

Лесоведение, 2023, № 3, стр. 269-276
Влияние колоний серой цапли на радиальный прирост деревьев в культурах сосны
Ю. П. Демаков a, *, Д. В. Тишин b, И. П. Демитрова a
a Поволжский государственный технологический университет
424000 Йошкар-Ола, пл. Ленина, д. 3, Россия
b Казанский (Приволжский) федеральный университет
420008 Казань, ул. Кремлевская, д. 18, Россия
* E-mail: Demakov@yandex.ru
Поступила в редакцию 27.09.2021
После доработки 15.03.2022
Принята к публикации 07.06.2022
- EDN: PSWKUO
- DOI: 10.31857/S0024114823030038
Аннотация
Приведены данные по влиянию колоний серой цапли (Ardea cinerea L., 1758) на динамику параметров годичного кольца деревьев в сосняках искусственного происхождения. Показано, что начало колонизации насаждений серой цаплей можно довольно точно установить при анализе динамики параметров годичных колец деревьев, особенно ширины их позднего слоя и оптической плотности древесины (значения первого из этих параметров в контрольном насаждении ниже, а второго выше). Колонии серой цапли начинают появляться в сосняках уже с 10–12-летнего возраста, когда высота деревьев достигает 5–6 м, а диаметр ствола на высоте 1.3 м от поверхности почвы – всего 7–8 см. В первые 15–20 лет они оказывают положительное влияние на годичный прирост деревьев, особенно на ширину позднего слоя древесины и толщину клеточных стенок, но затем картина меняется на противоположную. Общее время пребывания колоний цапли на одном месте составляет около 35–40 лет, после чего они переселяются на новые места в связи с расстройством или же полной гибелью насаждений. Для снижения вероятности появления колоний серой цапли, которые во многих случаях оказывают отрицательное влияние на состояние лесных биогеоценозов, необходимо отказаться от создания вблизи водоемов чистых сосняков, отдавая предпочтение еловым, березовым, тополевым или же липово-дубовым насаждениям.
Актуальность исследования обусловлена необходимостью совершенствования технологий и методов управления качеством окружающей среды и развитием лесных биогеоценозов, в функционировании которых большую роль играют позвоночные животные, в том числе и птицы, оказывающие как положительное, так и негативное влияние на их состояние (Рахилин, 1970; Иноземцев, 1978; Кулаков, Крылов, 2018). Средообразующая деятельность птиц наиболее сильно проявляется в местах их массовых скоплений (Ардамацкая, 1967; Бреслина, Карпович, 1969; Семаго, 1975; Тараненко, 1975; Захаренко, Романов, 2009; Лысенков, 2016). Яркий пример – крупные гнездовые поселения серой цапли, где птицы привносят значительные массы органического вещества при строительстве гнёзд и особенно при выкармливании птенцов, оказывая очень большое влияние на организацию и состояние заселенных ими биогеоценозов (Чугай, 1993; Недосекин, 2001, 2003), выступая в качестве своеобразных экосистемных “инженеров” (Jones et al., 1994; Wright, Jones, 2004). Исследователями показано, что колонии цапли приводят к формированию мощной лесной подстилки, затрудняющей прорастание семян и препятствующей процессу естественного лесовозобновления, изменяют структуру и химизм почв, а также населяющих их сообществ беспозвоночных. В итоге все это вызывает обеднение исходного состава фитоценозов и снижение проективного покрытия растениями вплоть до полного их исчезновения непосредственно под гнездами птиц. Длительное гнездование цапель на одних и тех же деревьях приводит к их усыханию, что вынуждает птиц перемещаться на новые участки. Воздействие цапель на биогеоценозы зависит от плотности населения птиц и длительности существования колоний, а также местных особенностей экотопов, что должно быть предметом детального изучения, актуальность которого в современных условиях, когда человек активно преобразовывает ландшафты, вполне очевидна. Цель исследования заключается в выявлении особенностей динамики радиального прироста деревьев в местах гнездования серой цапли, образовавшихся в искусственно созданных сосновых насаждениях Марийского ополья.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проведены в чистых сосновых насаждениях, созданных в 1973 г. на приовражных землях в бассейне реки Манаги, находящихся в центральной части Республики Марий Эл (рис. 1). Почва на опытном объекте, состоящем из нескольких участков, расположенных на разных берегах пруда и различающихся по составу древостоев, дерново-подзолистая суглинистая (Демаков и др., 2020). Поселение цапель, состоящее в настоящее время из 16-19 гнездящихся пар птиц, находится на правом берегу пруда в культурах сосны, примыкающих к пашне (координаты 56.748828° с.ш., 48.250289° в.д.). Текущая густота древостоя на участке составляет 1360 экз. га–1, полнота – 1.18, средняя высота – 24 м, а средний диаметр – 21 см (Демаков и др., 2017). В качестве контроля выбрано аналогичное по параметрам сосновое насаждение, расположенное на противоположной стороне пруда, где поселения цапли отсутствуют.

На каждом из участков с помощью бурава Пресслера у 12 деревьев, диаметр которых в коре изменялся от 14 до 24 см, отбирали на высоте 1.3 м от поверхности почвы керны древесины и помещали их в специальные пеналы для сохранности при транспортировке в лабораторию. Измерение ширины годичных колец проведено в лаборатории дендрохронологии Казанского федерального университета на полуавтоматической установке LINTAB-6 с точностью 0.01 мм стандартными методами дендрохронологии (Шиятов, 1973; Битвинскас, 1974; Ваганов и др., 1996). В дополнении к традиционным показателям годичного слоя деревьев было проведено измерение его оптической плотности (blue intensity, BI), величина которой зависит, как показано исследователями (McCarroll et al., 2002; Campbell et al., 2011; Rydval et al., 2014; Björklund et al., 2014, 2015; Долгова, 2017; Fuentes et al., 2017; Овчинников и др., 2018), от толщины клеточных стенок колец и количества в них лигнина, отрицательно коррелируя с физической (реальной) плотностью древесины. Подготовка кернов к измерениям заключалась в экстракции смол 99%-ным этанолом с помощью аппарата Сокслета, где их выдерживали в течение примерно 72 ч. Затем керны высушивали до воздушно-сухого состояния (влажность 9–12%), наклеивали на специальную подложку, аккуратно подрезали на микротоме и шлифовали мелкой наждачной бумагой с размером зерна менее 1000 мкм. Далее проводили сканирование годичных колец с разрешением 2400 точек на дюйм на профессиональном планшетном сканере Epson. Измерения оптической плотности древесины проведены с помощью специализированных программ CooRecorder® и CDendro® (http:// www.cybis.se/forfun/dendro/). Цифровой эмпирический материал обработан с использованием стандартных методов математической статистики (Лакин, 1990) и пакетов соответствующих прикладных программ.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Поселения серой цапли, которые птицы устраивали только на деревьях сосны, хотя на участке произрастает довольно много более крупных деревьев березы с раскидистыми кронами, а также менее крупных деревьев ели, появились на территории опытного объекта в 1985 году, что удалось установить довольно четко методом дендрохронологии. Возраст насаждений составлял в то время 12 лет, а высота деревьев достигала 5–6 м. Основной причиной образования колонии цапель явилось создание пруда, примыкающего к насаждению. Величина радиального годичного прироста деревьев на участке с колонией птиц начала с этого момента превышать аналогичные показатели контрольного насаждения (рис. 2 и 3, табл. 1). Наиболее значительные различия отмечались по значениям оптической плотности древесины (рис. 4), свидетельствующие о том, что изменения происходили в основном за счет стенок клеток годичных колец, которые в контрольном насаждении тоньше и содержат меньшее количество лигнина. По толщине слоя ранней древесины наибольшие различия между экотопами отмечались в 2004 г., а поздней древесины – в 2005. После этого различия стали уменьшаться, постепенно полностью исчезая. Различия экотопов по значениям остальных параметров годичного слоя древесины проявлялись не столь четко, а по значениям же доли поздней древесины они практически отсутствовали.
Рис. 2.
Динамика ширины раннего и позднего слоев годичных колец деревьев в заселенном серой цаплей (№ 1) и контрольном (№ 2) сосновых насаждениях.
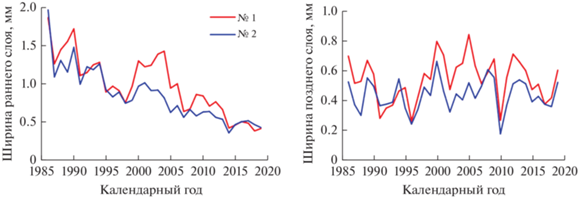
Рис. 3.
Динамика индексов общего радиального годичного прироста деревьев в заселенном серой цаплей (№ 1) и контрольном (№ 2) сосновых насаждениях.
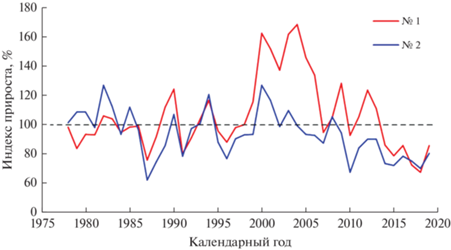
Таблица 1.
Разность параметров прироста деревьев сосны на объектах исследования с 1986 по 2015 гг.
| Оцениваемый параметр | Значения статистических показателей* | ||||
|---|---|---|---|---|---|
| M ± m | min | max | Sx | СV, % | |
| Общий годичный слой древесины, мм | 0.33 ± 0.05 | –0.01 | 0.85 | 0.23 | 68.6 |
| Индекс годичного прироста, % | 26.7 ± 4.1 | –0.7 | 69.0 | 18.5 | 69.1 |
| Слой ранней древесины, мм | 0.20 ± 0.04 | 0.00 | 0.61 | 0.16 | 82.9 |
| Слой поздней древесины, мм | 0.13 ± 0.02 | –0.02 | 0.32 | 0.08 | 63.4 |
| Доля поздней древесины, % | 0.92 ± 0.68 | –4.82 | 7.26 | 3.04 | 331.9 |
| Оптическая плотность, отн. ед. | 0.16 ± 0.01 | 0.11 | 0.20 | 0.03 | 15.7 |
Рис. 4.
Динамика оптической плотности годичного слоя деревьев в заселенном серой цаплей (№ 1) и контрольном (№ 2) сосновых насаждениях.
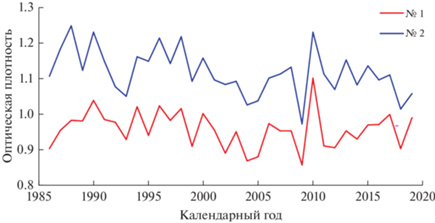
Изменения значений каждого из параметров годичного слоя происходили в экотопах довольно синхронно (r = 0.758–0.939), что свидетельствует об однотипности древостоев, одинаково реагирующих на флуктуации метеорологических условий, являющихся в данном случае ведущим фактором их вариабельности. Менее сходными между собой оказались ряды абсолютной ширины позднего слоя древесины (r = 0.758), а наиболее же сопряженными – ряды значений его доли в годичном кольце (r = 0.939). Коэффициент корреляции между рядами значений оптической плотности древесины годичных колец составил 0.779. На засуху 2010 года древостои отреагировали одинаково, резко снизив прирост позднего слоя годичного кольца и толщину клеточных стенок, что привело к увеличению значений оптической плотности древесины. Изменения же значений разных параметров годичного слоя происходили в экотопах несинхронно (табл. 2), что свидетельствует об их разном информативном значении. Наиболее сходен ход значений общей ширины годичного кольца и слоя ранней древесины, динамика же значений оптической плотности древесины наименее сопряжена с динамикой значений всех остальных параметров годичного кольца (r = –0.47–0.32).
Таблица 2.
Корреляционная связь между разными параметрами прироста деревьев сосны на объектах исследования с 1986 по 2019 гг.
| Оцениваемый параметр | Значение коэффициента корреляции между параметрами | ||||
|---|---|---|---|---|---|
| № 1 | № 2 | № 3 | № 4 | № 5 | |
| Участок с поселениями серой цапли | |||||
| 1. Общий годичный слой древесины | 1.00 | ||||
| 2. Индекс годичного прироста | 0.63 | 1.00 | |||
| 3. Слой ранней древесины | 0.95 | 0.48 | 1.00 | ||
| 4. Слой поздней древесины | 0.49 | 0.64 | 0.19 | 1.00 | |
| 5. Доля поздней древесины | –0.55 | –0.13 | –0.77 | 0.43 | 1.00 |
| 6. Оптическая плотность | –0.13 | –0.29 | 0.02 | –0.45 | –0.30 |
| Участок без поселений серой цапли | |||||
| 1. Общий годичный слой древесины | 1.00 | ||||
| 2. Индекс годичного прироста | 0.53 | 1.00 | |||
| 3. Слой ранней древесины | 0.96 | 0.39 | 1.00 | ||
| 4. Слой поздней древесины | 0.30 | 0.57 | 0.02 | 1.00 | |
| 5. Доля поздней древесины | –0.57 | –0.08 | –0.77 | 0.57 | 1.00 |
| 6. Оптическая плотность | 0.23 | –0.12 | 0.32 | –0.27 | –0.47 |
ВЫВОДЫ
1. Колонии серой цапли оказывают очень большое влияние на организацию и состояние заселенных ими лесных биогеоценозов, в том числе на динамику параметров структуры годичного кольца деревьев, постепенно приводя их к гибели, что вынуждает птиц перемещаться в новые места обитания.
2. Начало колонизации экотопов серой цаплей можно довольно точно установить при анализе динамики параметров структуры годичных колец деревьев, особенно ширины их позднего слоя и оптической плотности древесины. Значения первого из этих параметров в насаждениях, подвергшихся воздействию колоний цапли, выше по сравнению с контролем, а второго ниже.
3. Колонии серой цапли начинают появляться в 10–12-летних сосновых насаждениях, когда высота деревьев достигает 5–6 м, а диаметр ствола на высоте 1.3 м от поверхности почвы – всего 7–8 см.
4. В первые 15–20 лет колонии серой цапли оказывают положительное влияние на годичный прирост деревьев, особенно на ширину позднего слоя древесины и толщину клеточных стенок, но затем картина меняется на противоположную. Общее время пребывания колоний цапли на одном месте составляет около 35–40 лет.
5. Для снижения вероятности возникновения колоний серой цапли, которые во многих случаях оказывают отрицательное влияние на состояние лесных биогеоценозов, необходимо отказаться от создания вблизи водоемов чистых сосновых древостоев, отдавая предпочтение еловым, березовым, тополевым или липово-дубовым насаждениям.
Список литературы
Ардамацкая Т.Б. Влияние массовых колоний птиц на растительность и животное население о. Орлова // Структура и функционально-биогеоценотическая роль животного населения суши. М.: Наука, 1967. С. 113–114.
Битвинскас Т.Т. Дендроклиматические исследования. Л.: Гидрометеоиздат, 1974. 172 с.
Бреслина И.П., Карпович В.Н. Развитие растительности под влиянием жизнедеятельности колониальных птиц // Ботанический журн. 1969. № 5. С. 690–696.
Ваганов Е.А., Шиятов С.Г., Мазепа В.С. Дендроклиматические исследования в Урало-Сибирской Субарктике. Новосибирск: Наука, 1996. 246 с.
Демаков Ю.П., Нуреев Н.Б., Митякова И.И. Влияние характера хозяйственного использования приовражных земель на свойства почв // Вестник Поволжского государственного технологического университета. Сер.: Лес. Экология. Природопользование. 2020. № 4(48). С. 77–95.
Демаков Ю.П., Нуреева Т.В., Краснов В.Г., Рыжков А.А. Эколого-ресурсный потенциал лесных насаждений на приовражно-балочных землях Среднего Поволжья // Вестник Поволжского государственного технологического университета. Сер.: Лес. Экология. Природопользование. 2017. № 3(35). С. 73–87.
Долгова Е.А. Связь с засушливостью ширины колец ранней и поздней древесины и оптической плотности колец сосны (на примере Калужской области) // Засухи Восточно-Европейской равнины по гидрометеорологическим и дендрохронологическим данным. Санкт-Петербург: Нестор-История, 2017. С. 208–222.
Захаренко К.А., Романов В.В. О влиянии колониального поселения озерных чаек на особенности химического состава почв в условиях Владимирского ополья // Вестник Оренбургского государственного университета. 2009. № 6(110). С. 147–152.
Иноземцев А.А. Роль насекомоядных птиц в лесных биогеоценозах. Л.: ЛГУ, 1978. 264 с.
Кулаков Д.В., Крылов А.В. Влияние птиц на среду обитания // Природа. 2018. № 5(1233). С. 22–31.
Лакин Г.Ф. Биометрия. М.: Высшая школа, 1990. 352 с.
Лысенков Е.В. Средообразующая роль врановых в антропогенных ландшафтах // Русский орнитологический журн. 2016. Т. 25. № 1371. С. 4643–4647.
Недосекин A.A. Изменение химического состава почвы под влиянием колонии серых цапель // Актуальные проблемы экологии и природопользования. М.: РУДН, 2003. Вып. 3. С. 90–93.
Недосекин A.A. Изменения в распределении растительного покрова под гнездами в колонии серых цапель в Тульских засеках // Актуальные проблемы изучения и охраны птиц Восточной Европы и Северной Азии. Казань: Изд-во Казанского университета, 2001. С. 467–468.
Овчинников Д.В., Ерёмина А.Д., Овчинников С.Д., Кладько Ю.В. Потенциал оптической плотности древесины в дендроклиматологии // География и геоэкология на службе науки и инновационного образования. Красноярск: КГУ, 2018. Вып. 13. С. 68–70.
Рахилин В.К. О средообразующей роли птиц фауны СССР // Средообразующая деятельность животных. М.: Наука, 1970. С. 15–18.
Семаго Л.Л. К вопросу о средообразующей деятельности колониальных и стайных птиц // Проблемы изучения и охраны ландшафтов. Воронеж: Воронежское кн. изд-во, 1975. С. 45–47.
Тараненко Л.И. Влияние колониального гнездования грачей на окружающую среду // Роль животных в функционировании экосистем. М.: Наука, 1975. С. 104–106.
Чугай С. Роль колоний серой цапли в функционировании экосистем пойменных черноольшанников // Птицы бассейна Северского Донца. Донецк: ДГУ, 1993. С. 50–52.
Шиятов С.Г. Дендрохронология, её принципы и методы // Записки Всесоюзного ботанического общества. Вып. 6. Проблемы ботаники на Урале. Свердловск: УНЦ АН СССР, 1973. С. 53–81.
Björklund J.A., Gunnarson B.E., Seftigen K., Esper J., Linderholm H.W. Blue intensity and density from northern Fennoscandian tree rings, exploring the potential to improve summer temperature reconstructions with earlywood information // Climate of the Past. 2014. V. 10. P. 877–885.
Björklund J.A., Gunnarson B.E, Seftigen K., Zhang P., Fuentes M. Using adjusted Blue Intensity data to attain high quality summer temperature information: A case study from Central Scandinavia // The Holocene. 2015. V. 25. № 3. P. 547–556.
Campbell R., McCarroll D., Robertson I., Loader N.J., Grudd H., Gunnarson B.E. Blue intensity in Pinus sylvestris tree rings: A manual for a new palaeoclimate proxy // Tree-ring Research. 2011. V. 67. P. 127–134.
Fuentes M., Salo R., Björklund J., Seftigen K., Zhang P., Gunnarson B. et al. 970-year-long summer temperature reconstruction from Rogen, west-central Sweden, based on blue intensity from tree rings // The Holocene. 2017. V. 28. № 2. P. 254–266.
Jones C.G., Lawton J.H., Shachak M. Organisms as ecosystem engineers // Oikos. 1994. V. 69. P. 373–386.
McCarroll D., Pettigrew E., Luckman A., Guibal F., Edouard J.-L. Blue reflectance provides a surrogate for latewood density of high-latitude pine tree rings // Arctic, Antarctic and Alpine Research. 2002. V. 34. № 4. P. 450–453.
Rydval M., Larsson L.A., McGlynn L., Gunnarson B.E., Loader N.J., Young G.H., Wilson R. Blue intensity for dendroclimatology: should we have the blues? Experiments from Scotland // Dendrochronologia. 2014. V. 32. № 3. P. 191–204.
Wright J.P., Jones C.G. Predicting effects of ecosystem engineers on patch-scale species richness from primary productivity // Ecology. 2004. V. 85. № 8. P. 2071–2081.
Дополнительные материалы отсутствуют.