

Агрохимия, 2021, № 2, стр. 55-64
Влияние 24-эпибрассинолида на рост, содержание фотосинтетических пигментов, холодоустойчивость и антиоксидантную активность растений томата
А. Н. Дерябин 1, *, Т. А. Суворова 1, С. В. Сычева 2, С. Н. Деревщюков 2
1 Институт физиологии растений им. К.А. Тимирязева РАН
127276 Москва, ул. Ботаническая, 35, Россия
2 Воронежская овощная опытная станция – филиал Федерального научного центра овощеводства
369116 Воронежская обл., Верхнехавский р-н, п. НИИОХ, ул. Садовая, 3а, Россия
* E-mail: anderyabin@mail.ru
Поступила в редакцию 02.10.2019
После доработки 18.06.2020
Принята к публикации 10.11.2020
Аннотация
Исследовали реакцию растений томата (Solаnum lycopеrsicum Mill.) сортов Яхонт (среднеранний) и Кулон (скороспелый), на применение регулятора роста 24-эпибрассинолида (24-ЭБЛ) в концентрации 12.5 мкг/л, при действии низких положительных температур (НПТ). Предпосевная обработка семян 24-ЭБЛ не влияла на рост осевых органов проростков, но при действии НПТ оказала более выраженное антистрессовое действие на прорастание семян томата сорта Яхонт. 24-ЭБЛ на фоне НПТ способствовал увеличению содержания фотосинтетических пигментов в растениях сорта Кулон, а сорта Яхонт – сохранению их исходно высокого содержания. При НПТ листья растений томата сорта Кулон показали бóльшую отзывчивость на экзогенное (предпосевная обработка семян и/или опрыскивание растений за 1 сут до охлаждения) применение 24-ЭБЛ, что выражалось в снижении интенсивности процесса перекисного окисления липидов (ПОЛ). В этих условиях растения томата сорта Яхонт на 24-ЭБЛ реагировали сдержанно, о чем свидетельствовало отсутствие в их листьях достоверных изменений в интенсивности ПОЛ. 24-ЭБЛ не влиял на уровень антиоксидантной активности экстрактов листьев, при этом у сорта Яхонт он был исходно в 2 раза больше, чем у сорта Кулон. Сделан вывод, что меньшая отзывчивость растений томата сорта Яхонт на применение 24-ЭБЛ на фоне НПТ обусловлена их генетически детерминированной более высокой холодоустойчивостью, чем сорта Кулон.
ВВЕДЕНИЕ
В начальный период выращивания теплолюбивых культур (огурца, томата и др.) часто происходят резкие перепады дневных и ночных температур, а также кратковременные ночные заморозки, которые повреждают растения, замедляют их рост и развитие, приводят к снижению продуктивности. Одним из приемов снижения повреждающего действия абиотических стресс-факторов, в т.ч. низких температур, на растения является использование синтетических регуляторов роста, обладающих антистрессовой активностью [1–4].
Регулятор роста 24-эпибрассинолид (24-ЭБЛ) является синтетическим аналогом природного фитогормона брассиностероида и в ходе онтогенеза растений проявляет не только высокую регуляторную активность в отношении многих физиологических процессов (деления клеток, прорастания семян, ризогенеза, фотосинтеза, синтеза белков и нуклеиновых кислот и др.), но также повышает устойчивость к неблагоприятным факторам внешней среды [4–9]. 24-ЭБЛ регулирует внутриклеточные физиологические процессы во взаимодействии с эндогенными фитогормонами [10], поддерживает баланс между активными формами кислорода (АФК) и антиоксидантами [11] и индуцирует системную устойчивость растительного организма [8].
Томат – одна из основных сельскохозяйственных культур, которая по объему товарного производства и площади выращивания занимает 10-e место в мире [12]. В литературе имеются сведения о полифункциональном действии 24-ЭБЛ на томаты. Показано, что при применении 24-ЭБЛ повышались энергия прорастания и всхожесть семян [2, 13], биометрические показатели растений в фазе массового плодоношения [14], размер и масса плодов [2], урожайность [15–17], устойчивость растений к серой гнили [6], тяжелым металлам (Ni) [18], засухе [19]. Работы, в которых изучали реакцию растений томата на 24-ЭБЛ при действии низких положительных температур (НПТ), единичны. Например, опрыскивание 50-суточных растений томата раствором 24-ЭБЛ в последействии охлаждения способствовало усилению функционирования антиоксидантной системы и повышению содержания пролина в листьях [20]. Холодовое повреждение тканей плодов томата было менее выражено при предобработке их 24-ЭБЛ [21]. Вместе с тем при оценке эффективности действия 24-ЭБЛ на растения некоторые авторы отмечали неодинаковую реакцию разных сортов, особенно на фоне неблагоприятных факторов среды, что могло быть связано с различным гормональным статусом, антиоксидантной активностью тканей и др. [4, 16, 17, 20]. Например, сортовая специфичность действия 24-ЭБЛ выявлена при определении активностей антиоксидантных ферментов у пшеницы на фоне солевого стресса [22] и у томата при действии НПТ [20]. У проростков озимой пшеницы эндогенный уровень 24-ЭБЛ зависел от степени морозостойкости сорта [23]. Предпосевная обработка семян пшеницы 24-ЭБЛ способствовала повышению в условиях засухи интенсивности фотосинтеза в растениях засухоустойчивого сорта Саратовская 29 и не влияла на этот показатель у неустойчивого сорта Энита [24].
Цель работы – изучение физиолого-биохимических реакций 2-х сортов томата, различающихся по скороспелости, на применение 24-ЭБЛ на фоне действия НПТ. В задачи исследования входила оценка влияния экзогенного 24-ЭБЛ на сорта томатов по следующим показателям: всхожесть семян, морфометрические параметры осевых органов, содержание в листьях фотосинтетических пигментов, интенсивность перекисного окисления липидов (ПОЛ) и антиоксидантная активность (АОА) экстрактов листьев.
МЕТОДИКА ИССЛЕДОВАНИЯ
Объектами исследования были семена и молодые растения томата Solanum lycopersicum Mill. сортов Яхонт и Кулон селекции Воронежской овощной опытной станции. Растения обоих сортов имеют компактный детерминантный тип куста, но различаются по скороспелости: сорт Яхонт – среднеранний, тогда как сорт Кулон – скороспелый [25].
В работе использовали препарат эпин-Экстра (ННПП НЭСТ М, Россия), содержащий в качестве действующего вещества синтетический брассиностероид 24-ЭБЛ в концентрации 25 мг/л. Известно, что предпосевная обработка семян и опрыскивание вегетирующих растений раствором 24-ЭБЛ являются более эффективными способами, по сравнению с его внесением в почву [11]. В наших опытах 1 г семян томатов замачивали в водном растворе препарата эпин-Экстра в концентрации 0.5 мл/л (концентрация 24-ЭБЛ – 12.5 мкг/л) в течение 2-х ч. Контролем служила обработка семян дистиллированной водой. Семена проращивали в чашках Петри при 22 ± 0.5°С в темноте. Каждый вариант включал 4 повторности, по 100 семян в каждой. На 5-е сут оценивали энергию прорастания семян, на 10-е сут – их всхожесть, и измеряли длину осевых органов (корня и побега). Оценку биометрических показателей в каждом варианте проводили на 35–40 проростках.
Содержание фотосинтетических пигментов (хлорофилл a (Хл a), хлорофилл б (Хл б) и каротиноиды (кар)) в листьях томатов определяли спектрофотометрическим методом, при длинах волн, соответствующих максимумам поглощения Хл а, Хл б и кар в 80%-ном растворе ацетона – 663, 646 и 470 нм соответственно. Концентрации пигментов рассчитывали по следующим формулам:
где Сa, Сб, Скар – концентрации Хл а, Хл б и каротиноидов, мг/мл [26]. Содержание пигментов выражали в мг/г сухой массы листьев. Долю хлорофиллов в светособирающем комплексе (ССК) рассчитывали по формуле: ССК (%) = (Хл б + 1.2 × × Хл б)/(Хл а + Хл б) × 100, учитывая, что весь Хл б находится в ССК, а соотношение Хл а/Хл b в нем равно 1.2 [27].Содержание сухого вещества в листьях определяли после их высушивания в термостате при температуре 100–105°С до постоянной массы и выражали в % исходной сырой массы навески.
Оценку холодоустойчивости сортов томата проводили двумя способами: по прорастанию семян при НПТ и по интенсивности свободнорадикальных процессов в листьях растений. По первому способу контрольные и обработанные 24-ЭБЛ семена помещали в чашки Петри на фильтровальную бумагу, увлажненную дистиллированной водой, и проращивали при 10 ± 0.5°С в темноте в течение 25 сут. Каждый вариант включал 4 повторности, по 100 семян в каждой. Долю проросших при НПТ семян определяли каждые 5 сут.
По второму способу контрольные и обработанные 24-ЭБЛ семена высаживали в вазоны с субстратом на основе верхового торфа Агробалт-С (Росторфинвест, Россия) и проращивали в течение 3-х нед при 22 ± 0.5°С и 16-часовом фотопериоде с интенсивностью освещения 100 мкмоль квантов/(м2 с), лампы OSRAM L 80W/640 (ОСРАМ, Смоленск, Россия). Растения охлаждали в климатической камере MIR-153 (Sanyo, Япония), в темноте. Использовали краткосрочное (15 мин) охлаждение при 4 ± 0.5°С и длительное (24 ч) при 2 ± 0.5°С, при этом за 1 сут до охлаждения половину вазонов с растениями каждого варианта опрыскивали водным раствором препарата эпин-Экстра в концентрации 0.5 мл/л (концентрация 24-ЭБЛ – 12.5 мкг/л).
Холодоустойчивость растений оценивали по накоплению в листьях малонового диальдегида (МДА) – индикатора интенсивности процесса перекисного окисления липидов (ПОЛ). МДА в реакции с 2-тиобарбитуровой кислотой образует стойкий окрашенный триметиновый комплекс с максимумом поглощения при 532 нм. Неспецифическую адсорбцию определяли при длине волны 600 нм. Оптическую плотность растворов измеряли на спектрофотометре Genesys 10 UV (Termo Electron Corporation, США) с последующим пересчетом в концентрацию по величине молярной экстинкции (ε = 1.56 × 105 см–1 моль–1). Количество МДА рассчитывали в мкмоль/г сырой массы листьев [28]. Опыты проводили трижды в 3-х аналитических повторностях, каждая из которых включала навеску листьев, взятых из среднего яруса 3–4-х растений.
Антиоксидантную активность (АОА) экстрактов листьев томата анализировали по оптической плотности в реакции со стабильным радикалом 2,2-дифенил-1-пикрилгидразилом (ДФПГ) по методу [29]. Навеску листьев массой 1 г гомогенизировали в 10 мл 96%-ного этилового спирта и центрифугировали при 1500g в течение 15 мин. К 2 мл экстракта добавляли 1 мл 200 мкМ ДФПГ в 50%-ном этаноле, инкубировали в течение 30 мин при 21–23°С, после чего измеряли оптическую плотность на спектрофотометре в кюветах с толщиной поглощающего слоя 10 мм, при длине волны 517 нм против 50%-ного этанола. Радикал ДФПГ, взаимодействуя с молекулой антиоксиданта, переходит в нерадикальную форму, что сопровождается исчезновением максимума поглощения при 517 нм и изменением оптической плотности раствора [30]. Степень ингибирования ДФПГ (в %) рассчитывали по формуле:
степень ингибирования ДФПГ (%) = [(Ао – (А – – Аb)/Ао) × 100], где А – оптическая плотность экстракта с ДФПГ, Ао – оптическая плотность ДФПГ без экстракта, Аb – оптическая плотность экстракта без ДФПГ [31].
Статистическую обработку результатов экспериментов проводили с использованием программы Т-test (ISI, США). Значимость различий между вариантами оценивали по t-критерию Стьюдента для независимых выборок. В таблицах и на рисунках представлены средние арифметические величины и их стандартные ошибки, достоверные при 95%-ном уровне значимости. Различия средних между вариантами и сортами отмечены разными надстрочными латинскими буквами. В статье обсуждают только величины, достоверные при P ≤ 0.05.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Результаты влияния предпосевной обработки семян томата 24-ЭБЛ на энергию прорастания и их всхожесть, а также рост осевых органов (корень, побег) представлены в табл. 1. Из представленных данных видно, что всхожесть семян сорта Кулон в контрольном варианте была выше на 29%, чем сорта Яхонт. В варианте с предпосевной обработкой семян 24-ЭБЛ у сорта Яхонт энергия прорастания была на 15% больше, чем в контроле. Однако высокие (98–99%) показатели энергии прорастания и всхожести семян сорта Кулон не позволили оценить действие 24-ЭБЛ на их посевные качества.
Таблица 1.
Влияние предпосевной обработки семян томата регулятором роста 24-ЭБЛ на всхожесть и рост осевых органов проростков, на 10-е сут
| Вариант | Энергия прорас-тания (5-е сут) | Всхожесть | Длина, мм | |
|---|---|---|---|---|
| % | корня | побега | ||
| Cорт Яхонт | ||||
| Контроль | 52.3 ± 1.5a | 70.3 ± 0.8c | 56.1 ± 3.1g | 29.7 ± 1.1h |
| 24-ЭБЛ | 67.2 ± 1.4b | 77.0 ± 0.9d | 55.6 ± 2.7g | 26.5 ± 1.3h |
| Cорт Кулон | ||||
| Контроль | 98.0 ± 0.4e | 99.0 ± 0.2f | 46.3 ± 4.2i | 40.5 ± 2.3j |
| 24-ЭБЛ | 98.2 ± 0.3e | 99.0 ± 0.4f | 46.1 ± 2.0i | 40.0 ± 1.6j |
Сорта томата различались по биометрическим показателям (рост осевых органов) проростков: у сорта Кулон в контрольном варианте длина побега была больше на 35%, а корня меньше на 20%, чем у сорта Яхонт. Важно отметить отсутствие влияния 24-ЭБЛ на рост осевых органов проростков у обоих сортов томата. В литературе имеются сведения о том, что брассиностероиды, оказывая стимулирующее действие на прорастание семян, слабо влияют на линейный рост органов растений [32, 33].
В литературе 24-ЭБЛ характеризуется как регулятор роста, снижающий негативное действие низкотемпературного стресс-фактора у различных видов растений [2–4, 6, 8–10]. Установлено, что стресс-защитный эффект 24-ЭБЛ обусловлен адаптивными изменениями количественного состава и баланса фитогормонов, а также повышенным уровнем экспрессии генов стрессового ответа, кодирующих белки холодового шока [32, 34]. Например, обработанные 24-ЭБЛ растения перца при действии НПТ имели стабильные фотосинтетические показатели, более высокие активности антиоксидантных ферментов (супероксиддисмутазы (СОД), пероксидазы, каталазы и аскорбат-пероксидазы), накапливали больше пролина, растворимых сахаров, белка, а их ткани содержали меньше МДА – маркера окислительного стресса [35].
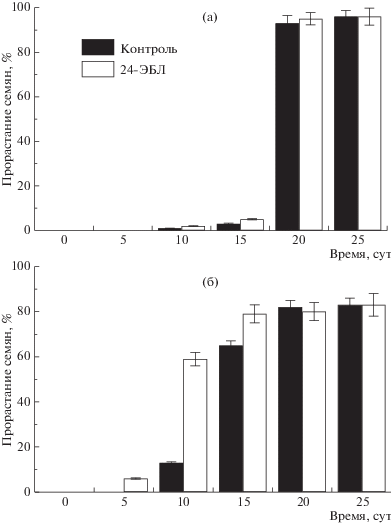
Принимая во внимание тот факт, что томат относится к теплолюбивым растениям, диапазон оптимальных температур для которых составляет 22 ± 7°С (при температуре <15°С растения не цветут, а при температуре <8°С прекращают рост) [12], для опытов по изучению стресс-протекторного действия 24-ЭБЛ на прорастание семян использовали температуру 10 ± 0.5°С. Согласно полученным данным (рис. 1), при НПТ семена томата прорастали медленнее, чем при 22 ± 0.5°С (табл. 1). Если у сорта Яхонт в контроле первые проросшие семена появились на 10-е сут, а в варианте с обработкой 24-ЭБЛ – на 5-е сут, то у сорта Кулон прорастание семян началось на 10-е сут, независимо от варианта (рис. 1). Следовательно, применение 24-ЭБЛ оказало более выраженное антистрессовое действие на прорастание семян томата сорта Яхонт, что проявилось в их быстром и дружном прорастании, в отличие от сорта Кулон. Предположили, что сорт Яхонт проявил себя как более устойчивый к НПТ, тогда как сорт Кулон – как менее холодоустойчивый.
Рис. 1.
Влияние предпосевной обработки семян томата 24-ЭБЛ на их прорастание при 10 ± 0.5°С, %: (а) – сорт Кулон, (б) – сорт Яхонт.
Известно, что у теплолюбивых растений при действии НПТ процесс фотосинтеза подавляется в первую очередь, при этом одним из параметров, используемых в качестве индикатора фотосинтетической активности, является количественный состав пигментов [36]. Содержание фотосинтетических пигментов в листьях является очень чувствительным показателем к действию НПТ. Результаты влияния предпосевной обработки семян 24-ЭБЛ на пигментный состав листьев растений томата при оптимальных температурных условиях и после 24 ч экспозиции при 2 ± 0.5°С в темноте представлены в табл. 2. Согласно полученным данным, до охлаждения контрольные растения обоих сортов не различались по содержанию пигментов, однако в варианте с применением 24-ЭБЛ у сорта Яхонт содержание Хл a и Хл б было больше на 23 и 29% соответственно по сравнению с сортом Кулон. Показатель Хл (a + б) и суммарное содержание пигментов у сорта Яхонт также были больше на 24% по сравнению с сортом Кулон.
Таблица 2.
Влияние предпосевной обработки семян 24-ЭБЛ на содержание фотосинтетических пигментов в листьях растений томата в норме (22 ± 0.5°С) и после охлаждения при 2± 0.5°С в течение 24 ч, мг/г сухой массы
| Показатель | Варианты | |||
|---|---|---|---|---|
| контроль | 24-ЭБЛ | контроль | 24-ЭБЛ | |
| 22 ± 0.5°С | 2 ± 0.5°С, 24 ч | |||
| Хл a | $\frac{{10.6 \pm {{{0.5}}^{{\text{a}}}}}}{{9.9 \pm {{{1.2}}^{{\text{a}}}}}}$ | $\frac{{9.9 \pm {{{0.8}}^{{\text{a}}}}}}{{12.2 \pm {{{1.3}}^{{\text{b}}}}}}$ | $\frac{{9.9 \pm {{{0.9}}^{{\text{a}}}}}}{{12.8 \pm {{{0.6}}^{{\text{b}}}}}}$ | $\frac{{13.8 \pm {{{0.8}}^{{\text{b}}}}}}{{14.6 \pm {{{1.7}}^{{\text{b}}}}}}$ |
| Хл б | $\frac{{3.7 \pm {{{0.3}}^{{\text{c}}}}}}{{3.4 \pm {{{0.3}}^{{\text{c}}}}}}$ | $\frac{{3.8 \pm {{{0.2}}^{{\text{c}}}}}}{{4.9 \pm {{{0.6}}^{{\text{d}}}}}}$ | $\frac{{3.3 \pm {{{0.1}}^{{\text{c}}}}}}{{4.4 \pm {{{0.1}}^{{\text{d}}}}}}$ | $\frac{{5.0 \pm {{{0.5}}^{{\text{d}}}}}}{{5.1 \pm {{{0.8}}^{{\text{d}}}}}}$ |
| Каротиноиды | $\frac{{2.4 \pm {{{0.1}}^{{\text{e}}}}}}{{2.2 \pm {{{0.3}}^{{\text{e}}}}}}$ | $\frac{{2.1 \pm {{{0.1}}^{{\text{e}}}}}}{{2.6 \pm {{{0.4}}^{{\text{e}}}}}}$ | $\frac{{2.3 \pm {{{0.2}}^{{\text{e}}}}}}{{3.3 \pm {{{0.1}}^{{\text{f}}}}}}$ | $\frac{{3.3 \pm {{{0.2}}^{{\text{f}}}}}}{{2.6 \pm {{{0.5}}^{{{\text{ef}}}}}}}$ |
| Хл (a + б) | $\frac{{14.2}}{{13.3}}$ | $\frac{{13.7}}{{17.1}}$ | $\frac{{13.8}}{{17.2}}$ | $\frac{{18.8}}{{19.6}}$ |
| Доля Хл ССК, % от Хл (a + б) | $\frac{{57.0}}{{55.9}}$ | $\frac{{61.7}}{{63.6}}$ | $\frac{{54.5}}{{56.3}}$ | $\frac{{58.9}}{{56.6}}$ |
НПТ способствовала некоторому снижению содержания хлорофиллов в листьях контрольных растений томата сорта Кулон, тогда как у сорта Яхонт, их содержание наоборот, возросло почти на треть. При применении 24-ЭБЛ и в последействии охлаждения у сорта Кулон наблюдали достоверное увеличение содержания Хл a и Хл б, на 40 и 20% соответственно. У обоих сортов после охлаждения в вариантах с 24-ЭБЛ, а также у контрольных растений сорта Яхонт содержание Хл a оставалось на высоком уровне. Отсутствие снижения содержания хлорофиллов у растений томата сорта Яхонт свидетельствовало о пониженной чувствительности их фотосинтетического аппарата к холоду. Данный факт мог свидетельствовать о возможной сбалансированной работе фотосинтетического аппарата (в частности, фотосистемы II) в последействии охлаждения, учитывая, что именно Хл a является преобразователем солнечной энергии, тогда как другие пигменты осуществляют ее поглощение и миграцию. Подобные изменения, связанные с увеличением содержания хлорофиллов и фотосинтетической активности листьев на фоне НПТ наблюдали у растений перца, предобработанных раствором 24-ЭБЛ [37].
При оптимальной температуре выращивания сорта томатов не различались по содержанию каротиноидов в листьях. После охлаждения в варианте с предобработкой семян 24-ЭБЛ содержание каротиноидов у сорта Кулон возросло на 50%, тогда как у сорта Яхонт оставалось на уровне неохлажденного варианта. Следует отметить, что в этих условиях содержание каротиноидов в листьях контрольных растений томата сорта Яхонт возросло на 26% по сравнению с неохлажденным вариантом. У обоих сортов предобработка семян 24-ЭБЛ на фоне НПТ способствовала увеличению в листьях суммарного содержания пигментов более чем на 40% (рис. 2). В этих условиях сорт Яхонт показал меньшую отзывчивость на применение 24-ЭБЛ: содержание пигментов в листьях оставалось на уровне контрольного варианта.
Рис. 2.
Влияние предпосевной обработки семян 24‑ЭБЛ на суммарное содержание фотосинтетических пигментов (Хл а, Хл б, каротиноиды) в листьях растений томата сортов Кулон и Яхонт при 22 ± 0.5°С (1) и после охлаждения при 2 ± 0.5°С в течение 24 ч (2). Достоверные различия средних величин при 95%-ном уровне значимости отмечены разными латинскими буквами. То же на рис. 3.
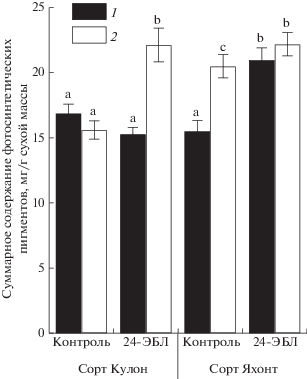
При оптимальных температурных условиях сорта не различались по такому показателю, как доля Хл б в ССК, тогда как в вариантах с применением 24-ЭБЛ он был больше – на 8% у сорта Кулон и на 13% у сорта Яхонт. После охлаждения доля Хл б в ССК снижалась у обоих сортов, а в вариантах с применением 24-ЭБЛ оставалась на уровне неохлажденного контроля. Следовательно, предобработка семян томата 24-ЭБЛ оказала существенное влияние на содержание фотосинтетических пигментов в листьях растений, при этом реакция сортов была неодинаковой. При оптимальных температурных условиях в варианте с 24-ЭБЛ более выраженные изменения в пигментном составе, связанные с увеличением содержания хлорофиллов, наблюдали у сорта Яхонт. При действии НПТ установлен стресс-защитный эффект 24-ЭБЛ на растения томата, направленный на адаптационные структурные изменения фотосинтетического аппарата, проявившиеся у сорта Кулон в виде увеличения содержания фотосинтетических пигментов, тогда как у сорта Яхонт – в сохранении их исходно высокого содержания на уровне неохлажденных растений.
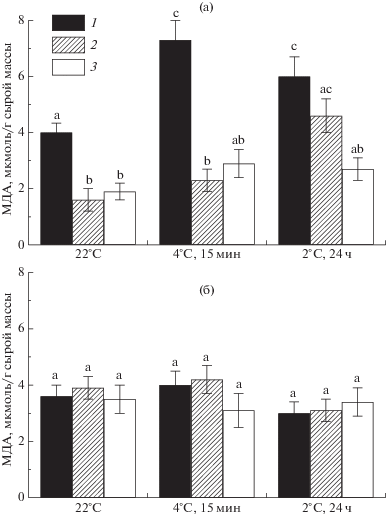
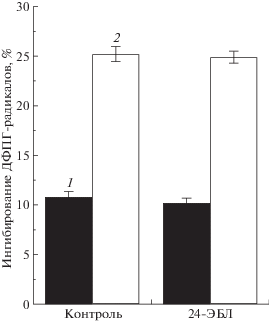
Одной из ранних неспецифических реакций растительной клетки на охлаждение является интенсификация окислительных процессов с участием АФК, накопление которых усиливает процесс ПОЛ мембран. МДА является одним из конечных продуктов ПОЛ и служит индикатором функционального состояния клеточных мембран [28]. Наши исследования показали, что в норме (без охлаждения) сорта томата не различались по содержанию МДА в листьях (рис. 3). Однако в вариантах с применением 24-ЭБЛ у сорта Кулон, в отличие от сорта Яхонт, наблюдали достоверное снижение содержания МДА. Различные режимы охлаждения способствовали накоплению МДА в листьях контрольных растений сорта Кулон, тогда как у сорта Яхонт этот показатель оставался на уровне неохлажденного контрольного варианта. Таким образом, при использовании 24-ЭБЛ (предпосевная обработка семян 24-ЭБЛ или предпосевная обработка семян + опрыскивание растений 24-ЭБЛ за 1 сут до охлаждения) содержание МДА в листьях растений сорта Кулон снижалось, тогда как у сорта Яхонт не изменялось. При НПТ растения томата сорта Кулон показали бóльшую отзывчивость на экзогенное применение 24-ЭБЛ, что выражалось в снижении у них интенсивности процесса ПОЛ. В этих условиях растения томата сорта Яхонт на 24-ЭБЛ реагировали сдержанно, о чем свидетельствовало отсутствие в их листьях достоверных изменений в содержании МДА относительно неохлажденного контроля. Полученные результаты подтверждают обнаруженные ранее (рис. 1) сортовые различия в холодоустойчивости: растения томата сорта Яхонт оказались более устойчивыми к охлаждению, чем растения сорта Кулон. Следовательно, при действии НПТ более высокая отзывчивость томата растений сорта Кулон на применение 24‑ЭБЛ была обусловлена их пониженной холодоустойчивостью, чем сорта Яхонт. Есть основания полагать, что повышенная устойчивость растений томата сорта Яхонт к охлаждению определялась наличием у них более эффективной антиоксидантной системы, чем у растений сорта Кулон. Для проверки данного предположения была проведена оценка АОА экстрактов листьев методом, основанным на взаимодействии стабильного хромоген-радикала ДФПГ с веществами (фенольные соединения, тиолы, аскорбиновая кислота и др.), проявляющих антиоксидантную активность [38]. Согласно методу, чем больше доля ингибирования ДФПГ-радикалов, тем больше АОА растительной ткани. Установлено, что экстракты листьев сортов томата различались по величине АОА, при этом доля ингибирования ДФПГ-радикалов у них была сходна в контроле и в варианте с применением 24-ЭБЛ (рис. 4). У сорта Кулон АОА экстрактов листьев была в 2 раза меньше, чем у сорта Яхонт. Таким образом, оценка АОА экстрактов листьев с помощью стабильного радикала ДФПГ не выявила влияние 24-ЭБЛ на уровень низкомолекулярных антиоксидантов, что указывало на возможное действие этого регулятора роста на ферментативную составляющую антиоксидантной системы. В литературе имеются сведения о том, что у растений томата экзогенный 24-ЭБЛ повышал активность антиоксидантных ферментов, при этом устойчивые к холоду сорта имели более высокие активности, чем неустойчивые [20]. Анализ изменений адаптивного потенциала разных сортов томата при применении препарата эпин-Экстра (0.1%-ный раствор) показал, что основной (более 50%) вклад в результаты опыта вносили особенности генотипов разных сортов [17]. Использованный в исследованиях сорт Яхонт в условиях агроценоза показал меньшую отзывчивость на применение 24-ЭБЛ по сравнению с другими сортами томата [16].
Рис. 3.
Влияние экзогенного 24-ЭБЛ на содержание МДА в листьях растений (возраст 3 нед) томата сортов Кулон (а) и Яхонт (б) при действии пониженных температур: 1 – контроль (вода), 2 – семена перед посадкой обработаны 24‑ЭБЛ, 3 – семена перед посадкой, а также растения за 1 сут до охлаждения обработаны 24-ЭБЛ.
ЗАКЛЮЧЕНИЕ
Таким образом, регулятор роста 24-ЭБЛ оказывал направленное влияние на отдельные этапы онтогенеза растений томата (прорастание семян, молодое растение), видимо, за счет мобилизации их генетических возможностей, что в итоге способствовало повышению их устойчивости к низким положительным температурам (НПТ). Данные по интенсивности окислительных процессов в листьях свидетельствовали о разной реакции сортов томата на 24-ЭБЛ в условиях действия НПТ: растения скороспелого сорта Кулон показали бóльшую отзывчивость, тогда как растения среднераннего сорта Яхонт на применение регулятора роста реагировали сдержанно. Применение 24-ЭБЛ приводило к более выраженным изменениям в пигментном составе у сорта Яхонт в отсутствии стресс-фактора, тогда как у сорта Кулон – при действии НПТ. 24-ЭБЛ не влиял на уровень антиоксидантной активности экстрактов листьев, при этом у сорта Яхонт он был в 2 раза выше, чем у сорта Кулон. Таким образом, одним из факторов, обусловливающих менее выраженную отзывчивость сорта Яхонт на обработку 24-ЭБЛ, являлось наличие у них более эффективной неферментативной антиоксидантной системы, чем у сорта Кулон. Следовательно, степень влияния 24-ЭБЛ на сорта томата была обусловлена уровнем их генетически детерминированной холодоустойчивости, который был выше у сорта Яхонт.
Список литературы
Прусакова Л.Д., Малеванная Н.Н., Белопухов С.Л., Вакуленко В.В. Регуляторы роста растений с антистрессовыми и иммунопротекторными свойствами // Агрохимия. 2005. № 11. С. 76–86.
Шаповал О.А., Вакуленко В.В., Прусакова Л.Д., Можарова И.П. Регуляторы роста растений в практике сельского хозяйства. М.: ВНИИА, 2009. 59 с.
Колмыкова Т.С., Лукаткин А.С. Эффективность регуляторов роста растений при действии абиотических стрессовых факторов // Агрохимия. 2012. № 1. С. 83–94.
Anwar A., Liu Y., Dong R., Bai L., Yu X., Li Y. The physiological and molecular mechanism of brassinosteroid in response to stress: a review // Biol. Res. 2018. V. 51. P. 46–61.
Singh I., Kumar U., Singh S.K., Gupta C., Singh M., Kushwaha S.R. Physiological and biochemical effect of 24-epibrassinoslide on cold tolerance in maize seedlings // Physiol. Mol. Biol. Plants. 2012. V. 18. P. 229–236.
Будыкина Н.П., Шибаева Т.Г., Титов А.Ф. Влияние Эпина-экстра – синтетического аналога 24-эпибрассинолида на стрессоустойчивость и продуктивность растений огурца (Сucumis sativus L.) // Тр. Карел. НЦ РАН. 2012. № 2. С. 47–55.
Ефимова М.В., Мануйлова А.В., Малофий М.К., Карташов А.В., Кузнецов Вл.В. Влияние брассиностероидов на формирование защитных реакций проростков рапса в условиях засоления // Вестн. Томск. гос. ун-та. Биология. 2013. № 1 (21). С. 18–128.
Колупаев Ю.Е., Вайнер А.А. Механизмы стресспротекторного влияния брассиностероидов на растения // Агрохимия. 2014. № 7. С. 69–84.
Cui L., Zou Z., Zhang J., Zhao Y., Yan F. 24-Epibrassinoslide enhances plant tolerance to stress from low temperatures and poor light intensities in tomato (Lycopersicon esculentum Mill.) // Funct. Integr. Genomics. 2016. V. 16. P. 29–35.
Вакуленко В.В. Регуляторы роста растений повышают стрессоустойчивость культур // Защита и карантин раст. 2015. № 2. С. 3–14.
Tanveer M., Shahzad B., Sharma A., Khan E.A. 24-Epibrassinolide application in plants: An implication for improving drought stress tolerance in plants // Plant Physiol. Biochem. 2019. V. 135. P. 295–303.
Мухортов С.Я. Частное овощеводство и грибоводство. Воронеж: ФГБОУ ВО Воронежский ГАУ, 2018. 247 с.
Гладышева О.В. Влияние Эпин-экстра и Циркона на энергию прорастания и лабораторную всхожесть семян некоторых сортов томата // Научн. альманах. 2019. № 2-2(52). С. 115–118.
Селиванова М.В., Романенко Е.С., Сосюра Е.А., Есаулко Н.А., Айсанов Т.С. Продуктивность томата при применении микроэлементов и биологически активных веществ // Овощи России. 2017. Т. 4. С. 91–95.
Деревщюков С.Н., Сычева С.В. Применение регуляторов роста при выращивании томата // Защита и карантин раст. 2007. № 11. С. 37–38.
Мухортов С.Я. Динамика адаптивной способности агроценозов томата при применении регуляторов роста // Плодоводство и ягодоводство России. 2014. Т. 40. № 1. С. 217–220.
Мухортов С.Я., Стазаева Н.В., Микулина Ю.С., Воробьев П.Н. Адаптивные свойства агроценозов при применении фитогормонов // Вестн. Воронеж. ГАУ. 2016. № 2(49). С. 66–73.
Nazir F., Hussain A., Fariduddin Q. Interactive role of epibrassinolide and hydrogen peroxide in regulating stomatal physiology, root morphology, photosynthetic and growth traits in Solanum lycopersicum L. under nickel stress // Environ. Exp. Bot. 2019. V. 162. P. 479–495.
Yuan G.F., Jia C.G., Li Z., Sun B., Zhang L.P., Liu N., Wang Q.M. Effect of brassinosteroids on drought resistance and abscisic acid concentration in tomato under water stress // Sci. Horticult. 2010. V. 126. P. 103–108.
Khan T., Fariduddin Q., Yusuf M. Lycopersicon esculentum under low temperature stress: an approach toward enhanced antioxidants and yield // Environ. Sci. Pollut. Res. 2015. V. 22. P. 14178–14188.
Aghdam M.S., Asghari M., Farmani B., Mohayeji M., Moradbeygi H. Impact of postharvest brassinosteroids treatment on PAL activity in tomato fruit in response to chilling stress // Sci. Horticult. 2012. V. 144. P. 116–120.
Shahbaz M., Ashraf M., Athar H.R. Does exogenous application of 24-epibrassinolide ameliorate salt induced growth inhibition in wheat (Triticum aestivum L.)? // Plant Growth Regul. 2008. V. 55. P. 51–64.
Janeczko A., Pociecha E., Dziurka M., Jurczyk B., Libik-Konieczny M., Oklestkova J., Novak O., Pilarska M., Filek M., Rudolphi-Skorska E., Sadura I., Siwek A. Changes in content of steroid regulators during cold hardening of winter wheat – steroid physiological/biochemical activity and impact on frost tolerance // Plant Physiol. Biochem. 2019. V. 139. P. 215–228.
Прусакова Л.Д., Чижова С.И., Агеева Л.Ф., Голанцева Е.Н., Яковлев А.Ф. Влияние эпибрассинолида и экоста на засухоустойчивость и продуктивность яровой пшеницы // Агрохимия. 2000. № 3. С. 50–54.
Сычева С.В., Бухаров А.Ф., Деревщюков С.Н., Востриков В.В. Селекция томата для открытого грунта Центрально-Черноземного Региона // Картофель и овощи. 2017. № 4. С. 37–40.
Wellburn A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution // J. Plant Physiol. 1994. V. 144. P. 307–313.
Lichtenthaler H.K. Chlorophylls and carotenoids pigments of photosynthetic biomembranes // Methods in Enzymol. V. 148 / Ed. Colowick S.P., Kaplan N.O. San-Diego E.A., 1987. P. 350–382.
Лукаткин А.С., Голованова В.С. Интенсивность перекисного окисления липидов в охлажденных листьях теплолюбивых растений // Физиология растений. 1988. Т. 35. С. 773–780.
Лукаткин A.C., Нарайкина Н.В. Оценка воздействия температурных стрессоров на растения кукурузы по изменению антиоксидантной активности // Докл. РАСХН. 2011. № 5. С. 8–10.
Kwon S.W., Hong S.S., Kim J.I., Ahn I.H. Antioxidant properties of heat-treated Hibiscus syriacus // Изв. РАН. Сер. биол. 2003. Т. 30. С. 20–21.
Кондратенко Е.И., Великородов А.В., Мохамад А.Э.С.А., Ломтева Н.А., Кондратенко К.Н. Химический состав и антиоксидантная активность экстрактов семян Nelumbo nucifera // Химия раст. сырья. 2012. № 3. С. 115–120.
Хрипач В.А., Лахвич Ф.А., Жабинский В.Н. Брассиностероиды. Минск: Наука и техника, 1993. 287 с.
Лукаткин А.С., Каштанова Н.Н., Духовскис П. Влияние эпибрассинолида на термоустойчивость проростков кукурузы // Агрохимия. 2013. № 6. С. 24–31.
Яхин О.И., Лубянов А.А., Калимуллина З.Ф., Яхин И.А., Вахитов В.А., Чемерис А.В., Гималов Ф.Р., Матниязов Р.Т. Антистрессовая активность регулятора роста растений Эпина-экстра // Докл. РАСХН. 2009. № 3. С. 25–27.
Li J., Yang P., Gan Y., Yu J., Xie J. Brassinosteroid alleviates chilling-induced oxidative stress in pepper by enhancing antioxidation systems and maintenance of photosystem II // Acta Physiol. Plant. 2015. V. 37. P. 222.
Shu S., Tang Y., Yuan Y., Sun J., Zhong M., Guo S. The role of 24-epibrassinolide in the regulation of photosynthetic characteristics and nitrogen metabolism of tomato seedlings under a combined low temperature and weak light stress // Plant Physiol. Biochem. 2016. V. 107. P. 344–353.
Li J., Yang P., Kang J., Gan Y., Yu J., Calderyn-Urrea A., Jian L., Zhang G., Feng Z., Xie J. Transcriptome analysis of pepper revealed a role of 24-epibrassinolide in response to chilling // Front. Plant Sci. 2016. V. 7. P. 1–16.
Miliauskas G., Venskutonis P.R., Van Beek T.A. Screening of radical scavenging activity of some medicinal and aromatic plant extracts // Food Chem. 2004. V. 85. P. 231–237.
Дополнительные материалы отсутствуют.