

Биоорганическая химия, 2021, T. 47, № 5, стр. 566-572
Зазеркалье биотехнологии: D-белки как объекты патентной охраны
И. Б. Никитина 1, *, И. В. Горетова 1, И. В. Федосеев 1
1 Федеральный институт промышленной собственности
125993 Москва, Бережковская наб., 30/1, Россия
* E-mail: otd1331@rupto.ru
Поступила в редакцию 10.11.2020
После доработки 28.11.2020
Принята к публикации 01.12.2020
Аннотация
Статья посвящена патентованию белков, включающих D-изоформы аминокислот, возможным проблемам и перспективам. Ввиду удешевления процесса твердофазного синтеза и появления модифицированных рибосом, способных использовать нестандартные аминокислоты для рибосомного синтеза, технологии биосинтеза белковых конструкций с нестандартными аминокислотами в их составе относятся к наиболее перспективным и быстроразвивающимся направлениям биотехнологии, наравне с технологиями редактирования генома. В статье приведены материалы, указывающие на актуальность и интерес изобретателей к этой области. Поскольку изменение аминокислотной последовательности известного белка, состоящей из цепочки L-аминокислот, путем замены исходных L-аминокислот на D-стереоизомеры приводит к существенным и часто неожиданным функциональным и физико-химическим изменениям белка, это открывает широкие перспективы для создания новых лекарственных препаратов, ферментов, пестицидов и иных белковых продуктов, использующихся в медицине, пищевой, сельскохозяйственной и легкой промышленности. Также в статье затронута тема изменения стандартов Всемирной организации интеллектуальной собственности (ВОИС), касающихся представления последовательностей, в части отображения D-аминокислот в последовательностях согласно стандарту ST.25 или ST.26 ВОИС. Описаны потенциальные проблемы, которые могут возникать в процессе патентной экспертизы при рассмотрении подобных заявок, например, в случае идентичных последовательностей D- и L-белков, обладающих разными биологическими свойствами.
ВВЕДЕНИЕ
Как известно, олигопептиды, полипептиды и белки состоят из аминокислотных остатков, ковалентно соединенных между собой пептидными связями [1]. В рибосомном синтезе белков преимущественно принимают участие 20 стандартных, так называемых канонических аминокислот, кодируемых триплетным кодом. Расположение заместителей вокруг α-атома углерода определяет хиральность (или конфигурацию) каждого аминокислотного остатка. В случае стандартных аминокислот (кроме глицина) существуют так называемые D- и L-стереоизомеры; в изолейцине и треонине присутствует второй хиральный центр в β-атоме углерода, что дает два дополнительных стереоизомера.
В живых организмах в рибосомном синтезе белков принимают участие L-изомеры. Строение рибосом и тРНК высококонсервативно и схоже для всех живых организмов, что определяет L‑изомерную чистоту природных белков и не допускает биологический синтез белков, построенных из D-аминокислотных остатков (D-белки). В то же время в некоторых белках было обнаружено присутствие D-аминокислот, что обусловливается уже последующими посттрансляционными модификациями L-аминокислот в белковой цепи.
Исследования, проводимые в последние десятилетия, показали, что биологическая активность L- и D-энантиомеров различается, причем отличия – довольно существенные [2, 3]. Впервые данные различия были установлены для полипептидов, специфично связывающихся с лигандами, среди которых D-полипептиды и природные L-полипептиды проявляли разные свойства хиральной специфичности на определенных субстратах. В частности, D-белки не связываются с лигандами, характерными для их L-изомеров, однако связываются с соответствующими D-изомерами таких лигандов. Было показано, что это свойство D-белков, скорее всего, обусловливает их устойчивость к протеолитическим ферментам [4].
Свойства D-белков – обширная и недостаточно изученная область белковой инженерии. Исследования в этой области могут привести к обнаружению большого множества потенциальных объектов патентной защиты.
Обнаруженные различные свойства хиральной специфичности L- и D-белков и полипептидов открыли, по меньшей мере, следующие перспективы в медицинской биотехнологии: создание более устойчивых к эндогенному протеолизу форм терапевтически активных белков, выступающих основой пролонгированных форм лекарственных средств с минимальными побочными эффектами; возможность создания новых терапевтически активных белков, обладающих уникальными свойствами и не имеющих известных L-аналогов.
Было показано, что введение D-аминокислотных остатков меняет спектр активности белков. Так, на основе микробных полипептидов, содержащих D-аминокислоты, были разработаны новые типы антибиотиков [5, 6], отличающиеся высокой эффективностью и продолжительностью действия. К таким антибиотикам относятся грамицидин, бацитрацин, актиномицин, валиномицин и многие другие. Они разрушают мембраны бактериальных клеток путем формирования ионных каналов [7].
Также созданы полипептиды, состоящие исключительно из D-аминокислот, способные специфично связываться с L-белком-мишенью [8]. Так, полипептид FOXO4 DRI (энантиомер фрагмента транскрипционного фактора fork head box O4), полностью состоящий из D-аминокислот, в отличие от пептида с идентичной аминокислотной последовательностью в L-изоформе, селективно устранял стареющие клетки дозозависимым образом посредством ингибирования ASK1 (apoptosis signal-regulating kinase 1), а также значительно усиливал способность вемурафениба и траметиниба уничтожать устойчивые к терапии клетки меланомы LOX-IMVI.
Создание белков, содержащих отдельные D-аминокислоты или полностью состоящих из D-аминокислот, стало возможным благодаря развитию технологий синтеза белковых конструкций. К ним относятся удешевление процесса твердофазного синтеза и развитие технологии зеркального фазового дисплея, а также появление модифицированных рибосом, способных использовать D-аминокислоты.
Зеркальный фаговый дисплей, базирующийся на классическом фаговом дисплее, представляет собой способ проведения скрининга больших библиотек D-пептидов, способных связываться с L-белками. Если L-пептид связывается с D-мишенью, то D-пептид связывается с L-мишенью [2, 9]. При проведении скрининга последовательность ДНК, кодирующую необходимый пептид, сливают с геном белков оболочки бактериофагов и вводят в вектор. Гены слитных белков экспрессируют, продукт биосинтеза очищают и наносят на поверхность, содержащую иммобилизованные D-белки-мишени. Несвязавшиеся пептиды смывают, а оставшиеся элюируют. Бактериофаги не способны продуцировать D-пептиды, поэтому иммобилизуют именно D-белки, синтезируемые химическим путем. Принцип работы зеркального фагового дисплея схематично представлен на рис. 1.
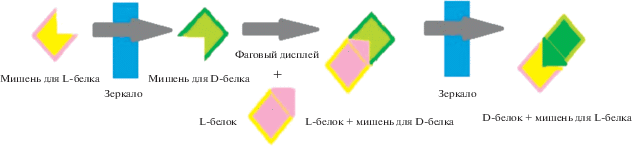
Ограничения твердофазного синтеза не позволяют создавать крупные белки, поэтому в последнее время особое внимание уделяется возможности рибосомного синтеза D-пептидов и белков. Так, исследования ведутся как в области модификации рибосом, которые позволяют последним взаимодействовать с нестандартными аминокислотами, в том числе с D-аминокислотами, например, за счет мутаций в пептидилтрансферазном центре 23S рРНК рибосом Escherichia coli [10], так и в области оптимизации тРНК, несущих необходимые D-аминокислоты, например, в части подбора соответствующих аминоацил-тРНК-синтетаз или рибозимов [11].
Таким образом, в ближайшем будущем возможно создание высокоэффективной системы рибосомного синтеза D-белков, что, естественно, будет стимулировать привлечение инвестиций для инновационных разработок и приведет к увеличению количества патентных заявок в данной области.
Цель данной работы – анализ возможностей патентования белковых объектов, включающих D-формы аминокислот, с выявлением проблематики и перспектив в данной области биотехнологий.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Открытие различий в хиральной специфичности L- и D-изомеров аминокислот стимулировало появление патентов на D-белки.
Наибольшую инновационную привлекательность обусловливает тот факт, что биологическая активность белка в D-форме, как правило, не будет идентична его биологической активности в L-форме. При этом невозможно однозначно прогнозировать характер изменений биологической активности D-энантиомеров белков только на основании гомологии их аминокислотных последовательностей с L-энантиомерами.
В качестве наглядного подтверждения можно привести результаты недавнего исследования свойств хиральной специфичности олигопептидных молекул, которые продемонстрировали неожиданное изменение биологических свойств пептидов на прямо противоположные [12]. Так, дипептиды в L-форме (L-Glu–L-Trp-OH, L‑Glu–(L-Trp-OH)-OH) и их стереоизомеры в D-форме (D-Glu–D-Trp-OH, D-Glu–(D-Trp-OH)-OH) демонстрировали противоположные свойства: L‑пептиды проявляли иммуностимулирующий эффект, в то время как D-пептиды – иммуносупрессивный.
Следовательно, весь имеющийся пласт информации о строении L-белков и о проявлении ими биологических функций, исходя из особенностей их структуры, не может быть экстраполирован на функционирование их D-изоформ при разработке новых структур D-белков, что открывает значительные перспективы в патентовании изобретений в данной области биотехнологий.
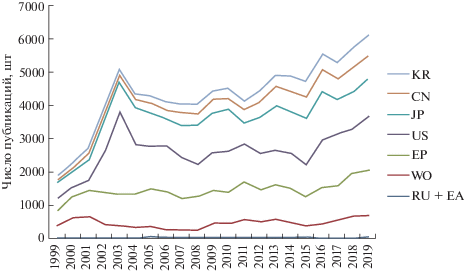
Анализ патентной активности в области изобретений, относящихся к белковым объектам, включающим D-формы аминокислот, показал стабильную динамику роста публикаций патентных документов, включая опубликованные заявки и патенты, за последние двадцать лет – с 1999 по 2019 гг. – по патентным ведомствам стран, входящих в IP-5 (пять офисов интеллектуальной собственности), а также России и международным патентным ведомствам – Всемирной организации интеллектуальной собственности (ВОИС) и Евразийскому патентному ведомству (рис. 2).
Рис. 2.
Динамика публикаций патентных документов на D-белки за 1999–2019 гг. по странам. KR – патентное ведомство Республики Корея; CN – патентное ведомство Китайской Народной Республики; JP – патентное ведомство Японии; US – патентное ведомство США; EP – Европейское Патентное Ведомство; WO – Всемирная организация интеллектуальной собственности (ВОИС); RU + EA – патентное ведомство России (Роспатент) и Евразийское патентное ведомство (ЕАПВ).
Выбранные для анализа патентной активности в рамках данной статьи патентные ведомства указанных стран наиболее полно отражают ситуацию на рынке интеллектуальной собственности. Так, ведомства IP5 совместно обрабатывают ~80% мировых патентных заявок и 95% международных заявок, поданных в соответствии с Договором о патентной кооперации (PCT). Члены IP5 – Европейское патентное ведомство (ЕПВ), Патентное ведомство Японии (JPO), Корейское ведомство интеллектуальной собственности (KIPO), Национальное управление интеллектуальной собственностью Китайской Народной Республики (CNIPA) и Ведомство США по патентам и товарным знакам (USPTO).
Выявлены два основных пика активности в начале и конце исследуемого временного периода и незначительные колебания в середине. Первый пик можно наблюдать с начала 2000-х гг., что, наиболее вероятно, связано с описанием различий в свойствах хиральной специфичности L- и D-аминокислот [2, 3], которое привело к интенсификации научных исследований в данной области биотехнологий и активной коммерциализации результатов в виде патентов на D-белки с новыми свойствами. Однако получение D-стереоизомеров белков в те годы было сопряжено со значительными трудностями их синтеза, сложно масштабируемого и требующего значительных финансовых затрат, что обусловливает спад активности и некоторую стагнацию в середине 2000-х гг.
Развитие технологий твердофазного синтеза привело к росту числа патентов о D-белках. С 2016 г. наметился значительный рост публикаций патентных документов в данной области, который сохраняется последние несколько лет. Увеличение за последние годы количества патентных документов на изобретения, относящиеся к D-белкам, связано с развитием технологий автоматизированного твердофазного синтеза полипептидов, благодаря которым становится широкодоступным создание непосредственно молекул D-полипептидов. С помощью технологии зеркального фагового дисплея можно проводить моделирование структур белков, включающих D-энантиомеры аминокислот. Также возможно создание полностью зеркальных белков, т.е. белков, характеризуемых одинаковой аминокислотной последовательностью, но отличающихся оптическими изомерами аминокислот и биологическими свойствами.
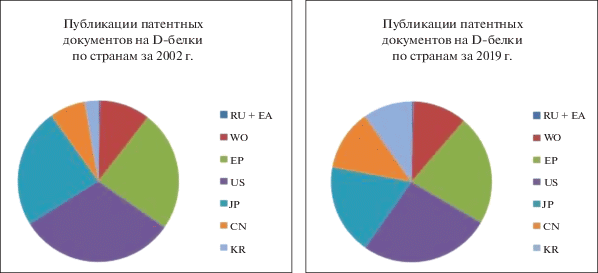
Динамика публикации патентов, посвященных D-белкам, в разных странах за последние 20 лет. Имеющиеся данные позволяют сделать вывод об огромном потенциале разработок с использованием D-белков и повышении интереса изобретателей к данной области. Так, если исходить из соотношения публикаций патентных документов, относящихся к изобретениям в области D-белков, по странам в периоды подъема активности в 2002 и 2019 гг., то можно судить о том, что бесспорные лидеры – изобретатели США. Чуть менее, но все же достаточно активны изобретатели, подающие заявки в Европейское патентное ведомство и Патентное ведомство Японии, за ними следуют Китай, Республика Корея, и, конечно, вызывает интерес подача международных заявок по системе РСТ. Можно заметить интерес к патентованию в России и на территориях, охватываемых патентами Евразийского патентного ведомства, но в значительно меньшей степени, что говорит о развивающемся рынке технологий (рис. 3).
Рис. 3.
Соотношения публикаций патентных документов на D-белки по странам в 2002 и 2019 гг. Расшифровку обозначений см. в подписи к рис. 2.
Отдельно обращает на себя внимание изменение соотношений публикационной активности по странам в периоды подъема интереса к патентованию изобретений в области D-белков, а именно в 2002 и 2019 гг., в процентах к общему числу публикаций по выбранным для анализа странам. Так, у ряда наиболее активных в патентовании D-белков стран несколько снизился интерес к данной области: США понизили активность с 32% в 2002 г. до 26% в 2019 г., как Япония – с 24 до 18% и Европа – с 24 до 22% соответственно. Однако значительно возросла активность в Республике Корея – с 3% в 2002 г. до 10% в 2019 г. и КНР – с 7 до 12%, число международных заявок также увеличилось – с 10 до 11% соответственно (рис. 4).
Рис. 4.
Патентная активность в области D-белков по странам в 2002 и 2019 гг. Расшифровку обозначений см. в подписи к рис. 2.
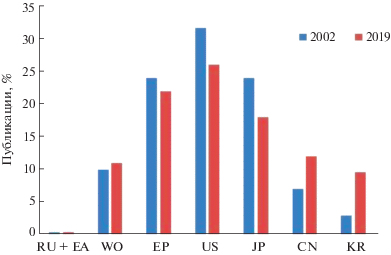
Как мы можем видеть, лидеры постепенно сдают позиции в инновационных разработках на тему зеркальных белков, уступая в росте активно развивающимся рынкам Республики Корея и КНР.
Сложности патентования D-белков. Ввиду того, что D-белки относятся к развивающейся области белковой инженерии, в процессе патентования таких объектов можно столкнуться с рядом специфических проблем, поскольку большая часть изобретений в области белковой химии и, соответственно, наработанная ранее практика патентной экспертизы были связаны преимущественно с природными L-белками, методами их синтеза и применения.
Так, основной характеризующий белок признак в формуле изобретения – его аминокислотная последовательность, которая, согласно национальному законодательству Российской Федерации (п. 53(6) и п. 51 Требований к документам заявки на выдачу патента на изобретение [13]), а также Правилу 5.2 Инструкции к Договору о патентной кооперации [14], должна быть представлена в Перечне последовательностей в описании изобретения. При этом Перечень последовательностей должен быть оформлен согласно соответствующему стандарту ВОИС.
В настоящий момент действует стандарт ST.25 ВОИС [15], и в качестве рекомендуемого указан стандарт ST.26 ВОИС [16], который вступит в силу с января 2022 г., прекратив действие предыдущего стандарта.
Согласно определению, изложенному в стандарте ST.25 ВОИС, п. 2 (iv), ““аминокислоты” – такие L-аминокислоты, которые обычно встречаются в натуральных белках и перечислены в табл. 3 Приложения 2 . Последовательности аминокислот, которые содержат, по крайней мере, одну D-аминокислоту, не предназначены для включения в данное определение”.
Следовательно, стандарт ST.25 ВОИС явно исключал D-полипептиды и D-белки из требований к перечню, предъявляемых к L-белкам, что делало необязательным предоставление аминокислотной последовательности.
Стандарт ST.26 ВОИС дает новое определение: ““Аминокислота” означает любую аминокислоту, которая может быть представлена любым из символов, указанных в Приложении I (см. раздел 3, табл. 3 ). Такие аминокислоты включают, среди прочего, D-аминокислоты и аминокислоты, содержащие модифицированные или синтетические боковые цепи. Аминокислоты будут истолкованы как немодифицированные L-аминокислоты, если они далее не описаны в таблице характеристик как модифицированные в соответствии с параграфом 30”.
Таким образом, ST.26 ВОИС, в противовес ST.25 ВОИС, вводит аналогичные требования к D-белкам, какие раньше предъявлялись только к L-белкам, с учетом указания, что аминокислоты в составе белка представлены в D-форме. Изменения, произведенные в ST.26 ВОИС, – следствие повышенного интереса изобретателей к тематике D-белков и белков, содержащих D-аминокислоты, в такой степени, что это повлекло преобразование стандартов представления материалов патентной заявки для экспертизы.
Следует отметить, что Международная патентная классификация (МПК) не содержит соответствующих рубрик для белков, включающих D-аминокислоты, в то время как в Совместную патентную классификацию (СПК) в 2013 г. введены такие рубрики к отдельным терапевтически активным белкам. Однако информационный поиск по данным рубрикам из СПК будет иметь отношение только к малому числу конкретных белков, в то время как стереохимическим модификациям могут быть подвергнуты тысячи других известных белков, рубрики по включению D-аминокислот в состав которых не предусмотрены в СПК.
Ввиду того, что в предыдущие годы большая часть заявок по новым полипептидам и белкам относилась только к L-белкам, то и существующий объем данных, касающийся их аминокислотных последовательностей, размещен в базах данных без учета оптических изомеров аминокислот в их составе, что в дальнейшем может привести к трудностям в проведении информационного поиска, связанным с проблемой в установлении отличий последовательности нового D-белка от известного L-белка с такой же аминокислотной последовательностью, но имеющего иные биологические свойства.
Учитывая вышеуказанную проблему, а также требования ST.26 ВОИС, возникает необходимость указания D-изомеров в примечаниях к предоставляемой аминокислотной последовательности, что должно быть отражено в существующих базах данных, таких как GenBank и UniProtKB/SwissProt. Отсутствие такой информации препятствует проведению корректного патентного поиска.
Кроме того, на сегодняшний день невозможно судить о биохимических свойствах D-белков по свойствам гомологичных им L-белков. Это связано с отсутствием массива информации о структурах и биологических свойствах D-белков, в отличие от природных L-белков.
Помимо этого, ввиду сложности рибосомного синтеза D-полипептидов, особенно в части оптимизации мРНК и использования стоп-кодонов, представляется проблематичной характеристика заявляемого D-белка через кодирующую его нуклеиновую кислоту ввиду невозможности однозначной интерпретации ее нуклеотидной последовательности для поиска в существующих базах данных.
В связи с переходом на ST.26 ВОИС появляются обнадеживающие перспективы представления наиболее точной и полной характеристики зеркальных белков в заявках на изобретения, с четкими обозначениями наличия аминокислот в D-форме в их аминокислотной последовательности. Данное обстоятельство в совокупности с доработкой имеющихся сведений по D-белкам в соответствующих базах данных должно в значительной степени повысить качество анализа массива данных при проведении информационного поиска и минимизировать риски нарушения объема прав изобретателей при выдаче патентов на зеркальные белки.
ЗАКЛЮЧЕНИЕ
Значительный потенциал разработок с использованием D-белков и рост интереса изобретателей к данной области биотехнологий требует своевременного и продуманного изменения подходов к экспертизе заявок, касающихся пептидной химии, связанных в том числе с подготовкой изменений не только стандартов представления материалов заявок, но и глобальных изменений поисковых систем, баз данных и классификации. Указанные изменения должны обеспечить оптимальные для всех участников рынка условия охраны изобретений на основе D-белков, что позволит снизить вероятность конфликта интересов и финансовые потери в результате оспаривания патентов.
Список литературы
Elliott W.H., Elliott D.C. // Biochemistry and Molecular Biology. Oxford: Oxford University Press, 2014. 648 p.
Schumacher T.N.M., Mayr L.M., Minor D.L., Milhollen M.A., Burgess M.W., Kim P.S. // Science. 1996. V. 271. P. 1854–1857. https://doi.org/10.1126/science.271.5257.1854
Milton R., Milton S., Kent S. // Science. 1992. V. 256. P. 1445–1448. https://doi.org/10.1126/science.1604320
Feng Z., Xu B. // Biomol. Concepts. 2016. V. 7. P. 179–187. https://doi.org/10.1515/bmc-2015-0035
Genchi G. // Amino Acids. 2017. V. 49. P. 1521–1533. https://doi.org/10.1007/s00726-017-2459-5
Adaligil E., Patil K., Rodenstein M., Kumar K. // ACS Chem. Biol. 2019. V. 14. P. 1498–1506. https://doi.org/10.1021/acschembio.9b00234
Chattopadhyay A., Kelkar D.A. // J. Biosci. 2005. V. 30. P. 147–149. https://doi.org/10.1007/BF02703693
de Keizer P.L.J. // International Application WO 2016/118014 A2, 28.07.2016.
Wiesehan K., Willbold D. // ChemBioChem. 2003. V. 4. P. 811–815. https://doi.org/10.1002/cbic.200300570
Jewett M.C., D’aquino A.E. // International Application WO 2020/010356 A1, 09.01.2020.
Terasaka N., Iwane Y., Geiermann A.S., Goto Y., Suga H. // Int. J. Mol. Sci. 2015. V. 16. P. 6513–6531. https://doi.org/10.3390/ijms16036513
Deigin V., Ksenofontova O., Yatskin O., Goryacheva A., Ignatova A., Feofanov A., Ivanov V. // Int. Immunopharm. 2020. V. 81. P. 106185. https://doi.org/10.1016/j.intimp.2020.106185
Требования к документам заявки на выдачу патента на изобретение (утверждены приказом Минэкономразвития России от 25 мая 2016 года № 316), https://rospatent.gov.ru/ru/documents/prikaz-ministerstva-ekonomicheskogo-razvitiya-rf-ot-25-maya-2016-g-316
Инструкция к Договору о патентной кооперации (текст, имеющий силу с 1 июля 2020 г.), https://wipolex.wipo.int/ru/text/577922
Стандарт SТ.25 ВОИС. Стандарт по представлению перечней нуклеотидных и аминокислотных последовательностей в заявках на патент, https://rospatent.gov.ru/ru/documents/st-25-ctandartpo-predstavleniyu-perechney-posledovatelnostey-nukleotidov-i-aminokislot-v-patentnyh-zayavkah/download
Стандарт SТ.26 ВОИС. Рекомендуемый cтандарт представления перечней нуклеотидных и аминокислотных последовательностей с использованием языка XML (расширяемого языка разметки), https://rospatent.gov.ru/ru/documents/st-26-rekomenduemyy-standart-predstavleniya-perechney-nukleotidnyh-i-aminokislotnyh-posledovatelnostey-s-ispolzovaniem-yazyka-xml-rasshiryaemogo-yazyka-razmetki/download
Дополнительные материалы отсутствуют.
Инструменты
Биоорганическая химия