особенно это касается размера капсулярного отверстия, степени
его открытости, а также наличия или отсутствия изрезанности краев.
476
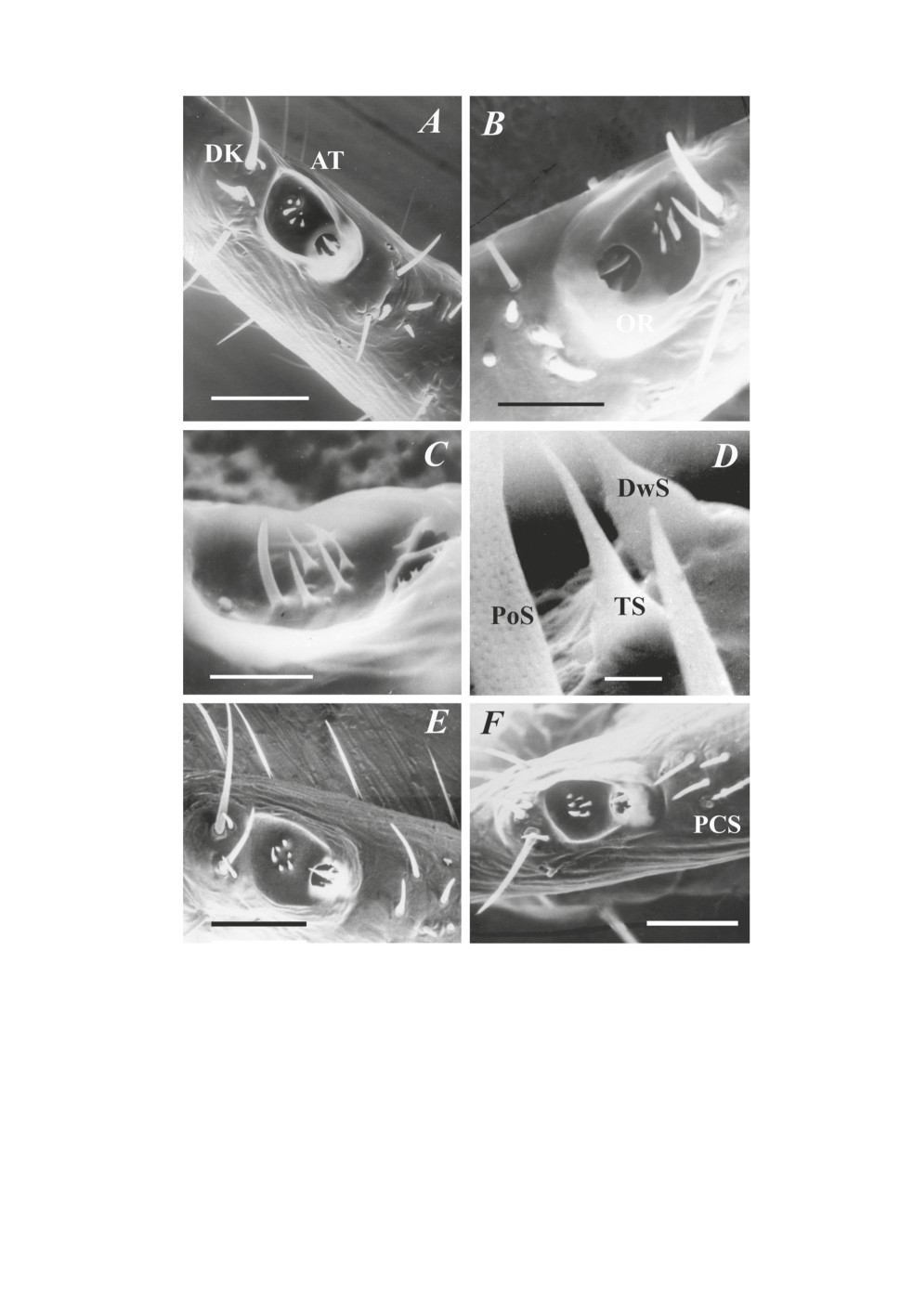
Рисунок. 1. Строение органа Галлера клещей подсемейства Ixodinae на примере таежного
клеща Ixodes persulcatus. A - внутреннее строение органа Галлера таежного клеща
по данным трансмиссивной электронной микроскопии (по: Леонович, 1977, 2005).
В - общий вид органа Галлера в растровом электронном микроскопе. С - схематическое
изображение органа Галлера (локализация сенсилл в основных отделах, форма переднего
углубления и капсулярного отверстия) органа Галлера таежного клеща, показанного
на рис. 1А и 1Б. Такие схемы использованы при иллюстрации текста (см. рис. 4-5).
AT - переднее углубление (anterior trough), в котором расположена передняя группа сенсилл
ATS (anterior trough sensilla); DK - дистальный бугорок (distal knoll); CA - капсула (capsule);
OR - капсулярное отверстие (capsular orifice); DKS - сенсиллы дистального бугорка (distal
knoll sensilla); CAS - капсулярные сенсиллы (capsular sensilla); PCS - посткапсулярные
сенсиллы (postcapsular senssilla); PoS - сенсилла с порами (porous sensillum) (обонятельная
сенсилла); DwS - сенсилла с дополнительными полостями (dowble-walled sensillum)
комбинированный хемо-терморецептор; BS - контактный хеморецептор, вкусовая сенсилла
(gustatory (upper-pore) sensillum); TS - терморецепторная сенсилла (thermo-sensitive
sensillum).
Figure 1. Structure of the Haller’s organ in ticks of the subfamily Ixodinae with an example of the
dog tick Ixodes ricinus. A - inner fine structure of the organ by the data obtained in transmissive
electron microscope (from: Leonovich, 1977; 2005). В - general view of the Haller’s organ in
scanning electron microscope. С - scheme of the organ (localization of sensilla in main parts of the
organ, shape of the anterior trough and capsular orifice) of the taiga tick I. persulcatus, shown in
Figs. 1A and 1B. Such schemes are used in graphic schemes in Figs 4 and 5).
477
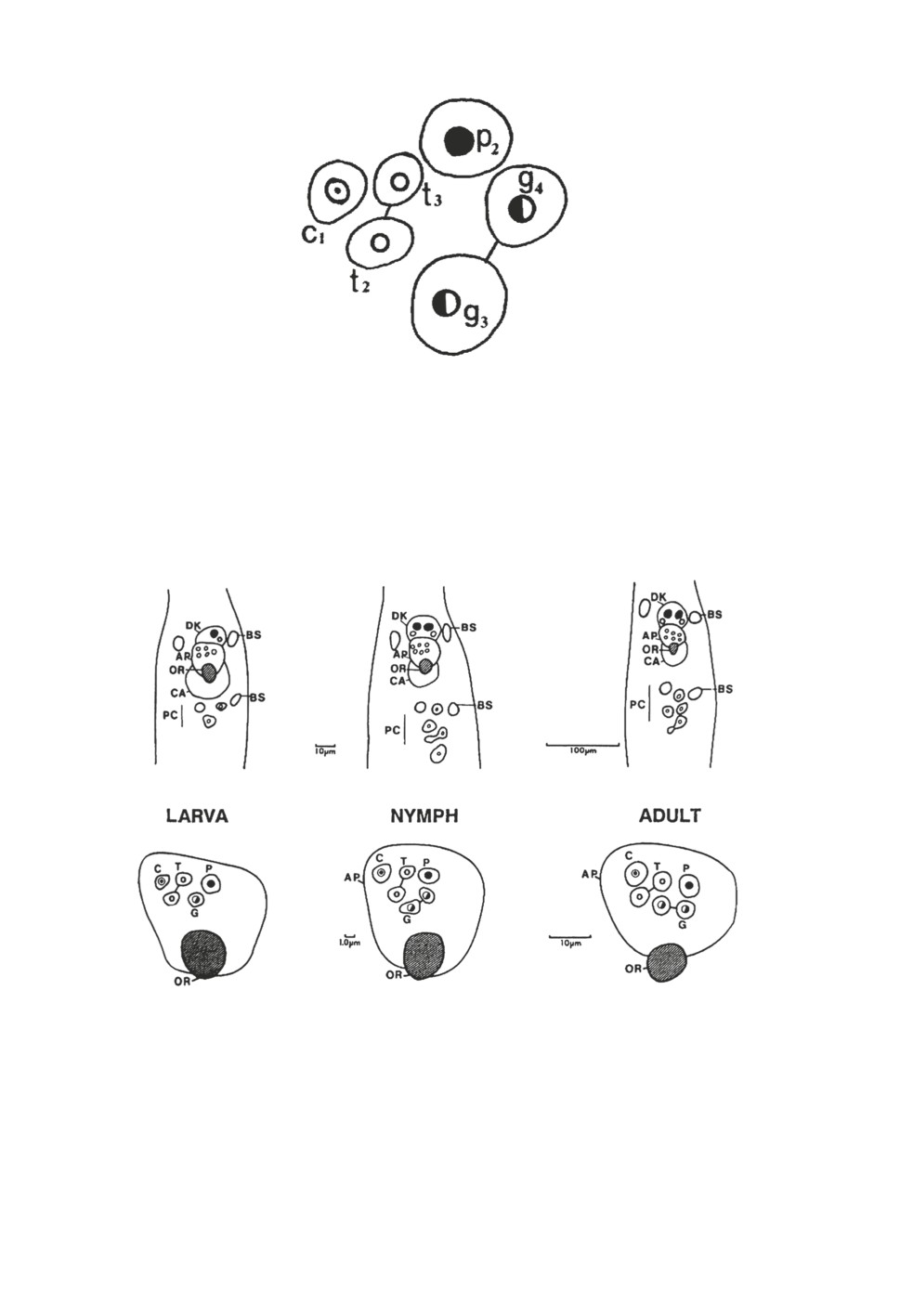
Рисунок 2. Топография сенсилл в передней группе (anterior trough sensilla) у клеща
Ixodes persulcatus, полученное путем наложения изображений 63 экземпляров (из работы
Leonovich, Belozerov, 1992). c1 - коническая сенсилла, p2 - пористая сенсилла, t2 тонкая
сенсилла; (t3) тонкая сенсилла; (g3) двухслойная сенсилла; (g4) двухслойная сенсилла.
Figure 2. Topography of anterior trough sensilla in the tick Ixodes persulcatus created
by superimposition of 63 images obtained in scanning electron microscope (from Leonovich,
Belozerov, 1992).
Рисунок 3. Топография сенсилл в передней группе (anterior trough sensilla) у клеща
Ixodes rubicundus (Neumann, 1904) на разных фазах онтогенетического развития, полученное
путем суперимпозиции (по: Belozerov et al., 1996). Условные обозначения как на рис. 1.
С - конический волосок (терморецептор); Т - тонкий волосок (гигрорецептор);
G - DwS (dowble-walled sensillum); Р - пористый волосок (обонятельная сенсилла).
Figure 3. Topography of anterior trough sensilla in the karoo paralysis tick Ixodes rubicundus
(Neumann, 1904) at different developmental stages obtained by superimposition of scanning
electron images (from Belozerov et al., 1996).
478
Рисунок 4. Строение органа Галлера у клещей рода Ixodes в растровом электронном
микроскопе: A - самка Ixodes ricinus (левая лапка); В - личинка I. ricinus (правая лапка);
С - I. redicorzevi, передняя группа сенсилл; D - I. persulcatus, часть передней группы сенсилл
с соновными типами сенсилл; E - I. persulcatus, самка; F - I. persulcatus, нимфа.
Масштабная линейка, мкм: A, E, F - 50; B - 20; C - 10 ; D - 1.
Figure 4. Structure of the Haller’s organ in ticks of the genus Ixodes in scanning electron
microscope: A - female of Ixodes ricinus (left tarsus); В - larva of I. ricinus (right tarsus);
С - I. redicorzevi, anterior group of sensilla; D - I. persulcatus, part of anterior pit sensilla
with main types of the latter; E - I. persulcatus, female; F - I. persulcatus, nymph.
Scale, µm: A, E, F - 50; B - 20; C - 10; D - 1.
479
Капсулярное отверстие может быть маленьким (составляющим менее пятой части
дорзо-проксимального размера углубления для сенсилл передней группы) (I. dentatus,
I. scapularis, I. loricatus), среднего размера (одна треть - одна четверть длины
углубления для передней группы сенсилл) (I. ricinus, I. persulcatus) или большим
(половина или треть длины углубления для сенсилл передней группы (I. muris,
I. boliviensis, I. asunumai). Задний край отверстия может быть изрезан (I. redocorzevi),
а может быть и гладким (I. scapularis, I. rubicundus).
Среди изученных видов особняком стоит I. luciae, отличающийся особым располо-
жением сенсилл передней группы (пористый волосок расположен отдельно от про-
чих, занимая самую дистальную позицию) и гантелевидным капсулярным отверстием
у взрослых клещей. При этом строение органа Галлера у личинки типично для под-
рода (Onofrio et al., 2010).
Также совершенно нетипично для подрода расположение сенсилл передней группы
у I. acututarsus, у которого сенсиллы передней группы расположены не компактно,
а в линию, при этом пористый волосок занимает центральное положение (Ronghang,
Roy, 2014)
Таким образом, для всех представителей подрода Ixodes s. str. характерны трапе-
циевидное углубление для сенсилл передней группы, как правило, окруженное вы-
раженным кутикулярным валиком, а также специфичная топография сенсилл перед-
ней группы (рис. 3, 4, 6A-6D). Вместе с тем различия в строении органа Галлера
у изученных представителей подрода Ixodes s. str. говорят о возможно сборном, поли-
филетическом характере данного подрода. При этим строение органа никак не связано
ни с характером паразитизма (нидобионты или немобионты), ни с природой хозяина.
Так, I. persulcatus, I. redicorzevi, I. ricinus - немобионты с самым широким кругом
хозяев (Филиппова, 1977); I. acutitarsus - распространенный в Юго-Восточной Азии
немобионт (Ronghang, Roy, 2014); I. boliviensis чаще всего паразитирует на собаках
в Новом Свете, I. dentatus - нидобионт, чаще всего встречается на кроликах, личинки
паразитируют на птицах (Smith, 1945); I. asanumai - паразит рептилий: I. granulatus -
нидобионт, паразитирует на тупайях; I. laguri - нидобионт, типичен в норах сусликов;
I. loricatus - паразит опоссумов в Южной Америке; I. luciae - нидобионт, также па-
разит опоссумов; I. scapularis - немобионт.
Таким образом, выявленные различия в строении органа Галлера данного подрода
косвенно говорят о его полифилии. Для решения этого вопроса необходимы даль-
нейшие таксономические исследования. Мнение о полифилии подрода Ixodes s. str.
разделяется и акарологами, использовавшими молекулярные методы (Xu et al., 2003).
Подрод Pholeoixodes (рис. 5А, 6Е-6Н).
Представлен исключительно нидобионтами, в основном паразитами птиц и,
в меньшей мере, млекопитающих.
480
Дистальный бугорок хорошо выражен, на нем располагаются 2 пары сенсилл дис-
тального бугорка (рис. 5A). Углубление для сенсилл передней группы округлое или
овальное (рис. 5A, 6E-6H), сенсилл передней группы 6, обычный набор. У всех изу-
ченных представителей подрода имеется одна весьма характерная черта, отличающая
клещей Pholeoixodes от всех остальных Ixodes. Капсулярное отверстие имеет особую
форму - оно вытянуто в дорзо-проксимальном направлении, обратно-каплевидное
(рис. 5A, 6E-6H). Сочетание округлого углубления для сенсилл передней группы и
изрезанного обратно-каплевидного капсулярного отверстия характерно для всех из-
ученных представителей подрода. У личинки отсутствует одна из двух пар сенсилл
дистального бугорка и одна из сенсилл передней группы (подобно тому, как это было
показано для клещей подрода Ixodes s. str. (рис. 6).
Строение органа Галлера у клещей подрода не зависит от круга хозяев. Так,
I. arboricola - паразит птиц-дуплогнездников (Heylen et al., 2014), I. lividus - встреча-
ется в норах ласточек-береговушек (Филиппова, 1977), а I. texanus - в норах мелких
млекопитающих (Darsie, Anastos, 1957).
Межвидовых различий в строении органа Галеры выявить не удалось.
Подрод Sternalixodes (рис. 6I-6L, 7А)
Представлен исключительно австралийскими видами (Австралия, Новая Зеландия,
Папуа) (9 видов) и характеризуется разнообразными местообитаниями и хозяевами.
Дистальный бугорок не выражен, сенсиллы переднего бугорка располагаются на
плоской поверхности лапки. В отличие от вышеописанных подродов, этот подрод
имеет всего одну пару этих сенсилл (рис. 6I-6L, 7А). Углубление для сенсилл перед-
ней группы овальное. В передней группе не 6, а 7 сенсилл, два пористых волоска,
очень крупных, один занимает самое дистальное, а второй - самое проксимальное
положение в группе (рис. 6I-6L, 7А). Капсулярное отверстие большое, в капсуле
5 обонятельных сенсилл.
У представителей этого подрода, также как у представителей Ixodes s. str., од-
нообразное, характерное для подрода строение органа Галлера связано только
с таксономией и не связано с характером паразитизма. Например, I. anatis - нидобионт,
паразитирующий на киви в Новой Зеландии; I. cordifer - паразитирует на сумчатых
дуплогнездниках (полосатый кускус) (т. е. тоже нидобионт), а I. hirsti и I. holocyclus -
немобионты, вызывающие паралич скота на пастбищах. Тем не менее строение органа
Галлера у них практически идентично.
Согласно некоторым современным работам, основанным на молекулярно-
генетических данных, Ixodes anatis не принадлежит к подроду Sternalixodes (Kwak
et al., 2017). Однако строение органа Галлера несомненно говорит о том, что этот
вид - типичный представитель указанного подрода (сравнить рис. 7А и рис. 6I-6L).
481
Рисунок 5. Строение органа Галлера у клещей рода Ixodes в растровом электронном
микроскопе: A - самка I. lividus (левая лапка); В - самка I.simplex; С - I. uriae, самка;
D - I. uriae, личинка; E - I. vespertilionis, самка; F - I. ovatus, самка.
Масштабная линейка, мкм: A, B - 30; C, D - 30; E, F - 50.
Figure 5. Structure of the Haller’s organ in ticks of the genus Ixodes in scanning electron
microscope: A - female of I. lividus (left tarsus); В - female of I.simplex; С - I. uriae, female;
D - I. uriae, larva; E - I. vespertilionis, female; F - I. ovatus, female.
Scale, µm: A, B - 30; C, D - 30; E, F - 50.
482
Рисунок 6. Схема внешнего строения органа Галлера у клещей рода Ixodes:
представители подродов Ixodes s. str. (A-D), Pholeoixodes (E-H), Sternalixodes (I-L):
A - I. ricinus; B - I. ricinus личинка; C - I. persulcatus; D - I. redicorzevi; E, F - I. lividus
(самка и личинка); G - I. arboricola; H - I. texanus (рисунок по фотографии из работы
(Homsher, Sonenshine, 1979); I - I. dengrolagi; J - I. confusus; K - I. hirsti; L - I. holocyclus
(все из работы Homsher et al., 1988).
Figure 6. Scheme of the structure of the Haller’s organ in ticks of the genus Ixodes:
subgenera Ixodes s. str. (A-D), Pholeoixodes (E-H), Sternalixodes (I-L):
A - I. ricinus; B - I. ricinus larva; C - I. persulcatus; D - I. redicorzevi; E, F - I. lividus
(female and larva); G - I. arboricola; H - I. texanus (drawing on the basis of image from
Homsher, Sonenshine, 1979; I - I. dengrolagi; J - I. confusus; K - I. hirsti; L - I. holocyclus
(all from Homsher et al., 1988).
483
Рисунок 7. Схема внешнего строения органа Галлера у клещей рода Ixodes:
представители подродов Sternalixodes (A), Ceratixodes (B), Multidentatus (C),
Trichotoixodes (D), Pomerantzevella (E), Endopalpiger (F), Ixodioipsis (G) и Eschatocephalus (H).
A - I. anatis; B - I. uriae; C - I. percavatus; D - I. frontalis; E - I. simplex; F - I. woyliei;
G - I. marxi; H - I. vespertilionis.
Figure 7. Scheme of the structure of the Haller’s organ in ticks of the genus Ixodes:
subgenera Sternalixodes (A), Ceratixodes (B), Multidentatus (C), Trichotoixodes (D),
Pomerantzevella (E), Endopalpiger (F), Ixodioipsis (G) and Eschatocephalus (H).
A - I. anatis; B - I. uriae; C - I. percavatus; D - I. frontalis; E - I. simplex; F - I. woyliei;
G - I. marxi; H - I. vespertilionis.
На примере этого подрода (как и вышеописанного подрода Pholeoixodes) особен-
но четко видно, что подродовые черты сохраняются у всех представителей подрода
(у исследованных 8 видов из 9 видов, входящих в состав подрода) и зависят только от
таксономического положения. Поэтому далее на рисунках автор приводит изображения
не всех изученных видов данного подрода, а только одного из них, имея в виду, что
у остальных орган Галлера устроен так же.
484
Подрод Ceratixodes (рис. 5С-5D, 7В)
Представлен единственным видом Ixodes uriae, который распространен
в приполярных областях Северного и Южного полушарий и является типичным оби-
тателем птичьих базаров.
Передний бугорок, как и углубление для сенсилл передней группы, отсутствует,
все сенсиллы обоих отделов располагаются на плоской поверхности передней лапки
(рис. 5С).
Сенсилл дистального бугорка две, но обе они представлены крупными пористыми
волосками, т.е. редуцированные вкусовые сенсиллы отсутствуют.
В передней группе 7 сенсилл, имеются два пористых волоска, как у представителей
подрода Sternalixodes (см. выше), также похож на представителей подрода Sternalixodes
и по характеру расположения этих сенсилл (рис. 7В). Капсула открытая, имеется не-
большое углубление, в котором видны 7 пористых волосков обонятельных сенсилл
(рис. 5С).
У личинки отсутствуют один пористый и один бороздчатый волоски в составе
передней группы сенсилл, в капсуле только 4 обонятельные сенсиллы (рис. 5D).
Подрод Multidentatus (рис. 7С)
Представители подрода связаны исключительно с птицами, преимущественно мор-
скими (Hoogstraal, Aeschlimann, 1982).
Возвышение дистального бугорка не выражено, две пары сенсилл, каждая из ко-
торых включает длинный пористый и короткий вкусовой волоски, располагаются на
прямой поверхности передней лапки.
В передней группе сенсилл 6 волосков, пористый занимает положение, показанное
на рис. 7С. Капсула открытая, содержит 7 обонятельных сенсилл (рис. 7С). У обоих
изученных видов (I. paranaensis и I. percavatus) строение органа Галлера практически
идентично.
Подрод Trichotoixodes (рис. 7D)
Паразиты птиц (Buczek et al., 2016).
У двух изученных видов (I. frontalis и I. brunneus) орган Галлера устроен сходно.
Передний бугорок хорошо выражен, крупный, на нем располагаются 6 сенсилл:
4 длинных пористых обонятельных волоска и 2 коротких вкусовых волоска (характер
расположения сенсилл показан на рис. 7D). Углубление сенсилл передней группы от-
четливое, окружено кутикулярным валиком, расположение сенсилл сходно с таковым
у Ixodes s. str. Капсулярное отверстие большое, округлое, число сенсилл в капсуле
определить не удалось.
485
Подрод Pomerantzevella (рис. 5В, 7Е)
Широко распространенные (Европа, Африка, Юго-Восточная Азия, Австралия,
Новая Зеландия) паразиты рукокрылых. Кроме изученного автором I. simplex, подрод
включает еще один вид - Ixodes spiculae Arthur, 1956, известный только по одному
самцу из Африки (без данных о месте находки и хозяине) (Филиппова, 1977).
Передний бугорок хорошо развит, с двумя парами сенсилл (рис. 5В, 7Е). Сенсил-
лы передней группы и сенсиллы капсулы находятся в одном общем углублении (рис.
5В). Расположение сенсилл передней группы сходно с таковым представителей Ixodes
s. str. (рис. 7Е).
Подрод Endopalpiger (рис. 7F)
Преимущественно австралийский род, в основном паразиты сумчатых
и однопроходных.
Углубление для сенсилл передней группы овальное, окружено валиком, волосков
дистального бугорка одна пара (рис. 7F). Сенсилл передней группы 7 (два пористых
волоска, расположение показано на рис. 7F). Капсулярное отверстие овальное, слегка
вытянутое в поперечном направлении, в капсуле 5 сенсилл.
Подрод Ixodiopsis (рис. 7G)
Голарктический подрод включает нидобионтов и немобионтов, для последних
обычны каменистые местообитания, нападают на хозяев с поверхности почвы (Фи-
липпова, 1977).
Строение органа Галлера близко таковому Ixodes s. str., однако углубление для
сенсилл передней группы слегка вытянуто в поперечном членику направлении, от-
верстие капсулы также почти щелевидное (рис. 7G). К сожалению, плохое качество
фотографий, приведенных в работе Homsher, Sonenshine, 1975, не позволяет рассмо-
треть строение органа более подробно.
Подрод Eschatocephalus (рис. 5Е, 7F)
Согласно данным Филипповой (1977), достоверно известен один вид - изученный
автором Ixodes vespertilionis, паразит рукокрылых. Недавно был описан второй вид
этого подрода I. ariadnae Hornok et al., 2014, орган Галлера которого был исследован
Хорноком с соавторами (Hornok et al., 2015). Строение органа Галлера у обеих видов
практически идентично.
Дистальный бугорок развит, передняя часть бугорка сглажена, на бугорке распола-
гаются две пары сенсилл (Рис. 5Е). Углубление для сенсилл передней группы узкое,
сильно вытянутое в дорзо-проксимально направлении. Сенсиллы передней группы
расположены в линию, пористый волосок находится примерно посередине линии (рис.
5Е, 7F). Капсула открытая, отделена от переднего углубления кутикулярным валиком.
Имеются 7 капсулярных сенсилл.
486
Подрод Partipalpiger (рис. 5F)
Строение органа Галлера очень близко к таковому Ixodes s. str. Для подрода
Partipalpiger характерны округлое капсулярное отверстие с неизрезанными краями,
сближенное положение волосков дистального бугорка и мелкие размеры редуциро-
ванных вкусовых сенсилл.
Подрод Afrixodes (рис. 3)
Строение органа Галлера на всех фазах развития показано на примере I. rubicundus
(Belozerov et al., 1996).
Таким образом, анализ строения органа Галлера у клещей подсемейства Ixodinae,
представленного в мировой фауне единственным родом Ixodes, показал, что суще-
ственные различия в морфологии касаются представителей отдельных подродов, в
то время как межвидовые различия в пределах отдельного подрода практически не-
существенны. Особенно это касается четко установленных и явно монофилетических
подродов, таких как Pholeoixodes, Sternalixodes, Ceratixodes, Multidentetus. В то же
время наибольшие внутриподродовые вариации наблюдаются в пределах подродов,
чей таксономический статус окончательно не определен (например, Ixodes s. str.). Эти
обстоятельства позволяют рекомендовать использование данных по строению органа
Галлера в систематике клещей рода Ixodes (и других родов, что будет показано во
втором сообщении). Отметим, что во всех работах, посвященных систематике клещей
рода Ixodes, данные по органу Галлера никак не используются. Использование этих
данных, наряду с данными, полученными молекулярными методами, в случае неясной
картины, позволили бы склонить чашу весов на ту или иную сторону.
Например, в работе Клиффорда с соавторами (Clifford et al., 1973) под-
род Trichotoixodes включен в состав Ixodes s. str., в то время как орган Галлера
у представителей этих подродов различается очень сильно (сравни рис. 6A-6D и рис.
7D). Подрод Phoileoixodes синонимизируется с подродом Ixodiopsis, что никак не со-
ответствует строению органа Галлера у представителей этих подродов (см. рис. 6Е-6Н
и рис. 7G). Еще один пример. В работе Charrier et al. (2019) филогенетические рекон-
струкции, выполненные по данным транскриптомного анализа представителей Ixodes
s. str., показали, что I. holocyclus (подрод Sternalixodes) и Ixodes uriae (Ceratixodes)
образуют общую кладу, отдельную от клады, образованной видами I. ricinus,
I. pertsulcatus, I. scapularis (подрод Ixodes s. str.). Это в целом соответствует особен-
ностям строения у них органа Галлера.
487
БЛАГОДАРНОСТИ
Работа выполнена по Государственному заданию «Разнообразие паразитарных систем,
адаптаций и путей эволюции паразитов» (номер темы: АААА-А19-119020690109-2).
В работе были использованы материалы коллекции Зоологического института РАН
(ЗИН РАН) (УФК ЗИН рег. № 2-2.20).
Список литературы
Леонович С.А. 1976. Морфология органа Галлера у клещей семейства Ixodidae (Acarina: Ixodoidea). Ис-
следования в растровом электронном микроскопе. Проблемы зоологии. Л., Наука, с. 59-61.
Леонович C.А. 1977. Электронно-микроскопическое исследование органа Галлера клеща Ixodes persulcatus
(Ixodidae). Паразитология 11 (4): 340-347.
Леонович С.А. 1985. Органы чувств. В кн. Таежный клещ Ixodes persulcatus Sch. (Acarina, Ixodidae). Мор-
фология, систематика, экология, медицинское значение. Л.: Наука, с. 77-83.
Леонович С.А. 2005. Сенсорные системы паразитических клещей. СПб., Наука, 235 с.
Леонович С.А. 2019. О типах паразитизма иксодовых клещей (Ixodidae). Паразитология 53 (5): 416-420.
DOI: 10.1134/S0031184719050053
Филиппова Н.А. 1977. Иксодовые клещи подсемейства Ixodinae. Фауна СССР. Паукообразные. Т. 4. Вып.
4. Л., Наука, 386 с.
Ash A., Elliot A., Godfrey S., Burmej H., Abdad M.Y., Northover A., Wayne A., Morris K., Clode P., Lymbery
A., Thompson R.C.A. 2017. Morphological and molecular description of Ixodes woyliei n. sp. (Ixodidae)
with consideration for co-extinction with its critically endangered marsupial host. Parasites and Vectors 10,
Article number: 70.
Barker S. C., Murrell A. 2004 . Systematics and evolution of ticks with a list of valid genus and species names.
Parasitology 129: S15-S36. 10.1017/S0031182004005207
Barros-Battesti D.M., Arzua M., Pichorim M., Keirans J.E. 2003. Ixodes (Multidentatus) paranaensis n. sp. (Acari:
Ixodidae) a parasite of Streptoprocne biscutata (Sclater 1865) (Apodiformes: Apodidae) birds in Brazil.
Belozerov V.N., Kok D.J., Fourie L.J., Leonovich S.A. 1996. The external structure of Haller’s sensory organ
in the Karoo paralysis tick Ixodes (Afrixodes) rubicundus Neum. (Ixodidae). Proceedings IX International
Congress of Acarology (Michigan), 1996, pp. 397-399.
Buczek A., Bartosik K., Olszewski T., Stepien K., Kubrak Т., Sałata М. 2006. Host specificity of ticks (Acari:
Ixodida). In: Arthropods. Epidemiological importance. Koliber, Lublin, pp. 37-54.
Charrier N.P., Hermouet A., Hervet C., Agoulon A., Barker S.C., Heylen D., Toty C., McCoy K.D.,
Plantard O., Rispe C. 2019. A transcriptome-based phylogenetic study of hard ticks (Ixodidae). Scientific
Reports 9, Article number 12923.
Clifford C.M., Sonenshine D.E., Keirans J.E., Kohls G.M. 1973. Systematics of the Subfamily Ixodinae (Acarina:
Ixodidae). 1. the Subgenera of Ixodes. Annals of the Entomological Society of America 66 (3): 489-
Darsie R.F., Anastos G. 1957. Geographical distribution and hosts of Ixodes texanus Banks (Acarina, Ixodidae).
Haller G. 1881 . Vorlaufige Bemerkungen Uber das Gehurorgan der Ixodiden. Zoologische Anzeiger 4: 165-167.
Hayashi F., Hasegawa M. 1983. Immature stages and reproductive characteristics of the lizard tick, Ixodes asanumai
aez.18.315
488
Heylen D.J.A., Van Oosten A.R., Devriendt N., Elst J. 2014. Seasonal feeding activity of the tree-hole tick, Ixodes
Homsher P.J., Sonenshine D.E. 1975. Scanning electron microscopy of ticks for systematic studies: fine structure
of Haller’s organ in ten species of Ixodes. Transactions of the American Microscopical Society 94 (3):
Homsher P.J., Sonenshine D.E. 1979. Scanning electron microscopy of ticks for systematic studies: 3. Structure
of Haller's organ in five species of the subgenus Multidentatus of the genus Ixodes. In Recent advances in
acarology, Vol II. N.Y., Academic Press, pp. 485 491.
Homsher P.J., Keirans J.E, Robbins R.G., Irwin-Pinkley L., Sonenshine D.E. 1988. Scanning electron microscopy
of ticks for systematic studies: structure of Haller's organ in eight species of the subgenus Sternalixodes of
jmedent/25.5.348
Homsher P.J., Sonenshine D.E. 1977. Scanning electron microscopy of ticks for systematic studies 2. Structure of
Hallerʼs organ in Ixodes brunneus and Ixodes frontalis. Journal of Medical Entomology 14 (1): 93-97. https://
doi.org/10.1093/jmedent/14.1.93
Honzаkovа E., Sixl W., Waltinger H. 1975. Scanning electron microscopy of the ticks Ixodes laguri and Ixodes
arboricola: surface structures of Haller's organ. Folia Parasitologica 22: 241- 243.
Hoogstraal H., Aeschlimann A. 1982. Tick-host specificity. Bulletin de la Société Entomologique Suisse 55: 5-32.
Hornok S., Kontschán J., Estrada-Peña A., de Mera I.G.F., Tomanović S., de la Fuente J. 2015. Contributions to
the morphology and phylogeny of the newly discovered bat tick species, Ixodes ariadnae in comparison
with I. vespertilionis and I. simplex. Parasites and Vectors 8: 47.
Kwak M.L., Beveridge I., Koehler A.V., Malipatil M., Gasser R.B., Jabbar A. 2017. Phylogenetic analysis of the
Australasian paralysis ticks and their relatives (Ixodidae: Ixodes: Sternalixodes). Parasites & Vectors 10,
Article number: 122.
Leonovich S.A. 2004. Phenol and lactone receptors in the distal sensilla of the Haller's organ in Ixodes ricinus
ticks and their possible role in host perception. Experimental and Applied Acarology 32 (1-2): 89-102.
DOI:10.1023/B:APPA.0000018200.24760.78
Leonovich S.A., Belozerov V.N. 1992 . Regeneration of Haller's sensory organ in the tick Ixodes ricinus L.
Experimental and Appliaed Acarology 15: 59-79.
Leonovich S.A., Dimov I. 2012. Sense organs on palps and fore tarsi of gamasid mites (Parasitiformes, Rhinonyssidae),
parasites of the nasal cavity of the great tit, the rock dove, and the Eurasian coot. Entomological Review 92
(46): 1012-1019. DOI: 10.1134/S0013873812090084
Leonovich S.A., Stanyukovich M.K. 2011. Sensory organs of mesostigmatic mites (Acarina, Mesostigmata) dwelling
in body cavities of mammals and birds. Proceedings of the Zoological Institute RAS 315 (3): 263-273.
Onofrio V.C., Labruna M.B., Faccini J.L.H., Barros-Battesti D.M.. 2010. Description of immature stages and
redescription of adults of Ixodes luciae Sénevet (Acari: Ixodidae). Zootaxa 2495: 53-64.
Ronghang B., Roy B. 2014. A Scanning electron microscopic observations on sensilla of Ixodes acutitarsus
recovered from semi-wild cattle Bos frontalis Lambert. Entomology and Applied Science Letters 1 (4): 8-15.
Shao R., Barker S.C., Mitani H., Aoki Y., Fukunaga M. 2005 . Evolution of duplicate control regions in the
mitochondrial genomes of Metazoa: a case study with Australasian Ixodes ticks. Molecular Biology and
Smith C.N. 1945. Biology of Ixodes dentatus Neumann (Ixodidae). Annals of the Entomological Society of America
Xu G., Fang Q.Q., Keirans J.E., Durden L. A. 2003. Molecular phylogenetic analyses indicate that the Ixodes
3395(2003)089[0452:MPAITT]2.0.CO;2
489
THE STRUCTURE OF THE HALLER’S ORGAN AND TAXONOMY
OF IXODID TICKS OF THE SUBFAMILY IXODINAE
(FAMILY IXODIDAE)
S. A. Leonovich
Keywords: Haller’s organ, scanning electron microscopy, Ixodinae, Ixodes, taxonomy
Summary
The analysis of the structure of the Haller’s organ, the main sensory organ of ixodid ticks, was
performed on the basis of own and literary data, obtained by methods of scanning electron microscopy.
A total of 43 hard tick species of the genus Ixodes Latreille, 1795, solitary genus of the subfamily
Ixodinae, were examined, including representatives of 13 subgenera of the genus: Ixodes s. str.
(14 species), Pholeoixodes Schulze, 1942 (3 species), Ixodiopsis Filippova, 1957 (4 species), Ceratixodes
Neumann, 1902 (1 species), Scaphixodes Schulze 1941 (2 species), Trichotoixodes Reznik, 1961
(2 species), Partipalpiger Hoogstraal, Clifford, Saito & Keirans 1973 (1 species), Pomerantzevella
Feider, 1965 (1 species), Eschatocephalus Frauenfeld, 1851 (2 species), Afrixodes Morel, 1966
(1 species), Sternalixodes Schulze, 1938 (8 species), Multidentatus Clifford, Sonenshine, Keirans &
Kohls, 1973 (3 species), Endopalpiger Schulze (1935) (1 species). Taxonomic characters in the structure
of the organ (the number and topography of sensilla in different parts of the organ, the presence and
shape of the capsular orifice, shape of the anterior trough, etc.) were revealed. It was demonstrated that
that the structure of the Haller’s organ is specific for each subgenus, whereas interspecific differences
within each certain subgenus are virtually very poor. Comparison of the obtained data with existing
views on taxonomy of the genus Ixodes was performed.
490