ПАРАЗИТОЛОГИЯ, 2022, том 56, № 6, с. 469-476.
УДК 593.5: 592/599
Myxidium rhodei Leger, 1905 (Cnidaria, Myxosporea):
сезонная, возрастная, пространственная
и многолетняя динамика зараженности
плотвы Rutilus rutilus
в Чивыркуйском заливе озера Байкал
© 2022 г. М. Д. Батуеваa,*, Ж. Н. Дугаровa, Л. Д. Сондуеваa
aИнститут общей и экспериментальной биологии СО РАН,
ул. Сахьяновой 6, Улан-Удэ, 670047 Россия
*e-mail: badmm_@rambler.ru
Поступила в редакцию 11.08.2022 г.
После доработки 30.10.2022 г.
Принята к публикации 03.11.2022 г.
Миксоспоридия Myxidium rhodei Leger, 1905 - распространенный паразит палеарктических
карповых рыб, локализующийся преимущественно в боуменовых капсулах почек рыб. В оз.
Байкал плотва Rutilus rutilus заражена миксоспоридией M. rhodei при высокой экстенсивности
(94-100%) и интенсивности инвазии (до 6000 псевдоцист) в 1999-2021 гг. В данной работе был
проведен анализ возрастной, сезонной, многолетней динамики и пространственного распреде-
ления зараженности M. rhodei плотвы в Чивыркуйском заливе оз. Байкал. Паразит начинает
регистрироваться у годовиков, затем интенсивность инвазии плотвы M. rhodei увеличивается,
достигая максимума у рыб среднего возраста, у старшевозрастных - снижается. M. rhodei
имеет годичный цикл развития в плотве. Колебания зараженности M. rhodei исследуемых рыб
в течение 25 лет наблюдений коррелируют с многолетними изменениями гидрологического
режима оз. Байкал.
Ключевые слова: Байкал, плотва, миксоспоридии, почки, псевдоцисты, Myxidium rhodei
DOI: 10.31857/S0031184722060035; EDN: FIGUMR
Миксоспоридии - широко распространенные эндопаразиты, имеющие двуххозяин-
ный жизненный цикл, где окончательными хозяевами являются олигохеты и полихеты,
а промежуточными позвоночные, преимущественно костистые рыбы. Разнообразие мик-
соспоридий велико, в мире насчитывается свыше 2500 видов (Okamura et al., 2018). Род
Myxidium Bütschli, 1882 представлен 230 видами, которые локализуются преимуще-
ственно в желчном пузыре рыб (Eiras et al., 2011; Espinoza et al., 2017; Fariya et al.,
2020). Myxidium rhodei Leger, 1905 - один из немногих видов из этого рода, имеющих
иную локализацию (боуменовы капсулы почек). Типовым хозяином M. rhodei явля-
469
ется обыкновенный горчак Rhodeus amarus (Léger, 1930), однако описание от этого
хозяина скудное. Позднее M. rhodei был отмечен более чем у 40 видов рыб (Шульман,
1984), но наиболее часто встречаются сведения о зараженности M. rhodei плотвы
обыкновенной Rutilus rutilus (Кашковский, 1967; Гуркина, 1983; Dyková et al., 1987;
Kepr, 1987; 1991; Athanassopoulou, Sommerville, 1993a, 1993b; Longshaw et al., 2005;
Dzika et al., 2006; Batueva et al., 2015). Кроме того, M. rhodei наряду с видами рода
Kudoa являлся удобным для исследователей объектом изучения интенсивности инва-
зии среди миксоспоридий (Oliva et al., 1992; Ware et al., 2014). Это вероятно, связано
с возможностью визуального подсчета плазмодиев в тканях под стереомикроскопом.
Так, M. rhodei в боуменовой капсуле почек локализуется по одному плазмодию (Dyk-
ová et al., 1987; Batueva et al., 2020).
Морфологические, молекулярные и патогистологические исследования M. rhodei от
плотвы из Чивыркуйского залива были проведены Батуевой и др. (Batueva et al., 2020).
В данной статье представлен анализ сезонной, возрастной и многолетней динамики
зараженности плотвы M. rhodei в Чивыркуйском заливе оз. Байкал.
МАТЕРИАЛ И МЕТОДЫ
Было исследовано 942 экз. плотвы в Чивыркуйском заливе оз. Байкал в районах Монахово
(53°67′N, 109°02′E), оз. Арангатуй (53°59′N, 109°05′E), бухта Онкогон (53°45′N, 109°10’E).
Сезонную, возрастную и многолетнюю динамику зараженности рыб исследовали в Монахово.
Для изучения сезонной динамики 360 рыб возраста от 5+ до 7+ исследовано в течение зимы
(январь), весны (март), лета (конец июня, начало июля) и осени (октябрь) в 2000-2002 гг.
Для анализа возрастной динамики исследовано 120 рыб в возрасте от 0+ до 8+. Для изучения
пространственного распределения 270 рыб возраста от 5+ до 7+ из районов Монахово, оз.
Арангатуй, бухта Онкогон исследовано в конце июня-начале июля. Возраст определяли по
чешуе (Hile, 1936).
Экстенсивность инвазии (ЭИ, %) определяли согласно Bush et al. (1997). Интен-
сивность инвазии и индекс обилия выявляли путем подсчета капсул (псевдоцист)
в почках, раздавленных между двумя стеклами размером 10 х10 см под стереомикроскопом
МБС-10 (Ломо, Россия) при увеличении x20.
Статистический анализ проведен путем сравнения различий методом Kruskal-Wallis test
(H-test) с p < 0.05 и множественных сравнений (post-hoc) для непараметрических данных
“Statistica, ver. 8, StatSoft Russia”.
РЕЗУЛЬТАТЫ
Возрастная динамика зараженности
В течение всего периода наблюдений экстенсивность инвазии плотвы M. rhodei
составляла 97% (918/942) M. rhodei начинает регистрироваться у годовиков плотвы
и ЭИ составляет 61%. У двухлетних рыб зараженность увеличивается до 95%, затем
достигает 100 % у рыб от 3+ до 8+. Индекс обилия M. rhodei постепенно увеличива-
ется у рыб в возрасте до 6+ и достигает максимума (1665 псевдоцист), затем индекс
обилия у старшего возраста достоверно (P ≤ 0.05) снижается у 7+ (635); 8+ (332)
и 9+ (208 псевдоцист, соответственно) (рис. 1).
470
Рисунок 1. Возрастная динамика зараженности плотвы R. rutilus миксоспоридией M. rhodei
в Чивыркуйском заливе оз. Байкал.
Figure 1. Age-related dynamics of the roach R. rutilus infestation with M. rhodei myxosporeans
in the Chivyrkui Gulf of Lake Baikal.
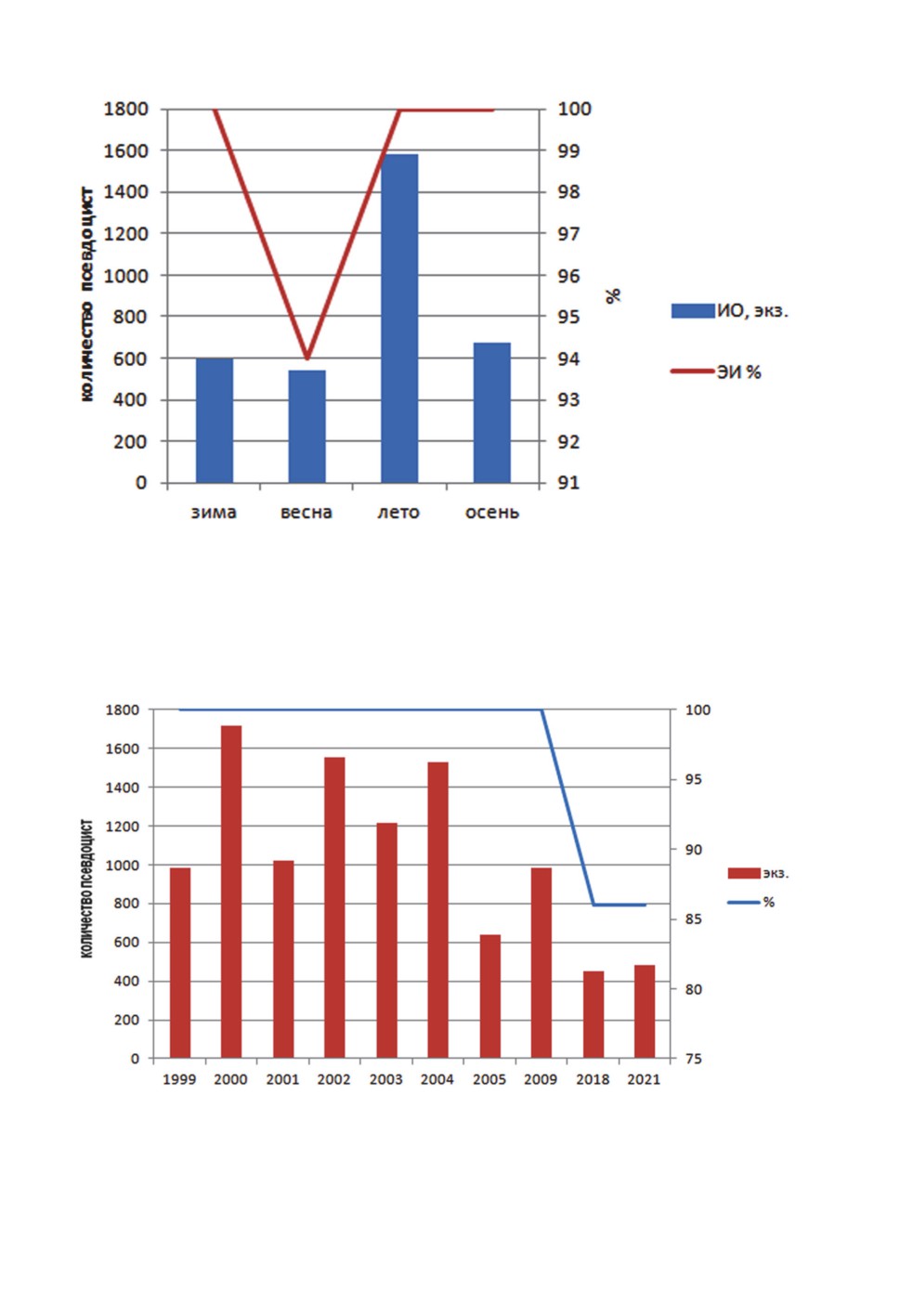
Сезонная динамика зараженности
В течение всего года экстенсивность инвазии была высокой (94 весной и 100%
в остальные сезоны). Более информативны показатели индекса обилия M. rhodei.
У плотвы ЭИ меняется в течение всего года с максимумом летом (1578 псевдоцист),
осенью, зимой и весной индекс обилия меньше (рис. 2). Летние показатели досто-
верно выше (P ≤ 0.05), чем в остальные сезоны года.
Пространственная динамика зараженности
Экстенсивность инвазии плотвы M. rhodei во всех точках Чивыркуйского зали-
ва летом составляла 100%. Различия наблюдаются только в интенсивности инвазии
плотвы.
Интенсивность заражения плотвы варьировала от максимальной (до 6000 псев-
доцист) в эвтрофной части Чивыркуйского залива (оз. Арангатуй), в средней части
залива Монахово (1578), до (600-20 псевдоцист) в олиготрофной части залива - бухты
Онкогон. Различия между точками исследования оз. Арангатуй - бухта Монахово -
бухта Онкогон достоверно различаются (P ≤ 0.05).
Многолетняя динамика зараженности
Нами исследована зараженность плотвы в периоды: 1999-2005, 2007, 2009, 2018,
2021. Экстенсивность инвазии M. rhodei у плотвы в течение всего времени исследо-
вания была высокой (рис. 3). Незараженные рыбы (4%) отмечались только в 2018
и 2021 гг. Индекс обилия был максимален (в среднем 1406 псевдоцист) в 2000-2004
годах, затем зараженность достоверно (P ≤ 0.05) уменьшалась в 2018 г. (449 псевдо-
цист) и 2021 г. (485 псевдоцист).
471
Рисунок 2. Сезонная динамика зараженности плотвы R. rutilus миксоспоридией M. rhodei
в Чивыркуйском заливе оз. Байкал.
Figure 2. Seasonal dynamics of the roach R. rutilus infestation with M. rhodei myxosporeans
in the Chivyrkui Gulf of Lake Baikal.
Рисунок 3. Многолетняя динамика зараженности плотвы R. rutilus миксоспоридией
M. rhodei в Чивыркуйском заливе оз. Байкал.
Figure 3. Long-term dynamics of the roach R. rutilus infestation with M. rhodei myxosporeans
in the Chivyrkui Gulf of Lake Baikal.
472
ОБСУЖДЕНИЕ И ЗАКЛЮЧЕНИЕ
M. rhodei доминирует в сообществе паразитов плотвы в оз. Байкал (Дугаров
и др., 2011). У плотвы наблюдается максимальная зараженность, по сравнению с за-
раженностью рыб M. rhodei по данным из литературных источников.
Анализ сезонной динамики зараженности плотвы M. rhodei были проведены
Кашковским (1967) в Ириклинском водохранилище, Гуркиной (1983) в оз. Врево
и в озерах Финляндии (Brummer-Korvenkontio et al., 1991). В Ириклинском водохра-
нилище у плотвы зараженность M. rhodei была максимальной зимой, а минимальной
летом. Гуркина (1983) отмечает высокую частоту встречаемости M. rhodei с июня по
сентябрь с максимумом в июле. В октябре происходит некоторое снижение, низкая
зараженность в марте и в апреле. В четырех озерах Финляндии колебания сезонной
зараженности рыб в озерах различной трофности не синхронны, но, тем не менее,
наблюдается небольшое снижение зараженности в конце весны-начале лета, в конце
лета и октябре-ноябре. Причины колебаний зараженности плотвы, по мнению этих
авторов, связаны со скоростью седиментации спор и особенностями питания рыб
в течение года. Однако, как было обнаружено, миксоспоридии имеют сложный жиз-
ненный цикл, поэтому причиной колебаний зараженности рыб могут быть особен-
ности их заражения актиноспорами. Также было выявлено, что температура воды
влияет на созревание и выход актиноспор во внешнюю среду (Yokoyama et al., 1993;
El-Matbouli et al., 1999).
Согласно нашим гистологическим исследованиям (Batueva et al., 2020), жизненный
цикл M. rhodei - годичный: летом плазмодии молодые с псевдоподиями, некоторые из
них локализуются в интерстициальной ткани. Созревание спор достаточно быстрое,
уже в конце лета-начале осени мы наблюдаем зрелые споры в соединительнотканных
капсулах. Вполне вероятно, что в это время созревшие споры могут быть вирулентны-
ми и могут попасть во внешнюю среду, а псевдоцисты могут подвергнуться лизису.
Осенью мы отмечаем уменьшение количества псевдоцист, по сравнению с началом
и серединой лета. Массовая дегенерация соединительнотканных капсул парази-
та отмечается зимой, продолжается весной, когда количество паразитов достигает
минимума. Полное освобождение от паразита в почках в течение года нами не от-
мечается, незначительная часть псевдоцист со зрелыми спорами может оставаться
в почках больше года, затем псевдоцисты постепенно элиминируются. Их количество
незначительно, в июне отмечается массовое заражение новой генерацией M. rhodei.
Сезонность в динамике зараженности плотвы в Чивыркуйском заливе сходна
с динамикой из оз. Врево (Гуркина, 1983) по сравнению с таковым в Ириклинском
водохранилище и в четырех озерах Финляндии. По мнению Кашковского (1966),
продолжительность жизненного цикла M. rhodei в Ириклинском водохранилище -
около трех месяцев и жизненный цикл может многократно повторяться в течение
года. Вероятно, одна из причин - теплые воды Ириклинского водохранилища, так
как температура воды играет существенную роль в созревании и выходе актиноспор
во внешнюю среду (El-Matbouli et al., 1999).
В озерах Финляндии (Brummer-Korvenkontio et al., 1991) отмечали уменьшение
зараженности у рыб старшего возраста, которое авторы связывали с усилением
473
иммунного ответа в этой возрастной группе. По нашему мнению, уменьшение за-
раженности плотвы M. rhodei в возрасте 7+ - 9+ в оз. Байкал вероятнее всего свя-
зано с тем, что в более глубоких слоях залива обитают популяции крупной плотвы,
а популяции плотвы младших возрастных групп - на мелководьях (Картушин, 1958).
Пространственные изменения зараженности рыб M. rhodei отмечались в различных
по трофности водоемах Финляндии и Польши (Brummer-Korvenkontio et al., 1991;
Dzika et al., 2006). Так, исследователями отмечена положительная связь между за-
раженностью и интенсивностью заражения M. rhodei у плотвы в зависимости от
трофического уровня водоема: в олиготрофном озере Пеурунка (Финляндия) интенсив-
ность заражения была ниже (65%), чем в эвтрофном озере Саравеси (79%) (Brummer-
Korvenkontio et al., 1991). В то же время в озерах Польши с разной степенью эв-
трофности зараженность плотвы была выше в слабо эвтрофном, с широкой зоной
фитолиторали, озере, чем в загрязненном гиперэвтрофированном озере с процессами
дегенерации фитолиторали (Dzika et al., 2006).
Многолетние изменения динамики зараженности данным паразитом рыб, вероятно,
связаны с изменением гидрологического режима оз. Байкал (маловодный период
1999-2015 и многоводный 2018-2021) (Гармаев, Цыдыпов, 2019; Никитин, 2021).
Связь между колебаниями уровня воды и изменениями структуры инфрасообществ
миксоспоридий отмечалась Маркоглисом и др. (Marcogliese et al., 2009; Marcogliese,
Cone, 2021). Низкий уровень воды и эвтрофирование способствовали увеличению
количества видов миксоспоридий. Заражению рыб миксоспоридиями способствуют
факторы, увеличивающие плотность актиноспор, - это малая глубина водоема, слабая
проточность и прогреваемость (Hallett, Bartholomew, 2007; Marcogliese et al., 2009;
Marcogliese, Cone, 2021).
Таким образом, результаты наших исследований подтверждают литературные дан-
ные, что в хорошо прогреваемых мелководных частях водоемов с медленным тече-
нием создаются благоприятные условия для обитания олигохет и велика вероятность
заражения рыб миксоспоридиями, в частности M. rhodei.
БЛАГОДАРНОСТИ
Работа выполнена в рамках темы гос. заданий (регистрационный номер
АААА-А17-117011810039-4) и «Влияние изменения уровня воды в озере Байкал на
состояние экосистемы озера, определение ущерба объектам экономики и инфраструк-
туры прибрежной территории Республики Бурятия, Иркутской области в зависимости
от уровней озера и сбросов с Иркутской ГЭС».
СПИСОК ЛИТЕРАТУРЫ
Гармаев Е.Ж., Цыдыпов Б.З. 2019. Уровенный режим оз. Байкал: состояние и перспективы в новых услови-
ях регламентации. Вестник Бурятского государственного университета. Биология, география 1: 37-44.
[Garmaev E.Zh., Tsydypov B.Z. 2019. Level regime of the Lake Baikal: state and prospects in the new
conditions of regulation. Vestnik of Buryat State University. Biology, Geography 1: 37-44. (in Russian)].
Гуркина Р.А. 1983. Сезонные изменения паразитофауны плотвы озера Врево. Сб. науч тр. ГосНИОРХ
197: 85-99. [Gurkina R.A. 1983. Seasonal fluctuations of roach parasite fauna of Lake Vrevo. Sbornik
nauchnykh trudov GosNIORKH 197: 85-99. (in Russian)].
474
Дугаров Ж.Н., Пронин Н.М., Сондуева Л.Д., Бурдуковская Т.Г., Батуева М.Д., Пронина С.В. 2011. Зависимость
структуры сообществ паразитов плотвы Rutilus rutilus (L.) от возраста хозяина. Биология внутренних вод
1: 86-97. [Dugarov Zh.N., Pronin N.M., Sondueva L.D., Burdukovskaya T.G., Batueva M.D., Pronina S.V. 2011.
Dependence that the community structure of the roach Rutilus rutilus (L.) Parasites Has on the Host Age.
Biology of Inner Water 1: 86-97. (in Russian)].
Картушин А.И. 1958. Биология сибирской плотвы, ельца, язя и карася в системе озера Байкал. В кн. Рыбы
и рыбное хозяйство в бассейне озера Байкал. Под ред. М.М. Кожова, К.И. Мишарина. Иркутск,
130-241. [Kartushin A.I. 1958. Biology of Siberian roach, dace, ide and gibel carp in the systems of Lake
Baikal. In: Fishes and fisheries in the Baikal basin. Eds Kozhov M.M., Misharin K.I. Irkutsk: 130-241.
(in Russian)].
Кашковский В.В. 1967. Сезонные изменения паразитофауны плотвы Rutilus rutilus (L.) Ириклинского во-
дохранилища. Вопросы ихтиологии 2: 378-386. [Kashkovsky V.V. 1967. Seasonal changes of parasite
fauna of roach Rutilus rutilus (L.) of Iriklinsky reservoir. Voprosy ikhtiology 2: 378-386. (in Russian)].
Никитин В.М. 2021. Влияние изменения уровня воды в озере Байкал на состояние экосистемы озера,
определение ущерба объектам экономики и инфраструктуры прибрежной территории Республики
Бурятия, Иркутской области в зависимости от уровней озера и сбросов Иркутской ГЭС. Итоги
Круглого стола «Актуальные вопросы регулирования уровня оз. Байкал» на площадке Научного
2021. The impact of changes in the water level in Lake Baikal on the state of the ecosystem of the lake,
determining the damage to economic facilities and infrastructure of the coastal territory of the Republic of
Buryatia, the Irkutsk region, depending on the levels of the lake and discharges from the Irkutsk HPP. The
results of the round table «Actual issues of regulation of the lake level. Baikal» at the site of the Scientific
Шульман С.С. (отв. ред.) 1984. Определитель паразитов пресноводных рыб фауны СССР. Т. 1. Паразитиче-
ские простейшие. Л., Наука, 431 с. (Определители по фауне СССР) [Shulman S.S. 1984. Class Myxo-
sporidia. Opredelitel Parasitov Presnovodnych Rib Fauny SSSR. Leningrad, Nauka, 431 pp. (in Russian)].
Athanassopoulou F., Sommerville C. 1993a. A comparative study of the myxosporeans Myxidium rhodei Léger,
1905 and Myxidium pfeifferi Auerbach, 1908 in roach, Rutilus rutilus L. Journal of Fish Diseases 16: 27-38.
Athanassopoulou F., Sommerville C. 1993b. The significance of myxosporean infections in roach, Rutilus rutilus
L., in different habitats. Journal of Fish Diseases 16: 39-51.
Batueva M.D., Pronin N.M., Pronina S.V. 2015. Prevalence of Myxidium rhodei (Cnidaria, Myxosporea) in the
Lake Baikal basin. Protistology 9: 67-74.
Batueva M.D., Pan X., Zhang J.Y., Liu X.H., Wei W., Liu Y. 2020. Morphological, histological and molecular
characterization of Myxidium cf. rhodei infecting the kidney of Rutilus rutilus”. Diseases of Aquatic Or-
Brummer-Korvenkontio H., Valtonen E.T., Pugachev O.N. 1991. Myxosporea parasites in roach, Rutilus ruti-
lus (Linnaeus), from four lakes in central Finland. Journal of Fish Biology 38: 573-586.
Bush A.O., Lafferty K.D., Lotz J.M., Shostak A.W. 1997. Parasitology meets ecology on its own terms: Margolis
et al. revisited. Journal of Parasitology 83: 575-583.
Dyková I., Lom J., Crupcheva G. 1987. Pathogenicity and some structural features of Myxidium rhodei (Myxozoa:
Myxosporea) from the kidney of the roach Rutilus rutilus. Diseases of Aquatic Organisms 2: 109-115.
Dzika E., Wlasow T., Hoffmann R.W. 2006. Myxidium rhodei Léger, 1905 (Myxozoa: Myxosporea) infection in
roach from four lakes of northern Poland. Bulletin of European Association of Fish Pathology 26: 119-124.
Eiras J.C., Saraiva A., Cruz C.F., Santos M.J., Fiala I. 2011. Synopsis of the species of Myxidi-
um Bütschli,1882 (Myxozoa: Myxosporea: Bivalvulida). Systematic Parasitology 80: 81-116.
El-Matbouli M., McDowell T.S., Antonio D.B., Andree K.B., Hedrick R.P. 1999. Effect of water temperature on
the development, release and survival of the triactinomyxon stage of Myxobolus cerebralis in its oligochaete
host. International Journal for Parasitology 29: 627-641. https:// doi: 10.1016/s0020-7519(99)00009-0
Espinoza L.L., Mertins O., Gama G.S., Fernandes Patta A.C.M., Mathews P.D. 2017. A new Myxidium species
(Myxozoa: Myxosporea) infecting the gallbladder of the turtle Podocnemis unifilis (Testudines: Podocnemi-
didae) from Peruvian Amazon. Acta Tropica 172: 75-79. https://doi: 10.1016/j.actatropica. 2017.04.021.
475
Fariya N., Kaur H., Abidi R. 2020. Myxidium tictoi n. sp., a myxozoan parasite infecting kidney of fresh water
barb Puntius ticto (Hamilton, 1822) from river Gomti, Lucknow (U.P). Journal of Parasitic Diseases 44:
Hallett S.L., Bartolomew J.L. 2007. Effects of water flow on the infection dynamics of Myxobolus cerebralis.
Hile R. 1936. Age Determination of Fish from Scales; Method and Application to Fish Cultural Problems. Pro-
gressive Fish-Culturist 3: 1-5.
Kepr T. 1987. Myxidium rhodei Léger, 1905 (Protozoa, Myxosporea) in the muscle and liver-tissue of the Roach,
Rutilus rutilus (Linne). Folia Parasitologica 34: 181-181.
Kepr T. 1991. Parasitic protozoa of cyprinid fishes: protozoa of the roach Rutilus rutilus (Linnaeus, 1758) in
Czechoslovakia. Folia Parasitologica 38: 11-21.
Léger L. 1930. Myxosporidies nouvelles ou peu connues du genre ‘‘Myxidium’’ chez les poissons d’eau douce.
Travaux du Laboratoire d’Hydrobiologie et Pisciculture de l’Universite´ de Grenoble 22: 231-243.
Longshaw M., Frear P.A., Feist S.W. 2005. Descriptions, development and pathogenicity of myxozoan (Myxozoa:
Myxosporea) parasites of juvenile cyprinids (Pisces: Cyprinidae). Journal of Fish Diseases 28: 489-508.
Marcogliese D.J., Cone D.K. 2021. Myxozoan Communities in Two Cyprinid Fishes from Mesotrophic and
Marcogliese D.J., Gendron A.D., Cone D.K. 2009. Impact of municipal effluents and hydrological regime
on myxozoan parasite communities of fish. International Journal for Parasitology 39: 1345-1351.
https:// doi: 10.1016/j.ijpara.2009.04.007
Okamura B., Hartigan A., Naldoni J. 2018. Extensive uncharted biodiversity: the parasite dimension. Integrative
and Comparative Biology 58: 1132-1145. https: //doi.org/10.1093/icb/icy039
Oliva M., Luque J.L., Teran L., Llican L. 1992. Kudoa sciaenae (Myxozoa: Multivalvulidae) cysts distribution
in the somatic muscules of Stellifer minor (Tschudi, 1844) (Pisces: Scienidae). Memórias do Instituto
Oswaldo Cruz 87 (1): 33-35.
Ware S., Roumillat W.A., Connors V.A., Buron I. 2014. Distribution of Kudoa inornata plasmodia in the mus-
culature of its host, the spotted seatrout Cynoscion nebulosus. Comparative Parasitology 81 (1): 10-14.
Yokoyama H., Ogawa K., Wakabayashi H. 1993. Some biological characteristics of actinosporeans from the oli-
gochaete Branchiura sowerbyi. Diseases Aquatic Organisms 17: 223-228. https:// doi:10.3354/dao017223
Myxidium rhodei Leger, 1905 (Cnidaria, Myxosporea):
age-related, seasonal, spatial dynamics
and long-term survey of common roach Rutilus rutilus
infection in Chivirkuy Bay Lake Baikal
M. D. Batueva*, Zh. N. Dugarov, L. D. Sondueva
Keywords: Lake Baikal, common roach, myxosporean, kidney, pseudocysts, Myxidium
rhodei
SUMMARY
The myxosporean Myxidium rhodei Leger, 1905 is a common parasite of Palearctic cyprinids.
The parasite occurs predominantly in Bowman’s capsules of the kidney of the common roach Rutilus
rutilus. In 1999-2021, these fish were infested with M. rhodei at a high prevalence (94-100%) and
intensity of invasion (up to 1578 pseudocysts). In this work, we analyzed the age-related, seasonal,
long-term dynamics, and spatial distribution of the roach M. rhodei infestation in the Chivyrkuisky
Bay of Lake Baikal. The parasite is primarily recorded in yearlings, then the intensity of M. rhodei
roach invasion increases, reaching the maximum in middle-aged fish, and decreasing in older ones.
M. rhodei has an annual developmental cycle in roach. Fluctuations in M. rhodei infestation of the
studied fish during 25 years of observations correlate with long-term changes in the hydrological
regime of Lake Baikal.
476