

Биология моря, 2021, T. 47, № 4, стр. 244-252
Новые данные о флоре диатомовых водорослей (Bacillariophyta) в заливе Кара-Богаз-Гол Каспийского моря
С. А. Булатов *
Международный институт моделирования и прогнозирования развития морских и гипергалинных экосистем
141603 Клин, Россия
* E-mail: mimge_rus@mail.ru
Поступила в редакцию 17.06.2020
После доработки 27.01.2021
Принята к публикации 01.04.2021
Аннотация
В результате изучения диатомовых водорослей (Bacillariophyta), собранных в 2000–2001 гг. из планктона гипергалинного зал. Кара-Богаз-Гол, выявлено 13 видовых и внутривидовых таксонов, относящихся к родам Campylodiscus, Cocconeis, Diploneis, Epithemia, Navicula, Nitzschia, Planothidium, Surirella и Tryblionella. Водоросли обитали при солености воды от 44 до 250‰. Для большинства таксонов расширены пределы солеустойчивости: 11 видов и разновидностей диатомовых водорослей впервые отнесены к галофилам. Створки видов Planothidium cf. lanceolatum, Epithemia sorex и Nitzschia dissipata из зал. Кара-Богаз-Гол имели меньше штрихов, чем в диагнозе (9–11, 10–12 и 25–35 штрихов в 10 мкм соответственно). Таксоны P. cf. lanceolatum, Surirella brebissonii var. kuetzingii и Tryblionella compressa впервые приведены для Каспийского моря.
Кара-Богаз-Гол является соленым заливом морского происхождения, который расположен в восточной части Каспийского моря на территории Туркменистана. Длина залива по меридиану 165 км и параллели 154 км в координатах 40°35′−41°50′ с.ш. и 50°40′−52°30′ в.д. (Аковецкий, Богданов, 1988). Залив соединяется с Каспийским морем через прол. Кара-Богаз-Гол. Площадь поверхности зеркала залива составляет около 18 000 км2 с уровнем ниже уровня Каспийского моря на 28 м. В период проведения исследований соленость в разных частях залива изменялась от 40 до 270‰ (Булатов, 2020). Средняя глубина залива составляла 4.7 м с колебаниями от 2.2 до 6.2 м (Булатов, 2002).
Первые сведения о таксономическом составе диатомовых водорослей в зал. Кара-Богаз-Гол появились в 1970-х годах (Караева, 1972), впоследствии по результатам проведенных электронно-микроскопических исследований он был дополнен новыми таксонами (Караева, Гаджиева, 1975; Караева, Бухтиярова, 2010). Исследования диатомовых водорослей зал. Кара-Богаз-Гол были продолжены с конца 1990-х годов (Булатов, 2002) и касались в основном изучения продукционных показателей альгофлоры залива (Булатов, 2004). Недавние исследования диатомовых водорослей в зал. Кара-Богаз-Гол позволили описать новый для науки вид диатомовых водорослей Brachysira carabogazgolensis Bulatov, 2020, обитающий при солености 50‰ (Булатов, 2020).
Цели настоящей работы − подробное исследование видового состава диатомовой флоры зал. Кара-Богаз-Гол с использованием светового и сканирующего электронного микроскопов, а также изучение экологии составляющих ее видов.
МАТЕРИАЛ И МЕТОДИКА
Материалом для исследования послужили 19 проб, собранных автором в феврале–июне и сентябре–октябре 2000 г., а также в мае–июне и августе–сентябре 2001 г. на трех прибрежных станциях западного побережья зал. Кара-Богаз-Гол: станция 1 – зона смешения вод, район с координатами 41°06′43″ с.ш. – 52°54′32″ в.д., глубина от 0.2 до 0.5 м; станция 2 – у косы Карасукут, координаты 41°32′51″ с.ш. – 52°52′13″ в.д., глубина от 0.2 до 0.7 м; станция 3 – южнее прол. Кара-Богаз-Гол, координаты 41°04′41″ с.ш. – 52°55′42″ в.д., глубина от 0.4 до 1.0 м (рис. 1).
Рис. 1.
Карта-схема расположения зал. Кара-Богаз-Гол с указанием солености воды в 2000–2001 гг. (Булатов, Шакирова, 2005) и расположение станций отбора планктонных проб (указаны точками): станция 1 – зона смешения вод, станция 2 – коса Карасукут, станция 3 – акватория южнее прол. Кара-Богаз-Гол.

Качественные пробы для изучения таксономического состава диатомовых водорослей в зал. Кара-Богаз-Гол отбирали в 50–300 м от берега планктонной сетью с диаметром входного отверстия 50 см и размером ячеи 40 мкм. Сборщик проходил 100 шагов и процеживал около 9.8 м3 воды при отборе каждой пробы. Собранный материал сливали в пластиковые емкости объемом 1 л и фиксировали 1–2 мл 40% раствора формалина. Соленость воды в местах отбора проб измеряли рефрактометром ATAGO (Япония) с точностью измерения 2.0‰ при температуре от 10 до 30°C. Пробы концентрировали методом центрифугирования (Clark, 1956) на центрифуге ОПН-3 с частотой вращения 2 тыс. об/мин. От органического содержимого створки освобождали методом сжигания концентрированной серной кислотой (Забелина и др., 1951). Электронно-микроскопические исследования проводили с использованием сканирующего электронного микроскопа (СЭМ) JEOL JSM-25S (Япония) при рабочем расстоянии 10 мм и ускоряющем напряжении 15 кВ на базе Института биологии внутренних вод им. И.Д. Папанина РАН (пос. Борок, Ярославская область). Для СЭМ препараты напыляли золотом с помощью установки Eico IB-3 (Япония). Для съемки использовали фотопленку Kodak T-MAX 1114 100/120 PRO и Свема Foto 100. Для изучения створок с помощью светового микроскопа (СМ) готовили постоянные препараты (Забелина и др., 1951), в которых створки заключали в среду “плевракс” (показатель преломления 1.9). В работе использовали световой микроскоп P.Z.O. (Польша), оснащенный 100-кратным масляно-иммерсионным объективом с числовой апертурой 1.25. Таксономическую принадлежность диатомовых водорослей определяли по: Забелина и др., 1951; Прошкина-Лавренко, 1955, 1963; Караева, 1972; Krammer, Lange-Bertalot, 1986, 1988, 1991; Генкал, 1992; Sims, 1996; Hofmann et al., 2011; Van de Vijver et al., 2013; Куликовский и др., 2016; Bertolli et al., 2020. Список диатомовых водорослей составлен по системе, предложенной Раундом с коллегами (Round et al., 1990), с учетом рекомендаций (см.: Guiry, Guiry, 2020). Вычисляли средние значения и стандартное отклонение (Лакин, 1980).
Исследованный материал, препараты и негативы микрофотографий хранятся в Лаборатории изучения гипергалинных экосистем Международного института моделирования и прогнозирования развития морских и гипергалинных экосистем (г. Клин, Московская область, Россия).
РЕЗУЛЬТАТЫ
В планктоне зал. Кара-Богаз-Гол было определено 13 видовых и внутривидовых таксонов диатомовых водорослей, принадлежащих к классу Bacillariophyceae.
Campylodiscus echeneis Ehrenberg ex Kützing, 1844 (рис. 2а, 2б)
Рис. 2.
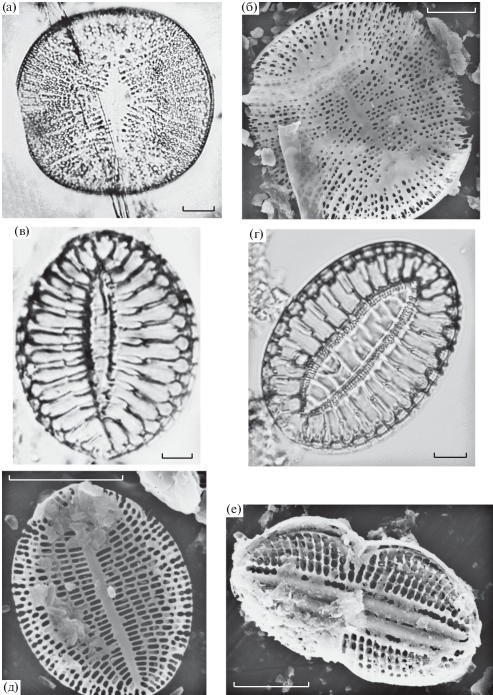
Электронные (СЭМ) и световые (СМ) фотографии микроводорослей из зал. Кара-Богаз-Гол: а, б – Campylodiscus echeneis; в, г – Campylodiscus neofastuosus; д – Cocconeis placentula var. euglypta; е – Diploneis didymus. Масштаб: а – 30 мкм; в, г – 10 мкм (СМ); б – 30 мкм; д, е – 10 мкм (СЭМ).
Караева, 1972: 224; Krammer, Lange-Bertalot, 1988: 213, pl. 175, fig. 1–2; pl. 176, fig. 1–3.
Рис. 3.
Электронные (СЭМ) и световые (СМ) фотографии микроводорослей из зал. Кара-Богаз-Гол: а – Cocconeis lineata; б – Diploneis bombus; в – Epithemia sorex; г, д – Navicula gregaria (г – вид со створки, д – вид структуры створки); е, ж − Nitzschia dissipata (вид со створки); з − Navicula peregrina. Масштаб: а, г, е, ж, з – 10 мкм и д – 1 мкм (СЭМ); б, в – 10 мкм (СМ).

Синонимы: Coronia (Campylodiscus) echeneis Ehrenberg, 1840: 206; Campylodiscus cribrosus W. Smith, 1853: 29, pl. 7, fig. 55 .
Описание. Створки почти круглые, диаметром 144.1–181.5 мкм, в среднем 162.6 ± 14.5 мкм (n = = 48 створок); покрыты расположенными в слаборадиальных рядах округлыми, округло-квадратными и эллиптическими гранулами, 2–3 гранулы в 10 мкм. Рядов гранул 1–3 в 10 мкм. Среднее поле ромбическое или ланцетное. Вид встречался в прибрежном планктоне зоны смешения вод (станция 1) и южнее прол. Кара-Богаз-Гол (станция 3) при солености 73.2–120.0‰.
Campylodiscus neofastuosus Ruck et Nakov, 2016 (рис. 2в, 2г)
Синонимы: Navicula fastuosa Ehrenberg, 1840: 214; Surirella fastuosa (Ehrenberg) Ehrenberg, 1843: 388, pl. 1–4; Прошкина-Лавренко, 1955: 207, рис. 102 , 1 ; табл. VIII , рис. 8 ; 1963: 203, табл. XVI , рис. 1–11 ; Караева, 1972: 224; Sims, 1996: 556, pl. 270, fig. 5 ; Novilla fastuosa (Ehrenberg) Cleve, 1868: 234, pl. 1–4; Suriraya fastuosa (Ehrenberg) Schaarschmidt, 1880: 161.
Описание. Створки широкоэллиптические, почти округло-четырехугольные, длиной 72.5–85.6 мкм, в среднем 78.9 ± 4.7 мкм, и шириной 55.8–64.6 мкм, в среднем 59.9 ± 3.0 мкм (n = 52 створки). Ребра грубые, радиальные, 1.5–2.0 в 10 мкм. Поперечных межреберных штрихов по краю створки 10–11 в 10 мкм. Осевое поле ланцетное, штрихов 10 в 10 мкм. Вид встречался в прибрежном планктоне косы Карасукут (станция 2) при солености 240–250‰.
Cocconeis lineata Ehrenberg, 1849 (рис. 3а)
Караева, 1972: 117, табл. I , рис. 15 ; Krammer, Lange-Bertalot, 1991: 87, pl. 49, fig. 1; pl. 50, fig. 2; pl. 52, fig. 1–13 ; Куликовский и др., 2016: 246, табл. 31 , рис. 1–9 .
Синонимы: Cocconeis placentula var. lineata (Ehrenberg) Van Heurck, 1885: 133, pl. 30, fig. 31 ; Cocconeis placentula f. lineata (Ehrenberg) Hustedt, 1957: 244, pl. 1.
Описание. Створки эллиптические, длиной 17.6–20.7 мкм, в среднем 18.65 ± 1.0 мкм, и шириной 12.5–14.4 мкм, в среднем 13.3 ± 0.6 мкм (n = 23 створки). Штрихи радиальные, ареолированные; на шовной створке 28–30 штрихов в 10 мкм, на бесшовной – 16–18 штрихов в 10 мкм. Вид встречался в прибрежном планктоне косы Карасукут (станция 2) при солености 240–250‰.
Cocconeis placentula var. euglypta (Ehrenberg) Grunow, 1884 (рис. 2д)
Караева, 1972: 117, табл. I , рис. 16 ; Krammer, Lange-Bertalot, 1991: 87, pl. 49, fig. 3; pl. 50, fig. 1; pl. 53, fig. 1–19 ; Куликовский и др., 2016: 245, табл. 32 , рис. 1–5 .
Базионим: Cocconeis euglypta Ehrenberg, 1854: pl. XXXIV, VI, fig. 2.
Синонимы: Cocconeis lineata var. euglypta (Ehrenberg) Grunow, 1880: 121, pl. 30, fig. 33 , 34 ; Cocconeis rouxii var. euglypta (Ehrenberg) J. Héribaud, 1893: 46; Cyclotella lineata var. euglypta Gutwinski, 1895: 155, pl. 2–3; Cocconeis placentula f. euglypta (Ehrenberg) Hustedt, 1957: 244, pl. 1.
Описание. Створки широкоэллиптические, длиной 18.4–37.6 мкм, в среднем 24.6 ± 6.3 мкм, и шириной 12.0–27.2 мкм, в среднем 18.3 ± 4.7 мкм (n = 102 створки). Штрихи радиальные, состоящие из удлиненных ареол; на шовной створке 15–19 штрихов в 10 мкм, на бесшовной – 20–22 штриха в 10 мкм. Микроводоросли отмечены в прибрежном планктоне зоны смешения вод (станция 1) и косы Карасукут (станция 2) при солености 60–210‰.
Diploneis bombus (Ehrenberg) Ehrenberg, 1853 (рис. 3б)
Прошкина-Лавренко, 1963: 138; Караева, 1972: 142, табл. III , рис. 34 .
Базионим: Pinnularia bombus Ehrenberg, 1844: 84.
Синонимы: Navicula bombus (Ehrenberg) Kützing, 1849: 83; Schizonema bombus (Ehrenberg) Kuntze, 1898: 552.
Описание. Створки гитаровидные, сильно суженные посередине, длиной 41.8–73.6 мкм, в среднем 59.4 ± 10.8 мкм; шириной 25.6–35.2 мкм, в среднем 29.6 ± 3.7 мкм; ширина створки на середине 14.4–20.8 мкм, в среднем 17.9 ± 1.7 мкм (n = 82 створки). Концы створок клиновидно-закругленные. Штрихи радиальные, 6–9 в 10 мкм. Вид встречался в прибрежном планктоне зоны смешения вод (станция 1), косы Карасукут (станция 2) и южнее прол. Кара-Богаз-Гол (станция 3) при солености 73.2–210.0‰.
Diploneis didymus (Ehrenberg) Ehrenberg, 1845 (рис. 2е)
Караева, 1972: 140, табл. III , рис. 39 ; Krammer, Lange-Bertalot, 1986: 292, pl. 112, fig. 7 .
Базионим: Pinnularia didymus Ehrenberg, 1844: 85.
Синоним: Schizonema didymus (Ehrenberg) Kuntze, 1898: 552.
Описание. Створки гитаровидные, слегка суженные посередине, длиной 32.0–62.4 мкм, в среднем 50.1 ± 7.3 мкм, и шириной 17.6–32.0 мкм, в среднем 24.9 ± 5.2 мкм; ширина створки на середине 13.0–24.3 мкм, в среднем 19.1 ± 3.8 мкм (n = 58 створок). Штрихи грубые, радиальные, 7–10 штрихов в 10 мкм. Вид отмечен в прибрежном планктоне зоны смешения вод (станция 1) и косы Карасукут (станция 2) при солености 70–242‰.
Epithemia sorex Kützing, 1844 (рис. 3в)
Караева, 1972: 117, табл. I , рис. 16 ; Krammer, Lange-Bertalot, 1988: 154, pl. 106, fig. 1–13 ; Куликовский и др., 2016: 395, табл. 146 , рис. 11 –18 .
Синонимы: Eunotia westermanni var. sorex (Kützing) G. Rabenhorst, 1846: 18; Eunotia sorex (Kützing) G. Rabenhorst, 1853: 18, pl. 1, fig. 7 ; Cystopleura sorex (Kützing) Kuntze, 1891: 891.
Описание. Створки дугообразные, с вогнутым брюшным и выпуклым спинным краями, к концам постепенно суживающиеся, длиной 35.2–46.4 мкм, в среднем 39.3 ± 3.8 мкм, и шириной 11.2–14.4 мкм, в среднем 13.0 ± 1.0 мкм (n = 43 створки). Ребра слабозаметные. Штрихи радиальные, ареолированные, 10–12 штрихов в 10 мкм. Вид встречался в прибрежном планктоне зоны смешения вод (станция 1) при солености 73.2‰.
Navicula gregaria Donkin, 1861 (рис. 3г, 3д)
Караева, 1972: 148, табл. V , рис. 61 ; Krammer, Lange-Bertalot, 1986: 116, pl. 38, fig. 10 –15 ; Куликовский и др., 2016: 330, табл. 55 , рис. 49 –53 .
Описание. Створки ланцетные, с сильно оттянутыми субголовчатыми концами, длиной 26.2–27.9 мкм, в среднем 26.8 ± 0.6 мкм, и шириной 6.3–7.3 мкм, в среднем 6.9 ± 0.4 мкм (n = 28 створок). Среднее поле слабовыраженное. Штрихи тонкие, линеолированные, слабо радиальные, к концам конвергентные, 16–17 штрихов в 10 мкм. Линеол в штрихах, хорошо заметных только в СЭМ, 28–32 в 10 мкм. Вид встречался в прибрежном планктоне зоны смешения вод (станция 1) при солености 44‰.
Navicula peregrina (Ehrenberg) Kützing, 1844 (рис. 3з)
Забелина и др., 1951: 316, рис. 178 , 1 ; Караева, 1972: 156, табл. IV , рис. 47 .
Базионим: Pinnularia peregrina Ehrenberg, 1843: 421, pl. 1, fig. 5 –6 ; pl. 2, fig. 22 ; pl. 3, fig. 3.
Синоним: Schizonema peregrinum (Ehrenberg) Kuntze, 1898: 554.
Описание. Створки ланцетные, длиной 50.0–105.6 мкм, в среднем 81.3 ± 18.3 мкм, и шириной 10.0–20.0 мкм, в среднем 15.6 ± 3.6 мкм (n = 128 створок). Концы широкозакругленные, тупые. Штрихи грубые, линеолированные, радиальные, у концов створки конвергентные, на середине створки более редкие, 7–8 в 10 мкм. Осевое поле ланцетное, среднее поле эллиптически-прямоугольное, расширенное в стороны. Вид отмечен в прибрежном планктоне зоны смешения вод (станция 1) и косы Карасукут (станция 2) при солености 68–250‰.
Nitzschia dissipata (Kützing) Rabenhorst, 1860 (рис. 3е, 3ж; 4а, 4б)
Hofmann et al., 2011: 441, pl. 109, fig. 8 –18 ; Куликовский и др., 2016: 401, табл. 142 , рис. 8 –15 .
Базионим: Synedra dissipata Kützing, 1844: 64, pl. 14, fig. 3.
Синонимы: Nitzschia palea f. dissipata (Kützing) Rabenhorst, 1864: 160; Homoeocladia dissipata (Kützing) Kuntze, 1898: 409; Nitzschia palea var. dissipata (Kützing) Schönfeldt, 1907: 225.
Описание. Створки линейно-ланцетные, с оттянутыми широкозакругленными субголовчатыми концами, длиной 17.7–58.3 мкм, в среднем 33.4 ± 11.4 мкм, и шириной 4.8–7.2 мкм, в среднем 6.3 ± 0.9 мкм (n = 72 створки). Фибул 9–13 в 10 мкм. Штрихи на створке слабозаметные, параллельные, 25–35 штрихов в 10 мкм, состоящие из мелких округлых ареол, хорошо различимых в СЭМ. Вид отмечен в прибрежном планктоне зоны смешения вод (станция 1), косы Карасукут (станция 2) и района южнее прол. Кара-Богаз-Гол (станция 3) при солености 50–240‰.
Planothidium cf. lanceolatum (Brébisson ex Kützing) Lange-Bertalot, 1999 (рис. 4в, 4г, 4д, 4е)
Рис. 4.
Электронные (СЭМ) микрофотографии: а, б – Nitzschia dissipata (а – вид структуры центральной части створки, б – вид структуры конца створки); в, г, д, е – Planothidium cf. lanceolatum (г – шовная створка; в, д, е – бесшовная створка); ж – Surirella brebissonii var. kuetzingii; з – Tryblionella compressa. Масштаб: а, б – 1 мкм; в, г, д, е, ж, з – 10 мкм.

Van de Vijver et al., 2013: 108, fig. 2–60 .
Базионим: Achnanthidium lanceolatum Brébisson ex Kützing, 1846: 247.
Синонимы: Achnanthes lanceolata (Brébisson ex Kützing) Grunow, 1880: 27, pl. XXVII, fig. 8 ; Забелина и др., 1951: 221, рис. 124 , 1 ; Krammer, Lange-Bertalot, 1991: 75, pl. 41, fig. 1–8 , 25 ; Генкал, 1992: 22, табл. 61 , рис. 1–3; Microneis lanceolata (Brébisson ex Kützing) Frenguelli, 1923: 72, pl. 6, fig. 18 –19 ; Achnantheiopsis lanceolata (Brébisson ex Kützing) Lange-Bertalot, 1997: 201, 207, fig. 11 –12 .
Описание. Створки ланцетные, длиной 16.8–31.8 мкм, в среднем 22.9 ± 5.9 мкм, и шириной 7.3–9.5 мкм, в среднем 8.5 ± 0.7 мкм (n = 75 створок), концы широкозакругленные. Створка со швом имеет линейное осевое поле; среднее поле прямоугольно расширено в стороны, не доходит до краев створки. Бесшовная створка с линейно-ланцетным осевым полем, которое иногда в центральной части ромбически расширено. В середине бесшовной створки имеется подковообразная структура. Штрихи радиальные, грубые, 9–11 в 10 мкм. В заливе вид отмечен в прибрежном планктоне зоны смешения вод (станция 1) и косы Карасукут (станция 2) при солености 50–240‰.
Surirella brebissonii var. kuetzingii Krammer ex Lange-Bertalot, 1987 (рис. 4ж)
Krammer, Lange-Bertalot, 1988: 180, pl. 127, fig. 1–8 .
Описание. Створки яйцевидные, верхний конец широкозакругленный, нижний – к концу суженный, остро закругленный, длиной 24.3 мкм и шириной 12.0 мкм (n = 1 створка). Ребра нежные, сильно радиальные, 6–8 в 10 мкм. Микроводоросли отмечали в прибрежном планктоне зоны смешения вод (станция 1) при солености 73.2‰.
Tryblionella compressa (Bailey) Poulin, 1990 (рис. 4з)
Bertolli et al., 2020: 6, fig. 2K−M, fig. 6E−G.
Базионим: Pyxidicula compressa Bailey, 1851: 40, pl. 2, fig. 13 –14 .
Синонимы: Dinopyxis compressa (Bailey) Stein 1883: 1, pl. 1, fig. 34 –38 ; Exuviaella compressa (Bailey) Ostenfeld, 1899: 59; Nitzschia compressa (Bailey) C.S. Boyer, 1916: 116, pl. 39, fig. 7 ; Krammer, Lange-Bertalot, 1988: 46, pl. 37, fig. 1–10 ; Prorocentrum compressum (Bailey) T.H. Abé ex J.D. Dodge, 1975: 110, fig. 2F, 4H; pl. 4E, 4F.
Описание. Створки широкоэллиптически-ланцетные со слегка оттянутыми остро закругленными концами, 57.6 мкм длиной и 30.4 мкм шириной (n = 1 створка). Фибул 6 в 10 мкм. Штрихов 7 в 10 мкм. Вид отмечен в прибрежном планктоне косы Карасукут (станция 2) при солености 240‰.
ОБСУЖДЕНИЕ
В результате проведенных исследований получены оригинальные сведения об обитании в условиях гипергалинных вод зал. Кара-Богаз-Гол 13 диатомовых водорослей, относящихся к родам Campylodiscus Ehrenberg ex Kützing, 1844; Cocconeis Ehrenberg, 1836; Diploneis Ehrenberg ex Cleve, 1894; Epithemia Kützing, 1844; Navicula Bory, 1822; Nitzschia Hassall, 1845; Planothidium Round ex L. Bukhtiyarova, 1996; Surirella Turpin, 1828 и Tryblionella W. Smith, 1853 (см.: Guiry, Guiry, 2020).
Все диатомовые водоросли являются бентосными, обрастающими прибрежные макрофиты и различные твердые субстраты (Караева, 1972; Krammer, Lange-Bertalot, 1988). Преобладание непланктонных (бентосных) форм в водах залива, по-видимому, связано с его мелководностью. Перемешивание водных масс при постоянном ветровом волнении (Аковецкий, Богданов, 1988) приводит к выносу большинства бентосных форм в толщу воды. В результате этого формируется состав планктоценозов водоема, наиболее широко представленных в его мелководной прибрежной зоне.
Преобладание непланктонных форм диатомовых в водах зал. Кара-Богаз-Гол отмечалось нами ранее (Булатов, 2004). Так, из 65 видовых и внутривидовых таксонов диатомовых доминирующее положение занимали бентосные формы (72%) и лишь незначительная часть приходилась на планктонные формы (28%). Из ранее выявленных в заливе другими исследователями 25 таксонов бентосных диатомовых водорослей (Караева, 1972; Караева, Гаджиева, 1975; Караева, Бухтиярова, 2010) в нашем исследовании отмечено только 5 таксонов (C. neofastuosus, C. lineata, Cocconeis scutellum, Navicula salinarum, Nitzschia scalpelliformis). На изменение таксономического разнообразия диатомовых в заливе, возможно, повлияло его полное перекрытие дамбой в 1980 г. (Булатов, 2020), изменившее соотношение сульфатов и хлоридов в воде (Лепешков и др., 1981).
Из встреченных в зал. Кара-Богаз-Гол диатомовых водорослей C. echeneis, C. neofastuosus, C. lineata, C. placentula var. euglypta, D. bombus, D. didymus, E. sorex, N. gregaria, N. peregrina и N. dissipata попали в залив из Каспийского моря (Караева, 1972), а P. cf. lanceolatum, S. brebissonii var. kuetzingii и T. compressa ранее не были отмечены в Каспийском море (Bagheri, Fallahi, 2014; Гусейнов и др., 2019). Известно, что C. neofastuosus встречается в Аральском море при солености от 93 до 134‰ (Сапожников, 2010) и в оз. Бакальское в Крыму при солености 60–110‰ (Неврова, Шадрин, 2005). Наши исследования показали, что этот вид способен обитать и при более критической солености 240–250‰.
В отношении видов C. echeneis, C. placentula var. euglypta, C. lineata, D. bombus, D. didymus, E. sorex, N. gregaria, N. peregrina, N. dissipata, S. brebissonii var. kuetzingii и T. compressa, ранее считавшихся обитателями пресных, солоноватых и морских вод с умеренной соленостью (Забелина и др., 1951; Караева, 1972; Krammer, Lange-Bertalot, 1986, 1988, 1991; Куликовский и др., 2016; Bertolli et al., 2020), полученные нами данные позволили расширить пределы их солевыносливости и отнести к галофилам.
Известно, что длина и ширина створок C. placentula var. euglypta из популяции в Каспийском море составляют соответственно 18.0–29.7 и 12.2–16.4 мкм, D. bombus – 31.0–60.8 и 14.3–23.4 мкм, D. didymus – 30.3–50.4 и 11.6–22.0 мкм, E. sorex – 11.0–33.2 и 5.5–7.7 мкм, N. dissipata – 16.5–33.0 и 3.3–5.5 мкм, а C. echeneis – 86.0–148.4 мкм (Караева, 1972). Створки C. placentula var. euglypta, D. bombus, D. didymus, E. sorex, N. dissipata и C. echeneis из зал. Кара-Богаз-Гол отличались большими размерами по сравнению со створками этих водорослей из Каспийского моря. Створки больших размеров у указанных видов диатомовых водорослей отмечены также в Черном море, соленость воды в котором выше, чем в Каспийском море (Прошкина-Лавренко, 1963).
На всех станциях отбора проб в зал. Кара-Богаз-Гол найден P. cf. lanceolatum, створки которого внешне были схожи со створками Planothidium lanceolatum. По литературным данным (Забелина и др., 1951; Krammer, Lange-Bertalot, 1991; Генкал, 1992) P. lanceolatum является пресноводным видом, ранее не встречавшимся в Каспийском море, но обитающим во впадающей в море р. Волга и ее водохранилищах – Иваньковском, Угличском, Горьковском и Чебоксарском (Генкал, 1992). Характерной особенностью створок P. cf. lanceolatum из зал. Кара-Богаз-Гол является стабильно низкое число штрихов. От протолога (Lange-Bertalot, 1999; Van de Vijver et al., 2013), а также волжской популяции P. lanceolatum (см.: Генкал, 1992) створки P. cf. lanceolatum отличаются меньшей частотой штрихов (9–11 против 12–17 в 10 мкм).
Число штрихов (10–12 в 10 мкм) на створках E. sorex из зал. Кара-Богаз-Гол также было меньше, чем указано в известных диагнозах (Забелина и др., 1951; Караева, 1972; Куликовский и др., 2016), в которых для створок E. sorex указано 12–15 штрихов в 10 мкм. Причем штрихи на створках у E. sorex из залива были грубее, поэтому при наблюдении в СМ характерные для данного вида радиальные ребра на створках (Куликовский и др., 2016) почти не были заметны.
Для N. dissipata из залива также отмечено уменьшение числа штрихов на створке по сравнению с таковым в известных диагнозах: 25–35 штрихов в 10 мкм против 39–50 в 10 мкм (Hofmann et al., 2011; Куликовский и др., 2016).
Таким образом, установлено, что соленость в зал. Кара-Богаз-Гол является фактором, определяющим морфологию створок диатомей: чем выше соленость воды, тем крупнее створки и меньше число структурных элементов на них. По-видимому, в создавшихся условиях соленость среды может выступать в качестве одного из факторов видообразования в заливе.
Список литературы
Аковецкий В.И., Богданов И.Я. Кара-Богаз-Гол: вчера, сегодня, завтра. Ашхабад: Ылым. 1988. 195 с.
Булатов С.А. Структура и состояние диатомовых сообществ высокоминерализованных вод залива Кара-Богаз-Гол // Тез. докл. VIII школы диатомологов России и стран СНГ “Морфология, экология и биогеография диатомовых водорослей”. Борок: Ярославский гос. тех. ун-т. 2002. С. 11–12.
Булатов С.А. Особенности экологии артемии ультрагалинного залива Кара-Богаз-Гол и некоторые аспекты ее питания в естественных условиях // Материалы докл. междунар. научно-исслед. конф. “Биоразнообразие Артемии в странах СНГ: современное состояние ресурсов и их использование”. Тюмень. ФГУП “Госрыбцентр”. 2004. С. 94–104.
Булатов С.А. Новый вид Bacillariophyta из залива Кара-Богаз-Гол (Каспийское море, Туркменистан) // Альгология. 2020. Т. 30. № 2. С. 211–218.
Булатов С.А., Шакирова Ф.М. Современное состояние популяции Artemia sp. (Crustacea, Branchiopoda) залива Кара-Богаз-Гол и перспективы ее рационального использования // Selevinia. 2005. С. 123–128.
Генкал С.И. Атлас диатомовых водорослей планктона реки Волги. Санкт-Петербург: Гидрометеоиздат. 1992. 128 с.
Гусейнов М.К., Гасанова А.Ш., Гусейнов К.М., Хлопкова М.В. К изучению видового разнообразия фитопланктона Среднего Каспия // Вестн. Дагестан. науч. центра. 2019. № 75. С. 11–17.
Забелина М.М., Киселев И.А., Прошкина-Лавренко А.И., Шешукова В.С. Определитель пресноводных водорослей СССР: Диатомовые водоросли. Вып. 4. М.: Советская наука. 1951. 618 с.
Караева Н.И. Диатомовые водоросли бентоса Каспийского моря. Баку: Элм. 1972. 258 с.
Караева Н.И., Гаджиева М.А. Новые данные к изучению диатомовых водорослей Каспийского моря // Новости системат. низш. раст. 1975. Т. 12. С. 123–129.
Караева Н.И., Бухтиярова Л.Н. Дополнения к флоре Bacillariophyta Каспийского моря. I // Альгология. 2010. Т. 20. № 4. С. 471–481.
Куликовский М.С., Глущенко А.М., Генкал С.И., Кузнецова И.В. Определитель диатомовых водорослей России. Ярославль: Филигрань. 2016. 804 с.
Лакин Г.Ф. Биометрия. М.: Высшая школа. 1980. 293 с.
Лепешков И.Н., Буйневич Д.В., Буйневич Н.А., Седельников Г.С. Перспективы использования солевых богатств Кара-Богаз-Гола. М.: Наука. 1981. 274 с.
Неврова Е.Л., Шадрин Н.В. Донные диатомовые водоросли соленых озер Крыма // Мор. екол. журн. 2005. Т. 4. № 4. С. 61–71.
Прошкина-Лавренко А.И. Диатомовые водоросли планктона Черного моря. М.; Л.: Изд-во АН СССР. 1955. 222 с.
Прошкина-Лавренко А.И. Диатомовые водоросли бентоса Черного моря. М.; Л.: Изд-во АН СССР. 1963. 243 с.
Сапожников Ф.В. Ценозы донных микроводорослей Большого Арала на этапе ультрагалинизации // Автореф. дис. … канд. биол. наук. М.: Ин-т океанологии им. П.П. Ширшова. 2010. 22 с.
Bagheri S., Fallahi M. Checklist of phytoplankton taxa in the Iranian Waters of the Caspian Sea // Caspian J. Env. Sci. 2014. V. 12. № 1. P. 81–97.
Bertolli L.M., Talgatti D.M., Nascimento T.M.S., Torgan L.C. The genus Tryblionella W. Smith (Bacillariaceae, Bacillariophyta) in southern Brazil salt marshes // Biota Neotropica. 2020. V. 20. № 1. e20190774.
Clark W.J. An evaluation of methods of concentrating and counting the phytoplankton of Bear Lake // A thesis submitted in partial fulfillment of the requirements for the degree of Master of Science. Utah. 1956. 51 p.
Hofmann G., Werum M., Lange-Bertalot H. Diatomeen im Sübwasser-Benthos von Mitteleuropa. Konigstein: A.R.G. Gantner Verlag K.G. 2011. 908 p.
Guiry M.D., Guiry G.M. AlgaeBase. World-wide electronic publication. National University of Ireland. Galway. 2020. https://www.algaebase.org. Cited November 06, 2020.
Krammer K., Lange-Bertalot H. Bacillariophyceae, Teil 1, Naviculaceae // Süsswasserflora von Mitteleuropa. Stuttgart, New York: Gustav Fischer Verlag. V. 2/1. 1986. 876 p.
Krammer K., Lange-Bertalot H. Bacillariophyceae, Teil 2. Bacillariaceae, Epithemiaceae, Surirellaceae // Süsswasserflora von Mitteleuropa. Jena: Gustav Fischer Verlag. V. 2/2. 1988. 596 p.
Krammer K., Lange-Bertalot H. Bacillariophyceae, Teil 4, Achnanthaceae, Kritische Ergaenzungen zu Navicula (Lineolatae) und Gomphonema // Süsswasserflora von Mitteleuropa. Stuttgart, Jena: Gustav Fischer Verlag. V. 2/4. 1991. 434 p.
Lange-Bertalot H. Neue kombinationen von taxa aus Achnanthes Bory (sensu lato) // Iconographia Diatomologica, Annotated Diatom Micrographs. V. 6. Vaduz: A.R.G. Gantner Verlag K.G. 1999. P. 270–283.
Round F.E., Crawford R.M., Mann D.G. The Diatoms: Biology and Morphology of the Genera. Cambridge, New York: University Press. 1990. 747 p.
Sims P.A. An atlas of British diatoms. Bristol: Biopress Ltd. 1996. 600 p.
Van de Vijver B., Wetzel C., Kopalová K. et al. Analysis of the type material of Achnanthidium lanceolatum Brébisson ex Kützing (Bacillariophyta) with the description of two new Planothidium species from the Antarctic Region // Fottea. Olomouc. 2013. V. 13. № 2. P. 105–117. https://doi.org/10.5507/fot.2013.010
Дополнительные материалы отсутствуют.