

Биология моря, 2019, T. 45, № 5, стр. 334-341
Использование спутниковых данных для оценки удельной скорости роста фитопланктона в поверхностном слое Черного моря
З. З. Финенко 1, *, И. В. Ковалёва 1, В. В. Суслин 2
1 Институт морских биологических исследований РАН
299011 Севастополь, Россия
2 Морской гидрофизический институт РАН
299011 Севастополь, Россия
* E-mail: zosim_finenko@mail.ru
Поступила в редакцию 19.06.2018
После доработки 18.10.2018
Принята к публикации 18.10.2018
Аннотация
Предложена региональная модель для оценки удельной скорости роста фитопланктона в поверхностном слое моря с использованием спутниковых данных. Модель основана на скорости фотосинтеза и биомассе фитопланктона в единицах органического углерода. Для расчета скорости фотосинтеза фитопланктона использованы эколого-физиологические характеристики, полученные ранее для Черного моря. Входными параметрами модели являются концентрация хлорофилла в поверхностном слое; температура воды; интенсивность фотосинтетически активной радиации, падающей на поверхность моря; глубина перемешанного слоя и коэффициент диффузного ослабления света. Сезонные изменения удельной скорости роста фитопланктона рассчитаны для глубоководной части моря и придунайского района. Максимальные значения получены в прибрежном районе в мае–июне, минимальные – в глубоководной части моря перед весенним максимумом развития фитопланктона. Максимальные и минимальные значения различались в 5 раз. Измеренные и расчетные значения удельной скорости роста, полученные для отдельных станций и обширных акваторий моря, оказались достаточно близкими. Модель можно использовать для оперативной оценки удельной скорости роста фитопланктона в поверхностном слое моря по спутниковым данным.
Удельная скорость роста планктонных микроводорослей наряду с первичной продукцией и биомассой фитопланктона относится к важнейшим показателям, отражающим состояние экосистемы. Знания об изменчивости этих параметров необходимы при изучении многих вопросов, связанных с влиянием климата на продуктивность экосистем, с формированием гидрохимического режима вод, биогеохимическими циклами углерода, азота, фосфора, кремния и других элементов минерального питания фитопланктона. Изменение условий среды приводит к изменению функциональных характеристик как отдельных видов водорослей, так и всего фитопланктонного сообщества.
В зависимости от выбранной единицы времени удельная скорость роста может быть часовой, суточной, месячной и годовой. В настоящее время чаще используются суточные значения, которые дают более точную информацию о динамике процессов роста численности и биомассы организмов в лабораторных и естественных условиях. Зная удельную скорость роста фитопланктонного сообщества, легко рассчитать его продукцию, которая будет равна произведению биомассы на удельную скорость роста. В зависимости от того, в каких единицах выражена биомасса фитопланктона (органический углерод, азот), продукция будет принимать ту же размерность, что расширяет возможности для анализа циклов биогенных элементов. Сравнивая величины удельной скорости роста фитопланктона в отдельных районах, можно судить об условиях среды, в которых развиваются водоросли.
В настоящее время в лабораторных условиях получены количественные зависимости, характеризующие действие света, основных биогенных веществ и температуры на скорость роста некоторых видов водорослей (Finenko et al., 2003; Chen, Liu, 2010; Мансурова, 2013). Установлено, что скорость роста водорослей зависит не только от внешних факторов среды, но и от размера клеток и их таксономического положения (Marañón, 2015; Стельмах, Мансурова, 2017). Так, у диатомовых и зеленых водорослей она выше, чем у водорослей других систематических групп. В целом результаты исследований, полученные для некоторых видов водорослей, трудно использовать для оценки скорости роста всего фитопланктонного сообщества по нескольким причинам. В природе скорость роста фитопланктона зависит, во-первых, от совместного действия отдельных факторов среды, во-вторых, от таксономического и размерного состава фитопланктонного сообщества, варьирующего во времени и пространстве. Данных по скорости роста фитопланктона в естественных условиях недостаточно, поэтому моделирование этого показателя с использованием параметров, которые могут быть измерены по спутниковым наблюдениям на больших акваториях и с высокой частотой, крайне важно для понимания процессов, определяющих биосинтез органического вещества автотрофными организмами в море.
Цель настоящего исследования состояла в оценке межгодовой и сезонной динамики биомассы и скорости роста фитопланктона в поверхностном слое глубоководных центральных районов и мелководной северо-западной части Черного моря. В связи с этим решали следующие задачи: 1) разработка модели для оценки первичной продукции в поверхностном слое по спутниковым данным и фотосинтетическим параметрам фитопланктона, измеренным в Черном море; 2) создание алгоритма для оценки удельной скорости роста фитопланктона в поверхностном слое моря.
МАТЕРИАЛ И МЕТОДИКА
Ключевыми параметрами в моделировании удельной скорости роста являются концентрация хлорофилла а, интенсивность солнечной радиации, достигающей поверхности моря, и количество фотосинтетически активной радиации, поглощенной фитопланктоном. Для определения концентрации хлорофилла в поверхностном слое использовали спутниковые наблюдения, полученные с аппаратов SeaWiFS, MODIS-Aqua и MODIS-Terra (http://doi.org/10.5067/ORBVIEW-2/SEAW IFS_OC.2014.0.,10.5067/AQUA/MODIS_OC.2014.0.,10.5067/TERRA/MODIS_OC.2014.0; https://oceancolor.gsfc.nasa.gov/atbd/rrs/; https://oceancolor.gsfc.nasa. gov/atbd/chlor_a/). Измерения спутниковым сканером SeaWiFS, которые проводились с 1998 по 2008 г. и с 2002 по 2015 г., были дополнены данными, полученными с помощью приборов MODIS-Aqua и MODIS-Terra. Расчет хлорофилла базировался на двух индексах: I510 = nLw(555)/nLw(510) и I490 = = nLw(510)/nLw(490), которые усреднялись на пространственной сетке 0.025 градуса по широте и 0.035 градуса по долготе. Алгоритм расчета концентрации хлорофилла и критерии отбора нормализованной спектральной яркости водной поверхности (nLw) на длинах волн 490, 510 и 555 нм подробно описаны в работе Суслина и Чуриловой (Suslin, Churilova, 2016). Индексы I510 и I490 учитывают изменение поглощения окрашенного растворенного вещества, слабо чувствительны к ошибкам атмосферной коррекции и обратному рассеянию света взвесью, а также более точно восстанавливают концентрацию хлорофилла по спутниковым данным по сравнению с алгоритмами, которые использовались ранее. Сравнение результатов, полученных с помощью трех спутниковых сканеров SeaWiFS, MODIS-Aqua и MODIS-Terra, показало, что алгоритм Морского гидрофизического института РАН дает наименьшую относительную ошибку (~30%) (Суслин и др., 2008), поэтому он был применен нами для восстановления концентрации хлорофилла а по спутниковым данным. Для расчета удельной скорости роста использовали входные параметры спутниковых наблюдений: Хл0 – концентрация хлорофилла в поверхностном слое, мг/м3; Т – температура воды, °С; Е0 – интенсивность фотосинтетически активной радиации, падающей на поверхность моря, моль квантов/(м2 сут); zp – глубина перемешанного слоя, м; kd – коэффициент диффузного ослабления света водой, м–1. В нашем случае это ежедневные с 4-километровым пространственным разрешением данные, усредненные за двухнедельный период.
Удельная скорость роста фитопланктона рассчитана для трех районов моря; два из них находились в глубоководной части, а один район располагался в прибрежной области северо-западной части Черного моря (рис. 1).

РЕЗУЛЬТАТЫ
Модель для оценки удельной скорости роста сообщества фитопланктона
Значения удельной скорости роста (µ) рассчитывали как:
Если весь органический углерод, образованный в процессе фотосинтеза, используется на рост водорослей, то ВТ можно определить как: ${{{\text{B}}}_{{\text{T}}}} = {{{\text{B}}}_{0}} + \Delta {\text{P}},$ где ΔP – чистая продукция фитопланктона за сутки, мгС/м3. Для фитопланктона поверхностного слоя моря ΔP равно:
Начальная биомасса фитопланктона (В0) рассчитана как: ${{{\text{B}}}_{0}} = {\text{\;}}{{{\text{Х}}{{{\text{л}}}_{0}}} \mathord{\left/ {\vphantom {{{\text{Х}}{{{\text{л}}}_{0}}} {\left( {{\text{Хл\;}}:{\text{C}}} \right)}}} \right. \kern-0em} {\left( {{\text{Хл\;}}:{\text{C}}} \right)}}$, где Хл : С – удельное содержание хлорофилла а в органическом углероде фитопланктона: ${\text{Хл}}~:{\text{C}} = {\text{r}}{{({{{\text{E}}}_{{{\text{ср}}}}}{{{\text{a}}}_{{{\text{ph}}}}})}^{{ - {\text{g}}}}}$), где Хл : С – безразмерная величина; r и g – коэффициенты, полученные в экспериментах с фитопланктоном, обитающим в поверхностном слое (r = 0.0072, g = 0.395); aph – удельное поглощение света фитопланктоном, нормированное на хлорофилл а (м2/мгХл0), рассчитанное как: ${{{\text{a}}}_{{{\text{ph}}}}} = {\text{dХл}}_{0}^{{ - {\text{f}}}}$, где коэффициент d = 0.015, коэффициент f = 0.29 (Churilova et al., 2004). Среднесуточная облученность в верхнем перемешиваемом слое рассчитана как: ${{{\text{Е}}}_{{{\text{ср}}}}} = \left( {{{0.95{{{\text{Е}}}_{0}}} \mathord{\left/ {\vphantom {{0.95{{{\text{Е}}}_{0}}} {{{{\text{k}}}_{{\text{d}}}}}}} \right. \kern-0em} {{{{\text{k}}}_{{\text{d}}}}}}} \right){{\left( {\exp \left( { - {{{\text{k}}}_{{\text{d}}}}){{{\text{z}}}_{{\text{p}}}}} \right)} \right)} \mathord{\left/ {\vphantom {{\left( {\exp \left( { - {{{\text{k}}}_{{\text{d}}}}){{{\text{z}}}_{{\text{p}}}}} \right)} \right)} {{{{\text{z}}}_{{\text{p}}}}}}} \right. \kern-0em} {{{{\text{z}}}_{{\text{p}}}}}}$, где Eср – средняя облученность в квазиоднородном слое, моль квантов/(м2 сут); zp – глубина перемешанного слоя, м; kd – коэффициент диффузного ослабления света водой, м-1.
Таким образом, используя полученные уравнения и спутниковые данные по концентрации хлорофилла, температуре воды, интенсивности солнечной радиации и коэффициенту диффузного ослабления света водой, были рассчитаны максимальные удельные скорости роста фитопланктона в поверхностном слое моря.
Сезонная динамика удельной скорости роста фитопланктонного сообщества в контрастных районах моря, рассчитанная по модели
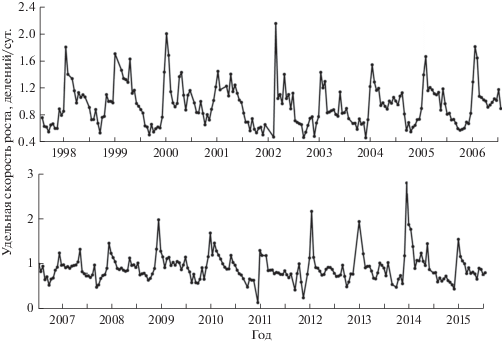
В западном циклональном круговороте глубоководной части Черного моря удельная скорость роста фитопланктона в поверхностном слое (0–1 м) в течение года варьировала в пределах трех раз (рис. 2). В этом районе межгодовая динамика сезонных изменений (1998–2015 гг.) практически не различалась. В течение года наблюдали три максимума удельной скорости роста: первый – в начале зимнего периода (декабрь), второй – весной в период весеннего развития (вторая половина марта, апрель), третий – в конце лета (август). Минимальные значения µ зарегистрированы перед началом весеннего цветения фитопланктона.
Рис. 2.
Сезонная и многолетняя динамика удельной скорости роста фитопланктона в западном (1) и восточном (2) круговоротах Черного моря.

В восточном циклональном круговороте скорость роста и ее сезонная динамика имели тот же характер (рис. 2). В то же время круговороты различались по температурному режиму. В западном циклональном круговороте зарегистрировано восемь холодных зим (температура воды в поверхностном слое ниже 7°С), тогда как в восточном циклональном круговороте – только три. Однако эти различия не отразились на годовой динамике скорости роста. В двух рассматриваемых глубоководных районах весной отмечены минимальные значения µ (в среднем за сезон 0.58/сут). Остальную часть года среднесезонные величины удельной скорости роста находились приблизительно на одном уровне, наибольших значений они достигали осенью в западном циклональном круговороте (0.78/сут) и летом – в восточном (0.77/сут).
В прибрежном районе, находящемся под действием стока р. Дунай, значения удельной скорости роста фитопланктона были заметно выше, чем в глубоководной части моря (рис. 3). В зимний период при колебании температуры от 3.5 до 12°С значения µ варьировали от 0.44 до 1.18/сут. После весеннего паводка скорость роста фитопланктона увеличивалась в несколько раз. Максимум зарегистрирован в июне, когда скорость роста в среднем за 18 лет составила 1.61 ± 0.26/сут, изменяясь от 1.07 до 1.98/сут. Летом она оставалась довольно высокой, в среднем 1.12/сут. Второй максимум скорости роста отмечен осенью. Как правило, он был ниже первого и наблюдался 13 раз за 18 лет. Зимний максимум удельной скорости роста зарегистрирован всего 4 раза за весь исследованный период, в среднем он составил 1.07 ± 0.09/сут. В целом в зимний период в прибрежном районе отмечены наиболее низкие значения µ по сравнению с таковыми в другие сезоны 1998–2015 гг. Среднемноголетние значения интенсивности солнечной радиации и температуры в глубоководной и прибрежной зонах моря были приблизительно одинаковы, тогда как концентрация биогенных веществ различалась на порядок (Гаркавая, Богатова, 2006). Среднегодовые значения скорости роста фитопланктона в течение исследованного периода в прибрежном районе были на 20–25% выше, чем в глубоководной части моря.
ОБСУЖДЕНИЕ
Особенности внутригодовой изменчивости скорости роста фитопланктона обусловлены сменой лимитирующих факторов и таксономического состава фитопланктона. В конце лета максимальные значения µ определяются высокой температурой, при которой повышается удельная скорость фотосинтеза при оптимальной облученности, и относительно высокими концентрациями неорганического азота (Pakhomova et al., 2014). С октября по декабрь температура снижается и ослабевает температурный градиент в столбе воды, а скорость поступления биогенных веществ из более глубоких слоев к поверхности увеличивается. В этот период наблюдаются высокие концентрации неорганического азота (Pakhomova et al., 2014) и развиваются диатомовые водоросли, характеризующиеся малым размером клеток и высокой скоростью роста. В результате биомасса фитопланктона и значения µ повышаются. В феврале–марте при высоком конвективном перемешивании и минимальной температуре воды в поверхностном слое содержание хлорофилла, биомасса фитопланктона и удельная скорость роста снижаются. В это время рост лимитируется низкой температурой, а уменьшение биомассы связано с переносом водорослей в более глубокие слои. В начале весны интенсивность конвективного перемешивания воды ослабевает, что позволяет фитопланктону находиться в поверхностном слое. Это приводит к кратковременному повышению концентрации хлорофилла и удельной скорости роста, которые снижаются в апреле. В мае при повышении температуры и формировании сезонного пикноклина значения µ вновь увеличиваются.
В настоящее время используется два подхода для определения удельной скорости роста фитопланктонного сообщества. Первый подход основан на определении скорости фотосинтеза, измеряемой методом С14, и биомассы фитопланктона в единицах органического углерода (Behrenfeld et al., 2005; Maraňón, 2008), второй – на методе разведения (Landry, 1993). Оба подхода имеют свои допущения. При определении скорости фотосинтеза фитопланктона радиоуглеродным методом не учитываются потери углерода в процессе дыхания и экскреции органического вещества водорослями. Предполагается, что радиоуглеродным методом определяется чистая первичная продукция, однако ее значения находятся в пределах между валовой и чистой продукцией. При втором подходе не учитывается фотоакклимация водорослей. Предполагается, что скорость выедания водорослей прямо пропорциональна разведению пробы чистой морской водой. В целом значения удельной скорости роста фитопланктона при использовании обоих подходов довольно близки (Calbet, 2001; Calbet, Landry, 2004; Landry, Calbet, 2005).
Сравнение значений удельной скорости роста фитопланктона, рассчитанных по разработанной нами модели и полученных методом разведения (Стельмах и др., 2004; Стельмах, 2010), показало идентичный характер динамики в течение года, а также достаточно близкие значения в северо-западной части моря и в прибрежных районах вблизи Севастополя. Среднегодовое значение скорости роста фитопланктона в придунайском районе по модельным расчетам составило 0.93 деления/сут, а в прибрежных водах Крыма, включая бухты, – 0.91 деления/сут. В глубоководных районах показатели несколько различались. Так, среднее значение µ в мае и августе в западной глубоководной части Черного моря, полученное методом разведения на небольшом материале, составило 1.5 деления/сут. По нашим модельным расчетам, основанным на огромном фактическом материале, скорость роста составила 0.7 деления/сут. Эти различия, вероятно, связаны с разным объемом выборок. Таким образом, модельные расчеты, основанные на скорости фотосинтеза и биомассе фитопланктона, дают верные представления об удельной скорости роста и ее пространственно-временной изменчивости.
Бехренфилд с соавторами (Behrenfeld et al., 2005) разработали модель расчета удельной скорости роста фитопланктона в Мировом океане, основанную на спутниковых данных по концентрации хлорофилла, а также на расчетах взвешенного органического углерода и скорости фотосинтеза. В умеренных широтах Атлантического океана максимальная удельная скорость роста фитопланктона достигала около 1 деления/сут и наблюдалась в летний период, зимой она составляла около 0.2 деления/сут. В тропических и субтропических районах удельная скорость роста фитопланктона в течение года изменялась от 0.2 до 0.5 деления/сут, в экваториальных районах – от 1 до 2 делений/сут. Следовательно, в Черном море удельная скорость роста фитопланктона в зимний период значительно выше, чем в океанических водах. Эти различия связаны с особенностями плотностной и гидрохимической структуры вод и определяются глубиной залегания основного пикноклина. Он является нижней границей обитания фитопланктона, от которой зависит увеличение вертикального потока биогенных веществ в зону фотосинтеза в зимний период (Виноградов и др., 1992; Финенко, Крупаткина, 1992; Ведерников, Демидов, 2002). Высокое положение основного пикноклина в Черном море, верхняя граница которого располагается вблизи зоны фотосинтеза, позволяет фитопланктону находиться внутри эвфотического слоя, а поток питательных веществ из более глубоких слоев приводит к быстрому размножению водорослей. В океанических водах умеренных широт основной пикноклин залегает на значительно бóльших глубинах. В результате часть фитопланктона постоянно увлекается турбулентной диффузией ниже зоны фотосинтеза, и роста биомассы зимой в поверхностном слое не наблюдается.
Близкие результаты получены при использовании модели, основанной на скорости фотосинтеза, измеренной по С14, на концентрации хлорофилла по спутниковым данным и на концентрации взвешенного органического углерода, измеренного in situ (Sathyendranath et al., 2009). Биомасса фитопланктона рассчитана по концентрации хлорофилла и отношению С : Хл. Для определения отношения С : Хл использован метод квантильной линейной регрессии между взвешенным органическим углеродом и хлорофиллом. Максимальные значения удельной скорости роста (0.07–0.1/ч) получены в мае у берегов Гренландии и Канады, в открытых районах океана значения были примерно в 2 раза ниже. Минимальные сходные значения в мае получены нами для прибрежных и глубоководных районов Черного моря.
Удельная скорость роста фитопланктона зависит как от абиотических факторов, так и от среднего размера клеток в фитопланктоне (Chen, Liu, 2010; Maraňón, 2015). В океанических водах умеренных широт максимальная скорость роста характерна для нанофитопланктона (Chen, Liu, 2010). В океане и Черном море удельная скорость роста фитопланктона уменьшается от прибрежных районов к открытой части. Различия между прибрежными и глубоководными районами зависят от сезона и района исследований. Можно полагать, что они обусловлены разными процессами, определяющими скорость поступления биогенных веществ в зону фотосинтеза, а также особенностями видового и размерного состава фитопланктона.
Таким образом, предложена региональная модель для оценки удельной скорости роста фитопланктона в поверхностном слое Черного моря с использованием спутниковых данных. Модель включает оценку скорости фотосинтеза и биомассы фитопланктона в единицах органического углерода. Входными параметрами модели являются концентрация хлорофилла в поверхностном слое; температура воды; интенсивность фотосинтетически активной радиации, падающей на поверхность моря, и коэффициент диффузного ослабления света, которые определены по спутниковым данным.
Модельные расчеты удельной скорости роста фитопланктона показали, что в глубоководных районах Черного моря величины варьировали от 0.37 до 1.07 деления/сут в течение 18-летнего периода. Максимальные значения, как правило, наблюдались в августе–сентябре и в декабре, минимальные – перед весенним максимумом развития фитопланктона. В придунайском районе удельная скорость роста изменялась от 0.44 до 1.98 деления/сут, максимальные значения отмечены в мае–июне. Измеренные и расчетные значения удельной скорости роста, полученные для отдельных станций и обширных акваторий моря, были достаточно близкими.
Модель можно использовать для оперативной оценки удельной скорости роста фитопланктона в поверхностном слое моря по спутниковым данным.
Список литературы
Ведерников В.И., Демидов А.Б. Долговременная и сезонная изменчивость хлорофилла и первичной продукции в восточных районах Черного моря // Комплексные исследования северо-восточной части Черного моря. М.: Наука. 2002. С. 212–234.
Виноградов В.И., Сапожников В.В., Шушкина Э.А. Экосистема Черного моря. М.: Наука. 1992. 112 с.
Гаркавая Г.П., Богатова Ю.И. Гидрохимические исследования // Северо-западная часть моря; биология и экология. Киев: Наукова думка. 2006. С. 60–83.
Мансурова И.М. Влияние света на удельную скорость роста динофитовых водорослей Чёрного моря // Мор. экол. журн. 2013. Т. 12. С. 73–78.
Стельмах Л.В. Удельная скорость роста фитопланктона в глубоководной части Черного моря в различные сезоны года // Мор. экол. журн. 2010. Т. 9. № 3. С. 83–87.
Стельмах Л.В., Губанов В.И., Бабич И.И. Сезонные изменения скорости роста и лимитирование фитопланктона питательными веществами в прибрежных водах Черного моря в районе Севастополя // Мор. экол. журн. 2004. Т. 3. № 4. С. 55–73.
Стельмах Л.В., Мансурова И.М. Унимодальная зависимость скорости роста от объема клеток в культурах черноморских видов микроводорослей // Вопр. современной альгологии. 2017. № 1 (13). URL: http://algology.ru/1101.
Суслин В.В., Чурилова Т.Я., Сосик Х.М. Региональный алгоритм расчета концентрации хлорофилла а в Черном море по спутниковым данным SeaWiFS // Мор. экол. журн. 2008. Т. 7. № 2. С. 24–42.
Финенко З.З., Крупаткина Д.К. Первичная продукция в Черном море в зимне-весенний период // Океанология. 1992. Т. 32. № 1. С. 97–104.
Финенко З.З., Суслин В.В., Чурилова Т.Я. Региональная модель для расчета первичной продукции Черного моря с использованием данных спутникового сканера цвета Sea WiFS // Мор. экол. журн. 2009. Т. 8. № 1. С. 81–106.
Behrenfeld M.J., Boss E., Siegel D.A., Shea D.M. Carbon-based ocean productivity and phytoplankton physiology from space // Global Biogeochem. Cycles. 2005. V. 19. P. 1–14.
Calbet A. Mesozooplankton grazing effect on primary production: a global comparative analysis in marine ecosystems // Limnol. Oceanogr. 2001. V. 46. P. 1824–1830.
Calbet A., Landry M.R. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems // Limnol. Oceanogr. 2004. V. 49. P. 51–57.
Chen B., Liu H. Relationships between phytoplankton growth and cell size in surface oceans: interactive effects of temperature, nutrients and grazing // Limnol. Oceanogr. 2010. V. 55. P. 965–972.
Churilova T., Berseneva G., Georgieva L. Variability in the biooptical characteristics of phytoplankton in the Black Sea // Oceanology. 2004. V. 44. № 2. P. 192–204.
Finenko Z.Z., Churilova T.Ya., Sosik H.M., Basturk O. Variability of photosynthetic parameters of the surface phytoplankton in the Black Sea // Oceanology. 2002. V. 42. № 1. P. 53–67.
Finenko Z.Z., Hoepffner N., Williams R., Piontkovski S.A. Phytoplankton carbon to chlorophyll a ratio: response to light, temperature and nutrient limitation // Мор. экол. журн. 2003. V. 2. № 2. С. 40–64.
Landry M.R. Estimating rates of growth and grazing mortality of phytoplankton by the dilution method / Handbook of methods in aquatic microbial ecology. Eds. Kemp P.F., Sherr B.F., Sherr E.B., Cole J.J. Ann Arbor: Lewis Publishers. 1993. P. 715–722.
Landry M.R., Calbet A. Reality checks on microbial food web interactions in dilution experiments: responses to the comments of Dolan and McKeon // Ocean Sci. 2005. V. 1. P. 39–44.
Maraňón E. Inter-specific scaling of phytoplankton production and cell size in the field // J. Plankton Res. 2008. V. 30. № 2. P. 157–163.
Maraňón E. Cell size as a key determinant of phytoplankton metabolism and community structure // Annu. Rev. Mar. Sci. 2015. V. 7. P. 241–264.
NASA Goddard Space Flight Center, Ocean Ecology Laboratory, Ocean Biology Processing Group. Sea-viewing Wide Field-of-view Sensor (SeaWiFS) Ocean Color Data; 2014 Reprocessing. https://doi.org/10.5067/ORBVIEW-2/SEAWIFS_OC.2014.0
NASA Goddard Space Flight Center, Ocean Ecology Laboratory, Ocean Biology Processing Group. Moderate-resolution Imaging Spectroradiometer (MODIS) Aqua Ocean Color Data; 2014 Reprocessing. NASA OB.DAAC, Greenbelt, MD, USA. https://doi.org/10.5067/AQUA/MODIS_OC.2014.0
NASA Goddard Space Flight Center, Ocean Ecology Laboratory, Ocean Biology Processing Group. Moderate-resolution Imaging Spectroradiometer (MODIS) Terra Ocean Color Data; 2014 Reprocessing. NASA OB.DAAC, Greenbelt, MD, USA. https://doi.org/10.5067/TERRA/MODIS_OC. 2014.0
NASA Goddard Space Flight Center, Ocean Ecology Laboratory, Ocean Biology Processing Group. Remote Sensing Reflectance (Rrs). https://oceancolor.gsfc.nasa.gov/atbd/rrs/
NASA Goddard Space Flight Center, Ocean Ecology Laboratory, Ocean Biology Processing Group. Chlorophyll a (chlor_a) / Algorithm Description. https://oceancolor.gsfc.nasa.gov/atbd/chlor_a /
Pakhomova S., Vinogradova E., Yakushev E. et al. Interannual variability of the Black Sea Proper oxygen and nutrients regime: the role of climatic and anthropogenic forcing // Estuar. Coast. Shelf Sci. 2014. V. 140. P. 134–145.
Sathyendranath S., Stuart V., Nair A. et al. Carbon-to-chlorophyll ratio and growth rate of phytoplankton in the sea // Mar. Ecol. Prog. Ser. 2009. V. 383. P. 73–84.
Suslin V., Churilova T.Ya. A regional algorithm for separating light absorption by chlorophyll-a and coloured detrital matter in the Black Sea, using 480–560 nm bands from ocean colour scanners // Int. J. Remote Sens. 2016. V. 37. № 18. P. 4380–4400.
Дополнительные материалы отсутствуют.