

Лесоведение, 2023, № 3, стр. 255-268
Площадь питания и рост деревьев в средневозрастных культурах сосны
М. В. Рогозин *
Пермский государственный национальный исследовательский университет
614990 Пермь, ул. Генкеля, д. 4, Россия
* E-mail: rog-mikhail@yandex.ru
Поступила в редакцию 05.02.2021
После доработки 18.02.2022
Принята к публикации 07.06.2022
- EDN: PVGIFC
- DOI: 10.31857/S0024114823030087
Аннотация
Изучены культуры 1Б класса бонитета полнотой 0.95 на площади 0.64 га, созданные на вырубке по схеме 1.82 × 1.10 м. На план наносили живые и отпавшие деревья и площади их питания в программе “ArcMap-ArcView”. Территорию разделили на 9 секций, с густотой 76–122% от среднего по насаждению. Доставшаяся деревьям в возрасте 30 лет площадь питания повлияла на диаметр ствола в 55 лет в редких местах древостоя с силой 13.3%, а в густых местах – с силой 5.0%. То есть подтвердилась гипотеза, что влияние площади питания дерева на диаметр ствола может быть слабым также и в среднем возрасте насаждений, и уровень его влияния зависит от густоты древостоя. Отпад деревьев к 55 годам коррелировал с частотой в классах площади питания (r = 0.96 ± 0.03), поэтому площадь питания менее среднего значения повышала вероятность отпада дерева к 55 годам всего лишь на 7%. Выдвинуто предположение, что снижение влияния площади питания при высокой густоте происходит из-за усиления кооперации деревьев. При имитационном разреживании культур, с увеличением площади питания у оставляемых деревьев в 2 раза было получено соответствующее увеличение диаметра только у 11% деревьев. Остальные 89% деревьев не воспользовались доставшейся им большей площадью питания и не увеличили свои размеры, несмотря на 25 лет развития при более свободном стоянии. Это указывает на то, что в культурах второго класса возраста увеличение площади питания деревьев уже не приводит к улучшению их развития в подавляющем числе случаев. Поэтому густоту следует снижать в намного более раннем возрасте, например, в 10–15 лет.
К настоящему времени получили развитие исследования, где рост деревьев изучают в многофакторных пространственных моделях (Стороженко, 2007; Колобов, 2014; Грабарник, Секретенко, 2015) с анализом волновых процессов смены поколений и структуры насаждений на фоне внешних и внутренних факторов (Гавриков, 2013; Усольцев, 2013), а также взаимодействие деревьев в биогруппах и микроценозах (Вайс, 2014; Рогозин, 2019). Также были начаты, но, к сожалению, не продолжены исследования еженедельной динамики прироста у соседствующих деревьев в девственных лесах (Горячев, 1999). Эти исследования в конечном счете формируют теоретические основы современного лесоводства, в котором должны соединиться “теория лесообразовательного процесса”, “популяционная биология” и “теоретическая экология”, частью которой является и недавно возникшая “лесная биогеофизика” (Тихонова, 2020).
В качестве факторов влияния на рост деревьев необходимо использовать независимые переменные величины. К их числу относится и площадь питания дерева. Она выступает в качестве внутреннего фактора, и лесовод может ее регулировать в течение всей жизни древостоя (Правила …, 2017). Так, средняя площадь питания дерева влияет на средний диаметр древостоя сосны с силой 88–94% (Нагимов, 2000). Однако касательно влияния индивидуальной площади питания дерева и густоты микроценоза дело обстоит иначе.
Поясним, что расчеты “средней” и “индивидуальной” площади питания дерева сильно отличаются. Первую рассчитывают делением площади на число растений на ней (в лесоводстве чаще используют обратную ей величину – густоту стояния деревьев на 1 га), и она отражает действие густоты на уровне всего фитоценоза (Рогозин, 2019). Исследований по этой теме достаточно, в особенности для лесных культур (Прокопьев, 1981; Плантационное …, 2007; Терехов, Усольцев, 2008; Мерзленко, Бабич, 2011), тогда как для “индивидуальной” площади питания, действующей на уровне микроценоза, исследований немного, а методы различны (Горячев, 1999; Борисов и др. 2014; Грабарник, Секретенко, 2015).
Разбор этих методов показал (Рогозин, 2019), что использование зависимых от размеров дерева так называемых “пропорциональных расстояний” приводит к завышенной оценке влияния площади питания дерева. Эта завышенная оценка была основанием для внедрения интенсивных рубок ухода, действующих и поныне (Правила …, 2017). Они основаны на господствовавшей весь ХХ в. идее тотальной конкуренции в растительных сообществах. Однако теории таких рубок подобны уже неким фантомам (Рогозин, 2017) и вполне отвечают понятию “нарратива” (Карабаева, 2003), т.е. объяснению явлений таким образом, что их содержание выстраивается в зависимости от задуманного автором финала (для рубок ухода это получение ликвидной древесины). Такой нарративный финал имела знаковая для своего времени книга “Рубки ухода за лесом в новом освещении” Г.Р. Эйтингена (1934). Однако ныне эти идеи уже не отвечают реальному ходу роста насаждений, где действуют не только закон естественного изреживания (единственный закон, на котором основаны Правила ухода за лесом), но и череда других законов: закон развития древостоев в зависимости от начальной густоты Г.С. Разина (1979), ранговый закон роста деревьев Е.Л. Маслакова (1984) и выдвинутые недавно (Рогозин, 2019) новые законы – закон генетического гомостаза частот правых и левых форм деревьев А.М. Голикова (Голиков, 2011, 2014), закон неравномерного размещения деревьев и закон ослабления конкуренции в микроценозах.
Свидетельства ослабления конкуренции в микроценозах есть в работе В.А. Усольцева с соавт. (2018), который на 20 пробных площадях в сосняках в возрасте 20–40 лет определил индексы конкуренции для центрального дерева и соседей, полученные 10 способами. Эти индексы повлияли на фитомассу и прирост дерева в естественных молодняках соответственно на 5 и 11%, а в 20-летних культурах – лишь на 0.2 и 3%. При этом на радиальный прирост в естественных насаждениях индексы конкуренции воздействовали с силой 36%, а в культурах – с силой 11% (Усольцев и др., 2018).
Вопросы конкуренции мы изучали ранее в массиве сосны в возрасте 184 лет, где влияние соседей на рост центрального дерева исследовали четырьмя методами (Рогозин, 2019). Выяснилось, что густота микроценозов размером 0.01 га в возрасте 120 лет повлияла на диаметр деревьев в возрасте 184 года с силой 10.1%, а площадь питания дерева – с силой 7.7%. Другие методы показали еще меньшие значения, и поэтому конкурентное давление на дерево со стороны соседей можно считать слабым. На основании этих данных выдвинуто положение, что в микроценозах сосны к спелому возрасту внутривидовая конкуренция ослабевает и меняется на взаимное партнерство. Последнее выражается в корреляции диаметров у соседствующих деревьев, которая имеет место до расстояния 4.5 м, и влияние такого партнерства оказалась до 10 раз сильнее конкуренции (Рогозин, 2019).
Принято считать, что конкуренция в насаждении наиболее высока при максимуме прироста в возрасте 40–50 лет (Анучин, 1982). Чрезвычайно сильны в этом возрасте также дифференциация и отпад деревьев (Чирков, 2004). Однако в моделях развития древостоев важна количественная оценка конкуренции. Для этого иногда используют оценки на основе густоты, подразумевая, что чем она больше, тем выше отпад. Однако здесь происходит незаметная подмена смыслов – давление конкуренции отождествляют с уровнем густоты либо с отпадом деревьев. Между густотой, отпадом и давлением конкуренции связь, конечно, существует, но эти явления разные. И тут возникает вопрос, в каких единицах надлежит измерять силу конкуренции и как оценивать ее действие?
Так, используемые “индексы конкуренции” дерева (Усольцев, 2013; Усольцев и др., 2018) не отражают какое-либо новое явление и содержат известные показатели – диаметр ствола, высоту дерева, размеры его кроны при их отношении к площади питания дерева или к расстояниям до соседствующих деревьев. Их переводят в относительные величины, т.е. измеряют по отношению к среднему значению, которое принимают за единицу, называя полученное значение “индексом”. Однако для измерения давления конкуренции нужно выделить долю (обусловленность или силу) этого давления на ростовые или физиологические показатели. В биологии априорно считается (Костерин, 2007; Баландин, 2010), что растения в сообществах взаимодействуют преимущественно как конкуренты за ресурсы и поэтому “давят” друг на друга тем сильнее, чем ближе между ними расстояние и больше их размеры.
Но между деревьями есть и явление партнерства, которое выражается, в частности, в виде разнесения на 1–2 недели максимумов прироста у деревьев-соседей во время вегетации, открытое В.М. Горячевым (1999). То есть деревья “приспосабливаются” друг к другу, вероятно, эпигенетически и “выбирают” для себя разное время отбора ресурсов питания, занимая разные экологические ниши во времени. Явление партнерства ослабляет конкуренцию и вполне объясняет успешный рост до 40–57% (!) деревьев в плотных биогруппах в молодняках (Маслаков, 1984; Марченко, 1995), в среднем возрасте (Ипатов, Тархова, 1975; Маслаков, 1999) и спелых насаждениях (Сеннов, 1999; Рогозин, 2019).
Явление, открытое В.М. Горячевым (1999), почти неизвестно лесоводам; однако оно имеет колоссальную важность и выступает как одно из главных положений в новой парадигме лесоведения (Рогозин, 2019). В насаждении мы всегда наблюдаем действие конкуренции и партнерства вместе. Не зная, как их разделять, причем желательно делать это быстрее, чем, например, при способе В.М. Горячева (измеряя прирост у деревьев каждую неделю во время вегетации), различия между размерами деревьев априорно приписывают действию конкуренции, а то, что она не может объяснить появление плотных биогрупп, относят к действию микроусловий (даже не пытаясь выяснить силу их влияния количественно).
На наш взгляд, в связи с неясностью единиц, в которых надлежит измерять собственно конкуренцию, следует отказаться от термина “индекс конкуренции”, в котором простые показатели роста дерева при их отношении к его пространственному размещению заменены абстрактными словами с иным смыслом, которого не было в исходных показателях. По-видимому, их введение было вызвано стремлением упростить многословные термины. В результате сложное явление было редуцировано до одной только конкуренции.
Все сказанное выше относится к “световой” конкуренции. И лишь немногие авторы пытаются выяснить влияние “корневой” конкуренции (Терехов, Усольцев, 2008, 2010; Санников, Санникова, 2014). Здесь возникают чисто технические трудности, ограничивающие объем выборок, что не позволяет пока рассчитать количественно влияние фактора корненасыщенности на рост отдельных деревьев в пределах их “световой” площади питания. При этом слабо изучается и давно известное, но совершенно парадоксальное явление – срастание корней деревьев, которое наблюдается иногда у 60% деревьев сосны и которое свидетельствует о явной кооперации деревьев (Демаков, 2018; Усольцев и др., 2018). При этом остается неизвестным, в каких лесорастительных зонах и типах леса срастание корней усиливается, а где его можно не заметить и по-прежнему априорно считать конкуренцию корневых систем доминантой во взаимоотношениях между особями как одного, так и разных видов деревьев. Эти вопросы представляют собой малоизученную проблему, однако и по вопросам “световой” конкуренции остается много неясностей.
Таким образом, по итогам обзора литературы можно сформулировать общую сверхзадачу – необходимо исследовать соотношение конкуренции и сотрудничества деревьев в период высокой полноты в среднем возрасте насаждений, для которого данных недостаточно. Конкуренцию можно изучать с использованием нескольких показателей, например, площадей питания, давления соседей на центральное дерево в виде суммы их диаметров, давления с учетом расстояний до них, суммарное давление соседей на единицу площади питания дерева в центре и т.д. Среди этих, а также более сложных методов с использованием прироста лесоводам пока не удалось выявить лучшие из них из-за слабых связей с продукционными показателями и недостаточности выборок (Усольцев, 2013). Поэтому следует использовать максимальные выборки и вначале простые методы, после чего переходить к более сложным. Для данной работы мы взяли показатели площади питания и диаметры стволов деревьев. Они будут отражать влияние фактора густоты на уровне микроценоза на размер дерева в его центре. В следующих статьях и монографии мы используем другие вышеперечисленные показатели конкуренции.
Так как большая часть исследователей считает конкуренцию зависимой от плотности ценоза, то следует выбрать древостои с разной густотой, где желательно изучить также отпад деревьев. Для этого вполне подходят культуры сосны с густотой посадки около 5 тыс. шт. га–1, в которых сильное естественное изреживание начиналось в возрасте 30–35 лет при полноте 1.2–1.3, которая затем неизбежно снижалась (Прокопьев, 1981; Рогозин, Разин, 2015).
С учетом этих соображений из общей сверхзадачи и конкретно для данной работы можно выдвинуть гипотезу о том, что влияние площади питания дерева на диаметр его ствола в среднем возрасте насаждений может быть слабым и будет зависеть от густоты древостоя.
В связи с этим целью работы было изучение влияния индивидуальной площади питания деревьев на их размеры и отпад в 55-летних культурах сосны на участках с разной густотой. Для среднего возраста насаждений эти данные будут получены впервые с достаточной статистической точностью.
ОБЪЕКТЫ И МЕТОДИКА
Исследовали рядовые культуры сосны обыкновенной (Рinus sylvestris L.) в возрасте 55 лет, состав 10С, созданные в 1967 г. на вырубке 1950-х годов в кв. 73 бывшего Нижне-Курьинского лесничества Закамского лесхоза (ныне леса Пермского городского лесничества). Территория предназначалась под строительство, и на ней провели раскорчевку, однако строительство отменили и на площади 3.2 га создали культуры сосны посадкой по схеме 1.82 × 1.10 м (5.0 тыс. шт. га–1). В них мы выбрали территорию без больших прогалин и заложили пробную площадь размером 80 × 80 м (0.64 га), где колебания густоты были вызваны в основном пропусками мест в рядах из-за остатков корчевки, а также разной приживаемостью культур. Местоположение участка относительно ровное, с перепадом высот от его центра на восток и запад в пределах 1.0–1.5 м. В первой половине ХIХ в. леса здесь были пройдены рубками (1830–1840 гг.), чтобы получить древесный уголь для заводов г. Перми, и поэтому повсеместно встречаются характерные следы в виде кольцевых канав в местах выжигания угля. Насаждение расположено на надпойменной террасе р. Камы, почва супесчаная, тип леса – сосняк кисличниковый.
Методика работ включала в себя картирование живых и отпавших деревьев по опорным и промежуточным визирам и прямоугольным координатам с итоговой точностью нанесения на план соседствующих деревьев в пределах ±5–10 см, с определением диаметра ствола с точностью ±0.1 см через его окружность на высоте 1.3 м. Перед этим у дерева в 2019 г., осенью, подрумянивали грубую корку, наносили номер и отмечали место измерения окружности ствола для повторных измерений в последующие годы с целью определения текущего прироста. План оцифровали в программе “ArcMap-ArcView” и пробную площадь виртуально разбили на 9 секций по 0.071 га для изучения флуктуаций густоты. Такие размеры секций обеспечивали получение выборки с числом деревьев 70–100 шт., которые образовывали минимальный по численности макроценоз. Методика построения полигонов питания вокруг каждого дерева известна (Мартынов, 1976; Тябера, 1978). В компьютерном варианте (Рогозин, 2019) ее особенность состояла в том, что расстояния до ближних 4–8 соседей делили пополам и проводили через эти точки перпендикулярные линии; их соединяли и получали полигон питания. Считалось, что все деревья в раннем возрасте имели равные шансы на использование доставшейся им площади питания. При выстраивании полигонов питания требовались соседи, поэтому использовали деревья и за границами секций.
Основные таксационные показатели определяли в два этапа. Вначале рассчитывали густоту по секциям и далее, ориентируясь на ее близкие значения, а также принимая во внимание рельеф территории, с целью определения средней высоты древостоя эти секции разделили на четыре неравные группы, в которых раздельно определяли высоты для построения затем графиков зависимости между высотой и диаметром ствола дерева. Всего было измерено 137 высот деревьев из всех ступеней толщины. Относительная полнота была определена для каждой из 9 секций размером по 0.071 га. Для ее точного определения вначале специально рассчитывали так называемую “верхнюю” высоту яруса по деревьям, имеющим диаметры выше среднего с целью определения высоты господствующей части древостоя, необходимой для входа в стандартную таблицу полноты и запасов насаждений (Разин, 1977; Рогозин, Разин, 2015). При рубках ухода в данных культурах убирали только усохшие деревья. Это позволило с учетом следов от старых пней ретроспективно определить площадь питания каждого дерева для возраста примерно 30 лет (далее – площадь питания).
Таксационные показатели все взаимосвязаны, и в качестве ведущих или “указывающих” на размер других используют лишь некоторые из них: возраст дерева, диаметр ствола, высоту дерева и размер кроны (Нагимов, 2000; Демаков, 2018). При этом последние два измеряют обычно всегда у модельных деревьев, выбирая их по частотам диаметра. Модели широко используют при изучении крон (Разин, 1979), корневых систем (Терехов, Усольцев, 2008; Санников, Санникова, 2014), площади листовой поверхности и фитомассы деревьев (Усольцев, 2013). Такие исследования требуют больших трудозатрат.
В биологическом плане диаметр ствола дерева вторичен, так как является следствием работы фотосинтезирующего аппарата. Тем не менее именно диаметр ствола занимает первые места в исследованиях роста деревьев (Маслаков, 1984; Кузьмичев, 2013). В данной работе мы также использовали диаметр ствола как основную характеристику развития дерева. Конечно, было бы желательно изучить также и текущие приросты деревьев. Однако этот углубленный анализ мы используем далее и уже достаточно скоро после повторных измерений диаметров у всех деревьев на пробной площади.
Анализ различий между показателями площади питания, диаметрами стволов и коэффициентами корреляций между ними проведен для уровня значимости t0.05 по принятым статистическим процедурам (Усольцев, 2013; Демаков, 2018).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Закладка пробной площади проведена в центре выдела культур, на удалении от прогалин и деревьев естественного возобновления. Расположение деревьев и полигонов питания на секциях пробной площади и её обрамлении показаны ниже (рис. 1).
Рис. 1.
Полигоны питания деревьев в 55-летних культурах сосны на пробной площади 0.64 га с ее обрамлением на общей площади 0.85 га.

На пробной площади учтено 858 живых и 238 отпавших деревьев (137 сухостойных и 101 старый пень). Из результатов таксации следует, что в возрасте 30 лет густота насаждения колебалась в пределах 72–127% от среднего значения, и к возрасту 55 лет ее колебания сократились до 76–122% (табл. 1).
Таблица 1.
Некоторые таксационные и иные показатели древостоя культур сосны в возрасте 30 и 55 лет на пробной площади размером 0.64 га и на 9 секциях по 0.071 га
| Показатели | Номера секций | Среднее | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | ||
| Число деревьев в возрасте 30 лет, шт. | 116 | 126 | 139 | 155 | 130 | 120 | 89 | 83 | 107 | 118.3 |
| Число деревьев в возрасте 55 лет, шт. | 85 | 96 | 105 | 111 | 106 | 95 | 69 | 72 | 81 | 91.1 |
| Густота в возрасте 55 лет, шт./га | 1195 | 1350 | 1477 | 1561 | 1491 | 1336 | 970 | 1013 | 1139 | 1281 |
| То же, % | 93 | 105 | 115 | 122 | 116 | 104 | 76 | 79 | 89 | 100 |
| Средний диаметр, см | 21.6 | 20.1 | 19.4 | 19.0 | 19.7 | 20.9 | 22.6 | 22.9 | 21.5 | 20.9 |
| То же, % | 103 | 97 | 93 | 91 | 94 | 100 | 108 | 110 | 103 | 100,0 |
| Высота средняя, м | 27.5 | 26.3 | 26 | 25.7 | 25.8 | 27.6 | 28 | 28.1 | 27.9 | 27.0 |
| Высота господствующая, м | 28.2 | 26.8 | 26.8 | 26.8 | 26.8 | 28.7 | 28.2 | 28.3 | 28.2 | 27.6 |
| Полнота, м2/га | 43.7 | 43.0 | 43.6 | 44.5 | 44.9 | 45.7 | 38.9 | 41.8 | 41.2 | 43.0 |
| Стандарт полноты, м2/га | 45.6 | 45 | 45 | 45 | 45 | 46 | 45.6 | 45.6 | 45.6 | 45.4 |
| Стандарт запаса, м3/га | 570 | 550 | 550 | 550 | 550 | 590 | 570 | 570 | 570 | 563 |
| Относительная полнота | 0.96 | 0.96 | 0.97 | 0.99 | 1.00 | 0.99 | 0.85 | 0.92 | 0.90 | 0.95 |
| Запас, м3 /га | 550 | 530 | 530 | 540 | 550 | 590 | 490 | 520 | 510 | 534 |
| Корреляция (r) между площадью питания и диаметром ствола | 0.31 | 0.25 | 0.21 | 0.14 | 0.15 | 0.25 | 0.34 | 0.45 | 0.27 | 0.26 |
| Коэффициент детерминации (r2) при переводе в % | 9.6 | 6.0 | 4.5 | 2.0 | 2.1 | 6.2 | 11.8 | 20.5 | 7.5 | 7.8 |
В целом на пробной площади средняя густота в 55 лет составила 1.32 тыс. шт. га–1, с колебаниями по секциям от 1.05 до 1.63 тыс. шт. га–1. Средние высоты изменялись от 25.7 до 28.1 м (размах 2.4 м), тогда как в господствующей части средние высоты колебались несколько меньше: от 26.8 до 28.7 м (размах 1.9 м). Несмотря на близкие значения последних высот, которые относительно среднего значения менялись от 97.1 до 104.0%, стандарты полноты и запаса из местных таблиц использованы разные. В результате относительная полнота и запас по секциям менялись соответственно от 0.85 до 1.00 и от 490 до 590 м3/га–1. В среднем относительная полнота равна 0.95 при запасе древесины 534 м3/га–1. Соотношение средних высот и возраста классифицирует древостои на всех секциях 1Б классом бонитета.
Колебания густоты в возрасте 30 лет по секциям вполне ожидаемо повлияли на средний диаметр ствола: показатель аппроксимации линии тренда по девяти точкам оценивает ее влияние равным R2 = 0.95; также достаточно сильно, при R2 = 0.83, она повлияла на корреляцию в секциях между площадью питания дерева и диаметром ствола (рис. 2).
Рис. 2.
Влияние густоты культур в возрасте 30 лет на средний диаметр древостоя в 55 лет и на корреляцию между площадью питания и диаметром ствола.
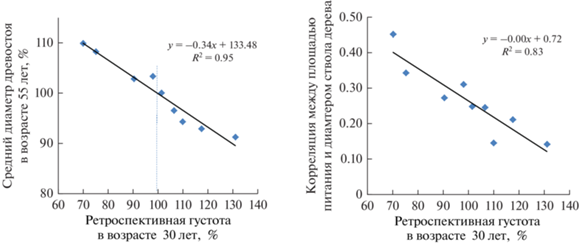
Подобное влияние густоты на диаметры деревьев отмечалось и у других исследователей (Разин, 1977; Нагимов, 1999; Вайс, 2014). Однако в соответствии с целью исследования нас прежде всего интересует, как воздействуют флуктуации густоты на аспект влияния индивидуальной площади питания. Для этого 9 секций разделили на две группы. Первую образовали в редких местах древостоя, где густота в 30-летнем возрасте была ниже средней и колебалась в пределах 70–98% от нее (секции 1, 7, 8, 9). Вторую сформировали из остальных пяти пробных площадей (секции 2, 3, 4, 5, 6), где густота была выше (101–131%). На рис. 2 слева эти группы разделены вертикальным пунктиром.
Выше в табл. 1 в редких местах культур (секции 1, 7, 8, 9) были приведены корреляции между площадью питания и диаметром дерева, которые колебались в пределах 0.14–0.45, и в густых местах они были наиболее слабыми (0.14–0.25). При возведении коэффициента корреляции в квадрат и переводе полученных долей единицы в проценты, можно получить коэффициент детерминации, который в среднем равен 7.8%, с колебаниями от 2.0 до 20.5%. Столь значительные колебания влияния густоты на изучаемую связь вынуждают провести далее анализ материала с разделением массива данных на две совокупности (рис. 3).
Рис. 3.
Влияние площади питания дерева в возрасте 30 лет на диаметр ствола в возрасте 55 лет в редких и густых местах древостоя.
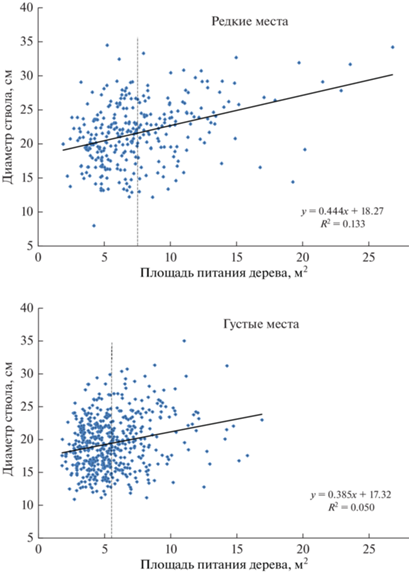
В редких и густых местах древостоя культур средние значения площади питания были равны соответственно 7.50 ± 0.21 м2 и 5.53 ± 0.11 м2 при среднеарифметическом диаметре ствола 21.6 ± ± 0.26 см и 19.3 ± 0.18 см. Различия между средними площадями питания и диаметрами составили 27 и 12% и были высоко достоверны: для площади питания t = 8.2 > t0.99 = 2.6 и для диаметра t = 7.2 > > t0.99 = 2.6.
Показатели аппроксимации R2 влияния площади питания на диаметр ствола в редких и густых местах были равны соответственно 0.133 и 0.050. С учетом объема выборок средний показатель аппроксимации для всего насаждения равен R2 = (0.133 × 308 + 0.050 × 518)/826 = 0.081 или 8.1%. Выше в табл. 1 это же влияние при оценке по коэффициенту детерминации соответствовало в среднем 7.8%. Полученные значения близки, и поэтому общую оценку влияния доставшейся дереву в 30-летнем возрасте площади питания на диаметр его ствола в возрасте 55 лет в этих культурах сосны можно считать в целом равной 8.1%.
Коэффициенты корреляции между площадью питания и диаметром ствола в редких и густых местах были равны соответственно r = 0.365 ± 0.049 и r = 0.224 ± 0.042. Различие достоверно при t = 2.17 > > t0.95 = 1.97. Это позволяет уверенно говорить о том, что влияние площади питания на диаметр ствола в густых местах достоверно слабеет; значения показателя R2 для трендов на рис. 3 также снижаются от 0.133 до 0.050 или в 2.7 раза.
В литературе (Усольцев, 2013; Усольцев и др., 2018) есть сведения о слабом (0.2–3.0%) влиянии площади питания на размер деревьев по индексам конкуренции, наблюдаемом в более густых культурах сосны во втором классе возраста, в их сравнении с естественными молодняками, где это влияние несколько усиливалось (до 5–11%). Полученные нами слабые влияния совпадают по уровню с упомянутыми данными В.А. Усольцева, характерными для 20–40-летнего возраста, что позволяет, с рядом оговорок на разные условия, начать выстраивать некоторую теоретически возможную динамику влияния площади питания на размеры деревьев в более старших возрастах (через 55-летний возраст наших культур). Далее такие же слабые влияния получены нами уже только в возрасте сосны 184 года (Рогозин, 2019), поэтому остаётся пробел в этой динамике для возраста сосняков 100–120 лет.
Обнаруженное ослабление влияния площади питания на размер деревьев в густых местах культур парадоксально для существующего ныне понимания внутривидовой конкуренции. Большинство биологов представляют ее вполне в духе идей Ч. Дарвина, как непрерывную “борьбу особей за существование” в сообществе себе подобных (Костерин, 2007; Баландин, 2010), и при таком ее понимании при увеличении плотности популяции конкуренция должна усиливаться и приводить к снижению размеров каждой особи. Именно так это и происходит в секциях (см. рис. 2), где на уровне древостоя деревья реагируют на повышение плотности их существования согласованным снижением размеров каждого его члена, и сила этой согласованной реакции на повышение общей густоты достигает 95%.
Однако на уровне отдельного дерева такого сильного влияния уже нет – здесь влияние густоты всего ценоза “рассыпается” на множество индивидуальных взаимодействий между соседствующими деревьями. Частями этого взаимодействия являются конкуренция и кооперация в освоении ресурсов питания, о чем уже говорилось, как о разделении экологических ниш питания во времени – явлении, обнаруженном В.М. Горячевым (1999). В этом свете совершенно по-иному можно интерпретировать резкое ослабление (в 2.7 раза) влияния площади питания на диаметр ствола в густых местах, здесь обнаруженное. Такое ослабление можно считать свидетельством возможного усиления положительного взаимодействия деревьев, которое проявляет себя, в том числе через образование биогрупп, в которых до перестойного возраста существуют 40% деревьев без существенного снижения их размеров (Рогозин, 2019).
Результатом конкуренции является также отпад деревьев и его зависимость от доставшейся дереву площади питания. Всего на девяти секциях нами учтено 238 отпавших деревьев. Были построены ряды распределения отпавших и живых деревьев по площади их питания с градуировкой между классами 1 м2 (рис. 4).
Рис. 4.
Площадь питания в возрасте 30 лет у всех учтенных для этого возраста деревьев и деревьев, отпавших к возрасту 55 лет. Вертикальные линии – средние значения.
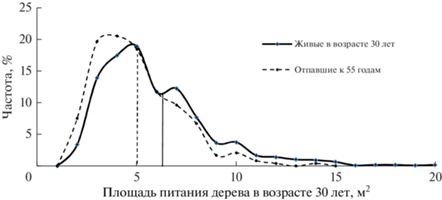
Вполне ожидаемо, что средняя площадь питания у отпавших деревьев была достоверно меньше (5.01 ± 0.20 м2), чем у деревьев, учтенных как бывшие живыми в возрасте 30 лет (для этого мы сложили вместе живые и отпавшие деревья), у которых площадь питания была определена равной 6.29 ± 0.10 м2. Сравнение рядов распределения показало, что в классе с наименьшей площадью питания (2 м2) погибло 50% деревьев, однако он имеет малую численность и слабо влияет на общую картину отпада, где главную роль играют модальные классы. Это хорошо видно при сравнении частот площадей питания у живых и отпавших деревьев, тесно коррелирующих при r = 0.96 ± 0.03 (рис. 4).
То есть деревья погибали почти одинаково часто во всех классах площади питания. Если взять шесть модальных классов со значениями от 3 до 8 м2 (81% значений площади питания), то в них погибло 18–23% деревьев в каждом при площади питания 5, 6, 7 и 8 м2 и несколько больше – 26 и 32% деревьев – в классах 4 и 3 м2 (в среднем по всем шести классам по 23%). Доля отпавших в этих модальных классах деревьев составила 87% от всех погибших к 55 годам особей. Поэтому в картине естественного отпада они играют основную роль. Если же оценивать отпад дерева по принципу “отпад произошел из-за площади питания менее среднего значения”, то для этого случая отпад составил 57%, а остальные 43% деревьев отпали при площадях питания выше средних значений. То есть “обладание” площадью питания менее среднего значения в 30-летнем возрасте увеличивает вероятность отпада дерева к 55 годам всего лишь на 7% в сравнении со случаем, если бы он не зависел от площади питания и был бы по 50% в ту и другую строну от ее среднего значения.
Отсюда следует такой же парадоксальный для конкуренции вывод, как и выше для диаметра ствола, а именно, отпад дерева в однородных экологических условиях в очень малой степени определяется конкурентным давлением соседей, и деревья погибают при самом разном расстоянии между собой и при разных площадях питания.
Располагая большими совокупностями деревьев в разных по густоте местах ценоза (выборки 308 и 518 шт. деревьев), их можно использовать далее в качестве имитационных моделей. Идея имитации состоит в ответе на вопрос, как бы реагировали диаметры стволов в возрасте 55 лет на то, если бы в возрасте 30 лет скопления деревьев разредили. Посмотрим, что произойдет с диаметрами стволов, если площади питания возрастут примерно в два раза. Для этого каждую из совокупностей поделили на две части, точно по среднему значению площади питания в них. Результаты представлены в таблице (табл. 2).
Таблица 2.
Имитация рубок ухода по площади питания (S пит) в возрасте 30 лет и их возможный результат при оценке по диаметру стволов в 55-летних культурах сосны
| Показатели | Вся совокуп–ность | Выборки | Увеличение | ||
|---|---|---|---|---|---|
| S пит малая | S пит большая | в число раз | % | ||
| Редкие места культур (секции 1, 7, 8, 9) | |||||
| Число наблюдений, шт. | 308 | 192 | 116 | – | – |
| Средняя площадь питания (Sср), м2 | 7.55 | 5.4 | 11.25 | 2.083 | 108.3 |
| Средний диаметр ствола (Дср), см | 21.6 | 20.71 | 23.15 | 1.118 | 11.8 |
| Эффект для диаметра ствола от увеличения площади питания дерева (Дср/Sср) | – | – | – | 0.109 | 10.9 |
| Доля стволов с диаметром выше среднего, % | 48.0 | 39.9 | 61.6 | – | – |
| Увеличение доли стволов с диаметром выше (+) или ниже (–) 50%, % | –2.0 | –10.1 | 11.6 | – | – |
| Густые места культур (секции 2, 3, 4, 5, 6) | |||||
| Число наблюдений, шт. | 518 | 313 | 205 | – | – |
| Средняя площадь питания (Sср), м2 | 5.54 | 4.03 | 7.84 | 1.945 | 94.5 |
| Средний диаметр ствола (Дср), см | 19.3 | 18.6 | 20.41 | 1.097 | 9.7 |
| Эффект для диаметра ствола от увеличения площади питания дерева (Дср/Sср) | – | – | – | 0.103 | 10.3 |
| Доля стволов с диаметром выше среднего, % | 47.6 | 41.6 | 56.7 | – | – |
| Увеличение этой доли выше или ниже 50%, % | –2.4 | –8.4 | 6.7 | – | – |
Так, из 308 деревьев в редких местах культур отделили выборку из 192 деревьев с площадью питания менее среднего значения (менее 7.55 м2), обозначенную в табл. 2 как “Sпит малая”, со средним значением 5.4 м2. Остальные 116 деревьев образовали вторую выборку “Sпит большая”, где средняя площадь питания составила 11.25 м2, что в 2.083 раза или на 108.3% больше, чем в первой. Если бы диаметры во второй выборке увеличились на такую же величину, что и площадь питания (т.е. на 108.3%), то эффект для диаметра от увеличения площади питания дерева был бы 100%. Однако он увеличился только на 11.8%, и отношение увеличения диаметра к увеличению площади питания составило 11.8/108.3 = = 0.109 или 10.9%. Эта оценка близка к показателю аппроксимации (R2 = 0.133) для тренда на рис. 3. Близкий результат в табл. 2 дают и расчеты по доле стволов (11.6%), которые сформировали диаметры выше среднего в этой выборке.
В густых местах древостоя аналогичные расчеты показали, что в совокупности из 518 деревьев можно образовать выборку из 313 деревьев с площадью питания менее средней (менее 5.54 м2), со средним значением 4.03 м2. Остальные 205 деревьев образуют выборку с площадями питания выше среднего значения и со средней площадью питания 7.84 м2, что в 1.945 раза или на 94.5% больше, чем в первой выборке. Средний диаметр увеличился здесь в 1.097 раза или на 9.7%. Отношение увеличения диаметра ствола к увеличению площади питания составило 9.7/94.5 = 0.103 или 10.3%. Такой же результат дают и расчеты по доле стволов (10.3%), которые выросли с диаметрами выше среднего там, где выборка имела увеличенные в 2 раза площади питания. Последняя оценка оказалась выше показателя R2 = 0.050 для соответствующего тренда на рис. 3; однако влияние увеличения площади питания на возрастание диаметра ствола все равно осталось на уровне первых десяти процентов, т.е. весьма слабым.
Полученные выше при имитации разреживаний данные можно интерпретировать как “правильную” реакцию диаметра ствола на увеличение площади питания только у 10.3–11.6% (в среднем у 11%) деревьев. Остальные 89% деревьев не воспользовались доставшейся им большей площадью питания и не увеличили диаметр ствола.
В литературе есть данные о разреживании древостоев в длительных опытах с рубками ухода, начиная с возраста 40 лет (Сеннов, 1984, 1999). Они показали их неэффективность для повышения производительности насаждений, т.е. наша имитация таких рубок вполне подтверждается строгой практикой. Однако даже столь длительные опыты не поколебали идею интенсивных разреживаний в насаждениях среднего возраста (Правила …, 2017), хотя еще в 1980-е годы уже было доказано, что в соответствии с “ранговым законом роста деревьев в молодняках” Е.Л. Маслакова (1984) регуляция густоты необходима намного раньше – в первом классе возраста, что подтвердил и длительный опыт плантационного выращивания леса (Плантационное …, 2007). Что действительно может быть связано с конкурентным давлением деревьев-соседей в наших культурах, так это отпад деревьев средних размеров. В числе усыхающих деревьев их менее 10%, но их гибель без видимых причин всегда вызывает вопросы. Возможно, это и влияние крупных соседей, и особенности литологии в точке их локализации или иные, пока малоизученные факторы.
Таким образом, подтвердилась гипотеза о том, что влияние площади питания дерева на диаметр ствола может быть слабым также и в среднем возрасте насаждений, а уровень этого влияния зависит от густоты древостоя.
ЗАКЛЮЧЕНИЕ
Современные методы пространственного анализа позволили сформировать две большие совокупности из деревьев в местах с разной густотой культур. На участках с редким расположением деревьев, обусловленным, по-видимому, пропуском посадочных мест из-за наличия порубочных остатков, доставшаяся дереву примерно в 30-летнем возрасте, площадь питания повлияла на диаметр его ствола в возрасте 55 лет с силой 13.3%, а в более густых местах ее влияние снизилось до 5.0%. Такое парадоксальное снижение и в целом слабое влияние площади питания на размер дерева (в среднем 7.8%) можно объяснить следующим образом. Вероятно, помимо конкуренции существует и партнерство деревьев; при этом в течение вегетации наступление максимума прироста у дерева может меняться, и деревья “приспосабливаются” друг к другу разнесением во времени пиков фотосинтеза и, следовательно, отбора элементов питания и тем самым резкого ослабления конкуренции между соседствующими деревьями. Явление это впервые установил В.М. Горячев (1999), и оно во многом объясняет слабое влияние площади питания в других наших исследованиях, где деревья росли в плотных биогруппах вплоть до перестойного возраста. Однако идея “разделения экологических ниш во времени” принимается в том случае, если презумпция априорного доминирования конкуренции будет заменена на ее равенство, а возможно, и на подчинение явлению партнерства деревьев, которое изучено крайне недостаточно.
При имитационном разреживании культур с увеличением площади питания у деревьев почти в 2 раза было получено соответствующее “правильное” увеличение диаметра только у 11% деревьев. Остальные 89% деревьев не воспользовались доставшейся им большей площадью питания и не увеличили свои размеры, несмотря на 25 лет развития при более свободном стоянии. Это указывает на то, что во втором классе возраста увеличение площади питания деревьев уже не приводит к их лучшему развитию в подавляющем числе случаев. Поэтому густоту следует снижать в намного более раннем возрасте, например, в 10–15 лет.
Список литературы
Анучин H.H. Лесная таксация. М.: Лесная промышленность, 1982. 552 с.
Баландин Р.К. Анти-Дарвин. Миражи эволюции. М.: Яуза, Эксмо, 2010. 350 с.
Борисов А.Н., Иванов В.В., Екимов Е.В. Метод оценки пространственного распределения ресурса в экологической нише // Сибирский лесной журн. 2014. № 5. С. 113–121.
Вайс А.А. Научные основы оценки горизонтальной структуры древостоев для повышения их устойчивости и продуктивности (на примере насаждений Западной и Восточной Сибири): автореф. дис. …д-ра с.-х. наук: 06.03.02. Красноярск, 2014. 33 с.
Гавриков В.Л. Рост леса: уровни описания и моделирования. Красноярск: Сибирский Федеральный Университет, 2013.176 с.
Голиков А.М. Эколого–диссимметрийный и изоферментный анализ структуры модельных популяций сосны обыкновенной // Лесоведение. 2011. № 5. С. 46–51.
Голиков А.М. Эколого-диссимметрический подход в генетике и селекции видов хвойных. LAP LAMBERT Academic Publishing, 2014. 162 с.
Горячев В.М. Влияние пространственного размещения деревьев в сообществе на формирование годичного слоя древесины хвойных в южнотаежных лесах Урала // Экология. 1999. № 1. С. 9–19.
Грабарник П.Я., Секретенко О.П. Анализ горизонтальной структуры древостоев методами случайных точечных полей // Сибирский лесной журн. 2015. № 3. С. 32–44.
Демаков Ю.П. Структура и закономерности развития лесов республики Марий Эл. Йошкар-Ола: Поволжский государственный технологический университет, 2018. 432 с.
Ипатов В.С., Тархова Т.Н. Количественный анализ ценотических эффектов в размещении деревьев по территории // Ботанический журн. 1975. Т. 60. № 9. С. 1237–1250.
Карабаева А.Г. Нарратив в науке и образовании // Серия “Symposium”. Инновации и образование. Выпуск 29. Сборник материалов конференции. Санкт-Петербург: Санкт-Петербургское философское общество, 2003. С. 89–96.
Колобов А.Н. Моделирование пространственно-временной динамики древесных сообществ: индивидуально-ориентированный подход // Лесоведение. 2014. № 5. С. 72–82.
Костерин О.Э. Дарвинизм как частный случай “бритвы Оккама” // Вестник ВОГиС. 2007. Т. 11. № 2. С. 416–431.
Кузьмичев В.В. Закономерности динамики древостоев. Новосибирск: Наука, 2013. 208 с.
Мартынов А.Н. Зависимость биометрических показателей сосны от площади питания // Лесоведение. 1976. № 5. С. 85–88.
Марченко И.С. Биополе лесных экосистем. Брянск: БГИТА,1995. 188 с.
Маслаков Е.Л. Формирование сосновых молодняков. М.: Лесная промышленность, 1984. 168 с.
Маслаков Е.Л. Генезис и динамика социальных структур сосны в фазе индивидуального роста // Таежные леса на пороге XXI в. СПб.: Санкт-Петербургский научно-исследовательский институт лесного хозяйства, 1999. С. 42–51.
Мерзленко М.Д., Бабич Н.А. Теория и практика искусственного лесовосстановления: учебное пособие. Архангельск: САФУ, 2011. 239 с.
Нагимов З.Я. Закономерности роста и формирования надземной фитомассы сосновых древостоев: дис. … д-ра с.-х. наук: 06.03.02. Екатеринбург: УГЛА, 2000. 409 с.
Плантационное лесоводство / Под ред. И. В. Шутова. СПб.: СПбГПУ, 2007. 366 с.
Правила ухода за лесами. М.: МПР РФ. Приказ № 626 от 22.11.2017.
Прокопьев М.Н. Культуры сосны в таежной зоне. М.: Лесная промышленность, 1981.136 с.
Разин Г.С. Эскизы таблиц хода роста древостоев (сосны, ели, березы и осины) с полнотой 1.0 по бонитетам // Основные положения организации и развития лесного хозяйства Пермской области. Гослескомитет СССР. В/О “Леспроект”. Пермь, 1977. С. 437–455.
Разин Г.С. Динамика сомкнутости одноярусных древостоев // Лесоведение. 1979. № 1. С. 23–25.
Рогозин М.В. Фантомы теорий рубок ухода // Бюллетень науки и практики. 2017. № 4(17). С. 48–55. https://doi.org/10.5281/zenodo.546284
Рогозин М.В. Структура древостоев: конкуренция или партнерство? [Электронный ресурс]. Пермь: ПГНИУ. 2019. 223 с. http://www.psu.ru/nauka/elektronnye-publikatsii/monografii-123123/rogozin-m-v-struktura-drevostoev-konkurentsiya-ili-partnerstvo
Рогозин М.В., Разин Г.С. Развитие древостоев. Модели, законы, гипотезы [Электронный ресурс]: монография. Пермь: ПГНИУ, 2015. 277 с. URL: http://k.psu.ru/library/ node/299086; http://elibrary.ru/item.asp?id=24420793
Санников С.Н., Санникова Н.С. Лес как подземно-сомкнутая дендроценоэкосистема // Сибирский лесной журн. 2014. № 1. С. 25–34.
Сеннов С.Н. Уход за лесом: экологические основы. М.: Лесная промышленность, 1984. 127 с.
Сеннов С.Н. Итоги 60-летних наблюдений за естественной динамикой леса // СПб.: СПбНИИЛХ, 1999. 98 с.
Стороженко В.Г. Устойчивые лесные сообщества: (теория и эксперимент). М.: Институт лесоведения РАН, 2007. 190 с.
Терехов Г.Г, Усольцев В.А. Формирование, рост и биопродуктивность опытных культур ели сибирской на Урале: Исследование системы связей и закономерностей. Екатеринбург: УрО РАН, 2008. 215 с.
Терехов Г.Г., Усольцев В.А. Морфоструктура насаждений и корненасыщенность ризосферы культур ели сибирской и вторичного лиственного древостоя на Среднем Урале как характеристика их конкурентных отношений // Хвойные бореальной зоны. 2010. Т. XXVIII. № 3–4. С. 1–7.
Тихонова И.В. Четыре опоры к фундаменту современного устойчивого лесоводства // Лесные экосистемы: современные вызовы, состояние, продуктивность и устойчивость. Материалы междунар. научно-практ. конф., посвященной 90-летию Института леса НАН Беларуси (Гомель, 13–15 ноября 2020 г.). Гомель: Институт леса НАН Беларуси. 2020. С. 97–101.
Тябера А.П. Площадь роста дерева и ее определение аналитическим способом // Лесной журн. 1978, № 2. С. 12-16.
Усольцев В.А. Продукционные показатели и конкурентные отношения деревьев. Исследование зависимостей. Екатеринбург: УГЛТУ, 2013. 556 с.
Усольцев В.А., Часовских В.П., Акчурина Г.А., Осмирко А.А., Кох Е.В. Фитомасса деревьев в конкурентных условиях: исследование системных связей средствами информационных технологий. Екатеринбург: УГЛТУ, 2018. 526 с.
Чирков Г.В. Закономерности формирования древесного отпада в хвойных древостоях Ленинградской области: автореф. дис. … к. с.-х. наук: 06.03.02. СПб. 2004. 24 с.
Эйтинген Г.Р. Рубки ухода за лесом в новом освещении. М., 1934. 224 с.
Дополнительные материалы отсутствуют.