

Акустический журнал, 2019, T. 65, № 6, стр. 853-860
Некоторые аспекты отражения ЧМ-сигналов (свистов) дельфинов в экспериментальном бассейне
В. А. Рябов *
Каpадагcкая научная станция им. Т.И. Вяземского – природный заповедник РАН
298188 Крым, п. Курортное, Феодосия, ул. Науки 24, Россия
* E-mail: ryabofff@inbox.ru
Поступила в редакцию 12.10.2018
После доработки 14.06.2019
Принята к публикации 09.07.2019
Аннотация
Впервые для зубатых китов записаны отражения ЧМ-сигналов (свистов) двух черноморских дельфинов афалин (Tursiops truncatus) от границ бассейна с размерами во много раз меньше протяженности сигналов в пространстве. Показано, что дельфины продуцируют “свисты” с уровнями звукового давления (около 2 Па), лишь незначительно (на 3–8 дБ) превышающими амплитуды акустических шумов бассейна. Установлено, что при распространении ЧМ-сигналы многократно отражаются от границ бассейна, форма излученного сигнала искажается в результате интерференции с сигналами, отраженными от границ бассейна и акустическими шумами. В отличие от временной области, спектр ЧМ-сигнала более устойчив к влиянию интерференции и поэтому отношение сигнал/шум спектральных плотностей мощности (СПМ) этих же сигналов и их отражений достигает 30–40 дБ. Полученные результаты указывают на преимущества анализа ЧМ-сигналов и их эха в спектральной области.
ВВЕДЕНИЕ
Акустические сигналы зубатых китов разнообразны и являются основным средством, необходимым им для осуществления сложного согласованного социального поведения [1]. Следует специально отметить, что в условиях недостаточной видимости эти сигналы становятся единственным сенсорным посредником животных. Однако, в научной литературе акустические сигналы животных описываются качественно – “щелчки, жужжание, скрип, кряки, хлопки, лай, визг, свист” и т.д. [2–4] и др.
До настоящего времени общепринято мнение, что у Odontoceti есть только один сонар. Зондирующие сигналы сонара – “щелчки”, или ультракороткие сверхширокополосные импульсы [5–7]. Эхолокационные “щелчки” бутылконосых дельфинов очень короткие, длительностью около 50 мкс, поэтому широкополосные и охватывают частоты 2–200 кГц с максимумом энергии на частотах около 120–130 кГц [8–10].
В то же время, были высказаны предположения о том, что ЧМ-сигналы (известные как “свисты”) дельфины могут использовать: в различных поведенческих контекстах [11, 12], для идентификации себя “свистами‑автографами” [13, 14], при определении направления движения “свистящего” дельфина [15], для поддержания акустического контакта между отдельными рассеянными особями в пространстве [16–21] на расстояниях до 10.5–25 км [17, 21].
ЧМ-сигналы зубатых китов изучаются с середины прошлого века, однако в большинстве работ регистрация сигналов выполнена в полосе частот только до 20 кГц. Характеристики сигналов и их функциональность не рассмотрены в свете теории сигналов и эхолокации. Эти недостатки в некоторой степени были восполнены в работах [5, 6], где они рассматриваются как многочастотные ЧМ-сигналы. У афалины эти сигналы охватывают область частот до 140 кГц, произведение длительности сигнала на ширину его спектра достигает 104–105. Результаты этих работ дают основания полагать, что “свисты” являются зондирующими сигналами ЧМ-доплеровского сонара и ЧМ-сонара со сжатием импульса, или CHIRP (compressed high intensity radiated pulse) сонара дельфинов.
Цель работы – изучение некоторых аспектов отражения ЧМ-сигналов черноморских дельфинов вида афалина (Tursiops truncatus) в условиях бассейна с размерами намного меньшими протяженности сигналов в пространстве.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Метод
Акустические сигналы двух взрослых черноморских дельфинов (Tursіops truncatus) c кличками “Яша” (самец) и “Яна” (самка) были записаны в закрытом бетонном бассейне размерами 27.5 × 9 × 4.5 м, ФГБУН “Каpадагcкая научная станция им. Т.И. Вяземского – природный заповедник РАН”. Дельфины находятся в бассейне около 25 лет и имеют нормальный слух. Сигналы регистрировались двухканальной системой записи (рис. 1). Расстояние между гидрофонами I и II каналов записи было выбрано 5 м, для получения достаточных межканальных различий уровней звукового давления (УЗД) и временных задержек каждого сигнала. Принадлежность записанных сигналов конкретным дельфинам установлена с учетом межканальных различий временных задержек и межканальных различий УЗД каждого сигнала, а также с учетом расстояний между дельфинами, гидрофонами и границами бассейна. Глубина погружения гидрофонов – 1 м. Гидрофон I канала был расположен ближе к середине бассейна (рис. 1, I), гидрофон II канала (рис. 1, II) находился близко у стенки бассейна. В некоторых случаях положение дельфинов в бассейне относительно гидрофонов регистрировала кинокамера, синхронно с записью сигналов.
Рис. 1.
Конфигурация эксперимента. I и II – гидрофоны I и II каналов записи, соответственно. 1 и 2 – дельфины, 3 – мостки. Ширина мостков – 3 м. Расстояние между гидрофонами – 5 м. Глубина гидрофонов – 1 м. Уровень воды – 4 м. Расстояние гидрофона II от стенки бассейна – 0.35 м.

Гидрофоны (I и II) – сферические, диаметром 14 мм, изготовлены из пьезокеpамики, имеют калиброванную чувствительность –203.5 и –206 дБ относительно 1 В/мкПа, или 66.5 и 50 мкВ/Па, соответственно. Частотная характеристика гидрофонов имела неравномерность ±3 дБ до частот около 160 кГц и ±10 дБ до частот около 220 кГц. Каждый канал записи сигналов состоял из гидрофона, фильтра верхних частот (0.1 кГц), усилителя напряжения (40 дБ) и одного из каналов многоканального 14-pазpядного аналого-цифрового преобразователя (АЦП) USB-3000. Динамический диапазон АЦП и тракта регистрации сигналов около 81 дБ (0.1–1600 Па), частота дискретизации каждого канала АЦП – 1 МГц. Оцифрованные сигналы дельфинов c АЦП записаны на жесткий диск ноутбука. Пакеты программ Power-Graph 3.3.8 и Adobe Audition 3.0 были использованы для записи, визуализации и обработки сигналов. Спектральные плотности мощности (СПМ) ЧМ-сигналов дельфинов во времени (спектрограммы) были рассчитаны c использованием быстрого преобразования Фурье на 4096 точек.
Эксперимент выполнялся без специальной дрессировки и без пищевого подкрепления дельфинов, в дневное время. Дельфины продуцировали акустические сигналы, свободно передвигаясь по бассейну, по-видимому, не подозревая, что их записывают. Всего было сделано 20 записей акустических сигналов дельфинов, одна запись в день, длительность каждой записи около 30 мин. Всего было записано несколько тысяч акустических сигналов дельфинов, пяти различных классов, в соответствии с работой [6]. ЧМ-сигналы были выбраны для анализа и обсуждения. Во время записи акустических сигналов дельфинов других животных в бассейне не было.
РЕЗУЛЬТАТЫ
Среди записанных акустических сигналов было около трехсот ЧМ-сигналов дельфинов. УЗД свистов обычно не превышал 2–3 Па, в редких случаях до 10 Па. Основная частота записанных ЧМ-сигналов изменялась от 3.5 до 42 кГц. Число гармоник могло составлять от единиц до нескольких десятков. Полоса частот ЧМ-сигналов вместе с гармониками могла составлять от 15 до 140 кГц. Спектрограммы ЧМ-сигналов описывали линейно или нелинейно возрастающие, или убывающие по частоте кривые разной крутизны, и их комбинации с повторениями. Эти кривые принято называть контуром свиста. В целом, формы контуров ЧМ-сигналов, как и основные частоты и соответственно частоты и количество гармоник сигналов, а также их длительность, согласуются с представленными в работах [3, 13–18, 21–23].
Кроме того, были зарегистрированы широкополосные шумоподобные ЧМ-сигналы с числом гармоник более 50 и максимальным УЗД около 10 Па (приведенные к 1 м от дельфина). Эти сигналы имеют достаточный уровень энергии в области звуковых частот, поэтому при воспроизведении человек хорошо слышит их как шумовые импульсы [6].
При визуальном анализе спектрограмм записанных ЧМ-сигналов дельфинов обнаружено их многократное эхо (рис. 2–4). Отражения повторяют форму контуров изменения основной частоты и гармоник ЧМ-сигналов, с различной временной задержкой относительно них (табл. 1). Задержка каждого последующего эха в целое число раз кратна задержке между соответствующим сигналом и первым эхо. В качестве примера на рис. 2 показан ЧМ-сигнал, продуцированный на ходу дельфином Яша вблизи (около 2–3 м) гидрофона I канала (обозначено 1 на рис. 1). Временные задержки между отражениями сигнала составляют около 9–12 мс (рис. 2, табл. 1). В этом случае уровень СПМ ЧМ-сигнала на I канале приблизительно на 20 дБ больше, чем на II канале. Число гармоник сигнала на I канале семь, на II канале – четыре. Полоса частот этого ЧМ-сигнала с гармониками около 119 кГц. На рис. 2 показана только область спектрограммы, содержащая эхо ЧМ-сигнала. Таблица учитывает только гармоники, относительно которых измерены временные задержки эхо.
Рис. 2.
ЧМ-сигнал и четыре “щелчка”, продуцированные дельфином Яша (спектрограмма). Расстояние от дельфина до гидрофона (I канала) 2–3 м. I и II – гидрофоны первого и второго каналов записи сигналов соответственно.

Рис. 3.
ЧМ-сигнал и “щелчки”, продуцированые дельфином Яна (спектрограмма). Расстояние от дельфина до гидрофонов около 12–15 м. I и II – гидрофоны первого и второго каналов записи сигналов соответственно.
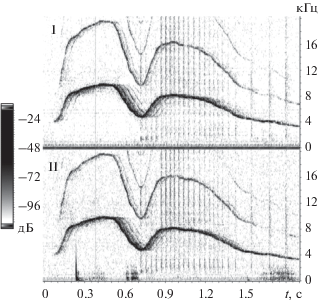
Рис. 4.
Два ЧМ-сигнала, продуцированные Яной (спектрограмма). Расстояние от дельфина до гидрофонов около 18–21 м. I и II – гидрофоны первого и второго каналов записи сигналов соответственно.

Рис. 5.
ЧМ-сигналы, показанные на рис. 4 (временная область). Ось абсцисс – время t, с. Ось ординат – уровень звукового давления в дБ относительно 1000 Па. I и II – гидрофоны первого и второго каналов записи сигналов соответственно.

Таблица 1.
Основные характеристики эхо ЧМ-сигналов дельфинов
| Расстояние от дельфина до гидрофонов, м | 2–3 | 12–15 | 18–21 | |||||
|---|---|---|---|---|---|---|---|---|
| Канал регистрации | I | II | I | II | I | II | I | II |
| первый ЧМ-сигнал | второй ЧМ-сигнал |
|||||||
| Число эхо основной частоты | 1 | 3 | 4 | 4 | 4 | 2 | 5 | 3 |
| Задержки эха основной частоты, мс | 12.8 ± 0.21 | 12.11 ± 0.19 | 35.95 ± 0.35 | 31 ± 1.3 | 35.23 ± 1.05 | 35.62 ± 0.65 | 34.7 ± 1.25 | |
| Число гармоник ЧМ-сигнала |
5 | 3 | 1 | 1 | 1 | 1 | ||
| Задержки эха гармоник, мс | 9.02 ± 0.32 | 12.18 ± 0.016 | 36 ± 0.35 | 36 ± 0.41 | 31.2 ± 0.65 | 33.94 ± 1.85 | ||
На рис. 3 в качестве примера показана спектрограмма ЧМ-сигнала Яны, который она продуцировала в движении с расстояния до гидрофонов около 12–15 м, как приблизительно обозначено 2 на рис. 1. В этом случае в области основной частоты спектрограммы сигнала, на I канале записи зарегистрированы четыре эха (табл. 1) с временной задержкой относительно основной частоты около 36 мс (первое эхо), 72 мс (второе эхо), 108 мс (третье эхо) и 144 мс (четвертое эхо). На втором канале зарегистрированы четыре эха с задержками, кратными около 31 мс. Задержка между эхами в целое число раз кратна задержке между основной частотой сигнала и первым эхо. В области первой гармоники свиста также зафиксировано ее отражение. Межканальных различий уровней СПМ сигнала не заметно.
На рис. 4 показан пример спектрограммы двух ЧМ-сигналов, продуцированных на ходу Яной с максимально возможного в нашем бассейне расстояния до гидрофонов, около 18–21 м. В области основной частоты и первых гармоник сигналов зарегистрировано различное количество эхо с различной временной задержкой (табл. 1). Межканальных различий уровней СПМ сигналов не заметно. Анализ записей акустических сигналов показал, что в частотной области ЧМ-сигналы дельфинов не имеют деформаций в результате интерференции с их отражениями. Более того, СПМ ЧМ-сигналов значительно больше (на 30–40 дБ) СПМ акустических шумов бассейна (рис. 2–4). В отличие от этого, во временной области эти же ЧМ-сигналы (рис. 4, 5) с трудом отличимы от акустических шумов бассейна. Амплитуда УЗД шума на I и II каналах составляет около –72 и –70 дБ относительно 1000 Па (рис. 5) соответственно. Амплитуда УЗД первого и второго ЧМ-сигнала превышает амплитуду УЗД шума на 3–5 дБ и около 8 дБ соответственно. Максимальные значения УЗД в начале и конце этих ЧМ-сигналов практически совпадают с максимальными уровнями окружающего акустического шума бассейна и только ближе к середине сигналов их УЗД достигают максимальных значений, превышающих уровень шумов на 3–8 дБ. Отражения ЧМ-сигналов во временной области визуально не обнаружены.
ОБСУЖДЕНИЕ
В настоящей работе отражения ЧМ-сигналов дельфинов зарегистрированы впервые для зубатых китов. Неожиданным также оказался тот факт, что эхосигналы четко видны в бассейне, размеры которого намного меньше протяженности сигналов в пространстве. Учитывая эти наиболее интересные результаты работы, обсудим некоторые аспекты отражения ЧМ-сигналов дельфинов в бассейне и возможности их анализа.
Учитывая, что длина бассейна около 27.5 м, а ширина в области расположения гидрофонов – около 9 м и скорость звука около 1500 м/с, время двойного пути звука между дальними стенками бассейна составит около 36 мс, и между боковыми стенками в области гидрофонов – около 12 мс.
Эхо ЧМ-сигналов дельфинов с различными временными задержками, меньшими 30 мс, по-видимому, представляют собой суперпозицию многочисленных эхо, которые видны на этих рисунках как утолщение линии контура сигнала и ее расширение, с разным уровнем серого (рис. 2–4). В этой области временных задержек, по-видимому, сосредоточено максимальное число однократных отражений ЧМ-сигналов и их гармоник, среди которых эхо от поверхности воды, дна и стенок бассейна. Эхо с большими временными задержками, очевидно, представляют собой многократные отражения ЧМ-сигналов от границ бассейна, при их распространении вдоль или поперек бассейна. Следовательно, задержки эхо, близкие к 36 и 12 мс (табл. 1), указывают на то, что эхосигнал проходит двойной путь вдоль или поперек бассейна, соответственно, каждый раз фиксируясь гидрофонами обоих каналов.
Анализ записей указывает также на то, что с увеличением расстояния (рис. 2–4) число наиболее высокочастотных гармоник и межканальная разница СПМ ЧМ-сигналов уменьшаются. Эти факты указывают на большую степень затухания высокочастотных составляющих ЧМ-сигналов и их направленность.
На рис. 4, 5 в качестве примера показаны одни и те же ЧМ-сигналы в частотной (спектрограмма) и временной области. Максимальные амплитуды УЗД этих сигналов достигают только 0.25–0.5 Па и лишь незначительно превышают УЗД акустических шумов бассейна. Протяженность рассматриваемых ЧМ-сигналов в пространстве (учитывая их длительность tw = 0.5–1.8 с и скорость звука в воде с0 ≈ 1500 м/с) составляет около Lw ≈ twсо ≈ ≈ 750–2700 м, что в десятки раз больше размеров экспериментального бассейна. ЧМ-сигналы дельфинов имеют низкие уровни звукового давления, близкие к уровню шумов бассейна (рис. 5). Распространяясь в бассейне, они многократно отражаются от границ бассейна и интерферируют со своими отражениями. Известно, что форма сигналов вследствие многократной суперпозиции с эхо значительно деформируется во временной области. Вместе с тем, их эхо существенно маскируется протяженными ЧМ-сигналами, имеющими больший уровень, и акустическими шумами бассейна, что значительно затрудняет анализ эхосигналов во временной области.
В отличие от этого, как следует из результатов настоящей работы, в частотной области ЧМ-сигналы не имеют деформаций в результате интерференции с отражениями. Маскировка отражений сигналами отсутствует и СПМ ЧМ-сигналов и их отражений значительно больше (на 30–40 дБ) СПМ акустических шумов бассейна (рис. 2–4). Более того, ЧМ-сигналы имеют длительность на три-четыре порядка большую длительности импульсных сигналов дельфина, что обеспечивает им относительно большую энергию, это подтверждается их значительным пробегом и многократными отражениями в бассейне (рис. 2–4). Учитывая результаты настоящей работы, можно полагать, что дельфины анализируют ЧМ-сигналы и их эхо в частотной области. В пользу этого также свидетельствует низкий уровень сигналов, соизмеримый с уровнем шума во временной области. Если предположить, что дельфин анализирует эхо ЧМ-сигналов, то возникает вопрос – какой механизм анализа может использовать для этого дельфин? В данном случае представляет интерес анализ временных задержек отражений ЧМ-сигналов. Для ответа на этот вопрос рассмотрим ЧМ-сигналы (свисты) в свете теории и техники сложных сигналов.
Акустические сигналы с произведением длительности на ширину спектра TW $ \gg $ 1 называют сложными, или шумоподобными, а также сигналами с расширенным спектром. Большая величина TW, где T – средняя длительность “свиста”, W – средняя ширина его спектра, означает сложную структуру и большое информационное содержание сигналов, тогда как малая величина этого произведения характеризует простые сигналы. Произведение TW ЧМ-сигнала дельфина, при учете только девиации основной частоты сигнала достигает 10 251, а с учетом гармоник сигнала – около 49 000, и еще больше у шумоподобных “свистов” – около 108 000 [6]. Сложные акустические сигналы дельфина в процессе эволюции развивались и совершенствовались на протяжении десятков млн лет вместе с его эхолокационной системой и слухом. Следовательно, оптимальная эффективность этих систем определяется физическими характеристиками сигналов и методами их обработки дельфином [6]. Преимущества, полученные эхолокационной системой дельфина от использования сложных сигналов и соответствующих им методов обработки, можно объяснить, исходя из известных концепций теории сложных сигналов.
Для оптимального приема эхосигналов на фоне шума в технике эхолокации используют согласованную фильтрацию или оптимальный корреляционный прием. В этом случае современные эхолокационные системы со сложными сигналами получают повышенную помехоустойчивость, которая определяется известным соотношением
где ρ2= Pс/Pп – отношение мощностей сигнал/помеха на входе приемника, q2= 2E/Nп – отношение сигнал/помеха на выходе согласованного фильтра или корреляционного приемника, где E – энергия сигнала, Nп – спектральная плотность мощности помехи в полосе частот сигнала. Для этого достаточно выбрать сложный сигнал с достаточным произведением TW (1). Прием сложных сигналов согласованным фильтром или корреляционным приемником усиливает сигналы в 2TW раз. Величину K = q2/ρ2 называют коэффициентом усиления сложного сигнала при обработке. Соотношение (1) является фундаментальным в теории систем связи со сложными сигналами. Вместе с тем, сложные сигналы используются в эхолокаторах при одновременном измерении дальности цели R и доплеровского сдвига эхосигнала fd.Точность измерения и разрешающая способность эхолокатора по дальности R возрастает с расширением полосы частот сложного сигнала и с возрастанием отношения сигнал/помеха q2. Чем больше q2 и длительность сигнала T, тем выше точность измерения и разрешающая способность определения относительной радиальной скорости Vr.
У сложных сигналов можно независимо изменять ширину спектра W и длительность T, при совместном измерении дальности R и относительной радиальной скорости цели Vr. Другими словами, форма контура свиста может быть различной, необходимо лишь, чтобы она соответствовала амплитудно-частотной и фазово-частотной характеристикам согласованного фильтра слуха для решения соответствующей эхолокационной задачи. Поэтому при анализе свистов обычным слухом различную форму контура свиста дельфины могут использовать в контексте задач, рассмотренных в работах [11–21]: как контекстно-специфические сигналы, свисты‑автографы, для определения ориентации и направления движения сигнализирующего дельфина, а также для установления акустического контакта, координации действий и поддержания сплоченности между рассеянными животными в группе.
При обработке сложного сигнала согласованным фильтром происходит сжатие сигнала во времени и его усиление вследствие синфазного сложения составляющих сигнала, поэтому можно обойтись без усилителя мощности сигнала. Важно, что сжатие импульса позволяет использовать относительно большую энергию длинного импульса с временным разрешением как у короткого импульса. Отклик согласованного фильтра для ЧМ-сигнала длительностью Т и шириной спектра W представляет собой узкий импульс или центральный пик и боковые импульсы или боковые пики. Амплитуда центрального пика V, его длительность τ0≈ 1/W. Амплитуда боковых импульсов vmax. Чем шире спектр сложного сигнала, тем короче длительность центрального пика. Сложные сигналы с большим произведением TW обладают свойствами, которые записываются следующим образом:
(3)
${V \mathord{\left/ {\vphantom {V {{{{v}}_{{{\text{max}}}}}}}} \right. \kern-0em} {{{{v}}_{{{\text{max}}}}}}} \approx {\text{ }}{{(\alpha TW)}^{{1{\text{/}}2}}},$Соотношение (2) определяет сжатие сложных сигналов – отношение длительности сигнала к длительности центрального пика. Сжатие приблизительно равно произведению TW сложного сигнала.
Соотношение (3) характеризует подавление боковых пиков. Чем больше TW сложного сигнала, тем больше подавление боковых пиков. Чем уже центральный пик АКФ зондирующего сигнала сонара, тем выше разрешающая способность сонара по времени – доплеровскому смещению и точности измерения дальности цели и радиальной скорости.
Отметим, что согласованный фильтр и корреляционный приемник эквивалентны с точки зрения обнаружения сигнала. Однако коррелятор является устройством с переменными во времени параметрами и может гибко подстраиваться под изменяющийся сигнал (как активный фильтр), тогда как согласованный фильтр настроен на один определенный сигнал. Тем не менее, вопрос о применении каждого устройства определяется в зависимости от простоты реализации.
Каким же образом может быть организован согласованный фильтр в слухе дельфина? Этот вопрос не простой и, очевидно, потребует дальнейших исследований. Однако можно предположить, что каждый раз при продуцировании ЧМ-сигнала (свиста) слух дельфина организует согласованный с конкретным сигналом фильтр слуха, с соответствующими амплитудно-частотной и фазо-частотной характеристиками. Поэтому все отражения сигнала, как зондирующего сигнала сонара дельфина со сжатием импульса или ЧМ-доплеровского сонара, будут фиксироваться на выходе этого фильтра в виде отдельных откликов, с разной временной задержкой (определяемой дальностью отражателей, и их относительной радиальной скоростью). Временное разрешение между откликами определяется произведением TW ЧМ-сигнала. Важно, что в этом случае существенно повышается помехозащищенность этих сонаров. Каждый дельфин может продуцировать различные типы своих сигналов одновременно в совместной полосе частот [6]. В этих условиях возникают значительные перекрестные помехи (как между сигналами одного дельфина, так и сигналами сородичей), поэтому анализ согласованным фильтром слуха является оптимальным средством борьбы с перекрестной помехой и другими помехами, некоррелированными с сигналом. При этом эхолокационная система дельфина будет утилизировать рассмотренные выше преимущества использования сложных сигналов.
Исходя из характеристик рассмотренных выше ЧМ-сигналов наших дельфинов, при TW = = 104–105 коэффициент усиления сложного сигнала при обработке, исходя из (1), K = q2/ρ2 = = 2TW ≈ 104–105. При этом сигнал длительностью Т (в нашем случае Т ≈ 0.5–1.8 с) сожмется до длительности отклика согласованного фильтра слуха и, исходя из (2), будет составлять τ0≈ Т/TW ≈ ≈ (0.5–1.8)/(104–105) ≈ 18–50 мкс. Отношение амплитуды центрального пика отклика V согласованного фильтра к амплитуде боковых импульсов оклика vmax, исходя из (3), в этом случае составит
При УЗД ЧМ-сигнала около 2.5 Па (приведенное к 1 м от дельфина) свисты пробегают вдоль бассейна более 300 м (рис. 4), пять раз двойной путь звука вдоль бассейна. Максимальные УЗД ЧМ-сигналов бутылконосых дельфинов, известные из литературы, – амплитудные значения – составляют около 180 дБ относительно 1 мкПа [24], и усредненные значения – около 170 дБ (rms) относительно 1 мкПа [20], соответственно, что почти в 1000 раз выше зафиксированных в бассейне (рис. 2–5). Учитывая это, максимальная дальность действия ЧМ-сонаров дельфина в открытом море, при максимальных УЗД ЧМ-сигналов и обработке эхосигналов согласованным фильтром слуха, как показали расчеты [25], может быть близкой к максимальному расстоянию акустического контакта между дельфинами 10.5–20 км [17, 20]. Характеристика направленности ЧМ-сигналов [26] и слуха дельфина [27] в этой области частот невысокая, благодаря этому ЧМ-сигналы облучают и слух соответственно принимает эхо практически из всего пространства вокруг дельфина. В то же время точность локализации источников звука на этих частотах высокая [28], достигает 1° за счет бинаурального определения положения источника звука (эхо). В свете этих данных можно полагать, что ЧМ-сонары имеют максимальную дальность действия и наилучшую разрешающую способность по времени (18–50 мкс) и соответственно по дальности 0.027–0.075 м, а также лучшую помехозащищенность среди сонаров дельфина.
Вместе с тем, зондирующие ЧМ-сигналы (свисты) предполагаемых сонаров дельфинов [5, 6] и эхолокаторов летучих мышей [29 и др.] используют одинаковую область частот 2–120 кГц, содержат гармоники, и длительность сигналов сопоставима. Подобны также контуры изменения их частоты (линейное или нелинейное возрастание, или понижение частоты от времени с разной скоростью, и их комбинации с повторениями). Принципиальных различий между ЧМ-сигналами дельфинов и летучих мышей нет. Подобные зондирующие сигналы (только без гармоник) используют ЧМ-доплеровские и ЧМ-со сжатием импульса гидролокаторы и радиолокаторы (если не учитывать несущую частоту последних). Эти сложные сигналы обладают высокими корреляционными свойствами, поэтому не удивительно, что используются эхолоцирующими млекопитающими различных отрядов царства животных (Cetacea и Chiroptera) в водной и наземно-воздушной средах, а также в технике для получения высокой точности определения дальности и доплеровского сдвига цели.
Рассмотренные в настоящей работе результаты дают дополнительные подтверждения предположению о функциональности ЧМ-сигналов дельфина в качестве зондирующих сигналов ЧМ-доплеровского сонара и ЧМ-сонара со сжатием импульса [5, 6].
При проведении исследований были соблюдены все применимые международные и национальные руководящие принципы по уходу и использованию животных, а также принципы организации, в которой выполнено исследование.
Список литературы
Lammers M.O., Schotten M., Au W.W.L. The spatial context of free-ranging Hawaiian spinner dolphins (Stenella longirostris) producing acoustic signals // J. Acoust. Soc. Am. 2006. V. 119(2). P. 1244–1250.
Watkins W.A., Wartzok D. Sensory biophysics of marine mammals // Mar. Mamm. Sci. 1985. V. 1. P. 219–260.
Caldwell M.C., Caldwell D.K. Intraspecific transfer of information via the pulsed sound in captive Odontocete Cetaceans // Animal Sonar Systems: Biology and Bionics / Ed. Busnel R.G. Laboratoire de Physiologie Acoustic, Jouy-en-Josas, France, 1967. P. 879–936.
Herzing D.L. Vocalizations and associated underwater behavior of free-ranging Atlantic spotted dolphins, Stenella frontalis and bottlenose dolphin, Tursiops truncatus // Aqua. Mamm. 1996. V. 22. P. 61–79.
Ryabov V.A. Some Aspects of Analysis of Dolphins’ Acoustical Signals // Open J. Acoustics. 2011. V. 1. P. 41–54. https://doi.org/10.4236/oja.2011.12006
Ryabov V.A. Acoustic Signals and Echolocation System of the Dolphin // Biophysics. 2014. V. 59(1). P. 135–147.
Романенко Е. В. Акустика дельфинов и рыб (обзор) // Акуст. журн. 2019. Т. 65. № 1. С. 82–92.
Дубровский Н.А., Заславский Г.Л. О роли костей черепа в формировании пространственно-временной структуры зондирующего импульса дельфина // Акуст. журн. 1975. Т 3. С. 409–414.
Au W.W.L. Echolocation signals of Atlantic bottlenose dolphin (Tursiops truncatus) in open waters // Animal sonar systems / Eds. Busnel R., Fish J. N.Y: Plenum press, 1980. P. 251–282.
Иванов М.П. Эхолокационные сигналы дельфина при обнаружении объектов в сложных акустических условиях // Акуст. журн. 2004. Т. 50. № 4. С. 550–561.
Lilly J.C. Distress call of the bottlenose dolphin: stimuli and evoked behavioral responses // Science. 1963. V. 139. P. 116–118.
Dreher J.J., Evans W.E. Cetacean communication // Marine Bioacousics / Ed. Tavolga W.N. Oxford: Pergammon Press, 1964. V. 1. P. 373–399.
Caldwell M.C., Caldwell D.K., Tyack P.L. Review of the signature-whistle hypothesis for the Atlantic bottlenose dolphin // The Bottlenose Dolphin / Eds. Leatherwood S., Reeves R.R. San Diego: Academic Press, 1990. P. 199–234.
Tyack P. Dolphins whistle a signature tune // Science. 2000. V. 289. P. 1310–1311.
Lammers M.O., Au W.W.L. Directionality in the whistles of Hawaiian Spinner dolphins Stenella Longirostris: A signal feature to cue direction of movement // Marine Mammal Sci. 2003. V. 2. P. 249–264.
Janik V.M. Whistle matching in wild bottlenose dolphins Tursiops truncatus // Science. 2000. V. 289. P. 1355–1357.
Janik V.M. Source levels and the estimated active space of bottlenose dolphin (Tursiops truncatus) whistles in the Moray Firth, Scotland // J. Comp. Physiol. A. 2000. V. 186. P. 673–680.
Rasmussen M.H., Miller L.A. Whistles and clicks from white-beaked dolphins, Lagenorhynchus albirostris, recorded in Faxaflo’i Bay, Iceland // Aqua. Mamm. 2002. V. 28. P. 78–89.
Rasmussen M.H., Miller L.A. // Echolocation in Bats and Dolphins / Eds Thomas J., Moss C., Vater M. Univ. of Chicago, Chicago, 2004. P. 50–53.
Rasmussen M.H., Lammers M.O., Beedholm K., Miller L.A. Source levels and harmonic content of whistles in white-beaked dolphins (Lagenorhynchus Albirostris) // J. Acoust. Soc. Am. 2006. V. 120(1). P. 510–517. https://doi.org/10.1121/1.2202865
Janik V., Slater P. Context-specific use suggests that bottlenose dolphin signature whistles are cohesion calls // Animal Behaviour. 1998. P. 829–838.
Беликов Р.А., Белькович В.М. Свисты белух в репродуктивном скоплении у острова Соловецкий в Белом море // Акуст. журн. 2007. Т. 55. С. 601–608.
Беликов Р.А., Белькович В.М. Высокочастотные тональные сигналы белух (Delphinapterus leucas) в летнем скоплении у острова Соловецкий в Белом море // Акуст. журн. 2006. Т. 52. № 2. С. 156–164.
Richardson W.J., Greene C.R., Malme C.I., Thomson D.H. Marine Mammals and Noise. San Diego: Academic Press, 1995. 576 p.
Рябов В.А. Гидроакустические закономерности пищевого поведения дельфинов // Морской биолог. журн. 2018. Т. 3. № 2. С. 81–97.
Branstetter B.K., Moore P.W., Finneran J.J., Tormey M.N., Aihara H. Directional properties of bottlenose dolphin (Tursiops truncatus) clicks, burst-pulse, and whistle sounds // J. Acoust. Soc. Am. 2012. V. 131(2). P. 1613–1621.
Ryabov V.A., Polyakov M.A., Bibikov N.G. Characterization of beam patterns of bottlenose dolphin in the transverse plane // Biophysics. 2011. V. 56. № 3. P. 529–534.
Popov V.V., Supin A.Ya., Klishin V.O., Bulgakova T.N. Monaural and binaural hearing directivity in the bottlenose dolphin: Evoked-potential study // J. Acoust. Soc. Am. 2006. V. 119. P. 636–644.
Mayberry H.W., Faure P.A. Morphological, olfactory, and vocal development in big brown bats // Biology Open. 2015. V. 4. P. 22–34. https://doi.org/10.1242/bio.201410181
Дополнительные материалы отсутствуют.
Инструменты
Акустический журнал