

Лесоведение, 2023, № 3, стр. 217-227
Плотность древесины и коры деревьев на климатических градиентах Евразии
В. А. Усольцев a, b, *, И. С. Цепордей a
a Ботанический сад УрО РАН
620144 Екатеринбург, ул. 8 Марта, д. 202а, Россия
b Уральский государственный лесотехнический университет
620100 Екатеринбург, ул. Сибирский тракт, д. 37, Россия
* E-mail: Usoltsev50@mail.ru
Поступила в редакцию 22.02.2022
После доработки 25.03.2022
Принята к публикации 07.06.2022
- EDN: PXLDUA
- DOI: 10.31857/S0024114823030117
Аннотация
В условиях изменяющегося климата возрастает биосферная роль лесного покрова и актуальность исследований углерододепонирующей способности мировых лесов. Эти исследования содержат оценку биологической продуктивности деревьев и древостоев, включающей не только фитомассу, но и базисную плотность (БП) древесины и коры стволов. В нашем исследовании разработаны аллометрические модели БП древесины и коры деревьев 9 лесообразующих древесных видов Северной Евразии, имеющие такие независимые переменные, как возраст дерева, диаметр ствола, а также среднюю температуру января и среднегодовые осадки. Применена структура модели смешанного типа, в которой принадлежность исходных данных к каждому из древесных видов кодируется набором фиктивных переменных. На основе принципа пространственно-временного замещения полученные закономерности изменения БП в пространственных климатических градиентах использованы для прогноза их изменения в темпоральных градиентах. Подтверждено действие закона лимитирующего фактора Либиха при прогнозировании БП в пространственных и темпоральных климатических градиентах. Выявленные закономерности изменения БП древесины и коры деревьев в градиентах температур и осадков полностью повторяют ранее установленные закономерности изменения фитомассы и чистой первичной продукции деревьев и древостоев Евразии в тех же градиентах. Это означает, что климатическая обусловленность биологической продуктивности имеет общий характер как для количественных, так и для квалиметрических показателей деревьев и древостоев.
В условиях изменяющегося климата возрастает биосферная роль лесного покрова и актуальность исследований углерододепонирующей способности мировых лесов. Сегодня формируются глобальные базы данных не только о фитомассе деревьев и древостоев (Falster et al., 2015; Kattge et al., 2020), но и о квалиметрических показателях фитомассы, в частности, базисной плотности (БП) стволовой древесины (Zanne et al., 2009; Usoltsev, 2020), составляющей основную часть надземной фитомассы. Эти базы данных предоставляют необходимую исходную информацию для глобальных прогнозов углерододепонирующей способности лесов. Показатель плотности древесины, используемый при преобразовании данных о запасе стволовой древесины в показатели фитомассы, является ключевым фактором, влияющим на точность оценки депонирования углерода (Fearnside, 1997; Усольцев, Цепордей, 2020). С другой стороны, изменения климата, в частности учащающиеся засухи в некоторых регионах, влияют на физиологические процессы, определяющие ксилогенез и БП древесины (Bouriaud et al., 2005; Vieira et al., 2020).
Закономерности изменения БП древесины были проанализированы в связи с географической широтой, типом леса, температурой и осадками (Howe, 1974; Полубояринов, 1976; Kellomäki, 1979; Мелехов и др., 2003; Swenson, Enquist, 2007; Wiemann, Williamson, 2002; St-Germain, Krause, 2008). Было показано (Zhang, Shi, 2003), что географическое положение является определяющим фактором роста деревьев. Положительная связь БП древесины сосны обыкновенной (Pinus sylvestris L.) с температурой была установлена разными авторами на территории Финляндии, но эта связь характеризовалась низкими коэффициентами детерминации (Mikola, 1950; Saikku, 1975; Kellomäki, 1979).
На глобальном уровне сделан анализ изменчивости базисной плотности древесины в широтном диапазоне от 52° с. ш. до экватора, и установлено увеличение плотности на 0.5% на каждый градус среднегодовой температуры и снижение на 0.02% на каждый сантиметр среднегодовых осадков (Wiemann, Williamson, 2002). Рассматриваемая отдельно среднегодовая температура оказалась лучшим предиктором базисной плотности в зоне умеренных лесов (в диапазоне от 3 до 22°C), объясняя 80% изменчивости искомого показателя, по сравнению со всем исследуемым температурным диапазоном с объясненной изменчивостью 62% или по сравнению с теплыми тропическими районами (>23°C) с объясненной изменчивостью 33%. Напротив, только уровень среднегодовых осадков был лучшим предиктором базисной плотности в теплых тропических регионах (62%) по сравнению со всем исследуемым температурным диапазоном (4%) (Wiemann, Williamson, 2002).
По результатам анализа более 4600 таксонов установлена положительная связь БП древесины покрытосеменных и голосеменных растений со среднегодовой температурой и максимальной месячной температурой. Отрицательная корреляция между БП древесины и осадками была обнаружена у покрытосеменных растений. Значительная отрицательная корреляция установлена между БП древесины и высотой над ур. м. (Swenson, Enquist, 2007).
Таким образом, или выполнялось исследование изменчивости БП под влиянием различных эндо- и экзогенных факторов для отдельных древесных видов в пределах некоторого региона, или имелись обобщения планетарного уровня для совокупности видов. При моделировании фитомассы деревьев и древостоев на трансконтинентальном уровне в последние годы показано действие закона лимитирующего фактора Либиха с применением принципа пространственно-временного замещения (Усольцев и др., 2021; Usoltsev et al., 2022). Насколько нам известно, аналогичные результаты исследований изменения БП древесины и коры в климатических градиентах Евразии в открытых источниках отсутствуют.
В нашей статье была поставлена цель ответить на следующие вопросы:
– действует ли закон лимитирующего фактора на трансконтинентальном уровне при моделировании изменений БП древесины и коры деревьев лесообразующих видов Евразии в связи с территориально обусловленными показателями температур и осадков;
– используя принцип пространственно-временнóго замещения, проверить возможность применения построенных моделей БП древесины и коры деревьев, чувствительных к территориальным градиентам температур и осадков, при прогнозировании изменений БП деревьев во времени;
– определить вклады независимых переменных построенных моделей в объяснение изменчивости БП древесины и коры деревьев;
– установить, реагирует ли БП древесины и коры деревьев на изменения температур и осадков аналогично реакции на них фитомассы деревьев и древостоев, одинаковой для всех видов, или же реакция БП на климатические изменения будет видоспецифичной и отличной от ранее установленных реакций фитомассы на климатические сдвиги.
Известно, что показатели БП варьируют не только вдоль по стволу, но и в поперечном сечении ствола (Полубояринов, 1976; Исаева, 1978; Fujimoto et al., 2008; Télles et al., 2011; Sousa et al., 2016; Billard et al., 2021). В настоящем исследовании мы уходим от анализа изменчивости БП вдоль и поперек ствола и ограничиваемся моделированием показателей БП, средних для всего ствола дерева.
ОБЪЕКТЫ И МЕТОДИКА
Для решения поставленных задач мы использовали авторскую базу эмпирических данных о БП лесообразующих пород Северной Евразии (Usoltsev, 2020). Из нее отобрано 3448 модельных деревьев, распределение которых по древесным видам (родам) представлено в табл. 1. Были отобраны наиболее многочисленные данные, которыми характеризовались основные лесообразующие виды, при условии наличия в числе измеренных характеристик деревьев полного набора дендрометрических и квалиметрических показателей. Деревья, не обладающие полным набором данных, в наших расчетах не участвовали. Упомянутая база данных содержит координаты пробных площадей, где были получены сведения о модельных деревьях. Используя эти координаты, мы определили соответствующие территориальные показатели средней температуры января и среднегодового количества осадков по соответствующим климатическим картам (World Weather Maps, 2007). Эти климатические карты были показаны в нашей предыдущей публикации (Usoltsev et al., 2022).
Таблица 1.
Статистики показателей 3448 модельных деревьев, включенных в регрессионный анализ
| Обозначение статистик | Анализируемые показатели | |||||
|---|---|---|---|---|---|---|
| A | d | DW | DB | T | PR | |
| Сосна обыкновенная и чёрная (Pinus sylvestris L., P. nigra subsp. Pallasiana (Lamb.) Holmboe) | ||||||
| Mean | 46 | 12.5 | 398.4 | 290.7 | –14 | 378 |
| Min | 5 | 1.1 | 254.0 | 264.3 | –25 | 317 |
| Max | 186 | 55.0 | 640.4 | 636.4 | –3 | 570 |
| SD | 33.5 | 9.5 | 52.1 | 61.9 | 3.8 | 87.9 |
| CV, % | 72.8 | 76.3 | 13.1 | 21.3 | –27.2 | 23.2 |
| n | 966 | 957 | 966 | 966 | 966 | 966 |
| Кедр сибирский и корейский (Pinus sibirica Du Tour., P. koraiensis S. et Z.) | ||||||
| Mean | 42 | 9.3 | 354.7 | 717.9 | –18 | 522 |
| Min | 15 | 1.5 | 245.8 | 298.7 | –19 | 500 |
| Max | 165 | 29.3 | 509.9 | 969.5 | –10 | 826 |
| SD | 37.5 | 6.7 | 51.1 | 214.4 | 2.3 | 82.4 |
| CV, % | 90.4 | 71.5 | 14.4 | 29.9 | –12.4 | 15.8 |
| n | 74 | 74 | 74 | 74 | 74 | 74 |
| Лиственница сибирская, Сукачева и Каяндера (Larix sibirica L., L. Sukaczewii N.Dyl., L. cajanderi Mayr.) | ||||||
| Mean | 81 | 13.7 | 511.7 | 335.8 | –26 | 401 |
| Min | 15 | 0.3 | 382.4 | 186.3 | –30 | 317 |
| Max | 400 | 52.8 | 735.5 | 545.3 | –15 | 444 |
| SD | 73.2 | 8.9 | 43.0 | 69.7 | 5.6 | 60.3 |
| CV, % | 90.5 | 64.7 | 8.4 | 20.8 | –21.5 | 15.1 |
| n | 193 | 194 | 194 | 194 | 194 | 194 |
| Ель европейская, сибирская и аянская (Picea abies (L.) H. Karst., P. obovata L., P. ajanensis (Lindl. еt Gord.) Fisch. еx Carr.) | ||||||
| Mean | 48 | 12.4 | 393.7 | 432.4 | –15 | 528 |
| Min | 11 | 1.0 | 255.0 | 176.7 | –17 | 500 |
| Max | 163 | 51.5 | 648.1 | 866.7 | 0 | 826 |
| SD | 32.6 | 10.2 | 64.1 | 125.3 | 4.1 | 58.7 |
| CV, % | 68.0 | 81.8 | 16.3 | 29.0 | –27.9 | 11.1 |
| n | 278 | 278 | 278 | 278 | 278 | 278 |
| Пихта белая, сибирская и цельнолистная (Abies alba Mill., A. sibirica L., А. holophуlla Maxim.) | ||||||
| Mean | 70 | 20.3 | 364.2 | 429.2 | –10 | 585 |
| Min | 8 | 1.6 | 272.8 | 299.1 | –17 | 500 |
| Max | 180 | 46.2 | 513.0 | 606.7 | –5 | 826 |
| SD | 40.2 | 11.0 | 41.3 | 71.1 | 5.5 | 107.7 |
| CV, % | 57.6 | 54.2 | 11.3 | 16.6 | –55.0 | 18.4 |
| n | 92 | 92 | 92 | 92 | 92 | 92 |
| Береза пушистая, повислая, ребристая, плосколистная и даурская (Betula verrucosa Ehrh., B. pendula Roth., B. costata Trautv., B. platyphylla Suk., B. dahurica Pall.) | ||||||
| Mean | 43 | 14.1 | 499.2 | 532.7 | –15 | 415 |
| Min | 4 | 1.0 | 352.3 | 215.1 | –30 | 317 |
| Max | 142 | 48.0 | 769.4 | 1083.3 | –5 | 826 |
| SD | 22.8 | 8.0 | 45.9 | 97.8 | 4.5 | 123.2 |
| CV, % | 52.7 | 56.9 | 9.2 | 18.4 | –29.4 | 29.7 |
| n | 898 | 900 | 900 | 900 | 900 | 900 |
| Осина, тополь Давида (Populus tremula L., P. davidiana Dode) | ||||||
| Mean | 32 | 15.7 | 417.3 | 468.0 | –15 | 379 |
| Min | 3 | 1.1 | 301.0 | 311.0 | –18 | 317 |
| Max | 57 | 45.7 | 500.0 | 692.0 | –5 | 826 |
| SD | 13.1 | 9.0 | 23.7 | 36.1 | 4.8 | 111.4 |
| CV, % | 41.3 | 57.1 | 5.7 | 7.7 | –32.0 | 29.4 |
| n | 317 | 317 | 317 | 317 | 317 | 317 |
| Липа мелколистная, амурская и маньчжурская (Tilia cordata Mill., T. amurensis Rupr., T. mandshurica Rupr. & Maxim.) | ||||||
| Mean | 52 | 17.7 | 415.8 | 483.8 | –15 | 588 |
| Min | 10 | 3.1 | 252.3 | 248.5 | –16 | 570 |
| Max | 115 | 38.6 | 647.8 | 900.0 | –10 | 826 |
| SD | 22.2 | 8.4 | 73.6 | 105.5 | 2.1 | 65.2 |
| CV, % | 42.4 | 47.7 | 17.7 | 21.8 | –14.2 | 11.1 |
| n | 202 | 202 | 202 | 202 | 202 | 202 |
| Дуб черешчатый и монгольский (Quercus robur L., Q. mongolica Fisch. ex Ledeb.) | ||||||
| Mean | 41 | 16.4 | 582.8 | 412.8 | –5 | 574 |
| Min | 6 | 1.9 | 450.0 | 269.6 | –10 | 570 |
| Max | 166 | 50.5 | 800.0 | 800.0 | –5 | 826 |
| SD | 23.9 | 9.4 | 50.6 | 71.4 | 0.6 | 32.6 |
| CV, % | 58.2 | 57.1 | 8.7 | 17.3 | –12.5 | 5.7 |
| n | 425 | 425 | 425 | 425 | 425 | 425 |
Примечание. Mean, Min и Max соответственно среднее, минимальное и максимальное значения; SD – стандартное отклонение; CV – коэффициент вариации; n – число наблюдений; A – возраст дерева, лет; d – диаметр ствола на высоте груди, см; T – средняя температура января, град.; PR – среднегодовые осадки, мм; DWи DB – соответственно базисная плотность древесины и коры, кг/м3. Необходимость анализа БП по средней температуре января, а не по среднегодовой или летней температуре была обоснована ранее (Usoltsev et al., 2020a, 2020b).
Эмпирические данные БП на пробных площадях получены по 3–10 дискам, выпиленным вдоль по стволу. БП (отдельно древесины и коры) рассчитана в результате обмера и взвешивания дисков, сушки до постоянной массы и повторного взвешивания. БП всего ствола рассчитана как средневзвешенная по площади поперечного сечения дисков.
В нашем предыдущем анализе фракционного состава фитомассы деревьев рода Лиственница (Larix Mill.) и ее связи с климатическими факторами была реализована следующая структура модели (Usoltsev et al., 2020b):
(1)
$\begin{gathered} \ln Pi = {{a}_{{0i}}} + {{a}_{{1i}}}\left( {\ln A} \right) + {{a}_{{2i}}}\left( {\ln d} \right) + {{a}_{{3i}}}\left( {\ln h} \right) + \\ + \,\,{{a}_{{4i}}}\left( {\ln d} \right)\left( {\ln h} \right) + {{a}_{{5i}}}\left[ {\ln \left( {T + 50} \right)} \right] + {{a}_{{6i}}}\left( {\ln PR} \right), \\ \end{gathered} $где h – высота дерева, м; Pi – фитомасса i-й фракции (стволы, ветви, хвоя), кг. При подборе структуры модели для БП мы взяли за основу модель (1), дополнив ее комбинированной независимой переменной [ln(T + 50)](lnPR), характеризующей совместное действие температуры и осадков. С целью учета нелинейности исследуемой закономерности применена аллометрическая форма модели, предполагающая процедуру линеаризации путем логарифмирования переменных. Поскольку в районах многолетней мерзлоты температура января иногда приближается к –40…–50°C, для выполнения процедуры линеаризации температура T модифицирована к виду (T + 50).
При моделировании биомассы деревьев получили распространение модели смешанного типа (Fu et al., 2012; Zeng, 2017). Аллометрическая модель смешанного типа включает два типа независимых переменных – численные, принимающие значения из непрерывного ряда чисел, и фиктивные переменные, представляющие дискретные качественные характеристики (Freese, 1964). Путем ввода в модель фиктивных переменных кодируются (опосредуются) свойства нескольких обособленных уровней (Дрейпер, Смит, 1973). Поскольку аллометрические закономерности относительно схожи у различных древесных видов (родов), строится модель биомассы смешанного типа для нескольких древесных видов одновременно (Fu et al., 2012; Zeng, 2017).
Древесные виды представлены в базе данных крайне неравномерно: от 966 по сосне обыкновенной до 74 деревьев по кедрам сибирскому и корейскому. Разная представленность видов как в климатических градиентах в пределах ареалов, так и по общему количеству наблюдений может быть причиной возможных отклонений от той общей реакции видов, которая обусловлена общностью воздействия на них биоэкологических и климатических факторов (Molteberg, Høibø, 2007). Исходя из качественного уровня имеющейся базы данных, мы предполагаем, руководствуясь принципом “гармонизации” видоспецифичных закономерностей (Jacobs, Cunia, 1980), построить аллометрические модели БП смешанного типа, включающие как численные (дендрометрические показатели и климатические переменные), так и фиктивные переменные, кодирующие видовую принадлежность исходных данных (табл. 2). Возможное влияние обеспеченности элементами питания и других связанных факторов на БП (Castro et al., 2021) в нашем исследовании не рассматривается.
Таблица 2.
Схема кодирования фиктивными переменными 9 лесообразующих видов Евразии
| Вид (род) | Блок фиктивных переменных | |||||||
|---|---|---|---|---|---|---|---|---|
| Х1 | Х2 | Х3 | X4 | X5 | X6 | X7 | X8 | |
| Сосна | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Кедр | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Лиственница | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ель | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Пихта | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Береза | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Осина | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| Липа | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Дуб | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
Мы приняли следующую структуру аллометрической модели смешанного типа:
(2)
$\begin{gathered} \ln \left( {DW} \right){\text{ и }}\ln \left( {DB} \right) = {{a}_{0}} + {{b}_{1}}\ln \left( A \right) + {{b}_{2}}\ln \left( d \right) + \\ + \,\,{{b}_{3}}\ln \left( h \right) + {{b}_{4}}\ln \left[ {\ln \left( {T + 50} \right)} \right] + {{b}_{5}}\ln \left( {\ln PR} \right) + \\ + \,\,{{b}_{6}}\left[ {\ln \left( {T + 50} \right)} \right]\left( {\ln PR} \right) + \Sigma {{a}_{i}}{{X}_{i}}, \\ \end{gathered} $РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В результате регрессионного анализа получены модели:
– для БП древесины ствола:
(3)
$\begin{gathered} \ln \left( {DW} \right) = 18.3231 + 0.0694\ln A - 0.0451\ln d - \\ - \,\,3.5227\ln \left( {T + 50} \right) - 2.1416\ln \left( {PR} \right) + \\ + \,\,0.6053\left[ {\ln \left( {T + 50} \right)} \right]\left( {\ln PR} \right) - 0.1235{{X}_{1}} + \\ + \,\,0.2638{{X}_{2}} - 0.0553{{X}_{3}} - 0.1670{{X}_{4}} + 0.2060{{X}_{5}} + \\ + \,\,0.0463{{X}_{6}} + 0.0121{{X}_{7}} + 0.2973{{X}_{8}}; \\ adj{{R}^{2}} = 0.621;\,\,\,\,SE = 0.11; \\ \end{gathered} $(4)
$\begin{gathered} \ln \left( {DB} \right) = 57.7517 - 0.0334\ln A - 0.0299\ln d - \\ - \,\,14.5700\ln \left( {T + 50} \right) - 8.4924\ln \left( {PR} \right) + 2.3879 \times \\ \times \,\,\left[ {\ln \left( {T + 50} \right)} \right]\left( {\ln PR} \right) + 0.8044{{X}_{1}} + 0.0468{{X}_{2}} + \\ + \,\,0.2878{{X}_{3}} + 0.2575{{X}_{4}} + 0.4970{{X}_{5}} + \\ + \,\,0.3526{{X}_{6}} + 0.4504{{X}_{7}} + 0.1445{{X}_{8}}; \\ adj{{R}^{2}} = 0.565;\,\,\,\,SE = 0.20. \\ \end{gathered} $Численные переменные в уравнениях (3) и (4) оказались значимыми на уровне p < 0.0001. Уравнения (3) и (4) действительны в пределах диапазона варьирования независимых переменных для каждого вида (табл. 1). Судя по знакам регрессионных коэффициентов переменных А и d, БП древесины имеет положительную корреляцию с возрастом дерева и отрицательную – с диаметром ствола. И та, и другая закономерности связаны с увеличением доли поздней древесины в годичном кольце и подтверждаются ранее выполненными исследованиями других авторов (Howe, 1974; Lachowicz et al., 2019). В отличие от БП древесины, БП коры имеет с возрастом отрицательную связь вследствие возрастного увеличения доли корки (Андреева, Родман, 2003).
Чтобы выполнить анализ изменений БП в координатах температур и осадков, мы подставляем в уравнения средние значения А и d из табл. 1 и делаем трехмерную геометрическую интерпретацию уравнений на примере сосны обыкновенной. Использование фиктивных переменных в моделях (3) и (4) означает, что различия БП разных древесных видов определяются лишь сдвигом свободного члена уравнений по оси абсцисс при постоянстве регрессионных коэффициентов численных переменных. Поэтому мы строим трехмерное изображение зависимости БП древесины и коры от климатических переменных только для одного вида, в частности, для сосны обыкновенной (рис. 1), поскольку для остальных древесных видов конфигурация трехмерных поверхностей изменения БП в координатах температур и осадков повторяется.
Рис. 1.
Изменение расчетных значений БП древесины (а) и коры (б) стволов сосны обыкновенной в территориальных градиентах зимних температур и осадков. Обозначения см. в табл. 1.
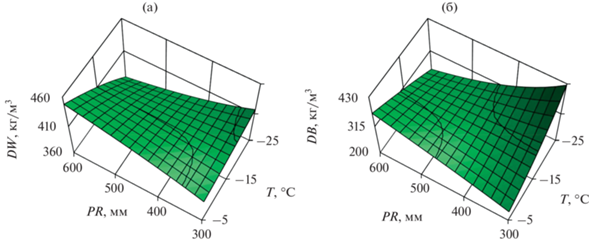
Результаты регрессионного анализа дали возможность оценить вклады дендрометрических и климатических переменных, а также видовой принадлежности деревьев в объяснение изменчивости БП древесины и коры стволов (Лиепа, 1980).
Судя по данным табл. 3, дендрометрические переменные объясняют изменчивость БП на уровне от 4 до 15%, климатические переменные – от 11 до 19%, и наибольший вклад в объяснение общей изменчивости БП вносит видовая принадлежность деревьев – от 74 до 77%. Это соответствует выводу о том, что внутривидовая изменчивость БП несопоставимо меньше по сравнению с межвидовой (Kerfriden et al., 2021).
Таблица 3.
Вклад дендрометрических переменных, климатических переменных и видовой принадлежности в объяснение изменчивости БП древесины и коры стволов, %
| Номер модели | ln A (I) |
ln d (II) |
(I) + (II) | ln(T + 50) (III) |
lnPR (IV) |
[ln(T + 50)](ln PR) (V) | (III) + (IV) + (V) | ΣaiXi |
|---|---|---|---|---|---|---|---|---|
| (3) | 8.7 | 6.6 | 15.3 | 3.3 | 3.5 | 3.6 | 10.6 | 74.1 |
| (4) | 2.0 | 2.0 | 4.0 | 6.2 | 6.2 | 6.3 | 18.7 | 77.3 |
На основе принципа пространственно-временного замещения (Усольцев, Цепордей, 2021б) мы использовали закономерности изменения базисной плотности древесины и коры в пространственных градиентах температур и осадков для прогнозирования изменений названных показателей при возможном увеличении средней температуры января на 1°C, предполагая неизменными осадки, и при возможном сокращении годичных осадков на 20 мм, предполагая неизменной температуру. Для этого мы взяли первую производную от 3D-зависимостей в координатах температур и осадков, показанных на рис. 1, и результаты графически представили на рис. 2 и 3. Подробности построения рис. 2 и 3 были изложены ранее (Усольцев, Цепордей, 2021а). Реакция БП на предполагаемые темпоральные изменения температур и осадков показана на рис. 2 и 3 в процентном выражении, и процент увеличения БП показан красным, а процент снижения – голубым цветом.
Рис. 2.
Изменение БП древесины (а) и коры (б) сосны обыкновенной при повышении температуры на 1°C в связи с ожидаемым изменением климата на разных территориальных уровнях температур и осадков. Здесь и далее: (1) – плоскость, соответствующая нулевому изменению БП при ожидаемом повышении температуры на 1°C; (2) – линия разграничения положительных и отрицательных изменений БП при ожидаемом повышении температуры на 1°C.
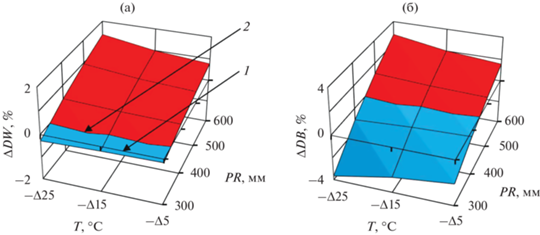
Рис. 3.
Изменение БП древесины (а) и коры (б) сосны обыкновенной при сокращении среднегодовых осадков на 20 мм в связи с ожидаемым изменением климата на разных территориальных уровнях температур и осадков.
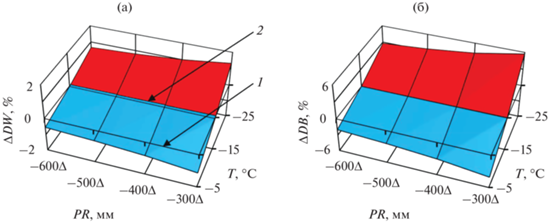
В нашем предыдущем исследовании, посвященном изменчивости надземной фитомассы деревьев и древостоев основных лесообразующих видов Евразии под влиянием зимних температур и годичных осадков, были показаны возможности применения климатически обусловленных моделей фитомассы при прогнозировании ее темпоральных изменений на основе принципа пространственно-временнóго замещения. В результате реализации принципов лимитирующего фактора Либиха и пространственно-временнóго замещения была установлена общая для основных древесных видов (родов) закономерность: в достаточно влагообеспеченных климатических поясах повышение температуры вызывает увеличение фитомассы, а во влагодефицитных поясах – ее снижение; в теплых климатических поясах сокращение уровня осадков вызывает снижение фитомассы, а в холодных – ее увеличение, т.е. в обоих случаях происходит смена лимитирующего фактора. Это означает, что в одних условиях лимитирующим фактором является как недостаток, так и избыток тепла, а в других – как недостаток, так и избыток осадков относительно некоторой нормы. Тем самым было показано проявление закона лимитирующего фактора по отношению к фитомассе деревьев и древостоев на трансконтинентальном уровне (Усольцев, Цепордей, 2021а, 2021б; Усольцев и др., 2021), причем по отношению не только к фитомассе деревьев и древостоев, но и к чистой первичной продукции (ЧПП) древостоев (Usoltsev et al., 2022).
Судя по рис. 1–3, закономерности изменения БП древесины и коры в градиентах температур и осадков у всех 9 древесных видов (родов) Евразии полностью повторяют закономерности изменения надземной фитомассы деревьев и древостоев. Таким образом, в градиентах температур и осадков на территории Евразии происходит смена лимитирующего фактора по показателям как фитомассы деревьев и фитомассы и ЧПП древостоев, так и БП древесины и коры основных лесообразующих видов. Подобная смена лимитирующего фактора была показана на примере радиального прироста березы (Фонти, 2020). На меридиональном трансекте в северной тайге Сибири лимитирующим фактором является температура, а в зоне степи – осадки, при этом смена лимитирующего фактора происходит в подзоне средней тайги.
ВЫВОДЫ
В ответ на поставленные задачи нашего исследования мы можем резюмировать:
– при моделировании изменений БП древесины и коры деревьев лесообразующих видов Евразии в связи с территориально обусловленными показателями температур и осадков на трансконтинентальном уровне подтверждено действие закона лимитирующего фактора;
– вклады независимых переменных в объяснение изменчивости БП древесины и коры составили: дендрометрических переменных – от 4 до 15%, климатических переменных – от 11 до 19% и видовой принадлежности деревьев – от 74 до 77%.
– показана возможность применения построенных моделей БП древесины и коры деревьев, чувствительных к территориальным изменениям температур и осадков, при прогнозировании изменений БП деревьев во времени на основе принципа пространственно-временнóго замещения;
– установлена общая для основных древесных видов закономерность смены лимитирующего фактора: в достаточно влагообеспеченных климатических поясах повышение температуры вызывает увеличение как БП древесины и коры, так и фитомассы деревьев и древостоев, а во влагодефицитных поясах – ее снижение; в теплых климатических поясах сокращение осадков вызывает снижение БП и фитомассы, а в холодных – ее увеличение. В одних условиях лимитирующим фактором является как недостаток, так и избыток тепла, а в других – как недостаток, так и избыток осадков относительно некоторой нормы. Таким образом, климатическая обусловленность исследуемых биопродукционных показателей имеет общий характер как для количественных, так и для квалиметрических показателей деревьев и древостоев.
Список литературы
Андреева И.И., Родман Л.С. Ботаника. М.: КолосС, 2003. 528 с.
Дрейпер Н., Смит Г. Прикладной регрессионный анализ. М.: Статистика, 1973. 392 с.
Лиепа И.Я. Динамика древесных запасов: Прогнозирование и экология. Рига: Зинатне, 1980. 170 с.
Полубояринов O.И. Плотность древесины. М.: Лесн. пром-сть, 1976. 160 с.
Исаева Л.Н. Метод расчета локальной и средней плотности абсолютно сухой древесины в стволах сосны и лиственницы // Лесоведение. 1978. № 4. С. 90-94.
Мелехов В.И., Бабич Н.А., Корчагов С.А. Качество древесины сосны в культурах. Архангельск: Изд-во АГТУ, 2003. 110 с.
Усольцев В.А., Цепордей И.С. Квалиметрия фитомассы лесных деревьев: плотность и содержание сухого вещества: монография. Екатеринбург: Уральский государственный лесотехнический университет, Ботанический сад УрО РАН, 2020. 182 с. (CD-ROM). https://elar.usfeu.ru/bitstream/123456789/10022/1/Usolcev_20.pdf
Усольцев В.А., Цепордей И.С. Климатически обусловленные территориальные изменения фитомассы деревьев лесообразующих видов Евразии и их прогнозирование // Сибирский лесной журн. 2021а. № 6. С. 72–90.
Усольцев В.А., Цепордей И.С. Принцип пространственно-временнóго замещения в экологии и прогнозирование биомассы Picea spp. при изменении климата // Хвойные бореальной зоны. 2021б. Т. 39. № 4. С. 269–275.
Усольцев В.А., Цепордей И.С., Азаренок М.В. Климатически обусловленные пространственные и темпоральные изменения биомассы рода Abies spp. Евразии в контексте закона лимитирующего фактора // Хвойные бореальной зоны. 2021. Т. 39. № 5. С. 392–400.
Фонти М.В. Климатический сигнал в параметрах годичных колец (плотности древесины, анатомической структуре и изотопном составе) хвойных и лиственных видов деревьев в различных природно-климатических зонах Евразии: дис. … докт. биол. наук: 03.02.08. Красноярск: СибФУ, 2020. 45 с.
Baskerville G.L. Use of logarithmic regression in the estimation of plant biomass // Canadian J. Forest Research. 1972. V. 2. P. 49-53.
Billard A., Bauer R., Mothe F., Colin F., Longuetaud F. Vertical variations in wood basic density for two softwood species // European J. Forest Research. 2021. V. 140. P. 1401–1416.
Bouriaud O., Leban J.-M., Bert D., Deleuze C. Intra-annual variations in climate influence growth and wood density of Norway spruce // Tree Physiology. 2005. V. 25. P. 651–660.
Castro V.R., Chambi-Legoas R., Filho M.T., Surdi P.G., Zanuncio J.C., Zanuncio A.J.V. The effect of soil nutrients and moisture during ontogeny on apparent wood density of Eucalyptus grandis // Scientific Reports. 2021. V. 11. № 1. Article 2530. https://doi.org/10.1038/s41598-020-59559-2
Falster D.S., Duursma R.A., Ishihara M.I., Barneche D.R., FitzJohn R.G., Vårhammar A., Aiba M., Ando M., Anten N., Aspinwall M.J., Baltzer J.L., Baraloto C., Battaglia M., Battles J.J., Bond-Lamberty B., van Breugel M., Camac J., Claveau Y., Coll L., Dannoura M., Delagrange S., Domec J.-C., Fatemi F., Feng W., Gargaglione V., Goto Y., Hagihara A., Hall J.S., Hamilton S., Harja D., Hiura T., Holdaway R., Hutley L.S., Ichie T., Jokela E.J., Kantola A., Kelly J.W.G., Kenzo T., King D., Kloeppel B.D., Kohyama T., Komiyama A., Laclau J.-P., Lusk C.H., Maguire D.A., le Maire G., Mäkelä A., Markesteijn L., Marshall J., McCulloh K., Miyata I., Mokany K., Mori S., Myster R.W., Nagano M., Naidu S.L., Nouvellon Y., O’Grady A.P., O’Hara K.L., Ohtsuka T., Osada N., Osunkoya O.O., Peri P.L., Petritan A.M., Poorter L., Portsmuth A., Potvin C., Ransijn J., Reid D., Ribeiro S.C., Roberts S.D., Rodríguez R., Saldaña-Acosta A., Santa-Regina I., Sasa K., Selaya N.G., Sillett S.C., Sterck F., Takagi K., Tange T., Tanouchi H., Tissue D., Umehara T., Utsugi H., Vadeboncoeur M.A., Valladares F., Vanninen P., Wang J.R., Wenk E., Williams R., Ximenes F. de Aquino, Yamaba A., Yamada T., Yamakura T., Yanai R.D., York R.A. BAAD: a Biomass And Allometry Database for woody plants // Ecology. 2015. V. 96. Article 1445.
Fearnside P.M. Wood density for estimating forest biomass in Brazilian Amazonia // Forest Ecology and Management. 1997. V. 90. № 1. P. 59–89.
Freese F. Linear regression methods for forest research. USDA Forest Service. Res. Paper FPL 17. Madison, 1964. 136 p.
Fu L.Y., Zeng W.S., Tang S.Z., Sharma R.P., Li H.K. Using linear mixed model and dummy variable model approaches to construct compatible single-tree biomass equations at different scales – A case study for Masson pine in Southern China // J. Forest Science. 2012. V. 58. № 3. P. 101–115.
Fujimoto T., Kita K., Kuromaru M. Genetic control of intra-ring wood density variation in hybrid larch (Larix gmelinii var. japonica × L. kaempferi) // Wood Science and Technology. 2008. V. 42. P. 227–240.
Howe J.P. Relationship of climate to the specific gravity of four Costa Rican hardwoods, an exploratory study // Wood Fiber. 1974. V. 5. P. 347–352.
Jacobs M.W., Cunia T. Use of dummy variables to harmonize tree biomass tables // Canadian J. Forest Research. 1980. V. 10. № 4. P. 483–490.
Kattge J., Bönisch G., Díaz S., Lavorel S., Prentice I.C., Leadley P., Tautenhahn S. et al. TRY plant trait database – Enhanced coverage and open access // Global Change Biology. 2020. V. 26. P. 119–188.
Kellomäki S. On geoclimatic variation in basic density of Scots pine wood // Silva Fennica. 1979. V. 13. № 1. P. 55–64.
Kerfriden B., Bontemps J.-D., Leban J.-M. Variations in temperate forest stem biomass ratio along three environmental gradients are dominated by interspecific differences in wood density // Plant Ecology. 2021. V. 222. № 3. P. 289–303.
Lachowicz H., Bieniasz A., Wojtan R. Variability in the basic density of silver birch wood in Poland // Silva Fennica. 2019. V. 53. № 1. Article id 9968.
Mikola P. On variations in tree growth and their significance to growth studies // Communicationes Instituti forestalis Fenniae. 1950. V. 38. № 5. P. 1–131.
Molteberg D., Høibø O. Modelling of wood density and fibre dimensions in mature Norway spruce // Canadian J. Forest Research. 2007. V. 37. № 8. P. 1373–1389.
Saikku O. The effect of fertilization on the basic density of Scots pine (Pinus silvestris L.). A densitometric study on the X ray chart curves of wood // Communicationes Instituti forestalis Fenniae. 1975. V. 85. № 3. P. 1-49.
Sousa V.B., Louzada J.L., Pereira H. Age trends and within-site effects in wood density and radial growth in Quercus faginea mature trees // Forest Systems. 2016. V. 25. № 1. Article e053.
St-Germain J.-L., Krause C. Latitudinal variation in tree-ring and wood cell characteristics of Picea mariana across the continuous boreal forest in Quebec // Canadian J. Forest Research. 2008. V. 38. № 6. P. 1397–1405.
Swenson N.G., Enquist B.J. Ecological and evolutionary determinants of a key plant functional trait: wood density and its community – wide variation across latitude and elevation // American J. Botany. 2007. V. 94. № 3. P. 451–459.
Télles J.R.G., Martínez A.V., de la Rosa A.B., Grande J.C., Mendoza C.P. Radial variation of basic density in Pinus patula Schltdl. et Cham. in three locations from Hidalgo state // Revista Mexicana de Ciencias Forestales. 2011. V. 2. № 7. P. 71–78.
Usoltsev V.A. Stem taper, density and dry matter content in biomass of trees growing in Central Eurasia: CD-monograph. Yekaterinburg: Ural State Forest Engineering University, Botanical Garden of Ural Branch of RAS, 2020. Available at: https://elar.usfeu.ru/handle/123456789/9649
Usoltsev V.A., Shobairi S.O.R., Tsepordey I.S., Chasovskikh V.P. On some differences in the response of Picea spp. and Abies spp. single-tree biomass structure to changes in temperatures and precipitation in Eurasia // Environment and Ecology. 2020a. V. 38. P. 300–315.
Usoltsev V.A., Shobairi S.O.R., Tsepordey I.S., Chasovskikh V.P. Additive model of aboveground biomass of larch single-trees related to age, DBH and height, sensitive to temperature and precipitation in Eurasia // J. Applied Sciences & Environmental Management. 2020b. V. 24. P. 1759–1766.
Usoltsev V., Zukow W., Tsepordey I. Climatically determined spatial and temporal changes in the biomass of Pinus sp. of Eurasia in the context of the law of the limiting factor // Ecological Questions. 2022. V. 33. № 1. P. 15–23. https://doi.org/10.12775/EQ.2022.007
Vieira J., Carvalho A., Campelo F. Tree growth under climate change: Evidence from xylogenesis timings and kinetics // Frontiers in Plant Science. 2020. V. 11. Article 90.
Wiemann M.C., Williamson G.B. Geographic variation in wood specific gravity: effects of latitude, temperature and precipitation // Wood and Fiber Science. 2002. V. 34. № 1. P. 96–107.
World Weather Maps, 2007. https://www.mapsofworld.com/ referrals/weather
Zanne A.E., Lopez-Gonzalez G., Coomes D.A., Ilic J., Jansen S., Lewis S.L., Miller R.B., Swenson N.G., Wiemann M.C., Chave J. Global wood density database. Dryad, 2009. Identifier: http://hdl.handle.net/10255/dryad.235
Zeng W.S. Developing tree biomass models for eight major tree species in China. In: Biomass volume estimation and valorization for energy. Chapter 1. Intech Publ., 2017. P. 3–21.
Zhang L., Shi H. Local modeling of tree growth by geographically weighted regression // Forest Science. 2003. V. 50. № 2. P. 225–244.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение