

Биология моря, 2021, T. 47, № 5, стр. 313-323
Эпифиты и эндофиты бурой водоросли Punctaria plantaginea (Roth) Greville 1830 (Ectocarpales: Chordariaceae) из залива Петра Великого Японского моря
О. С. Белоус 1, *, Т. В. Титлянова 1
1 Национальный научный центр морской биологии им. А.В. Жирмунского ДВО РАН
690041 Владивосток, Россия
* E-mail: ksu_bio@mail.ru
Поступила в редакцию 04.06.2020
После доработки 29.09.2020
Принята к публикации 22.03.2021
Аннотация
Изучены эпифиты и эндофиты бурой водоросли Punctaria plantaginea из зал. Петра Великого Японского моря. Обнаружено 57 видов водорослей-макрофитообрастателей из отделов Rhodophyta, Ochrophyta, Chlorophyta и Cyanobacteria. Пять видов эпифитов ранее не были указаны для флоры зал. Петра Великого, а четыре вида впервые приведены для флоры дальневосточных морей России. Среди обрастателей доминировали красные водоросли (35%), среди руководящих видов – бурые водоросли, которые составляли 50% от всех найденных видов. В приведенном списке преобладают факультативные голоэпифиты. Показана сезонная динамика видового состава эпифитона P. plantaginea в период с февраля по ноябрь. Количество фитообрастателей увеличивалось с октября по июль. Предполагается, что смена видов в сообществах обрастания P. plantaginea обусловлена сезонными изменениями светового и температурного режимов.
В большинстве таксономических работ, посвященных бентосным водорослям зал. Петра Великого, эпифитизму уделяется особое внимание. В первую очередь, это касается изучения видового состава и биомассы эпифитона культивируемых, промысловых (Паймеева, 1975; Суховеева, 1975; Перестенко, Чумаков, 2001; Михайлова, Штрик, 2007; Климова, 2015) или потенциально промысловых водорослей (Жигадлова, 2011), что обусловлено негативным влиянием эпифитов на продукцию хозяйственно-важных видов. Известно, что поселение эпи- и эндофитов часто вызывает морфологические аномалии у водоросли-хозяина, которые могут приводить к разрушению тканей базифита (Correa, McLachlan, 1992, 1994; Correa et al., 1994; Sánchez et al., 1996), снижению структурной прочности слоевищ и отрыву талломов от субстрата во время штормов (Lein et al., 1991). Кроме этого, густые эпифитные синузии препятствуют проникновению света к водоросли-хозяину, снижая его фотосинтетическую активность, скорость роста и конкурентную способность (Friedlander, 1992). Эндо- и эпифиты обнаружены у большинства видов водорослей зал. Петра Великого (Перестенко, 1980; Перестенко, Титлянова, 2001; Титлянов и др., 2016). В сборах водорослей, проведенных нами в бухтах Соболь и Троицы (Белоус и др., 2013, 2021), среди макрофитов, поросших эпифитами, особой плотностью поселения их талломов визуально выделялась бурая водоросль Punctaria plantaginea (Roth) Greville 1830. Ранее на слоевищах данной водоросли обнаружено 4 вида эпифитов, один из них был новым для науки (Перестенко, Титлянова, 2001).
На основании этого объектом для продолжения работы по изучению видового разнообразия современной бентосной флоры зал. Петра Великого, а также таксономии и экологии эпифитизма в Азиатско-Тихоокеанском регионе была выбрана бурая водоросль P. plantaginea – массовый вид, широко представленный в дальневосточных морях. Цель настоящей работы – получение сведений о составе фитообрастателей P. plantaginea в зал. Петра Великого, о сезонной динамике поселения, росте и развитии талломов эпифитона.
МАТЕРИАЛ И МЕТОДИКА
Объект исследования
Род Punctaria Greville 1830 широко распространен в Мировом океане и насчитывает 18 валидных видов (Guiry, Guiry, 2020). В юго-восточной Азии водоросли, относящиеся к этому роду, используются в пищу в сыром и обработанном виде (Дзизюров и др., 2008); выделенные из P. plantaginea биологически активные вещества обладают антикоагулянтной и антитромбической активностью (Ustyuzhanina et al., 2016). В зал. Петра Великого Японского моря встречается три вида данного рода: P. plantaginea (Roth) Greville 1830 (Перестенко, 1980), P. latifolia Greville 1830 (Клочкова, 1998; Коженкова, 2008) и P. flaccida Nagai 1940 (Клочкова, 1998), среди которых массовым является вид P. plantaginea.
В зал. Петра Великого P. plantaginea вегетирует с октября по июль (Дзизюров и др., 2008), растет в нижней литорали и сублиторали на скалистом, каменистом и илисто-песчаном с камнями грунтах, а также на створках моллюсков и на других водорослях. С февраля по июнь этот вид образует обширные практически чистые заросли или поселяется среди других крупных водорослей (Галышева, Христофорова, 2007; Калита, Скрипцова, 2014), определяя весенний аспект растительности (Перестенко, 1980). Как и другие массовые водоросли, P. plantaginea служит субстратом для многих эпифитных и эндофитных водорослей (Перестенко, Титлянова, 2001).
Сбор, идентификация и хранение материала исследования
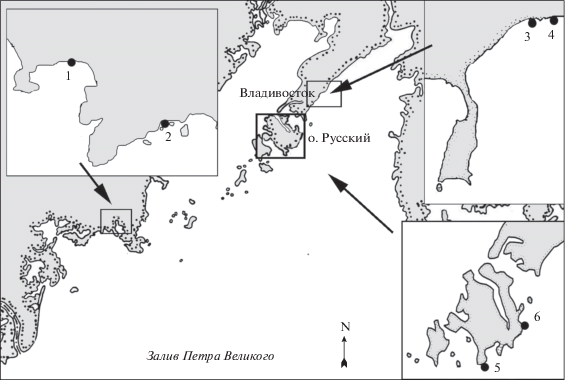
Образцы P. plantaginea собирали на литорали и в сублиторали с использованием легководолазного снаряжения, а также из выбросов вдоль побережья в разных частях зал. Петра Великого (рис. 1). Сборы проводили в июле 2006 г. (б. Песчаная) и 2018 г. (б. Рисовая), в мае 2019 г. (б. Соболь), в июне (б. Сухопутная, мыс Тобизина) и июле (б. Рисовая, мыс Ахлестышева) 2019 г.; в октябре–ноябре 2019 г. и в феврале–апреле 2020 г. (б. Соболь). Всего изучено более 500 слоевищ P. plantaginea.
Рис. 1.
Карта-схема мест сбора Punctaria plantaginea. 1 – б. Рисовая, 2 – б. Песчаная, 3 – б. Соболь, 4 – б. Сухопутная, 5 – мыс Тобизина, 6 – мыс Ахлёстышева.
Водоросли-макрофитообрастатели просматривали в световом микроскопе Carl Zeiss Jena и AxioVert 200M; микрофотографии выполняли с помощью цифровой фотокамеры AxioCam HRc. Эпифиты определяли по морфологическим и анатомическим признакам, а также по строению репродуктивных органов. Эндофиты определяли на срезах ткани водоросли-хозяина по морфологическим признакам и органам размножения. При идентификации водорослей использовали литературные источники (Косинская, 1948; Kuckuck, 1955; Зинова, 1967; Виноградова, 1979; Перестенко, 1980, 1994; Перестенко, Титлянова, 2001; Taşkin et al., 2006; Белоус и др., 2013; Титлянов и др., 2016). Названия таксонов приведены в соответствии с современными систематическими представлениями (Guiry, Guiry, 2020). Обработанный материал хранится в гербарии Лаборатории автотрофных организмов Национального научного центра морской биологии им. А.В. Жирмунского ДВО РАН.
Статистическая обработка
Для анализа сходства видового состава по сезонам использовали пакет статистических программ PRIMER v.6 (Clarke, Gorley, 2006). Степень видового сходства в разные сезоны устанавливали по коэффициенту Серенсена.
Терминология
В настоящей работе мы используем общепринятые термины. Эпифиты – организмы, поселяющиеся на поверхности растения-хозяина. Среди эпифитов принято различать голоэпифиты, когда водоросль поселяется непосредственно на поверхности слоевища другой водоросли, не проникая внутрь; амфиэпифиты – базальная часть эпифита проникает во внутренние ткани базифита, при этом его верхняя часть, выступающая над поверхностью хозяина, может быть представлена лишь репродуктивными структурами, а также эндофиты, которые полностью погружаются в тело базифита (Виноградова, 1989). Кроме этого мы выделяем факультативные эпифиты, растущие не только на водорослях, но и на других субстратах, облигатные эпифиты, живущие лишь на других водорослях, а также адельфоэпифиты, поселяющиеся на близкородственном базифите.
РЕЗУЛЬТАТЫ
Видовое разнообразие фитообрастателей на Punctaria plantaginea
Всего на P. plantaginea обнаружено 57 видов эпи- и эндофитов, из них красные водоросли (Rhodophyta, Rh) составляли 20 видов или 35% от всех видов, бурые (Ochrophyta, Phaeophyceae, Ph) – 15 видов (26%), зеленые (Chlorophyta, Ch) – 18 видов (32%) и синезеленые (Cyanobacteria, Cy) – 4 вида (7%). Перечисленные водоросли принадлежат 4 отделам, 7 классам, 15 порядкам, 23 семействам и 43 родам (табл. 1). Пять видов водорослей приведены для зал. Петра Великого впервые, это Microspongium globosum Reinke 1888, Myrionema foecundum (Strömfelt) Sauvageau 1897, Phaeostroma pustulosum Kuckuck 1893, Punctaria tenuissima (C. Agardh) Greville 1830 и Streblonema evagatum Setchell & N.L. Gardner 1922 (Ph). Виды M. globosum, M. foecundum, P. tenuissima и P. pustulosum ранее не указывались для дальневосточных морей России.
Таблица 1.
Список фитообрастателей, поселяющихся на слоевище Punctaria plantaginea
| Виды | Экологическая группа | Место сбора | Встречаемость по сезонам | |||
|---|---|---|---|---|---|---|
| октябрь–ноябрь | февраль–март | апрель–май | июнь–июль | |||
| RHODOPHYTA Класс Bangiophyceae Порядок Bangiales Семейство Bangiaceae |
||||||
| 1. Porphyra sp. | гэп | 3 | – | + | – | – |
| Класс Compsopogonophyceae Порядок Erythropeltales Семейство Erythrotrichiaceae |
||||||
| 2. Erythrotrichia carnea (Dillwyn) J. Agardh | гэп | 1, 5 | – | – | – | + |
| Класс Stylonematophyceae Порядок Stylonematales Семейство Stylonemataceae |
||||||
| 3. Stylonema alsidii (Zanardini) K.M. Drew | гэп | 1 | – | – | – | + |
| Класс Florideophyceae Порядок Acrochaetiales Семейство Acrochaetiaceae |
||||||
| 4. Acrochaetium humile (Rosenvinge) Børgesen | гэп | 3, 4 | – | + | + | + |
| 5. Acrochaetium moniliforme (Rosenvinge) Børgessen | гэп | 5, 3 | + | – | – | + |
| 6. Acrochaetium secundatum (Lyngbye) Nägeli | гэп | 1 | – | – | – | ед |
| Порядок Colaconematales Семейство Colaconemataceae |
||||||
| 7. Colaconema daviesii (Dillwyn) Stegenga | гэп | 1 | – | – | – | + |
| Порядок Corallinales Семейство Corallinaceae |
||||||
| 8. Pneophyllum fragile Kützing | гэп | 5, 6 | – | – | – | + |
| 9. Peyssonelia sp. | гэп | 6 | – | – | – | ед |
| Порядок Ceramiales Семейство Ceramiaceae |
||||||
| 10. Ceramium kondoi Yendo | гэп | 1 | – | – | – | ед |
| Семейство Dasyaceae | ||||||
| 11. Dasya sessilis Yamada | гэп | 1 | – | – | – | ед |
| 12. Dasysiphonia japonica (Yendo) H.-S. Kim [=Heterosiphonia japonica Yendo] | гэп | 6 | – | – | – | ед |
| Семейство Rhodochortonaceae | ||||||
| 13. Rhodochorton purpureum (Lightfoot) Rosenvinge | гэп | 1 | – | – | – | ед |
| Семейство Rhodomelaceae | ||||||
| 14. Chondriadasyphylla (Woodward) C. Agardh | гэп | 1 | – | – | – | + |
| 15. Laurencia nipponica Yamada | гэп | 6 | – | – | – | + |
| 16. Melanothamnus yendoi (T. Segi) Díaz-Tapia & Maggs [= Polysiphonia yendoi T. Segi] | гэп | 1 | – | – | – | ед |
| 17. Melanothamnus japonicus (Harvey) Díaz-Tapia & Maggs [= Neosiphonia japonica] | гэп | 1, 2, 3, 5, 6 | + | + | + | ++ |
| 18. Polysiphonia morrowii Harvey | гэп | 1 | – | – | – | ед |
| 19. Symphyocladia latiuscula (Harvey) Yamada | гэп | 4 | – | – | – | ед |
| 20. Symphyocladiamarchantioides (Harvey) Falkenberg | гэп | 1 | – | – | – | ед |
| OCHROPHYTA Класс Phaeophyceae Порядок Ectocarpales Семейство Chordariaceae |
||||||
| 21. Botrytella micromora Bory | гэп | 3, 5 | + | + | + | ед |
| 22. Microspongium globosum Reinke**♣ | гэп | 3 | – | ++ | ++ | – |
| 23. Myrionema foecundum (Strömfelt) Sauvageau**♥ | гэп | 5, 6 | – | – | – | + |
| 24. Myrionema sp. | гэп | 6 | – | – | – | + |
| 25. Phaeostroma pustulosum Kuckuck **♥ | гэп | 6 | – | – | – | + |
| 26. Protectocarpus speciosus (Børgesen) Kornmann | гэп | 3 | – | ++ | ++ | – |
| 27. Punctaria tenuissima (C. Agardh) Greville**♣ | гэп | 3 | – | + | + | – |
| 28. Sphaerotrichia divaricata (C. Agardh) Kylin | гэп | 3 | – | + | + | – |
| 29. Streblonema evagatum Setchell & N.L. Gardner*♥ | амф | 4 | – | – | – | + |
| 30. Streblonema sp. | амф | 4 | – | – | – | + |
| 31. Strepsithalia punctariae Perestenko & T.V. Titlyanova | амф | 1, 2, 5, 6 | – | – | – | ++ |
| Семейство Acinetosporaceae | ||||||
| 32. Feldmannia irregularis (Kützing) Hamel | гэп | 3 | – | + | – | – |
| 33. Pylaiella littoralis (Linnaeus) Kjellman | гэп | 5 | – | – | – | + |
| Семейство Ectocarpaceae | ||||||
| 34. Ectocarpus siliculosus (Dillwyn) Lyngbye | гэп | 4 | – | + | + | – |
| Порядок Sphacelariales Семейство Sphacelariaceae |
||||||
| 35. Sphacelariarigidula Kützing | гэп | 1,2,35, 6 | – | – | + | ++ |
| CHLOROPHYTA Класс Ulvophyceae Порядок Ulotrichales Семейство Ulotrichaceae |
||||||
| 36. Acrosiphonia heterocladia (Sakai) K.L. Vinogradova | гэп | 3 | – | – | + | – |
| 37. Protomonostroma undulatum (Wittrock) K.L. Vinogradova | гэп | 3 | – | – | ед | – |
| 38. Ulothrix flacca (Dillwyn) Thuret | гэп | 3 | – | + | + | – |
| 39. Ulothrix implexa (Kützing) Kützing | гэп | 3 | – | + | + | – |
| 40. Urospora penicilliformis (Roth) Areschoug | гэп | 3 | – | – | + | – |
| 41. Urospora wormskioldii (Mertens) Rosenvinge | гэп | 3 | – | ++ | + | – |
| Семейство Monostromataceae | ||||||
| 42. Monostroma grevillei (Thuret) Wittrock | гэп | 3 | – | ед | – | – |
| Порядок Cladophorales Семейство Cladophoraceae |
||||||
| 43. Cladophorastimpsonii Harvey | гэп | 5, 6 | – | – | – | + |
| 44. Cladophoraspeciosa Sakai | гэп | 3, 5 | – | + | + | + |
| Порядок Ulvales Семейство Bolbocoleonaceae |
||||||
| 45. Bolbocoleon piliferum Pringsheim | энд | 1, 6 | – | – | – | + |
| Семейство Kornmanniaceae | ||||||
| 46. Kornmannia leptoderma (Kjellman) Bliding | гэп | 3 | – | – | ед | – |
| Семейство Ulvaceae | ||||||
| 47. Ulva lactuca Linnaeus | гэп | 1, 3, 6 | – | – | + | + |
| 48. Ulva linza Linnaeus | гэп | 3 | – | ед | – | – |
| 49. Ulva sp. | гэп | 3 | – | – | – | + |
| Семейство Ulvellaceae | ||||||
| 50. Ulvella viridis (Reinke) R. Nielsen, C.J. O’Kelly & B. Wysor | энд | 1, 5, 6 | – | – | – | + |
| 51. Ulvella repens (Pringsheim) R. Nielsen, C.J. O’Kelly & B. Wysor | энд | 6 | – | – | – | + |
| 52. Ulvella sp. | энд | 1, 6 | – | – | – | + |
| Порядок Ulvophyceae incertae sedis Семейство Ulvophyceae familia incertae sedis (неопределенное систем. положение) |
||||||
| 53. Blastophysa rhizopus Reinke | энд | 1, 6 | – | – | – | ++ |
| CYANOBACTERIA Класс Cyanophyceae Порядок Nostocales Семейство Rivulariaceae |
||||||
| 54. Calothrix aeruginea Thuret ex Bornet & Flahault | гэп | 2, 6 | – | – | – | + |
| 55. Isactis plana Thuret ex Bornet & Flahault | гэп | 1, 2 | – | – | – | ++ |
| Порядок Oscillatoriales Семейство Oscillatoriaceae |
||||||
| 56. Oscillatoria limosa C. Agardh ex Gomont | гэп | 6 | – | – | – | ед |
| 57. Oscillatoria sp. | гэп | 1 | – | – | – | + |
Примечание. *Виды, впервые найденные в зал. Петра Великого; **виды, впервые зарегистрированные во флоре дальневосточных морей России; ♥ – подробное описание и фотографии этих видов опубликованы в работе Белоус и др., 2021; ♣ – подробное описание и фотографии этих видов приведены в настоящей работе. Условные обозначения: гэп – голоэпифит, амф – амфиэпифит, энд – эндофит; 1 – б. Рисовая, 2 – б. Песчаная, 3 – б. Соболь, 4 – б. Сухопутная, 5 – м. Тобизина, 6 – м. Ахлёстышева; “+” – вид встречается редко, “++” – часто, ед – единично, “–“ – вид не обнаружен.
Практически все найденные нами бурые водоросли, поселившиеся на слоевищах P. plantaginea, за исключением Sphacelaria rigidula Kützing 1843 (Ph), являлись адельфоэпифитами и принадлежали порядку Ectocarpales; 11 из них, как и водоросль-хозяин, относились к семейству Chordariacea (Ph).
По способу взаимодействия с тканями хозяина большинство найденных водорослей (86%) – голоэпифиты; виды S. evagatum, Streblonema sp. и Strepsithalia punctariae Perestenko & T.V. Titlyanova 2001 (Ph) (5%) – амфиэпифиты; к эндофитам отнесены 5 видов (9%): Bolbocoleon piliferum Pringsheim 1862, Ulvella viridis (Reinke) R. Nielsen, C.J. O’Kelly & B. Wysor 2013, Ulvella repens (Pringsheim) R. Nielsen, C.J. O’Kelly & B. Wysor 2013, Ulvella sp. и Blastophysa rhizopus Reinke 1889 (Ch). Среди поселенцев P. plantaginea преобладали факультативные голоэпифиты, из них наиболее часто встречались Acrochaetium spp., Melanothamnus japonicus (Harvey) Díaz-Tapia & Maggs 2017 (Rh), S. rigidula, Protectocarpus speciosus (Børgesen) Kornmann 1955 (Ph), Urospora wormskioldii (Mertens) Rosenvinge 1893 (Ch) и Isactis plana Thuret ex Bornet & Flahault 1886 (Cy). Девять видов – M. globosum, M. foecundum, Myrionema sp., P. pustulosum, S. evagatum, Streblonema sp., S. punctariae (Ph), Erythrotrichia carnea (Dillwyn) J. Agardh 1883 и Stylonema alsidii (Zanardini) K.M. Drew 1956 (Rh) – являлись облигатными эпифитами.
В группу руководящих видов входили водоросли из всех четырех отделов (табл. 2), лидирующее положение занимали бурые водоросли M. globosum, P. speciosus, S. punctariae и S. rigidula – 50% от всех руководящих видов. Затем следовали зеленые водоросли B. rhizopus и U. wormskioldii – 25%. Из красных водорослей наиболее часто встречался вид M. japonicus (12.5%), а из цианобактерий – I. plana (12.5%). В группе характерных видов доминировали отделы Chlorophyta (35%) и Ochrophyta (32%), затем следовали отделы Rhodophyta (27%) и Cyanobacteria (6%) с видами Calothrix aeruginea Thuret ex Bornet & Flahault 1886 и Oscillatoria sp. В группу редких видов входили водоросли из отделов Rhodophyta (67%), Chlorophyta (27%) и Cyanobacteria (6%).
Таблица 2.
Количественный состав эпи- и эндофитов, обнаруженных на Punctaria plantaginea
| Отдел | Общее количество видов | Руководящие виды | Характерные виды | Редкие виды | ||||
|---|---|---|---|---|---|---|---|---|
| число | % | число | % | число | % | число | % | |
| Rhodophyta | 20 | 35 | 1 | 12.5 | 9 | 27 | 10 | 67 |
| Ochrophyta | 15 | 26 | 4 | 50 | 11 | 32 | − | − |
| Chlorophyta | 18 | 32 | 2 | 25 | 12 | 35 | 4 | 27 |
| Cyanobacteria | 4 | 7 | 1 | 12.5 | 2 | 6 | 1 | 6 |
Сезонная динамика эпифитона Punctaria plantaginea
Отмечены значительные сезонные изменения в составе обрастателей P. plantaginea (рис. 2). В октябре–ноябре (осенний сезон) на ее молодых слоевищах найдено всего 3 вида водорослей: Acrochaetium moniliforme (Rosenvinge) Børgessen 1915, M. japonicus (Rh) и Botrytella micromora Bory 1822 (Ph). К зимнему и зимне-весеннему сезонам количество обрастателей значительно увеличилось. Общее число водорослей, которые росли на P. plantaginea в феврале–марте, составило 16 видов, из них Rhodophyta – 3 вида, Ochrophyta (Phaeophyceae) – 7 видов и Chlorophyta – 6 видов. Среди всех обрастателей доминировали 2 вида бурых водорослей (M. globosum и P. speciosus). Эти эпифиты образовывали многочисленные пятна на слоевище хозяина, занимая до 70% площади поверхности слоевища (рис. 3). Среди зеленых водорослей преобладала U. wormskioldii, в обрастаниях часто встречались Ulothrix flacca (Dillwyn) Thuret 1863 и U. implexa (Kützing) Kützing 1849. В апреле–мае видовой состав водорослей изменился незначительно. Обнаружено 18 видов эпифитов: 2 вида Rhodophyta, 7 видов Ochrophyta (Phaeophyceae) и 9 – Chlorophyta. Коэффициент Серенсена показал максимальное сходство видового состава эпифитов между этими сезонами на уровне 55%. К летнему сезону (июнь–июль) число обрастателей увеличилось до 41 вида преимущественно за счет красных водорослей (19 видов) с доминированием M. japonicus; 10 видов обнаружены как единственная находка. Водоросли Laurencia nipponica Yamada 1931 и Chondria dasyphylla (Woodward) C. Agardh 1817 найдены в ювенильной стадии. Отмечено по 9 видов бурых и зеленых водорослей. Среди бурых водорослей в значительном количестве найдены S. punctariae и S. rigidula, а среди зеленых преобладала эндофитная водоросль B. rhizopus, которая максимально занимала межклеточные пространства и полости, образовавшиеся после спороношения водоросли-хозяина. В летние месяцы также встречались 4 вида Cyanobacteria: C. aeruginea, I. plana, Oscillatoria limosa C. Agardh ex Gomont 1892 и Oscillatoria sp., среди которых доминировал вид I. plana. К осеннему сезону видовое разнообразие резко снизилось. Виды M. japonicus (Rh) и B. micromora (Ph) были отмечены в составе эпифитона P. plantaginea во все сезоны.
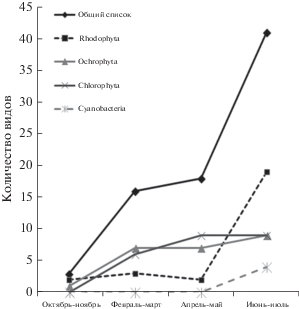
Рис. 3.
Эпифиты Protectocarpus speciosus и Microspongium globosum, образующие бурые пятна на поверхности слоевища Punctaria plantaginea.

Ниже приведены описания двух видов водорослей, ранее не зарегистрированных во флоре дальневосточных морей. Описание и фотографии трех остальных видов (M. foecundum, P. pustulosum, S. evagatum) опубликованы ранее (Белоус и др., 2021).
Punctaria tenuissima (C. Agardh) Greville 1830 (рис. 4а–г)
Рис. 4.
Растущие на Punctaria plantaginea водоросли-эпифиты Punctaria tenuissima (а–г) и Microspongium globosum (д–з): а – внешний вид P. plantaginea, один из талломов которой полностью оброс P. tenuissima (показан стрелками); б – молодое растение P. tenuissima, часть его слоевища с настоящими волосками, отходящими от края пластины (в), и поперечный срез слоевища (г); д – пучки таллома M. globosum с однорядными многогнездными спорангиями; е – разветвленные вертикальные нити M. globosum с латеральными и терминальными однорядными многогнездными спорангиями; ж – вертикальные нити, отходящие от однослойного основания M. globosum; з – часть слоевища M. globosum с пустыми спорангиями (показаны стрелками). Условные обозначения: h – настоящие волоски, s – спорангии, р – пиреноид.
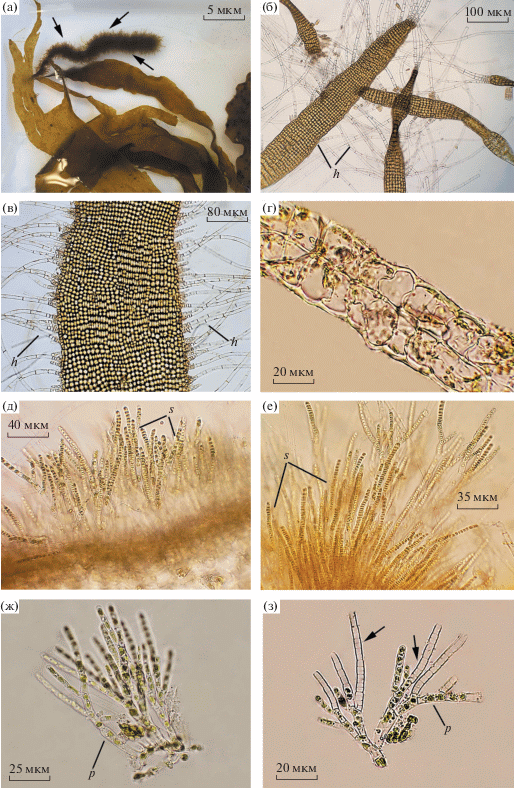
Растение имеет две стадии развития: нитевидную и пластинчатую. Нитевидная стадия представлена базальными стелющимися нитями, от которых поднимаются однорядные или многорядные нити с 1–2 волосками на вершине. Слоевище пластинчатое, узколентовидное, длиной 1–5 см и шириной 0.2–1.2 см, с вытянутой заостренной вершиной, состоит из 2–4 слоев клеток. Настоящие волоски густо развиваются по краям пластины.
Примечание. В дальневосточных морях России вид ранее не был отмечен. Встречается в Арктике (Баренцево море) (Малавенда, 2018), в Балтийском (Nielsen et al., 1995) и Черном (Зинова, 1967) морях, у берегов Северной Америки (Mathieson et al., 2001) и юго-восточной Азии (Мьянма) (Silva et al., 1996).
Microspongium globosum Reinke 1888 (рис. 4д–з)
Слоевище эпифитное, образует на водорослях темно-бурые пятна диаметром 0.5–3.0 мм. Состоит из базального диска и вертикальных разветвленных однорядных нитей высотой 240–500 (до 800) мкм. Клетки нитей длиной 50–60 мкм и шириной 4–6 мкм с одним или двумя пластинчатыми хлоропластами. Настоящие волоски присутствуют. Терминально и латерально отходящие от вертикальных нитей многогнездные спорангии однорядные, цилиндрические, их размер 50–90 × 5–6 мкм. Одногнездные спорангии не обнаружены.
Примечание. В дальневосточных морях России вид ранее не отмечен. Встречается в Арктике (Lee, 1980), в северной части Атлантического океана (Pedersen, 1976), у берегов Европы (Karlson et al., 2018), Северной Америки (Lindstrom, 1977) и Японии (Yoshida, 1998). Вид отмечен в Средиземном (Taşkin et al., 2006), Баренцевом и Белом морях (Михайлова, Штрик, 2007).
ОБСУЖДЕНИЕ
За время исследования на/в слоевище Punctaria plantaginea обнаружено 57 видов эпи- и эндофитов. Это 18% от общего числа видов водорослей, встречающихся в зал. Петра Великого (Скрипцова, 2019). Фитообрастатели (в основном тонконитевидные и микроскопические виды) появлялись на молодых пластинах P. plantaginea с октября и достигали обилия к концу вегетации водоросли (в июле–августе). Хрящеватые макроэпифиты L. nipponica и C. dasyphylla встречались в ювенильной стадии и не достигали зрелого состояния, так как слоевище P. plantaginea вегетирует несколько месяцев и является недолговечным субстратом. Несмотря на это, на некоторых ее талломах найдено более 20 видов водорослей-обрастателей (100 экз. и более).
Почти все обнаруженные на слоевище P. plantaginea бурые водоросли – адельфоэпифиты из порядка Ectocarpales семейства Chordariaceae. Это неудивительно, так как порядок Ectocarpales является самым многочисленным в классе Phaeophyceae и насчитывает 769 видов, из которых 501 вид входит в семейство Chordariaceae (Guiry, Guiry, 2020). Большинство представителей этого семейства – тонконитевидные эпифитные и эндофитные водоросли, которые широко представлены в нашем регионе (Перестенко, 1980) и поэтому являются основными обрастателями.
Результаты наблюдений показали, что сообщество обрастателей начинает формироваться уже на молодых стерильных талломах P. plantaginea. В зал. Петра Великого молодые пластины этой водоросли-хозяина длиной 4–8 см найдены в октябре с эпифитами A. moniliforme, M. japonicus (Rh) и B. micromora (Ph). Наши наблюдения не противоречат данным, полученным ранее. Известно, что инфицирование талломов ламинарий бурыми эндофитными водорослями происходит на стадии проростков, так как заражению взрослых талломов препятствует плотный слой коровых клеток, через который нити эндофитов не могут проникнуть в более глубокие слои (Peters, Schaffelke, 1996; Bernard et al., 2018). Обильное обрастание талломов P. plantaginea отмечено нами и в зимне-весенний сезон. В феврале–апреле на ее стерильных и фертильных слоевищах поселялись нитчатые эпифитные водоросли M. globosum и P. speciosus (Ph) в виде многочисленных бурых пятен, занимавших большую часть поверхности слоевища (рис. 3).
Согласно нашим данным, количество обрастателей и плотность их поселения на пластинах P. plantaginea увеличивались в течение всего периода вегетации водоросли, достигая максимума в июне–июле.
Смена видового состава эпифитона может быть связана с изменением температурных и световых условий, определяющих циклы развития как самих обрастателей, так и водоросли-хозяина. Ранее было показано, что в зал. Петра Великого в осенние месяцы многие виды водорослей заканчивают стадию вегетации, а в стадии репродукции находится минимальное количество видов (Гусарова, Бадыкина, 2002; Гусарова, 2003). В ранневесенний период при температуре ниже 4°С массово вегетируют эфемерные нитчатые зеленые водоросли, такие как Ulothrix spp. и Urospora spp. (Перестенко, 1980; Очеретяна, 2017), что, вероятно, и определяет структуру сообщества обрастания P. plantaginea в это время. К началу июня с повышением температуры воды заканчивается вегетация ранневесенних эфемеров, таких как Urospora spp., Ulothrix spp., Acrosiphonia heterocladia (Sakai) K.L. Vinogradova 1979, Monostroma grevillei (Thuret) Wittrock 1866, Protomonostroma undulatum (Wittrock) K.L. Vinogradova 1969, Kornmannia leptoderma (Kjellman) Bliding 1969 (Ch) и Porphyra sp. (Rh). Видовое богатство эпифитов на P. plantaginea достигает максимума в июне–июле, когда появляются относительно тепловодные виды эпифитов, как эфемеров, так и долгоживущих (однолетних) водорослей. В это время вегетирует большинство красных (E. carnea, S. alsidii, Acrochaetium spp., M. japonicus) и эндофитных зеленых (Ulvella spp., B. rhizopus, B. piliferum) водорослей. Они активно поселяются на/в P. plantaginea, замещая холодноводные бурые водоросли M. globosum, P. speciosus и Ectocarpus siliculosus (Dillwyn) Lyngbye 1819, которые составляли основу сообщества обрастания в зимне-весенний и весенний сезоны. Из бурых водорослей летом обильны только два вида – S. punctariae и S. rigidula. При максимальном прогреве мелководья в конце июля – начале августа у P. plantaginea заканчивается вегетация и ее слоевище разрушается (Перестенко, Титлянова, 2001).
Список литературы
Белоус О.С., Титлянова Т.В., Титлянов Э.А. Морские растения бухты Троицы и смежных акваторий (залив Петра Великого, Японское море). Владивосток: Дальнаука. 2013. 264 с.
Белоус О.С., Скрипцова А.В., Титлянова Т.В. Новые и малоизвестные виды макроводорослей дальневосточных морей России // Биол. моря. 2021. Т. 47. № 3. С. 202–208.
Виноградова К.Л. Определитель водорослей дальневосточных морей СССР. Зеленые водоросли. Л.: Наука. 1979. 146 с.
Виноградова К.Л. Эпифитизм водорослей: уточнение терминологии // Бот. журн. 1989. Т. 74. № 9. С. 1291–1293.
Галышева Ю.А., Христофорова Н.К. Среда и макробентос залива Восток Японского моря в условиях рекреационного воздействия // Изв. ТИНРО. 2007. Т. 159. С. 270–309.
Гусарова И.С., Бадыкина И.А. Сроки вегетации и размножения некоторых видов водорослей Приморья // Изв. ТИНРО. 2002. Т. 131. С. 340–346.
Гусарова И.С. Сезонная динамика растительности в бухте Прогулочной (зал. Петра Великого) // Изв. ТИНРО. 2003. Т. 133. С. 126–137.
Дзизюров В.Д., Кулепанов В.Н., Шапошникова Т.В. и др. Атлас массовых видов водорослей и морских трав российского Дальнего Востока. Владивосток: ТИНРО-Центр. 2008. 327 с.
Жигадлова Г.Г. Эпифиты и эндофиты водорослей рода Palmaria Stackhouse у берегов восточной Камчатки // Изв. ТИНРО. 2011. Т. 164. С. 300–311.
Зинова А.Д. Определитель зеленых, бурых и красных водорослей южных морей СССР. М.; Л.: Наука. 1967. 398 с.
Калита Т.Л., Скрипцова А.В. Сублиторальные сообщества макрофитов Уссурийского и Амурского заливов (Японское море) в современных условиях // Биол. моря. 2014. Т. 40. № 6. С. 427–434.
Климова А.В. Эпи- и эндофиты ламинариевых водорослей Авачинского залива (юго-восточная Камчатка) // Природные ресурсы, их современное состояние, охрана, промысловое и техническое использование. Материалы VI Всерос. научно-практ. конф. 2015. С. 81–86.
Клочкова Н.Г. Водоросли-макрофиты дальневосточных морей России: Автореф. дис. … д-ра биол. наук. Петропавловск-Камчатский. 1998. 48 с.
Косинская Е.К. Определитель морских сине-зеленых водорослей. М.; Л.: Изд-во АН СССР. 1948. 279 с.
Коженкова С.И. Ретроспективный анализ морской флоры залива Восток Японского моря // Биол. моря. 2008. Т. 34. № 3. С. 159–174.
Малавенда С.В. Флора водорослей макрофитов Кольского залива (Баренцево море) // Вестн. МГТУ. 2018. Т. 21. № 2. С. 245–252.
Михайлова Т.А., Штрик В.А. Макроэпифиты Laminaria hyperborea (Laminariaceae) Баренцева и Белого морей // Бот. журн. 2007. Т. 92. № 12. С. 1818–1828.
Очеретяна С.О. Видовой состав и структура альгосообществ “зеленых приливов” в Авачинской губе и устойчивость зеленых водорослей-макрофитов к неблагоприятному воздействию: Автореф. дис…. канд. биол. наук. Петропавловск-Камчатский. 2017. 23 с.
Паймеева Л.Г. Обрастания зостеры Японского моря // Изв. ТИНРО. 1975. Т. 98. С. 193–201.
Перестенко Л.П. Водоросли залива Петра Великого. Л.: Наука. 1980. 232 с.
Перестенко Л.П. Красные водоросли дальневосточных морей России. СПб.: Ольга. 1994. 330 с.
Перестенко Л.П., Чумаков А.А. Эпифиты Laminaria japonica Aresch. f. longipes (Miyabe et Tokida) Ju. Petrov с острова Сахалин // Новости системат. низш. раст. 2001. Т. 35. С. 26–30.
Перестенко Л.П., Титлянова Т.В. Дополнение к флоре Японского моря // Новости системат. низш. раст. 2001. Т. 35. С. 18–26.
Скрипцова А.В. Водоросли-макрофиты залива Петра Великого Японского моря // Биота и среда заповедных территорий. 2019. № 3. С. 14–57.
Суховеева М.В. Эпифиты ламинариевых дальневосточных морей // Изв. ТИНРО. 1975. Т. 98. С. 184–192.
Титлянов Э.А., Титлянова Т.В., Белоус О.С. Полезные морские растения стран Азиатско-Тихоокеанского региона. Владивосток: Дальнаука. 2016. 360 с.
Bernard M., Rousvoal S., Jacquemin B. et al. qPCR-based relative quantification of the brown algal endophyte Laminarionema elsbetiae in Saccharina latissima: variation and dynamics of host–endophyte interactions // J. Appl. Phycol. 2018. V. 30. P. 2901–2911.
Clarke K.R., Gorley R.N. PRIMER version 6: User manual/tutorial. Plymouth, U.K.: PRIMER-E. 2006. 192 p.
Correa J.A., Flores V., Garrido J. Green patch disease in Iridaea laminarioides (Rhodophyta), caused by Endophyton sp. (Chlorophyta) // Dis. Aquat. Org. 1994. V. 19. P. 203–213.
Correa J.A., McLachlan J.L. Endophytic algae of Chondrus crispus (Rhodophyta). IV. Detrimental effects on the host following infections by Acrochaete operculata and A. heteroclada (Chlorophyta) // Mar. Ecol.: Prog. Ser. 1992. V. 81. P. 73–87.
Correa J.A., McLachlan J.L. Endophytic algae of Chondrus crispus (Rhodophyta). V. Fine structure of the infection by Acrochaete operculata (Chlorophyta) // Eur. J. Phycol. 1994. V. 29. P. 33–47.
Friedlander M. Gracilaria conferta and its epiphytes: the effect of culture conditions on growth // Bot. Mar. 1992. V. 35. P. 423–428.
Guiry M.D., Guiry G.M. AlgaeBase. World-wide electronic publication. National University of Ireland, Galway. 2020. http://www.algaebase.org. Accessed June, 2020.
Karlson B., Andreasson A., Johansen M. et al. Nordic Microalgae. World-wide electronic publication. http://nordicmicroalgae.org. Accessed June, 2020.
Kuckuck P. Ectocarpaceen-Studien III Protectocarpus nov. gen. // Helgol. Wiss. Meeresunters. 1955. V. 5. P. 119–140.
Lee R.K.S. A catalogue of the marine algae of the Canadian Arctic // Canada National Museum of Natural Sciences Publications in Botany. V. 9. Ottawa: Natl. Mus. Can. 1980. P. 1–82.
Lein T.E., Sjøtun K., Wakili S. Mass-occurrence of a brown filamentous endophyte in the lamina of the kelp Laminaria hyperborea (Gunnerus) foslie along the southwestern coast of Norway // Sarsia. 1991. V. 76. P. 187–193.
Lindstrom S.C. An annotated bibliography of the benthic marine algae of Alaska // Alaska Department of Fish and Game Technical Data Report. V. 31. Juneau, Alaska: Alaska Dep. Fish Game. 1977. P. 1–172.
Mathieson A.C., Dawes C.J., Anderson M.L., Hehre E.J. Seaweeds of the Brave Boat Harbor salt marsh and adjacent open coast of southern Maine // Rhodora. 2001. V. 103. № 913. P. 1–46.
Nielsen R., Kristiansen A., Mathiesen L., Mathiesen H. Distributional index of the benthic marine macroalgae of the Baltic Sea area // Acta Bot. Fenn. 1995. V. 155. P. 1–70.
Pedersen P.M. Marine, benthic algae from southernmost Greenland // Medd. Groenl. 1976. V. 199. № 3. P. 1–80.
Peters A.F., Schaffelke B. Streblonema (Ectocarpales, Phaeophyceae) infection in the kelp Laminaria saccharina (Laminariales, Phaeophyceae) in the western Baltic // Hydrobiologia. 1996. V. 326/327. P. 111–116.
Sánchez P., Correa J.A., Garcia-Reina G. Host-specificity of Endophyton ramosum (Chlorophyta), the causative agent of green patch disease in Mazzaella laminarioides (Rhodophyta) // Eur. J. Phycol. 1996. V. 31. P. 173–179.
Silva P.C., Basson P.W., Moe R.L. Catalogue of the benthic marine algae of the Indian Ocean // Univ. Calif. Publ. Bot. V. 79. Berkley, Calif.: Univ. of California Press. 1996. P. 1–1259.
Taşkin E., Öztürk M., Wynne M.J. First report of Microspongium globosum Reinke (Phaeophyceae, Myrionemataceae) in the Mediterranean Sea // Nova Hedwigia. 2006. V. 82. P. 135–142.
Ustyuzhanina N.E., Bilan M.I., Gerbst A.G. et al. Anticoagulant and antithrombotic activities of modified xylofucan sulfate from the brown alga Punctaria plantaginea // Carbohydr. Polym. 2016. V. 136. P. 826–833.
Yoshida T. Marine algae of Japan. Tokyo: Uchida Rokakuho Publ. 1998. 1222 p. [in Japanese].
Дополнительные материалы отсутствуют.