

Лесоведение, 2023, № 3, стр. 228-243
Изменение парцеллярной структуры лиановых широколиственных лесов за 20 лет в Приморском крае
Т. А. Москалюк *
Федеральный научный центр биоразнообразия наземной биоты Восточной Азии ДВО РАН
692022 Владивосток, проспект 100-летия Владивостока, д. 159, Россия
* E-mail: tat.moskaluk@mail.ru
Поступила в редакцию 13.05.2022
После доработки 09.06.2022
Принята к публикации 18.10.2022
- EDN: PTRRVO
- DOI: 10.31857/S0024114823030075
Аннотация
Изучение динамики парцеллярной структуры лесов Приморского края необходимо для выявления закономерностей естественного восстановления уникальной формации региона – коренных хвойно-широколиственных лесов. В результате хозяйственной деятельности эти леса, насыщенные реликтовыми элементами, сменились производными на большей части ареала. Исследования проводились в водосборном бассейне р. Комаровки (Южное Приморье, Уссурийский район). В статье приведены результаты изучения парцеллярной структуры широколиственно-липового с лианами лещинового разнотравного типа леса – репрезентативного для региона и самого сложного в формации производных лесов. Метод исследований классический: на постоянной пробной площади (ППП) выполнено детальное описание и картирование в масштабе 1 : 100 всех ярусов фитоценоза с последующим попарным сравнением картосхем и выделением парцелл. Анализ трансформации фитоценоза за 20 лет восстановительной сукцессии показал, что основные лесоводственно-таксационные характеристики насаждения изменились незначительно. В древостое почти полностью выпала осина и значительная часть одной из главных пород – липы. Одновременно укрепились позиции подлеска и видов деревьев, характерных для коренных хвойно-широколиственных лесов. Изменения в парцеллярной структуре свидетельствуют об успешном восстановлении коренного типа леса: в фитоценозе число парцелл уменьшилось с 12 до 10; к шести условно-коренным добавилась еще одна. Общими остались 6 парцелл, все они условно-коренные. Произошло закономерное укрупнение и условно-коренных парцелл и выравнивание их границ. Отражена высокая индикаторная роль травяного яруса.
Сохранение функций и услуг лесных экосистем в условиях прогнозируемых изменений окружающей среды имеет решающее значение для всего человечества. Общая площадь лесов на планете, по данным Организации Объединенных Наций, сократилась с 1990 г. по 2015 г. на 3%, ежегодные темпы потери лесопокрытой площади в период с 2010 по 2015 год составили 3.3 млн га год–1 (Keenan et al., 2015). Проблемам, прямо или косвенно связанным с утратой биоразнообразия и предотвращением негативного антропогенного воздействия на леса, посвящены многочисленные публикации (Исаев и др., 1997; Caspersen, Pacala, 2001; Hector, Bagchi, 2007; Gao et al., 2014; Усольцев, 2019; Лукина и др., 2020; Тебенькова и др., 2021; Elliot и др., 2022; и др.).
В число целевых задач, обозначенных в Стратегическом плане Конвенции о биологическом разнообразии планеты, входит сохранение и восстановление коренных лесов с высоким потенциалом биоразнообразия (UNSPF, 2017). Это полностью согласуется с долгосрочными планами России, владеющей 46% площади лесов мира (Стратегия …, 2014) и обладающей самым высоким типологическим разнообразием лесных формаций. В их число входят хвойно-широколиственные леса Приморского края, определяющие уникальность и своеобразие природы региона. Отсутствие покровного оледенения в доисторические времена и благоприятные современные растительные условия обусловили высокую насыщенность приморских лесов реликтовыми и эндемичными видами. Всемирным фондом дикой природы они включены в список особо ценных природных объектов планеты.
Доминанты хвойно-широколиственных лесов: кедр корейский (Pinus koraiensis22) и пихта цельнолистная (Abies holophylla), их спутники: липа амурская (Tilia amurensis) и липа маньчжурская (T. mandshurica), граб сердцелистный (Carpinus cordata), орех маньчжурский (Juglans mandshurica), калопанакс, или диморфант, семилопастной (Kalopanax septemlobus), бархат амурский (Phellodendron amurense) и другие многочисленные реликты маньчжурской флоры неспособны быстро реагировать на изменения среды, вызываемые антропогенными факторами. Насаждения с перечисленными видами утрачивают средообразующие функции даже при малом нарушении системной целостности. В настоящее время им грозит полное исчезновение (Ковалев и др., 2020).
В результате длительных выборочных рубок и пожаров площадь дальневосточных кедровников с 1929 г. по 2001 г. уменьшилась в 1.9 раза (Корякин, Челышев, 2009). В Приморском крае с 1989 г. по 2010 г. она сократилась в 1.4 раза – с 3.04 до 2.16 млн га (Сибирина, 2014) и продолжает сокращаться. Главный современный фактор гибели лесов – пожары. В кедровниках края из-за пожаров произошла смена типов леса более чем на 1.5 млн га (Иванов и др., 2022). Перестойные кедровники в Приморье к концу минувшего столетия были уничтожены полностью (Петропавловский, 2004). Девственные чернопихтовые леса, северная граница которых проходит по Южному Приморью, остались только на заповедных территориях.
Одной из самых распространенных формаций в крае со второй половины минувшего столетия стала производная формация широколиственных лесов. В конце прошлого века на эти леса в южной части Приморского края приходилось около 20% площади гослесфонда (Кудинов, 2000). В настоящее время доля площади лесов с доминированием широколиственных пород составляет уже более 22% (Распределение площади …, 2021).
Для своевременного выявления негативных последствий, вызываемых разными причинами, и принятия правильных решений по их предупреждению и (или) ликвидации важное значение имеет мониторинг состояния экосистем и динамики экосистемных функций, или лесообразовательного процесса. Изучение лесообразовательного процесса необходимо также для выявления закономерностей естественного восстановления коренной хвойно-широколиственной формации и представляет одно из главных направлений дальневосточного лесоведения (Петропавловский, 2004; Ковалев и др., 2020).
В связи с этим особую актуальность приобретают изучение и мониторинг парцеллярной структуры, в которой отражается состояние сообщества в любой период его жизнедеятельности. Анализ структуры позволяет выявить важные функциональные особенности сообщества на каждой стадии онтогенеза в зависимости от лесорастительных условий, лесообразующих пород и воздействия экзогенных и эндогенных факторов (Дылис, 1974; Mestre et al., 2017; Spicer et al., 2020).
При относительно высокой общей изученности лесов Приморья многие аспекты их функционально-пространственной организации исследованы слабо. Начало исследованиям по горизонтальной структуре лесов Приморского края было положено в середине минувшего столетия известным геоботаником П.Д. Ярошенко (Иванова и др., 1963) в лесах Уссурийского заповедника. Через двадцать лет их продолжила Н.А. Крупянко (1983). В разные годы подобные исследования проводились в пихтово-еловых лесах Северного Сихотэ-Алиня (Комарова, 1978), кедрово-широколиственных лесах Сихотэ-Алиня (Максимова, 1987) и широколиственно-кедровых лесах Среднего (Комарова, 1993), Южного и Северного Сихотэ-Алиня (Возмищева, Перепелкина, 2015). Авторами перечисленных работ исследовались девственные леса. На всех объектах основное внимание уделялось мозаике травяного яруса, морфологической согласованности его структуры с внутриценотической средой. В конце минувшего и начале текущего столетий объектами изучения пространственной неоднородности стали производные леса Горнотаёжной станции ДВО РАН (Москалюк, 2006, 2021; Москалюк, Тарасова, 2015; и др.). Публикации по динамике парцеллярной структуры лесных фитоценозов в Приморском крае отсутствуют.
Целью наших исследований было изучение парцеллярной структуры и ее трансформации за 20 лет в широколиственно-липовом с лианами лещиновом разнотравном лесу – репрезентативном и самом сложном из производных типов леса на юге Приморского края. Для достижения этой цели запланировано выявление видового состава и особенностей пространственного распределения видов и их группировок с учетом приуроченности к конкретным микросайтам, изучение характера взаимоотношений между растительными ярусами, выделение и детальное описание ценоэлементов в пределах ярусов и фитоценоза, анализ изменения структурных элементов во времени и пространстве.
ОБЪЕКТЫ И МЕТОДИКА
Район исследований – окрестности пос. Горнотаёжное (Уссурийский район) в пределах бассейна р. Комаровки, входящего в систему водосборного бассейна Японского моря. По лесорастительному зонированию Д.И. Назимовой (2015), территория района исследований относится к зоне смешанных широколиственно-хвойных лесов с дубом Дальневосточного муссонно-континентального сектора, по известному ботанико-географическому районированию Б.П. Колесникова (1969), район исследований входит в южную подзону зоны смешанных хвойно-широколиственных лесов южной фации кедровников с пихтой цельнолистной бассейнов рек, впадающих в залив Петра Великого. Природные условия этой подзоны, как и всего Южного Приморья, определяются муссонным климатом, положением относительно Японского моря и преобладанием низкогорного рельефа с высотами до 1000 м н.у.м. Отсутствие покровного оледенения в доисторические времена и благоприятный современный климат обусловили очень высокое биологическое разнообразие как на видовом, так и на экосистемном уровнях.
Леса в районе исследований в основном производные, типичные для районов Южного Приморья (Кудинов, 2000). К инсолируемым склонам южных экспозиций приурочены монодоминантные сухие дубняки, к теневым склонам северных экспозиций с мягким микроклиматом и высоким плодородием равномерно увлажненных почв – полидоминантные широколиственные леса. В связи с прекращением температурной инверсии, в средних частях теневых склонов складываются оптимальные лесорастительные условия. Именно в них формируются полидоминантные леса с теплолюбивыми видами лиан – актинидией острой (Actinidia arguta) и актинидией коломикта (A. kolomicta) – чернопихтарники и грабовые варианты кедровников с запасами древесины до 800 м3 га–1 (Васильев, Колесников, 1962).
Объект исследований – широколиственно-липовый с лианами лещиновый разнотравный тип леса (рис. 1) – расположен в 1.5 км к северо-востоку от поселка в средней части северо-северо-восточного склона и занимает территорию с 200 до 400 м от подножия склона. Поверхность склона сравнительно ровная, с террасовидными элементами микрорельефа в виде небольших выпуклостей и понижений; уклон составляет в среднем около 15°, в местах перегиба может варьировать от 5° до 40°. Высотные отметки склона – от 170 (подножие) до 270 (водораздел) м н.у.м., протяженность – 610 м.
Рис. 1.
Широколиственно-липовый с лианами лещиновый разнотравный тип леса. Фрагмент “актинидиевой” парцеллы весной.

Методы исследований. В 1997–1998 гг. в “лиановом” фитоценозе заложена ППП размером 60 × 70 м, в 2018 г. проведена ее ревизия. Координаты северо-восточного угла пробной площади: 43°42′272″ с.ш. и 132°09′611″ в.д. Исследования проводились в соответствии с рекомендациями В.Н. Сукачева (1961) и А.И. Уткина (1982). Размеры и форма пробной площади отвечают требованиям ОСТа 56-69-83 (1983) – на ней представлены более 200 деревьев главных лесообразователей и все ценоэлементы структуры фитоценоза.
В 1997 г. были описаны местоположение и экологические условия пробной площади, выполнен сплошной перечет деревьев с указанием их жизненного состояния. Таксационные показатели пород рассчитывались по региональным справочным материалам (Справочник…, 1990). Предварительно пробная площадь была разбита на клетки – квадраты 10 × 10 м. Для растительных ярусов составлены картосхемы в масштабе 1 : 100 с указанием на них центров стволов и крон деревьев, проекций кустарников и контуров микрогруппировок травяного яруса. Картирование нижних ярусов проводилось следующим летом, так как при работе с древостоем травы были сильно измяты.
Травяной ярус – высокоинформативный индикатор экологических условий в фитоценозе (Назимова и др., 2015; Рубцов, Рыбакова, 2019). Большое внимание в работе уделялось изучению его горизонтальной структуры, в которой наглядно отражается взаимодействие факторов внешней и внутренней среды с ценопопуляциями видов. За основной структурный элемент яруса принята микрогруппировка – однородная по составу видов, проективному покрытию и жизненному состоянию совокупность растений одного участка. Контуры микрогруппировок на план наносились относительно границ клеток и расположения оснований деревьев и куртин кустарников. Картирование выполнялось с 3-ей декады мая до конца июня – периода, когда эфемероиды исчезли еще не полностью, а летние травы отросли настолько, что можно было установить границы их синузий. Характеристики микрогруппировок (приуроченность к микрорельефу, проективное покрытие, габитус и обилие растений каждого вида по шкале Друде) определялись по самому крупному и типичному фрагменту. Минимальная площадь выделения – 0.5 м2.
Названия микрогруппировок травяного яруса соответствовали названию вида-доминанта (шлемниковая, василистниковая, адиантовая) или группе видов-содоминантов со сходной экологией (осоковая, разнотравная, мелкотравная). При значении проективного покрытия менее 60% название микрогруппировки дополнялось словом “разреженная”, при менее 40% – словом “редкопокровная”.
Основным ценоэлементом фитоценоза принята парцелла в трактовке Н.В. Дылиса (1969), так как при ее выделении учитывались характеристики всех ярусов и условий экотопа. Парцеллы выделялись на основании попарного сравнительного анализа картосхем и описаний ярусов. Объективность выделения парцелл подтверждена методом графов. Флористическое сходство между парцеллами вычислялось с помощью критерия Жаккара, характеризующего степень сходства участков (клеток ППП) растительного покрова по качественным (число видов) и количественным (обилие видов, проективное покрытие) признакам (Дуденко, 2012). В названия парцелл – по аналогии с названиями типов леса – включались названия эдификаторных видов древостоя и подлеска (при сомкнутости кустарников выше 0.6.) и фонообразующей микрогруппировки травяного яруса.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Широколиственно-липовый с лианами лещиновый разнотравный тип леса33 представляет заключительную стадию восстановительной сукцессии. Он сформировался на месте чернопихтово-широколиственного леса, пройденного выборочной рубкой и огнем в середине прошлого века. Он включает более 110 видов высших сосудистых растений, отличаясь от ранее произраставшего насаждения лишь отсутствием пихты цельнолистной. Такие леса принято считать условно-коренными (Манько, 1984; цит. по работе Кудинова, 2000).
Трансформация фитоценоза, произошедшая с 1997 г. по 2018 г., обусловлена естественной динамикой древостоя (табл. 1). За 20 лет в насаждении почти полностью исчезла осина (Populus tremula), по 63% от прежнего числа выпало деревьев ясеня (ясеня маньчжурского (Fraxinus mandshurica) и ясеня носолистного (F. rhynchophylla)) и маакии амурской (Maackia amurensis). Из главных пород больше всего деревьев “потеряла” липа – около 32% от общего числа; в основном выпали перестойные и тонкомерные деревья. У дуба монгольского (Quercus mongolica) выпало только 2.5%, у ореха – не более 1%. При этом увеличилась численность пород, характерных для свиты хвойно-широколиственных лесов: граба, кленов – в большей степени клена ложнозибольдова (Acer pseudosieboldianum), ильма лопастного (Ulmus laciniata), мелкоплодника (Micromeles alnifolia). Формула состава (по запасу) изменилась мало: 1997 г. – 5Лп3Д1Км1Яс + И, Ор, ед. Бд, Дмр, Ос, Ма, Г; 2018 г. – 4Лп3Д1Км1Ор1Яс + И, Бд, Дмр, ед. Ма, Г. Прежними остались видовой состав древостоя – 22 вида и распределение деревьев по площади – равномерное. Средний диаметр древостоя увеличился у деревьев 1-го яруса (высота 16 м и выше) – на 2–3 ступени толщины, у деревьев 2-го яруса – на 1–2 ступени.
Таблица 1.
Таксационные показатели растущего древостоя
| Индекс вида | Число деревьев, шт. га–1 | Средние | Площадь сечения стволов, м2 га–1 | Запас, м3 га–1 | Полнота (рассчитана по запасу) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| диаметр, см | высота, м | |||||||||||
| 1997 г. | 2018 г. | 1997 г. | 2018 г. | 1997 г. | 2018 г. | 1997 г. | 2018 г. | 1997 г. | 2018 г. | 1997 г. | 2018 г. | |
| Лп | 276 | 188 | 22.6 | 27.1 | 16.7 | 17.5 | 11.1 | 10.81 | 92.4 | 87.3 | 0.40 | 0.30 |
| Д | 100 | 88 | 30.2 | 36.9 | 17.4 | 17.6 | 7.36 | 9.40 | 56.5 | 74.2 | 0.21 | 0.25 |
| Км | 129 | 148 | 17.3 | 14.2 | 12.0 | 11.1 | 3.01 | 3.00 | 20.9 | 20.3 | 0.21 | 0.19 |
| Ясм, Ясн | 50 | 19 | 18.6 | 26.2 | 15.1 | 18.2 | 1.37 | 1.03 | 9.55 | 9.70 | 0.07 | 0.05 |
| Ор | 24 | 19 | 24.5 | 33.1 | 15.5 | 16.9 | 1.12 | 1.64 | 8.01 | 12.82 | 0.03 | 0.08 |
| Иг, Ид | 24 | 31 | 15.9 | 15.4 | 14.2 | 14.0 | 0.47 | 0.58 | 5.58 | 5.26 | 0.21 | 0.20 |
| Ма | 19 | 12 | 13.1 | 18.2 | 11.6 | 12.9 | 0.26 | 0.31 | 1.63 | 2.12 | 0.02 | 0.02 |
| Гр | 10 | 16 | 11.4 | 11.3 | 8.3 | 8.3 | 0.25 | 0.39 | 1.28 | 1.99 | <0.01 | <0.1 |
| Дмр | 9 | 9 | 22.9 | 30.5 | 15.1 | 16.5 | 0.29 | 0.52 | 2.13 | 4.09 | 0.01 | 0.02 |
| Клжб | 40 | 52 | 7.6 | 8.6 | 6.7 | 7.0 | 0.16 | 0.31 | 0.67 | 1.3 | <0.1 | <0.1 |
| Тр | 40 | 24 | 6.6 | 7.8 | – | – | 0.14 | 0.11 | – | – | <0.01 | <0.01 |
| Мп | 5 | 7 | 7.0 | 8.2 | 9.0 | 9.7 | 0.02 | 0.04 | 0.07 | 0.16 | <0.01 | <0.01 |
| Бд | 5 | 2 | 22.9 | 51.0 | 16.7 | 19.2 | 0.38 | 0.47 | 2.78 | 5.22 | 0.01 | 0.02 |
| Ос | 10 | 2 | 18.9 | 22.2 | 15.7 | 16.6 | 0.27 | 0.09 | 1.87 | 0.65 | 0.01 | <0.01 |
| Итого | 741 | 617 | 21.3 | 24.6 | – | – | 26.2 | 28.7 | 203.4 | 225.0 | 1.19 | 1.18 |
Примечание. Породы: Лп – липа амурская, липа маньчжурская, Д – дуб монгольский, Км – клен мелколистный (Acer mono), Ясм – ясень маньчжурский, Ясн – ясень носолистный, Ор – орех маньчжурский, Иг – ильм лопастной (Ulmus laciniata), Ид – ильм японский (U. japonica), Ма – маакия амурская (Maackia amurensis), Гр – граб сердцелистный, Дмр – калопанакс семилопастной, Клжб – клен ложнозибольдов, Тр – трескун (Ligustrina amurensis), Мп – мелкоплодник ольхолистный (Micromeles alnifolia), Бд – береза даурская (Betula dahurica), Ос – осина.
Сомкнутость крон по-прежнему высокая – 0.95, но за счет отпада древостоя снизилась степень перекрытия крон. Как следствие, улучшилась освещенность под пологом леса, за исключением микросайтов с деревьями, в кронах которых расплелись лианы актинидии. Улучшение световых условий обеспечило повышение жизненности подроста и подлеска.
В подросте в достаточном количестве (около 7 тыс. экз. га–1) представлены все виды древостоя. В конце минувшего столетия на склоне единично росли особи мелкого подроста пихты. Они погибли рано весной 1998 г. после низового пожара. Лиственные кустарники за несколько лет восстановились полностью. Пробную площадь огонь не затронул.
Среди 11 видов кустарников на протяжении всего периода исследований преобладали лещина маньчжурская (Corylus mandshurica) и чубушник тонколистный (Philadelphus tenuifolius), обычными были трескун и особенно в 2018 г. калина бурейская (Viburnum burejanum). Общая сомкнутость подлеска увеличилась всего на 0.1 ед. – с 0.5–0.6 до 0.6–0.7, но в пределах фитоценоза усилилась неравномерность распределения кустарников по площади. Различия между показателями сомкнутости стали более разительными – от 0.1 (на сырых участках в основаниях террасовидных уступов и под деревьями с высокой сомкнутостью крон или с кронами, перевитыми лианами) до 1.0 (места вывала деревьев). Разреженность подлеска присуща также участкам с выпуклой поверхностью и потому более сухих. В таких экотопах, кроме разрозненных группировок чубушника и лещины, одиночными побегами высотой менее 1 м растут барбарис амурский (Berberis amurensis), калина Саржента (Viburnum sargentii) и смородина маньчжурская (Ribes mandshuricum).
В травяном ярусе выявлено 79 видов в 1998 г. и 74 – в 2018 г.; выпали светолюбивые и заносные виды. Доминируют лесной мак весенний (Hylomecon vernalis) разнотравье, папоротники, хвощ зимующий (Equisetum hiemale), и осоки (осока кривоносая (Carex campylorhina), о. возвратившаяся (C. cf reventa) и о. ржавопятнистая (C. siderosticta)). Преобладающими были и остались микрогруппировки разнотравного и – весной – эфемероидного комплексов, в совокупности занимающих почти 70% площади фитоценоза (Москалюк, 2021). Усиление эдификаторных функций подлеска и молодого поколения лиственных пород проявилось в повышении мозаичности травяного яруса – в горизонтальной структуре яруса число микрогруппировок за 20 лет увеличилось с 52 до 67.
Главной особенностью насаждения является наличие взрослых лиан актинидии острой, поднимающихся в кроны деревьев первого яруса, и коротких, как правило, стелющихся, лиан лимонника (Schizandra chinensis) и винограда (Vitis amurensis).
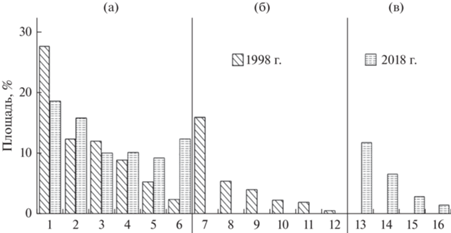
В коренных хвойно-широколиственных лесах с выработанной структурой в общей сложности насчитывается не более 4–5 парцелл (Максимова, 1987). Парцеллярная структура “лианового” фитоценоза окончательно еще не сформирована. В 1998 г. в нем выделены 12 парцелл (рис. 2а), из которых к 2018 г. прежними остались шесть. Вместо остальных шести появились четыре новых парцеллы (рис. 2б). Контуры парцелл стали более плавными, а распределение площади по парцеллам – более равномерным. Основными – преобладающими по площади (Дылис, 1969) в 1998 г. были пять парцелл с размерами от 9.0 до 27.8% от площади фитоценоза, а в 2018 г. их стало уже семь с размерами от 9.3 до 18.7%. Общая площадь этих парцелл различается несильно: 77.5% – в 1998 г. и 79.5 – в 2018 г. Самая большая из дополняющих парцелл в 1998 г. занимала 5.5%, в 2018 г. – 6.7%.
Рис. 2.
Парцеллярная структура широколиственно-липового с лианами лещинового разнотравного фитоценоза. Парцеллы “общие” для 1998 г. и 2018 г.: 1 – дубово-липовая лесно-маковая (27.8% от площади фитоценоза), 2 – дубово-липовая с кленом мелколистным трескуново-лещиновая редкопокровная (12.5%), 3 – широколиственная актинидиевая с трескуном и лещиной редкопокровная (12.1%), 4 – липово-дубовая с кленом мелколистным разнотравная с осокой редкопокровная (9.0%), 5 – липовая с клёном ложнозибольдовым разнотравно-осоковая (5.4%), 6 – ясенево-широколиственная хвощовая разреженная (2.5%). Парцеллы, исчезнувшие к 2018 г.: 7 – дубово-липовая с кленом мелколистным разнотравная с осокой (16.1%), 8 – кленово-мелколистно-липовая разнотравно-звездчатковая (5.5%), 9 – широколиственная актинидиевая лесно-маковая (4.1%), 10 – липово-дубовая василистниковая с разнотравьем (2.4%), 11 – маакиево-диморфантовая волжанковая (2.0%), 12 – берёзово-липовая бересклетовая разнотравная с осокой и василистником (0.6%). Парцеллы, появившиеся к 2018 г.: 13 – липовая-кленово-мелколистная чубушниковая редкопокровная (11.9%), 14 – маакиево-диморфантовая лещиновая редкопокровная (6.7%), 15 – широколиственная актинидиевая калиновая редкопокровная (3.0%), 16 – дубовая с лещиной редкопокровная (1.6%).
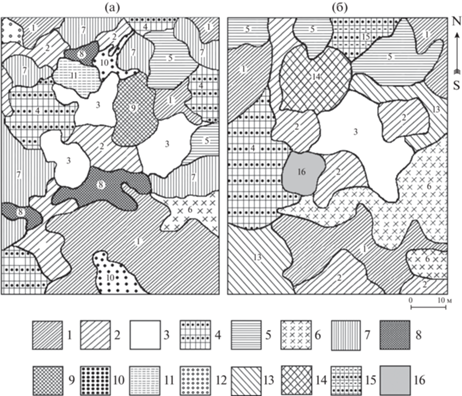
На первом месте по занимаемой площади в сообществе была и осталась условно-коренная дубово-липовая лесно-маковая (1, “лесно-маковая”)44 парцелла. Площадь лесно-маковой парцеллы к 2018 г. сократилась в 1.5 раза, но по-прежнему осталась самой большой (рис. 3). В ее древостое преобладают липа и дуб (формула состава 5Лп3Д1Яс1Ор + И, Тр). Небольшой уклон (5°–7°) – меньший, чем на смежных участках, способствует хорошему освещению поверхности не только весной, но и летом. Лесной мак до июня формирует густой покров с проективным покрытием 90–100%. Кроме него в этой парцелле обычна ветреница дальневосточная (Anemonoides extremiorientalis). К середине лета лесной мак полностью исчезает и фонообразующей становится разнотравная редкопокровная микрогруппировка с проективным покрытием 20–25%.
Рис. 3.
Распределение площади фитоценоза по парцеллам в 1998 г. (а и б) и 2018 г. (а и в). Названия и номера парцелл указаны в легенде к рис. 2.
На самом большом участке парцеллы в нижней части пробной площади выпали одновременно несколько деревьев липы. В окна “вклинились” фрагменты двух парцелл: смежной дубово-липовой с кленом мелколистным трескуново-лещиновой редкопокровной и новой липово-кленово-мелколистной с чубушником редкопокровной.
Дубово-липовая с кленом трескуново-лещиновая редкопокровная (2, “трескуново-лещиновая редкопокровная”) парцелла (рис. 4) – одна из основных, условно-коренная, занимала и занимает несколько участков с уклоном 5°–7° и более. Формула состава древостоя – 5Лп3Д1Км1Яс + + Ос, Гр, ед. Ма). Эта парцелла выделялась особенно густым ярусом лещины и немногочисленными куртинами клена бородчатого (Acer barbinerve). За 20 лет в ней сильно разросся трескун. Высота кустарников достигает 4 м. Выступая соэдификатором древостоя, подлесок сильно затеняет поверхность и сдерживает разрастание трав. В фонообразующую разнотравную редкопокровную микрогруппировку (проективное покрытие до 20%) вкраплены мелкие синузии разнотравья и одиночные особи крупнотравья. На некоторых микросайтах трескун принимает стелющуюся форму, заменяя травяной ярус. К 2018 г. площадь парцеллы увеличилась в 1.2 раза.

Широколиственная актинидиевая с трескуном и лещиной редкопокровная (3, “актинидиевая”) парцелла относится к наиболее характерным в фитоценозе. В 1998 г. она занимала три участка в центральной части пробной площади, которые примыкали к впадине, расположенной поперек склона. Формула состава древостоя 4Д3Лп2Яс1Ор, ед. Гр, Км, Тр, Ма. Многие деревья оплетены лианами актинидии. Толщина некоторых лиан у основания достигает 10–12 см. У крупных лиан образуется множество дочерних. Малочисленные деревца подроста деревьев и кустарники в 1998 г. были сильно угнетены. Летом в парцелле всегда сумрачно и сыро. Фон создают разнотравная редкопокровная микрогруппировка и листовой опад. Весной аспектирует лесной мак, сквозь опад пробиваются эфемероиды (адокса мускусная (Adoxa moschatellina), лук одноцветковый (Allium monanthum), лютик Франше (Ranunculus franchetii) и др.).
Между двумя участками парцеллы за счет бокового освещения со стороны большой прогалины, образовавшейся в результате вывала перестойной липы, проективное покрытие трав увеличилось до 60%. В этом месте в 1998 г. была выделена производная широколиственная актинидиевая лесно-маковая (9) парцелла с очень густыми микрогруппировками лесного мака. После зарастания прогалины лещиной парцелла снова стала широколиственной актинидиевой с трескуном и лещиной редкопокровной.
После очередного вывала нескольких деревьев в соседних парцеллах освещение в “актинидиевой редкопокровной” парцелле улучшилось, и на двух ее участках сформировались группировки из лещины и трескуна. После этого к названию парцеллы было добавлено сочетание слов “с трескуном и лещиной”. На третьем участке разрослась калина буреинская (Viburnum burejanum), образовав куртину из 22 кустов высотой от 0.8 до 1.6 м. Это стало основанием для выделения в 2018 г. одной из самых маленьких парцелл (см. рис. 2б и 3) – производной широколиственной актинидиевой калиновой редкопокровной парцеллы (16).
Липово-дубовая с кленом мелколистным разнотравная с осокой редкопокровная (4, “разнотравная с осокой редкопокровная”) парцелла – условно-коренная и последняя из пяти основных в 1998 г. Формула состава древостоя 4Лп3Д1Км1И1Ос + Гр, ед. Тр. В 1998 г. она располагалась в разных местах пробной площади. Для нее характерна слегка выпуклая поверхность с направлением уклона не только вниз, но и к востоку, и потому более сухая по сравнению с остальной территорией фитоценоза. Присущие парцелле разреженность подлеска и редкопокровность травяного яруса обусловлены, по-видимому, не только высокой сомкнутостью крон, но и конкуренцией за влагу и питательные элементы в почве. В подлеске парцеллы, кроме обычных видов, единичны особи смородины и барбариса.
За 20 лет осина полностью выпала. От четырех фрагментов парцеллы остался один (см. рис. 2), но площадь парцеллы не только не уменьшилась, а увеличилась – за счет присоединившегося к ней фрагмента производной “разнотравной с осокой” парцеллы. В редком травяном ярусе вместе с разнотравьем росли и растут мезофитные осоки и василистник нитчатый (Thalictrum filamentosum).
Липовая с кленом ложнозибольдовая разнотравно-осоковая (5, “кленово-ложнозибольдовая”) парцелла – еще одна из условно-коренных. Она из числа немногих парцелл, в формировании которых четко прослеживается связь с эдафическими условиями, главным образом, с влажностью почв. На пробной площади самый большой участок парцеллы представлен блюдцеобразным микропонижением, ограниченным с восточной и южной сторон пологими откосами террасовидного уступа. Высота откосов – 1.2–1.5 м, крутизна – до 20°. В западном направлении и вниз микропонижение плавно переходит в основную поверхность склона. В связи с перераспределением склонового стока в сторону уклона откосов влажность почв в парцелле гораздо выше, чем на смежной территории.
Древостой парцеллы разрежен и представлен малым числом видов – формула состава 8Лп1Д1Кл. В нем растет и плодоносит самое крупное на пробной площади дерево клена ложнозибольдова. Ажурные кроны кленов и высоких лип пропускают достаточно света, и на большей части парцеллы сформировался густой травяный ярус (проективное покрытие до 100%). Подлесок редкий, представлен небольшими кустами жимолости ранней (Lonicera praeflorens), чубушника и кустовидного клена ложнозибольдова.
Высокую влажность почв индицирует осока кривоносая – доминант травяного яруса парцеллы (рис. 5). Обычны мезофитные осоки – подобные осоке возвратившейся и ржавопятнистой. Фонообразующие микрогруппировки разнотравно-осоковая и осоковая с разнотравьем разреженная.

За 20 лет облик парцеллы почти не изменился, за исключением снижения обилия василистника, который в 1998 г. был содоминантом осок. В то же время площадь парцеллы увеличилась в 1.7 раза (см. рис. 3). В нее вошли два фрагмента производной “разнотравно-осоковой” парцеллы – из-за разрастания осоки кривоносой и вся “бересклетовая”.
К особо значимым трансформациям парцеллярной структуры “лианового” фитоценоза относится образование условно-коренной ясенево-широколиственной хвощовой разреженной (6, “хвощовой”) парцеллы. Состав ее древостоя 4Яс3Лп1Д1Км ед., Ма, Тр.
В 1998 г. в одном месте – по тальвегу самой большой впадины, была выделена одна густопокровная (обилие cop3-soc) и несколько редкопокровных микрогруппировок с доминированием хвоща зимующего (Equisetum hiemale). Вместе они занимали всего 2.5% площади фитоценоза, но к 2018 г., после вывала нескольких деревьев ясеня и липы, площадь их увеличилась в 5 раз. Потеснив “разнотравную с осокой” и “лесно-маковую” парцеллы, хвощовая заняла всю впадину и вошла в число основных парцелл.
В настоящее время идет экспансия хвоща в смежные парцеллы, указывая на повышение влажности почв. В периоды таяния снега и тайфунов в некоторых местах впадины из откоса террасовидного уступа на дневную поверхность выходят струйки воды.
Фон в парцелле создают микрогруппировки с максимальным обилием хвоща cop2–3 (рис. 6). В этой парцелле выделена единственная на пробной площади мелкотравная майничная с разнотравьем разреженная микрогруппировка с обилием майника (Maianthemum bifolium) cop1 (по шкале Друде). Майник и хвощ – виды-спутники коренных лесов, встречаются во многих микрогруппировках, всегда рассеянно – обилие каждого до sol.

Дубово-липовая с кленом мелколистным разнотравная с осокой (7, “разнотравная с осокой”) парцелла – производная; в 1998 г. – вторая по величине в фитоценозе. В прошлом ее местообитания на склоне были пройдены сильным пожаром. По составу древостоя она одна из самых сложных (4Лп4Д1Яс1Км, + Ос, ед. И, Ор, Гр, Дмр) с перестойными деревьями и корнеотпрысками осины. Разреженный травяный ярус (проективное покрытие – 40%) отличался высокой мозаичностью (17 микрогруппировок). Фонобразующие микрогруппировки: осоковая с василистником и разнотравьем разреженная и разнотравная редкопокровная.
К 2018 г., после выпадения осины и осветления экотопа, парцелла исчезла. Площадь ее распределилась между смежными парцеллами (см. рис. 2). Самый большой фрагмент отошел к условно-коренной “разнотравной с осокой редкопокровной” парцелле. В его подлеске наряду с лещиной и чубушником стал обычным элеутерококк и появились одиночные особи-побеги барбариса и калины Саржента; число микрогруппировок в травяном ярусе сократилось до 9.
К микросайтам со свежими нарушениями были приурочены две производные парцеллы, исчезнувшие к 2018 г.: кленово-мелколистно-липовая разнотравно-звездчатковая (8, “звездчатковая”) и маакиево-диморфантовая волжанковая (11, “волжанковая”). “Звездчатковая” парцелла занимала несколько участков: один – на небольшом участке свежей гари, “зацепившей” край пробной площади, остальные – в местах вывала деревьев. Она хорошо выделялась микрогруппировками звездчатки (Stellaria bungeana): чистыми или в смеси с разнотравьем. В 2018 г. парцеллы уже не было, но звездчатка встречалась в других микрогруппировках.
“Волжанковая” парцелла (рис. 7а) сильно отличалась от окружающих парцелл. Она сформировалась на месте вывалившейся очень старой большой липы. Вероятно, до вывала липы “волжанковая” парцелла была фрагментом соседней “актинидиевой редкопокровной” парцеллы. Состав древостоя как прост, так и необычен: 7Дмр2Ма1Тр. В парцелле росли стройная высокая маакия и огромный высоковозрастный диморфант в окружении нескольких тонкомерных деревьев трескуна. Кроны упавшей липы и диморфанта были густо переплетены лианами актинидии. В 1998 г. сухие сучья липы возвышались большой кучей хвороста, от которой к диморфанту шли туго натянутые отмершие лианы.
Рис. 7.
Парцеллы маакиево-диморфантовая волжанковая (7а) и сменившая её маакиево-диморфантовая лещиновая редкопокровная (7б).

В редком подлеске отмечены чубушник, лещина, жимолость и элеутерококк. Кустарники и подрост были сильно угнетены, всходы и самосев отсутствовали. Волжанка двудомная (Aruncus dioicus) явно препятствовала внедрению других видов в парцеллу. Фонообразующая микрогруппировка – чистая волжанковая с проективным покрытием 100%. Лишь в ее периферийной части росли василистник и звездчатка с разнотравьем.
По мере смыкания крон деревьев, окружающих прогалину, заросли волжанки стали вытесняться лещиной. Спустя 20 лет “волжанковую” парцеллу сменила маакиево-диморфантовая лещиновая редкопокровная (14), дополнительно захватившая участки соседних “звездчатковой”, “василистниковой” и частично “актинидиевой” парцелл (рис. 7б). Под высокосомкнутым пологом лещины дополнительно затеняют поверхность слабо развитые особи элеутерококка. Во-лжанковую микрогруппировку заменила разнотравная редкопокровная с вкраплениями фрагментов прежних микрогруппировок. От кучи “хвороста” и огромного валежного ствола не осталось и следа.
Липово-дубовая василистниковая с разнотравьем (10, “василистниковая”) парцелла в 1998 г. была одной из немногочисленных парцелл с густым травяным ярусом. В прошлом она, как и “разнотравная с осокой” парцелла, значительно пострадала от пожара, о чем свидетельствует присутствие в составе древостоя осины (формула состава 6Д3Лп1Ос + Км, ед. Ма). На участках под василистниковыми микрогруппировками зафиксировано значительное количество остатков разложившейся древесины, что позволяет предположить тяготение василистника к повышенному содержанию органики в почве. Высокое обилие василистника отмечалось и на участке с хорошим боковым освещением – близ прогалины с волжанкой. Василистник нитчатый – характерный вид хвойно-широколиственных лесов. В “лиановом” фитоценозе он и через 20 лет присутствовал во многих микрогруппировках, но с повышением сомкнутости подлеска обилие василистника повсеместно снизилось. “Василистниковая” парцелла объединилась с одной из условно-коренных парцелл или с производной дубовой с лещиной редкопокровной (14). Самый большой фрагмент василистниковой парцеллы (см. рис. 2) “поглотила” условно-коренная “трескуново-лещиновая редкопокровная” парцелла. В производной парцелле почти всю площадь под кронами лещины занимает василистниковая редкопокровная микрогруппировка.
Самой маленькой в фитоценозе в 1998 г. была березово-липовая бересклетовая разнотравная с осокой и василистником (12, “бересклетовая”) парцелла. Она располагалась в локальном микропонижении, аналогичном описанному в “кленово-ложнозибольдовой” парцелле. Формула состава древостоя в 5Лп5Бд, ед. Тр. На бровке откоса микропонижения росло и продолжает расти единственное в парцелле перестойное дерево березы (Betula davurica). Бересклет Максимовича (Еuonymus maximovicziana) в этой парцелле формировал группы по два-три тонкомерных дерева высотой до 6 м. К 2018 г. из пяти лип выпали три, в кроне березы усохло много скелетных ветвей; полностью усохли бересклет и трескун. В травяном ярусе резко снизилось обилие разнотравья и василистника, господство перешло к осоке кривоносой. Бересклетовая парцелла стала одним из фрагментов “кленово-ложнозибольдовой” парцеллы.
Липово-кленово-мелколистная чубушниковая редкопокровная парцелла (13) – последняя из семи условно-коренных парцелл, выделена в 2018 г. Главная порода ее древостоя – клен мелколистный. От парцелл с густым лещиновым подлеском эта парцелла отличается меньшей влажностью местообитаний, что способствовало после улучшения освещенности разрастанию в ней чубушника (рис. 8), образующего как чистые группировки, так и в смеси с элеутерококком. Сомкнутость подлеска 0.8–0.9, высота кустов достигает 3.0 м. Фонообразующая микрогруппировка – разнотравная редкопокровная.

ВЫВОДЫ
1. Широколиственно-липовый с лианами лещиновый разнотравный тип леса представляет производную формацию смешанных широколиственных лесов, восстановившихся на месте коренных кедрово- и чернопихтово-широколиственных лесов в южной части Приморского края. Он сформировался в оптимальных лесорастительных условиях и обладает сложным флористическим составом, включающим 22 вида деревьев, 11 – кустарников, 3 – лиан и более 80 видов трав.
2. Анализ результатов исследований показал, что широколиственно-липовый тип леса – устойчиво-производный вступил в завершающую стадию восстановительной сукцессии. В составе древостоя с 1997 г. по 2018 г. снизилось участие главных пород (липа и ясень), почти полностью выпала осина, укрепились позиции видов, характерных для хвойно-широколиственных лесов (граб сердцелистный, дуб монгольский, орех маньчжурский и др.), произошло усиление эдификаторных функций подлеска (лещина, чубушник, калина буреинская). Лесоводственно-таксационные показатели древостоя при этом изменились мало.
3. Трансформация роли древесно-кустарниковых ярусов, произошедшая к 2018 г., обусловила серьезную перестройку парцеллярной структуры “лианового” фитоценоза, указывая на продвинутость лесообразовательного процесса:
а – за период исследований было выявлено 16 парцелл; в 1998 г. – 12 (6 условно-коренных и 6 производных), в 2018 г. – 10 (7 и 3 соответственно), общими остались 6 парцелл, все они условно-коренные, в 2018 г. к ним добавилась еще одна;
б – произошло закономерное укрупнение парцелл и выравнивание их границ;
в – число основных – преобладающих по площади – парцелл возросло с 5 до 7, площадь увеличилась с 77.5 до 88.8% от площади фитоценоза;
в – все условно-коренные парцеллы в 2018 г. стали основными, занимая от 9.3 до 18.7% площади фитоценоза;
г – наиболее значимыми трансформациями в парцеллярной структуре следует считать исчезновение самой большой производной парцеллы – “разнотравной с осокой” (“растворилась” в смежных парцеллах), сильное увеличение площади типичной для хвойно-широколиственных лесов “хвощовой” парцеллы и смену “волжанковой” парцеллы “лещиновой редкопокровной”.
4. Формирование парцеллярной структуры лесных сообществ на северных склонах в Южном Приморье определяется в первую очередь экотопическими условиями (микрорельеф поверхности и влажность почв), во вторую – ценотическими факторами (ограничение эдификаторами сообщества доступа света к поверхности). Связь между видовым составом древесно-кустарниковых ярусов и размещением парцелл отсутствует.
5. Травяной ярус – самый динамичный компонент фитоценоза является надежным индикатором трансформации внутриценотических условий. В ответ на повышение сомкнутости древесно-кустарниковых ярусов площадь парцелл с густопокровными микрогруппировками (проективное покрытие >60%) за 20 лет уменьшилась в 5 раз; сократилось число парцелл с светолюбивыми видами (волжанка двудомная, осока возвратившаяся, диспорум зеленеющий (Disporum viridescens), лесной мак весенний, звездчатка Бунге и др.) и увеличилось – с теневыносливыми и влаголюбивыми (хвощ зимующий, адиантум стоповидный (Adiantum pedatum), осока кривоносая, стеблелист мощный (Caulophyllum robustum) и др.).
6. От ранее произраставшего коренного чернопихтово-широколиственного типа леса широколиственно-липовый отличается только отсутствием пихты цельнолистной и кедра корейского, что объясняется воздействием периодических низовых пожаров и удаленностью источников обсеменения. Для окончательного восстановления коренного типа леса необходимы профилактика пожаров и меры по содействию естественному возобновлению, которые следует планировать с учетом парцеллярной структуры фитоценоза.
Список литературы
Васильев Н.Г., Колесников Б.П. Чернопихтово-широколиственные леса Южного Приморья. М.–Л.: АН СССР, 1962. 147 с.
Возмищева А.С., Перепёлкина П.А. Влияние синузий травяно-кустарникового яруса на параметры подроста в северных и южных широколиственно-кедровых лесах // Современные проблемы науки и образования. 2015. № 6. [https://www.science-education.ru/ru/article/ view?id=23338]
Дуденко Г.А. Эколого-ценотическая структура вторичных лесов южного Приморья: автореф. дис. … канд. биол. наук: 06.03.02. Уссурийск: ФГБОУ ВПО “ПГСХА”, 2012. 23 с.
Дылис Н.В. Структура лесного биогеоценоза. М.: Наука, 1969. 55 с.
Дылис Н.В. Структурно-функциональная организация биогеоценотических систем и ее изучение // Программа и методика биогеоценологических исследований. М.: Наука, 1974. С. 14–23.
Иванов А.В., Иванова Е.В., Гамаева С.В. Изменение разнообразия хвойно-широколиственных лесов Южного Приморья в связи с рубками и пожарами // Экология. 2022. № 2. С. 1–8
Иванова И.Т., Ярошенко П.Д., Берстюкова К.П. Микрофитоценозы некоторых сообществ хвойно-широколиственных лесов Приморья // Комаровские чтения. Владивосток: ДВФ СО АН СССР, 1963. Вып. XI. С. 58–85.
Исаев А.С., Носова Л.М., Пузаченко Ю.Г. Биологическое разнообразие лесов России – предложения к программе действий // Лесоведение. 1997. № 2. С. 3–13.
Ковалев А.П., Алексеенко А.Ю., Лашина Е.В., Качанова Т.Г. О необходимости смены приоритетов при рубках в производных кедрово-широколиственных лесах Дальнего Востока // Сибирский лесной журн. 2020. № 2. С. 3–13.
Колесников Б.П. Растительность // Южная часть Дальнего Востока. М.: Наука, 1969. С. 206–250.
Комарова Т.А. Изменение синузиальной структуры кустарничково-травяного яруса в ходе послепожарных сукцессий // Ботанический журн. 1993. Т. 78. № 6. С. 86–95.
Комарова Т.А. О горизонтальной неоднородности пихтово-еловых мелкотравно-папоротниковых лесов северного Сихотэ-Алиня // Экология. 1978. № 1. С. 42–49.
Корякин В.Н., Челышев В.А. Современная характеристика лесов // Современное состояние лесов Российского Дальнего Востока и перспективы их использования. Хабаровск: ДальНИИЛХ, 2009. С. 15–34.
Крупянко Н.А. Синузиальная структура травяного яруса кедровников Уссурийского заповедника // Ботанический журн. 1983. Т. 68. № 10. С. 1385–1392.
Кудинов А.И. Дубово-кедровые леса Южного Приморья и их динамика. Уссурийск: ПГСХА, 2000. 182 с.
Лукина Н.В., Гераськина А.П., Горнов А.В., Шевченко Н.Е., Куприн А.В., Чернов Т.И., Чумаченко С.И., Шанин В.Н., Кузнецова А.И., Тебенькова Д.Н., Горнова М.В. Биоразнообразие и климаторегулирующие функции лесов: актуальные вопросы и перспективы исследований // Вопросы лесной науки. Т. 3. № 4. 2020. С. 1–90.
Максимова В.Ф. Элементы мозаики травяного яруса в кедрово-широколиственных лесах Среднего Сихотэ-Алиня // Кедрово-широколиственные леса Дальнего Востока (биогеоценотический аспект). Владивосток: ДВО АН СССР, 1987. С. 79–85.
Москалюк Т.А. Трансформация травяного яруса за 20 лет восстановительной сукцессии в широколиственных лесах с лиановой растительностью на юге Приморского края // Комаровские чтения, 2021. Вып. LXIX. С. 26–78.
Москалюк Т.А. Ценотическая структура и мониторинг лесов Дальнего Востока // Растения в муссонном климате: Материалы IV Междунар. конф. Владивосток, 2006. С. 78–82.
Москалюк Т.А., Тарасова И.С. Структура травяного яруса в широколиственных лесах Южного Приморья в летний период // Животный и растительный мир Дальнего Востока. Владивосток: ДВФУ, 2015. Вып. 24. С. 37–54.
Назимова Д.И., Коновалова М.Е., Данилина Д.М., Пономарев Е.И., Сташкевич Н.Ю., Бабой С.Д. Исследования долговременной динамики лесов в пергумидном климате Западного Саяна (Ермаковский стационар Института леса им. В.Н. Сукачева СО РАН) // Сибирский лесной журн. 2015. № 4. С. 3–17.
ОСТ 56-69-83. Площади пробные лесоустроительные. Метод закладки. М.: Государственный комитет СССР по лесному хозяйству, 1983. 60 с.
Петропавловский Б.С. Леса Приморского края: (Эколого-географический анализ). Владивосток: Дальнаука, 2004. 317 с.
Распределение площади лесов и запасов древесины по преобладающим породам и группам возраста на 01.01.2021. Форма № 2-ГЛР // Информация о порядке предоставления сведений из гос. лесного реестра Приморского края [https://primorsky.ru/authorities/executive-agencies/departments/forestry/folder2/]
Рубцов М.В., Рыбакова Н.А. Динамика парцеллярной структуры лесных фитоценозов в процессе восстановления популяции ели в южнотаежных березняках // Лесоведение. 2016. № 5. С. 323–331.
Сибирина Л.А. Риск потери местообитаний кедрово-широколиственных лесов при нерациональном лесопользовании // Природа без границ: Мат. VIII Междунар. эколог. форума (23–24 октября 2014 г., Владивосток). Владивосток: ДВФУ, 2014. С. 404–406.
Сосудистые растения советского Дальнего Востока: в 8 т. / Под ред. С.С. Харкевича. СПб. (Л.).: Наука, 1985–2006. Т. 1–8.
Справочник для таксации лесов Дальнего Востока / Отв. сост. В. Н. Корякин. Хабаровск: ДальНИИЛХ, 1990. 526 с.
Стратегия и План действий по сохранению биологического разнообразия Российской Федерации. М.: Министерство природных ресурсов и экологии Российской Федерации, 2014. 256 с. [https://www.cbd.int›doc › world › ru-nbsap-v2-ru].
Сукачев В.Н., Зонн С.В. Общие принципы и программа изучения типов леса // Методические указания к изучению типов леса. М.: АН СССР, 1961. С. 1–104.
Тебенькова Д.Н., Лукина Н.В., Чумаченко С.И., Данилова М.А., Кузнецова А.И., Горнов А.В., Шевченко Н.Е., Катаев А.Д., Гагарин Ю.Н. Мультифункциональность и биоразнообразие лесных экосистем // Лесоведение. 2019. № 5. С. 341–356.
Усольцев В.А. Биоразнообразие в экосистемах: краткий обзор проблемы // Эко-Потенциал. 2019. № 1(25). Биология. С. 9–47.
Уткин А.И. Методика исследований первичной продуктивности лесов // Биологическая продуктивность лесов Поволжья. М.: Наука, 1982. С. 59–71.
Caspersen J.P., Pacala S.W. Successional diversity and forest ecosystem function // Ecological Research. 2001. V. 16. № 5. P. 895–903.
Elliot T., Goldstein B., Gómez-Baggethun E., Proença V., Rugani B. Ecosystem service deficits of European cities // Science of The Total Environment. 2022. V. 837: 155 875 [https://www.sciencedirect.com/science/article/abs/pii/ S0048969722029722].
Gao T., Hedblom M., Emilsson T., Nielsen A.B. The role of forest stand structure as biodiversity indicator // Forest Ecology and Management. 2014. V. 330. P. 82–93.
Hector A., Bagchi R. Biodiversity and ecosystem multifunctionality // Nature. 2007. V. 448. № 7150. P. 188–190.
Keenan R., Reams G., Achard F., De Freitas J., Grainger A., Lindquist E. Dynamics of global forest area: results from the FAO Global Forest Resources Assessment 2015 // Forest Ecology and Management. Science to Sustain the World’s Forests, 2015. V. 352. p. 9–20.
Mestre L., Toro-Manríquez M., Soler R., Huertas-Herrera A., Martínez-Pastur G., Lencinas M.V. The influence of canopy-layer composition on understory plant diversity in southern temperate forests // Forest Ecosystems. 2017. № 4. Iss. 6. 13 p. [https://forestecosyst.springeropen.com/articles/10.1186/s40663-017-0093-z]
Spicer M.E., Mellor H., Carson W.P. Seeing beyond the trees: a comparison of tropical and temperate plant growth forms and their vertical distribution // Ecology. 2020. V. 101. №. 4. P. 1–9.
UNSPF: United Nations strategic plan for forests 2017–2030 and quadrennial programme of work of the United Nations Forum on Forests for the period 2017–2020: Resolution adopted by the Economic and Social Council on 20 April 2017. 24 p. Distrib. General 07.2017. [https:// www.iucn.org/sites/dev/files/content/documents/strategic_ plan_2017_2030.pdf]
Дополнительные материалы отсутствуют.