

Лесоведение, 2023, № 3, стр. 244-254
Генеративная сфера сосны обыкновенной как индикатор климатически детерминированной смены жизненных состояний популяций
Н. Ф. Кузнецова *
Всероссийский научно-исследовательский институт лесной генетики, селекции и биотехнологии
394087 Воронеж, ул. Ломоносова, д. 105, Россия
* E-mail: nfsenyuk@mail.ru
Поступила в редакцию 03.02.2022
После доработки 25.03.2022
Принята к публикации 07.06.2022
- EDN: PTGBOV
- DOI: 10.31857/S0024114823030063
Аннотация
Обсуждаются проблемы перехода сосны обыкновенной (Pinus sylvestris L.) из состояния устойчивого к неустойчивому равновесию, смены равновесной популяции на неравновесную в результате потепления климата. Цель исследований – изучить состояние генеративной сферы южной и центральной лесостепных популяций сосны обыкновенной; по признаку полнозернистости оценить их жизненное состояние в 2020 г. по сравнению с 2016 – годом дестабилизации сосновых лесов, а также относительно состояния степной популяции в урожайном 2017 г. Объектами исследования служили центральная и южная лесостепные популяции сосны, произрастающие на экологически благоприятной территории Воронежской и Белгородской областей. В 2015 г. зафиксирован переход их из равновесия в слабо неравновесную систему. Уровень полнозернистости ступинской и белгородской популяций снизился соответственно на 25.6 и 24.6%, число семян – на 37.7 и 22.4%, уровень смертности семяпочек повысился в 3.9 и 4.0 раза. Сравнение показателей семенной продуктивности в 2013, 2016 и 2020 гг. показало, что в настоящее время оба насаждения представляют равновесные системы. Центральная популяция вернулась к региональной норме (устойчивого равновесия), южная находится в состоянии неустойчивого равновесия. Дисперсионный анализ выявил существенное (57.8%) влияние фактора “ГТК” на генеративную сферу белгородской популяции по признаку полнозернистости. В последнее десятилетие число оптимальных лет в ЦЧР уменьшилось с 7–8/10 лет до 5. Для возвращения сосновых лесов в равновесие требуется 3 оптимальных года. Высокие темпы потепления климата могут повлечь за собой нарушение равновесия и повторную дестабилизацию. Дальнейшее сокращение числа оптимальных лет делает почти невозможным возвращение сосновых лесов к региональной норме.
Изменение климата не раз происходило в истории Земли. В эти периоды ускорялось движение видовых ареалов, одни виды-эдификаторы сменялись на другие. Сегодня мир вновь вступил в эпоху климатической нестабильности. В разных странах изменение климата происходит неравномерно (Dias et al., 2015; IPCC, 2017). В России повышение среднегодовой температуры составляет 0.47°С/10 лет, что в 2.5 раза выше роста глобальной температуры (Доклад …, 2021). Биология видов перестает соответствовать среде обитания, что является причиной ослабления лесов, перехода в более низкое жизненное состояние и даже гибели (Allen et al., 2010; Касимов и др., 2011; Alberto et al., 2013; Gauthier et al., 2015; Kuznetsova et al., 2019).
Изменение климата Среднерусской лесостепи началось во второй половине ХХ в. За первые 25 лет летняя температура повысилась на 0.6°С, к концу века – на 1.9°С (Акимов, Задорожная, 2018). В ХХI веке темпы потепления возросли, что подтверждается ростом активных температур (Виноградова, 2014; Виноградова и др., 2015). С 2000 по 2012 гг. их уровень достиг 2600–3000°С, что выше многолетних показателей на 250–350°С. Установлено, что нарушение термического режима связано с блокированием переноса западных воздушных масс, перемещением жаркого тропического воздуха и длительным его стационированием на юге Европейской России (Акимов, Задорожная, 2018). Индекс SCEI снизился с –1 до –1.5, ГТК – с 1.0 до 0.5–0.6 (Виноградова, 2014; Виноградова и др., 2015). Увлажненные территории стали превышать засушливые всего на 10–15%. В 2007–2014 гг. в Центрально-Черноземном районе произошла мощная тепловая волна, которая включала 4 сильные засухи (2007, 2010, 2012, 2014 гг.) (Кузнецова и др., 2021). Началась “раскачка” сосновых лесов, которая вызвала их дестабилизацию.
В данном исследовании мы использовали две концепции, согласно которым лесные экосистемы на территории ареала могут находиться в равновесном и неравновесном состоянии (Крамер, Ван дер Верф 2010; Iglesias, Whitlock, 2020). Каждое из них, в свою очередь, подразделяется на две формы – устойчивую и менее устойчивую. Сосновые леса как сложные динамичные системы представляют в порядке убывания следующий ряд: устойчиво равновесные, неустойчиво равновесные, слабо неравновесные, неравновесные, мертвые. Дифференциация, по-видимому, является следствием падения энергетического статуса: максимальный – у равновесных популяций, более низкий – у неравновесных, нулевой – у мертвых. Преимущество данного системного подхода заключается в том, что он дает возможность анализировать динамику изменчивости оптимально структурированных равновесных популяций, которые под воздействием климатического стресса могут переходить из одной формы в другую, а при достижении предельно допустимого порога скачком спускаться в более низкое жизненное состояние.
В центре ареала большинство популяций сосны обыкновенной находятся в состоянии устойчивого или неустойчивого равновесия. Общее жизненное состояние (ОЖС) сосновых лесов на индивидуальном и популяционном уровне принято оценивать по комплексу вегетативных и генеративных признаков (Боголюбов и др., 1999). Генеративная сфера сосны более чувствительна к среде обитания, часто используется как биоиндикатор для оценки ее изменения (Alaquori et al., 2020). В оптимальные годы диапазон изменчивости показателей биопродуктивности шишек низкий, отклонения относительно флуктуаций внешней среды обратимы (Кузнецова и др., 2021). Это является подтверждением того, что популяции развиваются в режиме равновесной системы. В данном жизненном состоянии (ЖС) сосновые леса, как упорядоченные системы, характеризуются высоким бонитетом и устойчивостью к региональным стрессорам.
Смена равновесного жизненного состояния на более низкое (слабо неравновесное) существенно изменяет функционирование предшествующих структур и функций, ведет к реорганизации метаболических процессов (Раутиан, 1993). В неравновесных популяциях нормы реакции деревьев перекрываются, что придает устойчивость как самому растению, так и популяции в целом. Поддержание гомеостаза и основных систем жизнеобеспечения растений в условиях, не соответствующих их биологии, невозможно без дополнительных энергетических затрат (Чудинова, Орлова, 2016). Переход лесных экосистем на менее затратный путь развития сопряжен с перестройкой энергетических процессов, что, с одной стороны, обеспечивает выживание растений в изменившихся условиях, а с другой – снижает их жизненный статус. При этом страдает вегетативная и генеративная сфера деревьев. Неравновесные популяции, как менее упорядоченные системы, в большей степени зависят от факторов внешней среды, имеют более низкую продолжительность жизни, бонитет, урожайность, качество семян.
Цель исследований – изучить состояние генеративной сферы южной и центральной лесостепных популяций сосны обыкновенной; по признаку полнозернистости оценить их жизненное состояние в 2021 г. по сравнению с 2015 г. дестабилизации сосновых лесов, а также относительно состояния степной популяции в урожайном 2017 г.
ОБЪЕКТЫ И МЕТОДИКА
Ступинский тест-объект (Воронежская обл., Рамонский р-н, 55–60-летние лесные культуры, контроль) – типичное по вегетативной и семенной продуктивности насаждение сосны в центре лесостепной зоны, объект 34-летнего мониторинга и комплексных эколого-генетических исследований (Kuznetsova, 2012; Клушевская, Кузнецова, 2016). Граничит с Воронежским заповедником, где эмиссионная нагрузка в 3.7 раза ниже, чем в Воронежской области (Куролап и др., 2010). Опытный объект – южная лесостепная популяция сосны (Белгородская обл., Красненский р-н, 30-летние лесные культуры) является объектом мониторинга с 2010 г. Насаждение произрастает на серых лесных почвах широкой приовражной полосы в 50–60 км от пессимальной зоны ареала сосны.
Выборка включает 30 случайно взятых деревьев, объем одной пробы – 10–15 шишек типичной для дерева формы и размера. Интенсивность семеношения определялась по шкале В.Г. Каппера и трем признакам семенной продуктивности – полнозернистости (%), числу семян на шишку (шт.), смертности оплодотворенных семяпочек (%). Эти экологически зависимые признаки имеют разную природу и порог чувствительности к засухе (Kuznetsova, 2012). Признак полнозернистости характеризует выживание зародышей в эмбриогенезе, начинает реагировать на раннюю засуху, вычисляется как отношение количества полных семян к общему их числу (полные + пустые). Мелкие пустые капсулы из анализа исключаются. Второй показатель – число семян на шишку более устойчив к засухе. Его величина зависит от объема фертильной зоны шишки (Свинцова, 2002). Для определения частотного распределения деревьев диапазон изменчивости признаков разбит на 4 класса. Их границы по признаку полнозернистости: I – 0–24.9%; II – 25.0–49.9%; III – 50.0–74.9%; IV – 75% и выше; по числу семян: I – 0–9.9; II – 10.0–19.9; III – 20.0–29.9; IV – 30.0 семян и больше. Смертность оплодотворенных семяпочек определялась как соотношение пустых капсул к числу выполненных семян (Иванов и др., 2015). Размах изменчивости признака оценивался по классам в границах: I – 0–29.9%; II – 30.0–69.9%; III – 70.0–99.9%; IV – 100.0% и выше. Характеристика признаков и показателей ЖС, связанных со сменой ЖС популяций сосны обыкновенной, приведена в предыдущей работе (Кузнецова и др., 2021).
Статистическая обработка данных выполнена с помощью пакета программного обеспечения MS Excel-2010. Для сравнения выборок и оценки силы влияния факторов использовали однофакторный дисперсионный анализ (ANOVA). С помощью двухвыборочного t-критерия с различными дисперсиями осуществляли оценку достоверности между выборками.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Среднерусская лесостепь – зона физико-географического оптимума вида сосны обыкновенной (Обновленский, 1962). Средний уровень ГТК составляет 1.0–1.2. Его величина между лесостепью и степью переходит через единицу. Периодичность засух – 2–3 в десятилетие. Зональная граница сосны на данном участке ареала совпадает с южным пределом зоны видового оптимума. Сосновые леса (естественные, производные) веками на ее территории развивались по принципу взаимодействия “генотип-среда”, и большинство из них до конца ХХ в. находилось в состоянии устойчивого равновесия (Романовский, 2002; Kuznetsova et al., 2019). Сбалансированное развитие (средний бонитет I,7-II,0) обеспечивали благоприятные природно-климатические условия. В оптимальные годы интенсивность семеношения стабильно высокая (около 80%).
Сосновые леса развиваются в режиме равновесной системы в том случае, если они на следующий год после возмущения возвращаются к исходному состоянию и природно-климатические условия экологической ниши соответствуют биологии вида сосны обыкновенной (Кузнецова, 2021). Ступинская и белгородская лесостепные популяции на начальном этапе мониторинга также являлись равновесными системами. Их отличала устойчивость и сбалансированное развитие вегетативной и генеративной сферы большинства деревьев. В этом случае реакция на засуху представляла генетически обусловленный отклик, протекающий на базе неспецифической и специфической реакции (Kuznetsova, 2012). На следующий год их показатели всегда возвращались к региональной норме. Так, после неурожайного засушливого 2012 г. уровень полнозернистости ступинской популяции составил 82.1 ± 2.62%, белгородской – 80.4 ± 2.08% (табл. 1). Показатели смертности оплодотворенных семяпочек имели близкие значения. Число семян на шишку у деревьев белгородской популяции достоверно превышало уровень контрольного объекта.
Таблица 1.
Показатели семенной продуктивности шишек и качества семян ступинской и белгородской популяций сосны обыкновенной
| Год исследований | Полнозернистость, % |
N семян на шишку, шт. | Смертность оплодотворенных семяпочек, % | Лабораторная всхожесть, % |
|---|---|---|---|---|
| Ступинский тест-объект | ||||
| 2013, opt 2016, дестабилизации 2020, opt |
82.1 ± 2.62 61.1 ± 3.40* 80.8 ± 1.34 |
23.6 ± 1.39 14.7 ± 1.21** 28.0 ± 1.36 |
21.4 ± 1.85 83.6 ± 13.17** 25.6 ± 2.49 |
82.1 ± 1.6 70.4 ± 3.3* 86.5 ± 1.2 |
| Белгородская популяция | ||||
| 2013, opt 2016, дестабилизации 2020, opt |
80.4 ± 2.08 60.6 ± 3.64* 61.4 ± 3.02* |
29.9 ± 1.64 23.2 ± 2.70* 32.5 ± 1.83 |
23.6 ± 1.74 94.0 ± 17.24** 76.3 ± 12.23** |
81.8 ± 2.7 64.6 ± 4.6* 79.7 ± 2.4 |
Тепловая волна 2007–2014 гг. вызвала на территории ЦЧР ослабление сосновых лесов и массовое размножение вредителей. В последний год ее протекания в регионе произошли две засухи. Особенно жесткой была поздняя засуха 2014 г., во время которой за три осенних месяца выпало всего 20% от нормы осадков. Деревья ушли в зиму сильно ослабленными. Вероятно, уровень внутренней разбалансировки (дестабилизации) достиг критической точки, или точки невозврата. Деревья потеряли устойчивость, и весной следующего года при выходе из покоя они скачком перешли из основного равновесного состояния в качественно более низкий режим функционирования – слабо неравновесную систему. Подтверждением является то, что все показатели генеративной сферы сосны в оптимальном 2015 г. и в последующие годы были ниже региональной нормы (Кузнецова и др., 2021). В годы дестабилизации у ступинской популяции отмечены следующие изменения: снижение годового прироста терминальных побегов, урожайности деревьев, семенной продуктивности шишек и лабораторной всхожести семян. Обратный процесс – возврат к региональной норме продолжался в течение 3-х лет. В Воронежской области этому способствовали благоприятные погодные условия вегетационных сезонов 2015–2017 гг. В 2018 г. ступинская популяция вновь находится в состоянии равновесия и реагирует на засуху как равновесная система. Что касается Белгородской области, то летом 2016 г. на ее территории произошла слабая засуха, что не позволило опытной популяции вернуться к состоянию устойчивого равновесия.
В оптимальные годы пустосемянность лесостепных популяций составляет около 20% (Свинцова, 2002; Kuznetsova, 2012). Такие количественные показатели шишек, как полнозернистость и число семян на шишку, на популяционном уровне маркируют генетическую структуру равновесных популяций с приблизительно одинаковым для каждого параметра соотношением между классами продуктивности (Kuznetsova, 2012). Несмотря на то, что данные показатели имеют разную природу, они связаны друг с другом обратно пропорциональной зависимостью. Признак полнозернистости более чувствителен к погодному стрессу и начинает реагировать на слабую засуху. Количество семян снижается в сильную засуху. Параметр “смертность оплодотворенных семяпочек” позволяет оценить на индивидуальном и популяционном уровне число погибших семяпочек в засушливые и оптимальные годы и, что более важно, степень разбалансировки равновесных лесных экосистем в результате потепления климата.
Данные изучения семенной продуктивности деревьев опытной и контрольной популяций в три оптимальных для развития генеративной сферы сосны года (2013, 2016 и 2020 гг.) приведены в табл. 1. Популяционный анализ показал, что в урожае второго 2016 года дестабилизации по отношению к 2013 у обоих насаждений сохранялось достоверное снижение всех анализируемых признаков. При этом отметим, что во время дестабилизации особенно высокие отличия имели место по смертности семяпочек: превышение составило 3.9 раз у ступинской и 4.0 раз у белгородской популяции (табл. 1). Данные различия подтверждаются высокой вариабельностью признака. Так, у ступинской популяции коэффициент вариации (CV, %) выборки повысился с 9.2 до 86.3%. Мониторинг систем семенного размножения позволил сделать вывод, что популяции, произрастающие в центре и на юге лесостепной зоны, в течение 3 оптимальных лет (2015–2017 гг.) находились в состоянии слабо неравновесной системы (Кузнецова и др., 2021).
Анализ состояния генеративной сферы популяций в 2020 г. выявил, с одной стороны, существенные отличия по уровню полнозернистости и смертности семяпочек, а с другой – среднее количество семян в шишках белгородской популяции (32.5 ± 1.83 шт.) несколько превысило уровень 2013 г. (табл. 1). Сравнение показателей смертности оплодотворенных семяпочек в урожае 2013 и 2020 гг. показало, что у белгородской популяции между годами сохраняются высокие различия. Его модальное значение превышает уровень 2013 г. в 3.2 раза (CV = 70.3%, диапазон изменчивости – 12.5–280%).
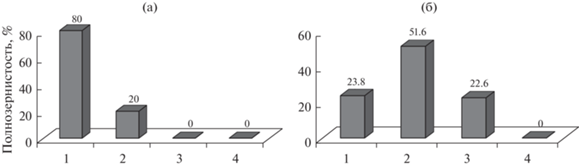
Полученные данные можно объяснить тем, что популяции находятся в равновесном состоянии, но в разных ее формах: ступинская – устойчивого, а белгородская – неустойчивого равновесия. Их различия по ЖС подтверждаются данными частотного распределения деревьев по уровню смертности семяпочек (рис. 1). Выборка ступинской популяции представлена растениями I и II класса (рис. 1а). В III класс входит 1 дерево, а объем I класса больше в 2.75 раза, чем II-го. Белгородская популяция включает деревья всех четырех классов, с модой признака в области II класса (рис. 1б). Принимая во внимание, что уровень смертности семяпочек опытного объекта во время дестабилизации составлял 94.0 ± 17.24%, то текущий уровень ее внутренней разбалансировки (76.3 ± 12.23) близок к критической точке перехода на более низкий режим функционирования – неравновесной системы.
Рис. 1.
Частотное распределение деревьев сосны обыкновенной по классам уровня смертности оплодотворенных семяпочек на ступинской и белгородской популяциях, урожай 2020 г.
После трех оптимальных лет климатическая ситуация в регионе вновь коренным образом изменилась. 2020 и 2021 гг. оказались самыми теплыми за всю историю метеонаблюдений. В 2020 г. среднегодовая температура Воронежской области достигла нового рекорда – 9.8°С. Произошла сильная почвенная засуха, которая продолжалась с середины июля до конца октября. ГТК снизился до 0.7, что соответствует влагообеспеченности центра района степей. Засуха негативно отразилась на состоянии вегетативной сферы сосны. В сосновых лесах наблюдалось массовое пожелтение и опад хвои. При этом погодный стресс не затронул генеративную сферу сосны.
Учитывая все сказанное выше, проанализируем текущее состояние генеративной сферы южной и центральной лесостепных популяций. В Воронежской области термо- и влагообеспеченность первой половины вегетационного сезона 2020 г. были близкими к региональной норме. Наиболее чувствительные стадии репродуктивного цикла сосны обыкновенной приходятся на конец апреля, май и июнь. Это определило высокий уровень интенсивности семеношения ступинской популяции (4 балла). Урожайность и кривая распределения деревьев внутри выборки соответствуют уровню оптимальных лет: полнозернистость семян 80.8 ± 1.34%, при размахе варьирования признака 56.6–90.4% (табл. 2). Диапазон изменчивости признака ниже, чем в предыдущие годы: коэффициент вариации равен 9.2%. Сила влияния ГТК составляет всего 12.5%, что свидетельствует о слабом влиянии данного фактора на конечный результат половой репродукции.
На диаграмме рис. 2 приведены данные частотного распределения деревьев ступинской и белгородской популяций по классам продуктивности признака полнозернистости. У ступинской популяции кривая изменчивости имеет асимметричный, типичный для оптимальных лет тип распределения. Растения низких I–II классов продуктивности отсутствуют (рис. 2а). Выборка представлена двумя продуктивными классами, из них 4/5 деревьев относятся к высокопродуктивному IV классу. Лабораторная всхожесть семян высокая – 86.5 ± 1.24% при размахе варьирования 73.5–98%.
Рис. 2.
Частотное распределение деревьев сосны обыкновенной по классам продуктивности признака полнозернистости на ступинской (а) и белгородской (б) популяциях сосны обыкновенной, урожай 2020 г.
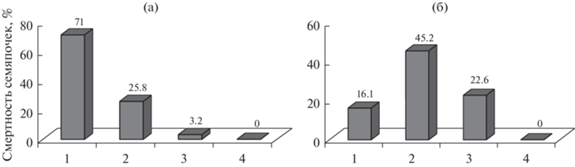
Несколько отличные результаты получены у белгородской популяции. Уровень полнозернистости составил 61.4 ± 3.0%, что достоверно ниже контроля (табл. 2). Коэффициент вариации по сравнению со Ступино в 2.3 раза выше – 21.3%. Диапазон изменчивости реализованной нормы реакции увеличился до 62.6% за счет смещения показателей деревьев в область нижних значений: 88.9–26.3%. Половина деревьев выборки относится к III классу (рис. 2б). Резко снизилась доля растений высокопродуктивного IV класса. Появился низко продуктивный II класс. Его объем сопоставим с количеством высокопродуктивных форм в выборке. Растения с уровнем полнозернистости ниже 25% отсутствуют. Согласно данным дисперсионного анализа, доля изменчивости, обусловленная фактором “ГТК”, составляет 57.8% от общей изменчивости, и он, как основной стрессор переходной зоны лесостепи, существенно влияет на снижение урожайности белгородской популяции.
Дополнительно проведен дисперсионный анализ изучения влияния фактора “жизненное состояние популяции” (ЖС) на изменчивость признака полнозернистости у трех объектов сосны в разные годы: центрального лесостепного (2016 – год дестабилизации, 2020 – оптимальный год), южного лесостепного (2016 – год дестабилизации), кантемировского степного (2017 – оптимальный год) – по отношению к параметру белгородского насаждения (2020 г.). Как сказано выше, ступинская и белгородская популяции перешли в слабо неравновесное состояние в 2015 г. Степная популяция (Воронежская область, Кантемировский район, 35-летние лесные культуры) произрастает в районе степей на экологически благоприятной территории (Сердюкова, 2020). Это пессимальная зона ареала сосны обыкновенной, где природно-климатические условия соответствуют биологии вида лишь частично. Насаждение представляет слабо неравновесную систему. В оптимальном 2017 году оно характеризовалось пониженной семенной продуктивностью: полнозернистость –71.2 ± 3.48%, число семян на шишку – 18.5 ± 1.03 шт. (Сердюкова, 2020; Degtyareva, 2021). Показатели коэффициентов вариации, полнозернистости и результатов однофакторного дисперсионного анализа их повариантного сравнения приведены в табл. 3.
Таблица 3.
Влияние жизненного состояния (ЖС) популяции на признак полнозернистости белгородского объекта сосны обыкновенной в 2020 г.
| Популяция | Год | Тип ЖС | Полнозернис- тость, % |
CV, % | Сила влияния фактора, % |
|---|---|---|---|---|---|
| Белгородская | 2020 | Неустойчиво равновесная | 61.4 ± 3.0 | 2.7 | – |
| Белгородская | 2016 | Слабо неравновесная | 60.6 ± 3.6 | 18.5 | 1.2 |
| Ступинская | 2020 | Устойчиво равновесная | 80.8 ± 1.3* | 9.2 | 54.3 |
| Ступинская | 2016 | Слабо неравновесная | 61.1 ± 3.4 | 3.0 | 1.1 |
| Кантемировская | 2017 | Слабо неравновесная | 71.2 ± 3.5 | 26.6 | 7.2 |
Сравнение показателей лесостепной белгородской (2020 г.) со степной кантемировской популяцией (2017 г.) показал, что сила влияния фактора “ЖС” на уровень полнозернистости незначительна и составляет 7.2% (табл. 3). Минимальное влияние фактора выявлено при сопоставлении данных ступинской и белгородской популяций в 2016 г. – 1.1–1.2%. Существенные различия (54.3%) установлены лишь в варианте сравнения контрольного и опытного объектов в 2020 г. Из проведенного анализа следует, что в настоящее время только сосна ступинского тест-объекта представляет устойчивую равновесную систему.
Второй признак – число семян на шишку – характеризуется высокой изменчивостью и зависит от генотипических особенностей материнских растений, индивидуальной продуктивности мегастробилов дерева и величины фертильной зоны шишек. В районе исследований потенциальная продуктивность мегастробилов составляет 14–76 семяпочек (Свинцова, 2002), что существенно выше, чем у сосны обыкновенной лесной зоны (Романовский, 1997). Заморозки, недоопыленность и повреждение насекомыми являются основными причинами их элиминации (Романовский, 1997; Свинцова, 2002; Kuznetsova, 2012). Прогамная фаза репродуктивного цикла достаточно устойчива к флуктуациям внешней и внутренней среды. Гибель семяпочек невысокая – ~5% (Свинцова, 2002), так как погодный стресс в период заложения семенных чешуй и формирования семяпочек, как правило, не превышает верхний порог их устойчивости. У ступинской популяции число семян на шишку в 2020 г. составило 28.0 ± 1.36 шт. (17.3–46.2 шт.) (табл. 1). Это новый максимум, который не укладывается в диапазон изменчивости модальных значений выборки, полученных в ходе мониторинга 22 оптимальных лет (19.6–26.1 шт.). Мода признака находится в области деревьев III класса (20–30 шт.). Группа низко продуктивного II класса представлена 5 растениями. Деревья с числом менее 10 семян отсутствуют. У опытного объекта модальное значение признака еще выше – 32.5 ± 1.83 шт. Различия между ступинской и белгородской популяцией недостоверны. Норма реакции смещена в область высокопродуктивных форм и включает три класса продуктивности. 2/3 деревьев имеют больше 30 семян. Лабораторная всхожесть – 79.9 ± ± 2.83%, при более широком размахе варьирования признака – 42.0–98.0%.
Высокие показатели числа семян и лабораторной всхожести указывают на то, что белгородская популяция, в отличие от степной кантемировской, является равновесной системой. Пониженный уровень полнозернистости свидетельствует о том, что насаждение находится в состоянии неустойчивого равновесия, что, по-видимому, характерно для лесостепных популяций сосны, произрастающих в переходной зоне ареала.
Из приведенного анализа данных следует, что повышенный вдвое уровень пустосемянности, по-видимому, результат действия сил стабилизирующего отбора, который обеспечивает высокое качество семян в условиях более теплого и сухого климата. Стабилизирующий отбор является важной функцией полового воспроизведения (Щербаков, 2005). Согласно литературным данным, линейные размеры зародышей на начальной стадии роста характеризуются высокой изменчивостью на индивидуальном уровне и в пределах кроны деревьев. В ходе формирования семян их изменчивость снижается и достигает минимума на стадии завершения роста (Свинцова, 2002). Климатическая нестабильность усиливает разбалансировку экологически зависимых процессов и количественных признаков в пределах диапазона изменчивости их норм реакции. Повышенная изменчивость уменьшается в ходе стабилизирующего отбора, который ограничивает избыточную изменчивость путем удаления из семенного генофонда генетически нарушенных, инбредных, несовместимых с эндоспермом, отклоняющихся по размерам и форме генотипов. Морфологически данное явление выражается в виде увеличения пустосемянности шишек в урожаях оптимальных лет.
Потепление климата выводит биоклиматическую систему региона из состояния гомеостаза. На территории Среднерусской лесостепи нестабильность со стороны климатической ее составляющей реализуется путем сокращения более чем на треть числа оптимальных лет и увеличения вдвое частоты экстремальных событий (засух, тепловых волн). Это увеличивает вероятность перехода высокопродуктивных сосновых лесов в более низкое жизненное состояние, а шансы для их возврата к региональной норме, наоборот, падают.
ЗАКЛЮЧЕНИЕ
Ступинская и белгородская лесостепные популяции в 2013 г. находились в состоянии устойчивого равновесия. 8-летняя тепловая волна и засуха 2014 г. весной следующего года вызвали их переход на другой режим функционирования – слабо неравновесной системы. В оптимальном 2016 г. состояние дестабилизации сохранялось: уровень полнозернистости относительно 2013 г. снизился соответственно на 25.6 и 24.6%, число семян – на 37.7 и 22.4%, уровень смертности семяпочек повысился в 3.9 и 4.0 раза.
Благоприятные погодные условия Воронежской области в течение трех лет обеспечили возврат центральной ступинской популяции к региональной норме (устойчивого равновесия). Анализируемые признаки семенной продуктивности в урожаях 2013 и 2020 гг. характеризуются небольшой амплитудой изменчивости и близки между собой по количественным параметрам.
Южная белгородская популяция из-за погодного стресса смогла вернуться только в состояние неустойчивого равновесия, в котором находится до настоящего времени. Число семян и их качество высокое. Уровень смертности семяпочек снизился, но он по-прежнему очень высокий (76.3 ± 12.23%). Полнозернистость ниже уровня 2013 г. на 23.6%. Сила влияния фактора “ГТК” составляет 57.8%. Причиной данного явления, по-видимому, является стабилизирующий отбор. Его действие в системе “родители-потомки” обеспечивает высокое качество ее семян, сохранение и передачу от поколения к поколению региональной нормы вида, что достигается за счет повышенной пустосемянности шишек и поддержания гомеостаза внутренней среды в условиях климатической нестабильности.
Из трех признаков семенной продуктивности смертность оплодотворенных семяпочек характеризуется самой высокой амплитудой изменчивости в оптимальные годы, отражая степень внутренней разбалансировки равновесных лесных экосистем сосны обыкновенной на индивидуальном и популяционном уровне их организации. Для оценки степени экологического неблагополучия лесостепных популяций в результате потепления климата данный признак может служить предиктором приближения ЖС популяций к нижнему пределу устойчивости равновесного состояния сосны обыкновенной.
Среднерусская лесостепь характеризуется высокими темпами потепления климата, что негативно отражается на ОЖС сосновых лесов. В последнее десятилетие количество оптимальных лет в регионе сократилось с 7–8/10 лет до пяти. Гидротермический стресс во время засухи повышает вероятность перехода сосны на более низкий уровень функционирования. Для возвращения из неравновесного состояния в равновесие требуется энергия, которая может накапливаться только в оптимальные годы. Дальнейшее сокращение их числа делает практически невозможным возврат сосновых лесов к региональной норме.
Список литературы
Акимов Л.М., Задорожная Т.Н. Особенности распределения трендов температуры воздуха на Европейской территории России и сопредельных государств // Вестник ВГУ. Сер. География. Геоэкология. 2018. № 4. С. 5–14. https://doi.org/10.17308/geo.2018.4/2260
Боголюбов А.С., Буйволов Ю.А., Кравченко М.В. Методика оценки жизненного состояния леса по сосне. М.: Экосистема, 1999. 25 с.
Виноградова В.В. Волны тепла на Европейской территории России в начале ХХI века // Известия РАН. Серия географическая. 2014. № 1. С. 47–55. https://doi.org/10.15356/0373-2444-2014-1-47-55
Виноградава В.В., Титкова Т.Б., Черенкова Е.А. Динамика увлажнения и теплообеспеченности в переходных ландшафтных зонах по спутниковым и метеорологическим данным в начале ХХI в. // Современные проблемы дистанционного зондирования Земли из космоса. 2015. Т. 12. № 2. С. 162–172.
Доклад об особенностях климата на территории Российской Федерации за 2020 г. Москва, 2021. 104 с.
Иванов В.П., Марченко С.И., Иванов Ю.В. Влияние погодных условий на женскую генеративную сферу сосны обыкновенной (Pinus sylvestris L.) // Вестник Томского государственного университета. Биология. 2015. № 3(31). С. 114–129. https://doi.org/10.17223/19988591/31/9
Касимов Н.С., Кислов А.В., Чернышев А.В., Семин В.Н., Аляудинов А.Р. Эколого-географические последствия глобального потепления климата ХХI в. на Восточно-Европейской равнине и в Западной Сибири. М.: МАКС Пресс, 2011. 496 с.
Клушевская Е.С., Кузнецова Н.Ф. Оценка устойчивости сосны обыкновенной к засухе по физиологическим характеристикам хвои // Лесоведение. 2016. № 3. С. 216–222.
Крамер К., Ван дер Верф Д.С. Концепции равновесия и неравновесия в генетическом моделировании лесов: популяционные и индивидуальные подходы. Forest Systems. 2010. № 19. С. 100–112. https://doi.org/10.5424/fs/201019S-9312
Кузнецова Н.Ф. Взаимодействие генотип-среда и адаптивная селекция Pinus sylvestris L. на засухо- и стрессоустойчивость // Труды КубГАУ. 2021. № 91. С. 181–186. https://doi.org/10.21515/1999-1703-91-181-186
Кузнецова Н.Ф., Клушевская Е.С., Аминева Е.Ю. Высокопродуктивные сосновые леса в условиях изменяющегося климата // Лесной журн. 2021. № 6. С. 9–23. https://doi.org/10.37482/0536-1036-2021-6-9-23
Куролап С.А., Нестеров Ю.А., Епринцев С.А. Типизация территории Воронежской области по уровню техногенного воздействия на среду обитания // Вестник ВГУ. Серия География. 2010. № 1. С. 5–11.
Обновленский В.М. Физико-географический оптимум произрастания сосны обыкновенной и использование его в семеноводстве // Тез. докл. XI науч.-техн. конф. по итогам науч.-иссл. работ за 1961 г. Брянского технол. ин-та. Брянск, 1962. С. 27–29.
Раутиан А.С. О природе генотипа и наследственности // Журн. общей биологии. 1993. Т. 54. № 2. С. 132–149.
Романовский М.Г. Формирование урожая семян сосны обыкновенной в норме и при мутагенном загрязнении. М.: Наука, 1997. 111 с.
Романовский М.Г. Продуктивность, устойчивость и биоразнообразие равнинных лесов Европейской России. М.: МГУЛ, 2002. 92 с.
Свинцова В.С. Характер изменчивости признаков репродуктивной сферы Pinus sylvestris L. в условиях среднерусской лесостепи: Воронежская область: автореф. дис. … канд. биол. наук: 03.00.05. Воронеж, 2002. 20 с.
Сердюкова А.П. Реакция сосны обыкновенной на изменение климата в условиях степной зоны Воронежской области // Организация и регуляция физиолого-биохимических процессов. Вып. 22. ВГУ, 2020. С. 177–181.
Чудинова Л.А., Орлова Н.В. Изменчивость физиолого-биохимических показателей в зависимости от жизненного состояния модельных деревьев сосны обыкновенной // Экологический мониторинг и биоразнообразие. 2016. Т. 11. № 2. С. 168–172.
Щербаков В.П. Эволюция как сопротивление энтропии. II. Консервативная роль полового размножения // Журн. общей биологии. 2005. Т. 66. № 5. С. 300–309.
Alaquori H.A.A., Ozer Genc C., Aricak B., Kuzmina N., Cetin M. The possibility of using Scots pine (Pinus sylvestris L.) as biomonitor in the determination of heavy metal accumulation // Applied Ecology and Environmental Research. 2020. V. 18. № 2. P. 3713–3727. https://doi.org/10.15666/aeer/1802_37133727
Alberto F.J., Aitken S.N., Alia R. et al., Potential for evolutionary responses to climate change – evidence from tree populations // Global Change Biology. 2013. V. 19. № 6. P. 1645–1661. https://doi.org/10.1111/gcb.12181
Allen C.D., Macalady A.K., Chenchouni H. et al. A global overview of drought and heat induced tree mortality reveals emerging climate change risks for forests // Forest Ecology and Management. 2010. V. 259. № 4. P. 660–684. https://doi.org/10.1016/j.foreco.2009.09.001
Degtyareva A.P. Characteristics of Pinus sylvestris L. seeds under drought conditions in the steppe region of the Central Chernozem region // J. Agriculture and Environment. 2021. № 3(19).https://doi.org/10.23649/jae.2021.3.19.1
Diaz S., Demissew S., Carabias J. et al. The IPBES conceptual framework – connecting nature and people // Current Opinion in Environmental Sustainability. 2015. V. 14. № 3–4. P. 1–16. https://doi.org/10.1016/j.cosust.2014.11.002
Gauthier S., Bernier P., Kuuluvainen T., Shvidenko A.Z., Schepaschenko D.G. Boreal forest health and global change // Science. 2015. V. 349. № 6250. P. 819–822. https://doi.org/10.1126/science.aaa9092
Iglesias V., Whitlock C. If the trees burn, is the forest lost? Past dynamics in temperate forests help management strategies // Philosophical Transactions of the Royal Society B: Biological Sciences. 2020. V. 375. № 1794. P. 20190115. https://doi.org/10.1098/rstb.2019.0115
IPCC. Climate Change 2022: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Montreal, Canada, 2017. P. 3–29.
Kuznetsova N.F. Self-fertility in Scots pine as a system for regulating close relationships and species survival in advance environment // Advances in Genetics Research. V. 9. N.Y.: Nova Science Publ., 2012. P. 83–106.
Kuznetsova N.F., Semenov M.A., Sautkina M.Yu. Pine forests of East European plain: distribution trends, functions and development problems // Pinus: Growth, Distribution and Uses. N.Y.: Nova Science Publ., 2019. P. 1–47.
Kramer K. Equilibrium and non-equilibrium concepts in forest genetic modeling: population- and individually-based approaches // Forest Systems. 2011. № 3(4). P. 100–109. https://doi.org/10.5424/fs/201019S-9312
Дополнительные материалы отсутствуют.