

Биология моря, 2022, T. 48, № 2, стр. 75-89
Строение и морфофункциональный анализ вторичной верхней челюсти в связи с особенностями добычи корма у костистых рыб (Teleostei)
Е. С. Громова 1, *, В. В. Махотин
1 Московский государственный университет (МГУ)
119991 Москва, Россия
* E-mail: zhenya_s@inbox.ru
Поступила в редакцию 10.04.2021
После доработки 23.07.2021
Принята к публикации 17.09.2021
- EDN: QYRZZA
- DOI: 10.31857/S0134347522020048
Аннотация
В работе рассмотрено функционирование вторичной верхней челюсти в связи с особенностями ее морфологии у разных видов Teleostei на дефинитивной стадии развития. Выделены основные варианты кинетизма maxillare и praemaxillare в сагиттальной и поперечной плоскостях. Показано, что внешне сходное устройство аппарата питания Teleostei в основе может иметь разное строение вторичной верхней челюсти.
Аппарат питания Teleostei представляет собой высокомобильную скелетно-мускульную систему, конструкцию которой формируют несколько составляющих: вторичные передние челюсти (Гуртовой и др., 1976; Дзержинский, 2005; Дзержинский и др., 2013), суспензориум, гиоид и аппарат глоточных челюстей (pharyngeal jaw apparatus − PJA). Кости жаберной крышки, нейрокраниум и плечевой пояс вносят опосредованный вклад в процесс начального схватывания корма животным (Liem, 1978; Lauder, 1982; Westneat, 2006). Данные, характеризующие работу структур черепа, используются для изучения взаимоотношений между анатомией висцерального механизма и экологией вида (Westneat, 1995). Путем модификации конструкции составляющих его элементов аппарат питания Teleostei приобретает новые функциональные возможности, благодаря чему рыбы могут расширить спектр потребляемого корма. Механизм вторичных передних челюстей включает maxillare и praemaxillare сверху, а dentale снизу (Sanford, Lauder, 1990). У примитивных таксонов группы в составе верхней челюсти могут присутствовать supramaxillare (Weitzman, 1974; Fink, Weitzman, 1982; Fink, 1985; Sasaki et al., 2006), а у некоторых родов сельдевидных Clupeoidei, таких как Harengula, Pliosteostoma и Pellona, также имеются озубленные hypomaxillare (Berry, 1964). Перемещения dentale обусловлены мобильностью anguloarticulare − элемента первичной нижней челюсти, однако у большинства видов Teleostei эти две кости (вместе с retroarticulare и меккелевым хрящом) функционируют как единый элемент (Дзержинский, 2005), который можно обозначить нижней челюстью и условно внести в состав вторичных передних челюстей (Гуртовой и др., 1976; Дзержинский и др., 2013). Изучение строения и особенностей работы данного механизма, в частности закономерностей его кинетизма (подвижности), является одним из важных и распространенных направлений морфофункциональных исследований в ихтиологии (Schaeffer, Rosen, 1961; Alexander, 1967а; Gosline, 1973, 1996; Liem, 1978; Lauder, 1982; Anker, 1987 и др.). Опубликованы данные о том, что кинетизм ротового аппарата способствует повышению манипуляционных возможностей челюстей и оптимизации распределения сил, действующих в черепе при удержании добычи (Иорданский, 2011). Наличие подвижности спланхнокраниума способствует “подгонке” челюстей рыбы к проглатываемому пищевому объекту, а также амортизации толчков в случае схватывания подвижной жертвы. Кинетизм может быть пассивным или обеспечивается системой собственного мускульного аппарата (Иорданский, 1980).
Исследованию механизмов действия вторичной верхней челюсти Teleostei посвящено значительное количество публикаций (Kirckhoff, 1958; Osse, 1969; Motta, 1988; Waltzek, Wainwright, 2003; Westneat, 2006), однако лишь в некоторых из них сделаны попытки анализа широкого набора данных, сочетающих анатомию ротового аппарата и движение его скелетных элементов в процессе ловли добычи (Schaeffer, Rosen, 1961; Gosline, 1987). Необходимо отметить, что особенности механизмов, лежащих в основе смещений maxillare и praemaxillare, обсуждаются до сих пор. Во многих публикациях значительная вариация анатомии, присущая двум вышеуказанным костям у представителей Teleostei, позволяет авторам проанализировать лишь часть морфологических признаков вторичной верхней челюсти без учета других (Staab et al., 2012). Классическими работами в данном направлении считаются исследования Александера (Alexander, 1967a, 1967b), который описывает конструкцию и выдвижение praemaxillare у некоторых форм группы Teleostei. В обзоре Мотта (Motta, 1984) сделана попытка классификации механизмов, обусловливающих протракцию praemaxillare, и описаны примеры их сочетания в рамках одного аппарата питания в зависимости от систематического положения представителей Teleostei. Однако в настоящее время вследствие быстро меняющейся таксономии данной группы (Nelson, 2006) при систематизации вариантов работы вторичной верхней челюсти Teleostei большое значение приобретает учет экологии питания рыб и способов начального захвата корма. Так, было показано, что одинаковые “жизненные формы” рыб (Шарова, 1973), обладающие конвергентно сходным устройством вторичных передних челюстей, могут формироваться у неродственных таксонов (Криволуцкий, 1971; Решетников, 1993). Необходимо отметить, что в русскоязычной литературе исследования по указанной тематике (функционирование вторичной верхней челюсти в связи с ее устройством) немногочисленны (Воскобойникова, 1986; Сиделева, Механикова, 1990; Gromova, Makhotin, 2018). Так, Еремеева (1948) описывает развитие в онтогенезе и устройство челюстного аппарата у взрослых особей некоторых видов карпообразных Cypriniformes с заключениями о его функционировании. В нашем обзоре сделана попытка обобщения имеющейся информации об анатомии аппарата maxillare и praemaxillare у Teleostei и о способах его работы.
Цель настоящего обзора − разработка классификации вариантов подвижности, которые свойственны вторичной верхней челюсти половозрелых особей Teleostei на качественном уровне с указанием основных особенностей строения ее элементов. В рамках этой цели поставлены следующие задачи: 1) подробно описать признаки костно-мышечной системы вторичной верхней челюсти у широкого ряда представителей Teleostei; 2) дать функциональную интерпретацию выявленным чертам анатомии челюстного аппарата; 3) выполнить сравнительный анализ собранных данных для характеристики особенностей добычи корма у спектра таксонов Teleostei. Признаки морфологии maxillare и praemaxillare использованы для понимания механизмов их функционирования при разных способах питания (всасывании, укусе и таране) у большого количества видов группы. Приведены примеры доминирования и редукции роли вторичной верхней челюсти в процессе начального схватывания корма.
В тексте использованы следующие сокращения: m. – мускул (musculus), lig. – связка (ligamentum), pr. – отросток (processus), car. – cartilago (хрящ).
Изменение вторичной верхней челюсти в ходе эволюции Pisces
Впервые озубленные praemaxillare и maxillare дифференцировались у палеонисков Palaeonisciformes, однако они обладали иными по сравнению с Teleostei функциональными свойствами (Cloutier, Arratia, 2004). Praemaxillare неподвижно срасталось с покровными костями крыши нейрокраниума, а его сочленение с maxillare было плохо выражено. Последнее вместе с praeoperculum, quadratojugale, dermohyale и окологлазничными костями являлось составляющим костного блока “щеки” рыбы, задний край которого во время открывания рта демонстрировал некоторое смещение вниз и наружу. Изнутри maxillare крепилось к небноквадратному хрящу (car. palato-quadratum) (Gardiner, 1960). Высокоамплитудные латеральные движения суспензориума новоперых рыб Neopterygii способствовали разъединению элементов наружного костного блока их щеки и, в частности, отделению maxillare и praeoperculum (Gardiner, 1960). Гардинер (Gardiner, 1967) полагал, что этому процессу способствовало функционирование m. levator arcus palatini (LAP), возросшая мускульная масса которого обеспечивала вышеописанные изменения, а также укрупнение m. adductor mandibulae (AM). При этом задний конец maxillare приобретал свободу смещения антеровентрально относительно первичной верхней челюсти, сочленяясь лишь с ее передним отделом (Gardiner, 1960). Таким образом, отверстие рта новоперых рыб сместилось вперед, что привело к увеличению объема их ротовой полости. Эти перестройки анатомии обеспечили возникновение нового способа питания, отличного от прямого укуса − всасывания, которое значительно расширяло спектр потребляемых типов добычи, включая ранее недоступных мелких жертв (Schaeffer, Rosen, 1961; Дзержинский, 2005).
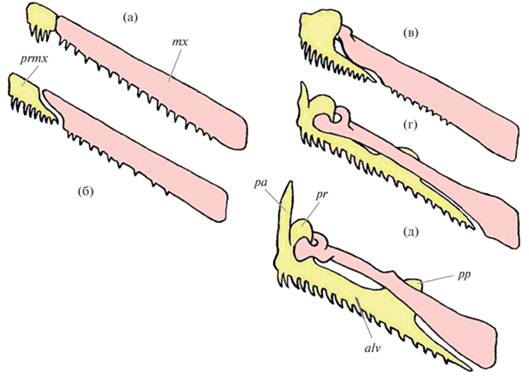
Путь эволюционных изменений вторичной верхней челюсти у Teleostei, состоящий из нескольких этапов (рис. 1), в ходе которых степень ее подвижности возрастала, описал Розен (Rosen, 1982). По мнению автора у мезозойских таксонов этой группы озубленные maxillare и praemaxillare следовали друг за другом, были прочно соединены между собой и могли вместе поворачиваться (“качаться”) ростровентрально вокруг сустава praemaxillare с этмоидным отделом нейрокраниума. Размер praemaxillare по сравнению с длинной рукояткой maxillare, которое образовывало край ротового отверстия сверху, был маленьким. В процессе дальнейшего развития аппарата вторичной верхней челюсти амплитуда смещения maxillare вперед и вниз увеличивалась; эта кость образовывала самостоятельное подвижное прикрепление относительно этмоида и суспензориума (Konstantinidis, Harris, 2011). Длина альвеолярной ветви praemaxillare возрастала, накладываясь на рукоятку maxillare, а головка praemaxillare усложнялась, формируя сочленовный и восходящий отростки (Delsman, 1925). В наиболее продвинутом состоянии у Teleostei maxillare и praemaxillare выстроены параллельно друг другу (Regan, 1911). Maxillare утрачивает зубы (Branch, 1966), его верхний конец приобретает комплексную структуру с несколькими отростками, в том числе для управления движением головки praemaxillare (Ishida, 1994). Последнее образует край пасти рыбы, поэтому первым входит в контакт с добычей, осуществляя ее укус и функционально фактически заменяя maxillare (Delsman, 1925; Schaeffer, Rosen, 1961; Johnson et al., 1996) (рис. 1).
Рис. 1.
Эволюционное развитие вторичной верхней челюсти (а−д) у Teleostei: cвязь между maxillare (mx) и praemaxillare (prmx) (по: Rosen, 1982). Остальные условные обозначения: alv – ramus alveolaris praemaxillare, pa – pr. ascendens praemaxillare, pp – pr. postmaxillaris, pr – pr. articularis praemaxillare.
Морфофункциональные особенности вторичной верхней челюсти Teleostei
Основным скелетным элементом, выполняющим крепление вторичной верхней челюсти на черепе рыбы, является maxillare, второстепенная роль принадлежит praemaxillare. У большинства Teleostei maxillare имеет сочленение или поддерживает контакт с mesethmoideum/supraethmoideum/preethmoideum, этмоидным хрящом или vomer (Weitzman, 1974; Hartel, Stiassny, 1986; Ishida, 1994; Johnson et al., 1996), хотя иногда это бывает и ectoethmoideum, как у полурыла Zenarchopterus kampeni (см.: Vandewalle et al., 2002) или жабуна Opsanus и сапу Batrachoides (см.: Gosline, 1996). У многих Teleostei, особенно продвинутых (Osse, 1969), maxillare имеет сочленение с palatinum и/или соединено с ним связками (Weitzman, 1974; Johnson et al., 1996; Hilton, 2007; Grande, Poyato-Ariza, 2010). Нередко взаимодействие этих двух костей обусловливает подвижность maxillare (Ishida, 1994; Vandewalle et al., 2002). Часто palatinum при формировании сустава с maxillare образует цилиндрический вырост – пальцевидный отросток (Ishida, 1994). Однако у некоторых представителей контакт maxillare c palatinum отсутствует, как, например, у фрактолема Анзорга Phractolaemus ansorgii (см.: Grande, Poyato-Ariza, 2010) и некоторых аностомовых рыб Anostomidae (см.: Sidlauskas, Vari, 2008), а взамен его имеется сочленение palatinum c praemaxillare, как у бомбиля Harpadon (см.: Baldwin, Johnson, 1996). У ряда форм отсутствует контакт обеих костей верхней вторичной челюсти с суспензориумом, например, у хоботнорыловых Mastacembelidae (см.: Gosline, 1983) и чаудхуриевых Chaudhuriidae (см.: Britz, Kottelat, 2003).
Разнообразие вариантов конструкции вторичной верхней челюсти у ряда таксонов Teleostei огромно (Weitzman, 1974; Bertelsen, Struhsaker, 1977; Travers, 1984; Fink, 1985; Cancino, Burgos, 2009). Путем видоизменения строения, формирующего его костные, хрящевые и соединительно-тканные элементы, аппарат питания Teleostei приобретает новые функциональные возможности, благодаря чему рыбы могут расширить спектр потребляемых типов добычи (Michel et al., 2015). Строение и ориентация maxillare, praemaxillare и нижней челюсти преобразуются в зависимости от местонахождения корма, его размера и способа захвата, создавая разные направления ротового отверстия (верхний, конечный и нижний рот) (Sidlauskas, Vari, 2008). Внешне сходное устройство аппарата питания, обусловленное одинаковыми пищевыми предпочтениями, в основе может иметь разную конструкцию вторичной верхней челюсти. Так, длинное рыло обыкновенной длиннорылой рыбы-бабочки Forcipiger longirostris сформировано чрезвычайно протяженными praemaxillare и dentale (Ferry-Graham et al., 2001a), тогда как удлиненный ростральный отдел черепа макрорамфосодеса Macrorhamphosodes platycheilus характеризуется крошечным аппаратом вторичных передних челюстей, смещенным на значительное расстояние от этмоида нейрокраниума (Tyler, 1980).
Упрощение конструкции вторичной верхней челюсти может происходить вследствие 1) потери функциональной нагрузки у одного/обоих составляющих ее элементов (Chapman, 1942; Branch, 1966; Johnson et al., 1996; Konstantinidis, Johnson, 2016) и/или 2) необходимости обеспечения силового укуса добычи (Roberts, 1969; Eagderi, 2010). Ярким примером утраты роли maxillare в схватывании добычи и по сути изъятия его из состава вторичных передних челюстей может служить висцеральный аппарат большинства сомообразных Siluriformes (см.: Vigliotta, 2008). Редукция костей вторичной верхней челюсти в процессе онтогенеза наблюдается у самцов цератиеподобных удильщиков Ceratioidea. Взамен исчезнувших составляющих у них формируются дополнительные озубленные элементы для прикрепления к телу самки (Berry, 1964). У ряда видов Teleostei наблюдается тенденция к формированию объединенной “монолитной” вторичной верхней челюсти путем (1) скрепления maxillare с praemaxillare при помощи неподвижных швов (Tyler, 1980) или (2) полного слияния в единый элемент (Berry, 1964; Travers, 1984; Britz, Kottelat, 2003). У индийской пилайи Pillaia indica и нагаихтиса Nagaichthys filipes (см.: Britz, Kottelat, 2003) происходит функциональная и анатомическая замена вторичной верхней челюстью составляющих первичной верхней челюсти (Travers, 1984). Их крупное, робустное maxillopraemaxillare по величине значительно превышает тонкий редуцированный передний отдел суспензориума, представленный ectopterygoideum, с ним не сочленяется и освобождает его от участия в схватывании добычи (Britz, Kottelat, 2003).
С увеличением степени кинетизма вторичной верхней челюсти в ходе эволюции Teleostei возрастала необходимость тонкой регулировки движений входящих в нее элементов. Основным приспособлением, осуществляющим этот динамический контроль, являются подразделения внутренней (у костнопузырных Ostariophysii и паракантоптеригий Paracanthopterygii) или наружной доли (у акантоптеригий Acanthopteygii) m. adductor mandibulae (AM) (Rosen, 1982). В примитивном варианте строения этот мускул оканчивается на нижней челюсти, выполняя ее аддукцию (Gosline, 1989). Стоит отметить что, хотя эти мускульные подразделения негомологичны у разных таксонов Teleostei, чаще всего их обозначают как “максиллярные порции” или компоненты “порции А1” (Alexander, 1967c; Rosen, Patterson, 1969; Gosline, 1989). Область окончания волокон мигрирует вдоль всей длины рукоятки maxillare (Tchernavin, 1947), а также может перемещаться на соседние связки – примордиальную (lig. articulo-maxillare), как у некоторых видов сомообразных (Schaefer, Lauder, 1986) и продвинутых макрурусовых Macrouridae (см.: Tracy, 1976), или на lig. maxillo-mandibulare, как у обыкновенной морской иглы Syngnathus acus (см.: Branch, 1966). У большинства представителей Teleostei сокращение этих подразделений АМ будет ограничивать передневентральное смещение рукоятки maxillare (Tchernavin, 1947; Gosline, 1996) или контролировать повороты данной кости внутрь и наружу (Schaeffer, Rosen, 1961; Hernandez et al., 2009; Vilasri, 2013). Ряд авторов отмечают роль подразделений порции А1 не только в запуске (Alexander, 1967b; Liem, 1978; Anker, 1987), но и в ограничении/блокировке (Alexander, 1967c; Motta, 1982; Westneat, 2006) подвижности maxillare. Реже в управление вторичной верхней челюстью вступают порции А2 или А3 (Ishida, 1994). Иногда в контроль смещений maxillare включаются дополнительные висцеральные мускулы, например, компоненты m. adductor arcus palatini (AAP) (m. extensor tentaculi) у ряда видов сомов (Diogo, Vandewalle et al., 2002). Стоит отметить, что вследствие усложнения движений аппарата вторичной верхней челюсти вышеуказанные мускульные образования в свою очередь подвергаются дальнейшей дифференцировке: в частности, у морских ворчунов Haemulidae и луциановых рыб Lutjanidae конечное сухожилие порции А1 может подразделяться на латеральную и медиальную ветви, осуществляя поворот рукоятки maxillare по или против часовой стрелки (Rosen, Patterson, 1969). У австралийской вельветки Neopataecus waterhousii на вышеуказанной кости оканчиваются целых три терминальных сухожилия порций АМ (Ishida, 1994). Отсутствие или редукция порции А1 могут быть связаны с утратой подвижности вторичной верхней челюсти (Vandewalle, 2002). Мобильность praemaxillare, независимая от смещений maxillare, контролируется специализированными подразделениями АМ, такими как m. retractor praemaxillae у лорикариевых сомов Loricariidae (см.: Schaefer, Lauder, 1986).
Кинетизм вторичной верхней челюсти в сагиттальной (рис. 2) (состояния a−d) и поперечной плоскостях
Рис. 2.
Варианты кинетизма (состояния a–d) вторичной верхней челюсти у Teleostei (см. раздел о кинетизме вторичной верхней челюсти). Условные обозначения, как на рис. 1.

В большинстве литературных источников понятие “подвижность” вторичной верхней челюсти в сагиттальной плоскости у Teleostei равнозначно понятию ее “выдвижение” и используется только в случае присутствия существенного смещения praemaxillare вперед и вниз относительно этмоидного отдела нейрокраниума в сагиттальной плоскости (a) (рис. 2, 3а) (Liem, 1967; Waltzek, Wainwright, 2003). Однако данная точка зрения не учитывает иные варианты кинетизма вторичной верхней челюсти, играющие важную роль в процессе питания рыбы, такие как (b) смещение praemaxillare вместе с maxillare как единого целого (т.е. при условии их взаимной неподвижности), (c) смещение одного maxillare, а также (d) подвижность между отдельными частями praemaxillare.
Рис. 3.
Составляющие вторичной верхней челюсти, (а) демонстрирующей подвижность в сагиттальной плоскости (рис. 2, состояние а1) у лофота Lophotus lacepede (вид сбоку), praemaxillare которого способно к значительной протракции вследствие смещения maxillare (по: Olney et al., 1993); (б) выполняющей смещения в поперечной плоскости при схватывании добычи у тауматихта Акселя Thaumatichthys axeli, вид спереди (аппарат “капкана” praemaxillare) (по: Bertelsen, Struhsaker, 1977). Стрелки − направление смещения костей. Условные обозначения здесь и на рис. 4 и 5: alv – ramus alveolaris praemaxillare, art – anguloarticulare, ce – c. ethmoidalis, cp – c. palatinum, cr – os coronomeckeli, d – dentale, ech – ectoethmoideum, ect – ectopterygoideum, ent – entopterygoideum, es – eska, hm – hyomandibulare, lpr – lig. praemaxillare, mt – metapterygoideum, mx – maxillare, neur – neurocranium, o – operculum, pa – pr. ascendens praemaxillare, pal – palatinum, par – parasphenoideum, pop – praeoperculum, pp – pr. postmaxillaris, ppt – рalatopterygoideum, pr – pr. articularis praemaxillare, pret – praemaxilloethmoideum, prmx – praemaxillare, q – quadratum, ra – retroarticulare, rc – c. rostrale, so – suboperculum, sy – symplecticum, v – vomer.

Кроме того, необходимо помнить о наличии повсеместно распространенной мобильности вторичной верхней челюсти в поперечной плоскости совместно с абдукцией и аддукцией суспензориумов, которая наблюдается даже в случаях малоподвижного крепления к нейрокраниуму maxillare и praemaxillare (Lesiuk, Lindsey, 1978; Fink, 1985). Помимо добывания корма это смещение наблюдается и в ходе дыхания. Особенно большой амплитуды подобное отклонение достигает у большерота Eurypharynx pelecanoides и используется этим животным во время специфического способа питания (заглатывания), а также для локомоции по типу “дирижабль” (Nielsen et al., 1989). В большинстве работ под отведением вторичной верхней челюсти латерально подразумевается смещение дистальных концов рукояток maxillare и praemaxillare вбок относительно нейрокраниума и суспензориума (Field, 1966; Gosline, 1973; Parmentier et al., 1998; Pietsch, Orr, 2007; Greenfield et al., 2008). Такое движение приводит к формированию более широкой пасти, что важно для схватывания крупной добычи. Поэтому латеральное смещение нижнего отдела рукояток maxillare и praemaxillare часто выражено у широкоголовых хищников из окуневидных Percoidei, таких как групер Epinephelus. Однако более примитивные Teleostei могут конвергентно развивать такой же механизм работы вторичной верхней челюсти для формирования огромной пасти, как это делают рыба-солдат Holotrachys или скорпенопс Scorpaenopsis (см.: Gosline, 1966). Отведение рукояток maxillare и praemaxillare во время протракции последнего отмечено у карапуса Carapus acus, рыб-жемчужниц Encheliophis boraborensis и E. homei (см.: Parmentier et al., 1998), а также у большинства карповых Cyprinidae (см.: Gosline, 1973). В некоторых случаях амплитуда переднелатерального смещения дистальных концов вышеупомянутых костей достигает чрезвычайно большой величины, как у самцов щучьей морской собачки Neoclinus blanchardi (см.: Hongjamrassilp et al., 2018). Наиболее ярким примером подвижности вторичной верхней челюсти в поперечной плоскости является кинетизм висцерального аппарата тауматихта Акселя Thaumatichthys axeli (рис. 3б) (см.: Smith, Radcliffe, 1912). Его крупные praemaxillare могут отводиться и приводиться (открываться и закрываться) в латеро-медиальном направлении и способны зажимать между собой добычу (Bertelsen, Struhsaker, 1977), действуя как капкан у венериной мухоловки Dionaea muscipula (см.: Volkov et al., 2012). Praemaxillare способны поворачиваться на полусферической выпуклой поверхности этмоидного хряща примерно на 180° (!) относительно продольной оси головы рыбы (Bertelsen, Struhsaker, 1977).
Важно отметить, что разные способы кинетизма вторичной верхней челюсти Teleostei и механизмы, лежащие в их основе, могут сочетаться друг с другом и присутствовать в одном висцеральном аппарате (рис. 2) (Motta, 1984).
Смещения praemaxillare относительно подвижного maxillare (рис. 2, состояние а) с учетом их морфологии
Протракция вторичной верхней челюсти в ее классическом понимании (Liem, 1967) вызвана смещением praemaxillare относительно maxillare, которое чаще всего опосредовано подвижностью (активной или пассивной) последнего (рис. 2 и 3а: а1). В случае присутствия сравнительно высокоамплитудного выдвижения praemaxillare и maxillare относительно переднего конца нейрокраниума эти кости выделяют в качестве максиллярного аппарата (Olney et al., 1993; Baldwin, Johnson, 1996). В развитом максиллярном аппарате головки maxillare и praemaxillare связаны друг с другом, передним отделом нейрокраниума и суспензориумом набором связок (Branch, 1966; Rosen, 1982; Johnson et al., 1996), в частности крестообразными связками (Rosen, Patterson, 1969; Ishida, 1994). В редких случаях перемещение praemaxillare в составе вторичной верхней челюсти происходит без участия maxillare (рис. 2, а2), как это имеет место в аппарате питания четырехглазой рыбы Anableps anableps (см.: Michel et al., 2015) и других продвинутых видов карпозубообразных Cyprinodontiformes (см.: Hernandez et al., 2009), бентопелагических особей тихоокеанской желеносной рыбы Ateleopus japonicus (см.: Sasaki et al., 2006) и у некоторых представителей лорикариеподобных сомов Loricarioidea (см.: Schaefer, Lauder, 1986). У представителей Teleostei изменчивость в степени выдвижения вторичной верхней челюсти часто связана с дивергенцией в экологии питания (Motta, 1984). Многие Teleostei выдвигают только вторичную верхнюю челюсть, однако некоторые представители, такие как рыбы-бабочки Chaetodontidae (см.: Ferry-Graham et al., 2001b), губан-обманщик Epibulus insidiator (см.: Westneat, Wainwright, 1989) и цихлиды Cichlidae (см.: Waltzek, Wainwright, 2003), могут единовременно протрагировать и нижнюю челюсть. Мотта (Motta, 1988) сообщает о наличии асимметрии рта у рыб-бабочек, поскольку степень участия челюстей в формировании выдвинутого ротового отверстия у разных видов может меняться: например, протракция обеспечивается смещением вперед только нижней челюсти на 70%, а верхней челюсти на 30%, тогда как у других представителей это соотношение составляет 50 : 50.
У многих видов Teleostei, обладающих сильно выдвижимым praemaxillare, данная кость имеет ряд характерных признаков. Один из них − наличие гипертрофированного восходящего отростка (рис. 1, рис. 3а) (Rosen, Patterson, 1969; Westneat, Wainwrigth, 1989). Во многих случаях значительная длина восходящего отростка способствует увеличению амплитуды протракции несущей его кости (Tracy, 1976; Westneat, Wainwright, 1989; Ishida, 1994), однако это справедливо не для всех таксонов Teleostei (см.: Greenfield et al., 2008; Michel et al., 2015) ввиду сочетания множества морфологических факторов, оказывающих влияние на процесс перемещения вторичной верхней челюсти (Gibb, 1997). У определенных представителей наряду с подвижностью praemaxillare наблюдается редукция восходящего отростка (Hernandez et al., 2009).
Розен и Паттерсон (Rosen, Patterson, 1969) отмечали, что возникновение рострального хряща в максиллярном аппарате Teleostei связано с увеличением мобильности praemaxillare. По-видимому, изначально это образование служило упругой “подушкой” между данной костью и этмоидом, играя роль новой точки опоры для ее поворотов. В ходе онтогенеза ныне живущих Teleostei ростральный хрящ обособляется от хрящевого зачатка нейрокраниума − этмоидной пластинки, формирующейся на основе трабекул (Nel, Swanepoel, 1984). У современных таксонов ростральный хрящ опосредует протракцию praemaxillare (Ishida, 1994; Pietsch, Orr, 2007; Vilasri, 2013). У большинства представителей с ним прочно связаны восходящие отростки этих костей контрлатеральных сторон головы (Delsman, 1925), хотя иногда между ними и ростральным хрящом сохраняется подвижность (Sasaki et al., 2006). У части видов значительная амплитуда выдвижения praemaxillare взаимосвязана с хрящом крупного размера, как у восточноатлантического лофота Lophotus lacepede (см.: Olney et al., 1993) (рис. 3а) или звездочетовых Uranoscopidae (см.: Vilasri, 2013). Хартел и Стисни (Hartel, Stiassny, 1986) полагали, что истинный ростральный хрящ имеется только у Acanthomorpha, тогда как у других групп Teleostei присутствует спектр разнообразных, но нефункциональных “ложных” вариантов, которые не обусловливают протракцию praemaxillare. Эти образования негомологичны друг другу и настоящему ростральному хрящу Neoteleostei. Подобные единичные ложные хрящи можно обнаружить у диплофуса Diplophos (см.: Fink, Weitzman, 1982) и ящероголова Synodus (см.: Hartel, Stiassny, 1986), парные структуры имеются у зеленоглазки Chlorophthalmus, алепизавра Alepisaurus (см.: Hartel, Stiassny, 1986) и ипнопса Ipnops (см.: Theisen, 1966). Отсутствие рострального хряща не всегда означает потерю выдвижения вторичной верхней челюсти (Pietsch, 1978).
Механизм выдвижения praemaxillare (1−4 способа) неоднократно развивался независимо в разных группах Teleostei (см.: Schaeffer, Rosen, 1961; Lauder, 1982, 1985). (1) В качестве одного из распространенных способов можно указать поворот рукоятки maxillare вокруг собственной оси, который инициируется сокращением порции А1 (Alexander, 1967b; Liem, 1978) или запускается пассивно опусканием нижней челюсти (Alexander, 1967b, 1967c; Motta, 1982). Чаще всего последствием этого процесса является перемещение praemaxillare вперед и вниз, что вызывает уменьшение диаметра ротового отверстия рыбы. Однако у некоторых форм в результате описываемого движения древко praemaxillare, наоборот, приподнимается вверх, увеличивая амплитуду раскрывания вторичной верхней челюсти, как это происходит у сарганощуки Belonesox belizanus (см.: Ferry-Graham et al., 2010).
(2) Абдукция dentale путем натягивания lig. maxillo-mandibulare, вызывающая перенос рукоятки maxillare передневентрально, которое, в свою очередь, протрагирует praemaxillare; это самый распространенный способ выдвижения последнего у Teleostei (см.: Gosline, 1973; Liem, 1978; Motta, 1984; Anker, 1987; Waltzek, Wainwright, 2003).
(3) У многих таксонов поднятие вверх черепа служит для ориентации зубов praemaxillare вперед на добычу, поскольку вторичная верхняя челюсть прочно связана с этмоидом (Janovetz, 2005). Однако у некоторых видов Teleostei со слабо закрепленными praemaxillare и maxillare поднятие нейрокраниума будет вызывать соскальзывание этих костей с рыла рыбы передневентрально, тем самым производя их небольшую протракцию вне зависимости от движений dentale (Motta, 1982; Westneat, 2006).
(4) Многие авторы отмечают в той или иной степени выраженное влияние суспензориума на процесс перемещения вторичных передних челюстей, чаще всего передающееся посредством maxillare (Delsman, 1925; Alexander, 1967c; Waltzek, Wainwright, 2003). Существование этого механизма взаимодействия первичной верхней челюсти с максиллярным аппаратом обусловлено сдвигом конструкции суспензориума вперед, в том числе в результате перемещения рострально челюстного сустава в ходе эволюции Teleostei (см.: Дзержинский, 2005; Gardiner, 1960, 1967; Schaeffer, Rosen, 1961). У части представителей отведение суспензориумов блокирует ретракцию ранее выдвинутого praemaxillare путем “запирания” пальцевидным отростком palatinum головки maxillare (Alexander, 1967c). По мнению Лиема (Liem, 1978), у цихлид подобная абдукция напрямую вызывает протракцию восходящих отростков praemaxillare путем выдавливания их со ската нейрокраниума вперед с помощью натянутого lig. palato-palatinum ввиду отсутствия контакта премаксиллярного мыщелка maxillare с сочленовным отростком praemaxillare. Однако по другим данным (Barel et al., 1977) у цихлид последнее сочленение хорошо развито, обеспечивая взаимодействие вышеуказанных двух костей как по механизму выдвижения praemaxillare при опускании нижней челюсти (а) (Motta, 1984), так и (б) путем “выжимания” восходящих отростков костей рострально натяжением lig. intermaxillare (Conith et al., 2019), связывающего контрлатеральные головки maxillare, аналогично варианту, предложенному Лиемом (Liem, 1978). В последнем случае участие palatinum и lig. palato-palatinum выступает в качестве дополнения. Подвижность суспензориума в сагиттальной плоскости относительно нейрокраниума, а также смещение одних его костей относительно других обеспечивают протракцию максиллярного аппарата совместно с нижней челюстью у обыкновенной длиннорылой рыбы-бабочки и желтой длиннорылой рыбы-бабочки Forcipiger flavissimus (см.: Motta, 1988; Ferry-Graham et al., 2001a). В черепе у этих видов рыб челюстной сустав обладает способностью к переднезадним перемещениям.
Иные варианты кинетизма вторичной верхней челюсти (рис. 2, состояние b−d) и его отсутствие
Тенденция к увеличению использования укуса и/или тарана тела (Nelson, 1976; Muller, Osse, 1984) в процессе схватывания добычи взаимосвязана с видоизмениями устройства вторичной верхней челюсти и характером ее кинетизма (b−d) у определенных таксонов Teleostei. Однако у некоторых представителей сочетание специфической анатомии и использования указанных ниже способов мобильности maxillare приводит к росту эффективности всасывания, как, например, у топорика Мюллера Maurolicus muelleri (см.: Weitzman, 1974).
(b) Часто praemaxillare неподвижно/малоподвижно относительно maxillare и перемещается вместе с ним как единый элемент относительно нейрокраниума (рис. 4а). При этом данные кости расположены последовательно (Weitzman, 1974) или параллельно друг другу (Tyler, 1980; Parmentier et al., 2000). Между ними отсутствуют связки и мениски. Следствием подобного устройства вторичной верхней челюсти в большинстве работ авторы указывают “отсутствие или ограниченную степень ее выдвижения” (Branch, 1966; Tyler, 1980; Wassenbergh et al., 2011), упуская из виду иной вариант кинетизма.
Рис. 4.
Конструкция вторичной верхней челюсти (вид сбоку), способной к (а) смещениям неподвижных друг относительно друга maxillare и praemaxillare у полурыла Zenarchopterus kampeni (рис. 2, состояние b) (по: Vandewalle et al., 2002); (б) перемещениям одного maxillare, тогда как praemaxillare неподвижно причленено к этмоиду у магостернархуса Magosternarchus duccis (рис. 2, состояние c) (по: Lundberg et al., 1996); демонстрирующей (в) внутренний кинетизм расчлененного praemaxillare у глазчатого макрогнатуса Macrognathus aculeatus (рис. 2, состояние d) (по: Travers, 1984) или (г) отсутствие кинетизма у грязевого угря Pythonichthys macrurus (по: Eagderi, 2010). Стрелки показывают направление смещения костей, синие круги условно обозначают точку, относительно которой происходят смещения элементов. Области неподвижного прирастания maxillare/praemaxillare показаны штрихами. Условные обозначения, как на рис. 3.

При открывании dentale верхний конец maxillare пассивно поворачивается в сочленении с нейрокраниумом/palatinum, а нижний отдел рукоятки кости смещается передневентрально за счет натяжения lig. maxillo-mandibulare (как правило, это движение не достигает большой амплитуды) (Weitzman, 1974; Fink, Weitzman, 1982). У некоторых видов сустав с этмоидом формирует praemaxillare, как у жемчужницы-охотника Onuxodon fowleri (см.: Parmentier et al., 2000) и танганикского хоботнорыла Mastacembelus tanganicae (см.: Gosline, 1983), или оба – maxillare и praemaxillare (Weitzman, 1974). Praemaxillare поворачивается вверх и назад, ориентируя зубы навстречу захватываемой добыче, например, у тигровой рыбы Hydrocynus sp. (см.: Vari, 1979) и королевской макрели Scomberomorus cavalla (см.: Ferguson, 2014). Оригинальный случай кинетизма вторичной верхней челюсти выявлен у палочкохвоста Stylophorus chordatus, у которого прочно скрепленные друг с другом praemaxillare и maxillare в процессе питания способны смещаться вниз относительно нейрокраниума на расстояние, равное его длине, хотя при этом остаются неподвижными относительно нижней челюсти. Главной движущей силой данного механизма является абдукция dentale (Pietsch, 1978).
У части представителей Teleostei имеют место переднезадние перемещения всей вторичной верхней челюсти относительно нейрокраниума. Ретракция прочно скрепленных между собой maxillare и praemaxillare у танганикского хоботнорыла M. tanganicae происходит благодаря сокращению АМ, оканчивающегося на рукоятке maxillare; при этом происходит скольжение обеих костей назад по нижней стороне этмоида (Gosline, 1983). Вторичная верхняя челюсть эустомиаса Eustomias obscurus обладает способностью к смещению рострально относительно этмоидного отдела нейрокраниума, выполняемому посредством протракции переднего отдела суспензориума (Schnell, Johnson, 2017). При этом maxillare и praemaxillare выезжают вперед по вентральной поверхности vomer “на салазках” palatinum. Такая мобильность возможна благодаря наличию значительной подвижности между передним и задним отделами суспензориума данного представителя, соединенными между собой лишь связкой (Fink, 1985). При опускании вниз нижней челюсти рукоятки praemaxillare и maxillare эустомиаса относительно нее остаются неподвижными (Schnell, Johnson, 2017).
(c) При опускании dentale может наблюдаться смещение одного maxillare, тогда как praemaxillare неподвижно причленено к этмоиду для создания прочного укуса (рис. 4б), что характерно для ножетелки муравьеда Orthosternarchus tamandua (см.: Hilton, 2007), электрического угря Electrophorus electricus и гимнотов Gymnotus (см.: Datovo, Vari, 2014). У перечисленных видов praemaxillare напрямую не контактирует с maxillare, будучи связанным с последним лишь соединительной тканью. При этом maxillare осуществляет смещения своим нижним концом передневентрально, поддерживая верхнюю губу рыбы и прикрывая небольшое ротовое отверстие с боков (Hilton, 2007). Классическим примером подвижности одного maxillare, опосредуемой смещением суспензориума, вне зависимости от аддукции/абдукции dentale является кинетизм небно-максиллярного механизма большинства сомов. Palatinum, полностью обособленное от заднего отдела суспензориума, способно к самостоятельным перемещениям и служит для управления движениями maxillare. Praemaxillare обычно неподвижно прикреплено к этмоидному отделу нейрокраниума (Diogo, Vandewalle, 2003).
(d) У некоторых представителей Teleostei praemaxillare демонстрирует внутренний кинетизм одних своих отделов относительно других (рис. 4в). В качестве адаптации к увеличению ширины пасти животного в латеральном направлении альвеолярное древко вышеупомянутой кости приобретает способность отводиться вперед и наружу путем поворота в шарнирном суставе, который формируется между ним и восходящим отростком. Это смещение происходит в процессе протракции praemaxillare у некоторых удильщикообразных Lophiiformes (см.: Pietsch, Orr, 2007), таких как европейский удильщик Lophius piscatorius (см.: Field, 1966), а также у рыб-жаб семейства Batrachoididae (см.: Greenfield et al., 2008), например, у жабуна и сапу (Gosline, 1996).
У хоботнорылов рода Macrognathus praemaxillare расчленяется на длинную серию расширенных латерально и гнущихся мелкоозубленных костных пластинок, которые тянутся вдоль нижней поверхности “хобота” животного (рис. 4в). Они способны смещаться относительно друг друга и постепенно уменьшаются в размере по направлению к переднему концу головы рыбы. Количество пластинок варьирует от 9−12 пар у более примитивных видов рода Macrognathus до 14–28 пар у продвинутых форм с более длинным “хоботом” (Travers, 1984). Поддержку конструкции рыла осуществляет длинный упругий хрящ, вентрально соединенный с группой костных элементов praemaxillare соединительно-тканными волокнами. В процессе поиска пищи этот хрящ изгибается, заставляя пластинки кости смещаться относительно друг друга синхронно с этим движением, тем самым формируя подвижный слаженный механизм высокоманевренного “хобота” (Sufi, 1956; Gosline, 1983).
Настоящее (полное) отсутствие кинетизма костей вторичной верхней челюсти встречается в группе Teleostei не так часто (рис. 2, рис. 4г). Специализация к укусу во многих случаях способствует развитию жесткой верхней челюсти и поэтому не совместима с какими-либо формами ее мобильности. Отмечено (Gosline, 1973), что только у определенных Neoteleostei, например губановых рыб Labridae, наблюдается сочетание выдвижимого praemaxillare с умеренно прочным укусом. У грязевого угря Pythonichthys macrurus удлиненная преорбитальная часть черепа – praemaxilloethmoideum – служит основанием для неподвижного прирастания maxillare, несущего крупные зубы (Eagderi, 2010). У самцов пеликановой ножетелки Compsaraia samueli maxillare и praemaxillare неподвижно прикреплены к хрящу palatinum и к этмоиду (Albert, Crampton, 2009). У быстроходных видов, которые ловят добычу в акваториях со скоростным течением, применяющих при схватывании жертв элементы тарана тела, выдвижение вторичной верхней челюсти становится гидродинамически невыгодным и редуцируется. В связи с этим у меч-рыбы Xiphias gladius maxillare и praemaxillare включены в состав рострума нейрокраниума (Cancino, Burgos, 2009).
Уменьшение роли вторичной верхней челюсти в схватывании добычи (рис. 5)
Рис. 5.
Уменьшение роли вторичной верхней челюсти в схватывании добычи у Teleostei на примере устройства черепа (вид сбоку) у гигантуры Gigantura chuni (по: Konstantinidis, Johnson, 2016) и пеликановой ножетелки Compsaraia samueli (самец) (по: Albert, Crampton, 2009). Условные обозначения, как на рис. 3.

У определенных форм Teleostei, по-видимому, вследствие специализированного устройства аппарата питания роль maxillare и praemaxillare в начальном схватывании добычи снижается в пользу задействования в этом процессе других отделов черепа. Поэтому структурные элементы вторичной верхней челюсти не всегда соответствуют существующей в действительности ее “функциональной версии”, непосредственно выполняющей прием корма. Так, у некоторых видов Teleostei, использующих во время питания укус, значение костей суспензориума в захвате добычи в различной степени увеличивается и может превышать вклад вторичной верхней челюсти. Часто это сопровождается редукцией maxillare и/или praemaxillare (Johnson et al., 1996; Konstantinidis, Johnson, 2016; Воскобойникова, Назаркин, 2017). Таким образом, устройство верхней челюсти вторично приобретает предковые черты, поскольку доминирование первичной верхней челюсти в захвате добычи является исходным (анцестральным) признаком для Pisces (Cloutier, Arratia, 2004). Однако в данном случае мы имеем дело со специализацией челюстей. Функциональная замена praemaxillare и maxillare элементами суспензориума присутствует у южноокеанского кинжалозуба Anotopterus vorax: размер его длинного гипертрофированного palatinum превышает величину каудальной части первичной верхней челюсти. Описываемая кость на всем протяжении медиальной стороной прочно прикреплена к хрящу этмоидного отдела нейрокраниума и, таким образом, приобретает в нем крепкую опору для выполнения укусов добычи. Palatinum несет крупные клыки, размер которых превышает размер зубов на нижней челюсти; praemaxillare и maxillare редуцированы до тонких, грацильных косточек (Воскобойникова, Назаркин, 2017). Похожее строение черепа у гигантуры Gigantura chuni (см.: Berry, 1964; Konstantinidis, Johnson, 2016) (рис. 5а) и у воинственного батизавра Bathysaurus ferox (см.: Konstantinidis, Johnson, 2016). Длинное крупное palatinum этих рыб обладает большими клыковидными зубами, по протяженности превышает задний отдел суспензориума и выполняет главную роль в схватывании добычи. У гигантуры укус описываемой кости дополняется клыками, сидящими на entopterygoideum; praemaxillare отсутствует, а maxillare имеет вид небольшой пластиночки, находящейся в углу рта животного. У воинственного батизавра к наружной стороне palatinum прилегает длинное тонкое озубленное praemaxillare, выполняющее второстепенную роль при схватывании добычи; у взрослых форм maxillare практически полностью исчезает. Исследования Балдвин и Джонсон (Baldwin, Johnson, 1996) показали, что у личинок гигантуры и воинственного батизавра имеются хорошо развитые praemaxillare и maxillare, расположенные параллельно друг другу, а также palatinum небольшого размера. Однако в ходе дальнейшего онтогенеза челюстной аппарат данных рыб претерпевает значительные структурные изменения. Результаты исследования этих авторов подтверждают, что у двух описываемых видов схватывание добычи при помощи первичной верхней челюсти приобретено вторично.
У некоторых видов Teleostei протяженный передний отдел нейрокраниума вносит значимый вклад в схватывание добычи. Так, у глазчатого макрогнатуса Macrognathus aculeatus при приведении нижней челюсти напротив нее оказывается преорбитальный отдел черепной коробки, тогда как верхняя вторичная челюсть сдвинута рострально, выполняя в основном функцию опоры для конструкции “хобота” рыла, и слабо участвует в удержании жертвы (Travers, 1984). У многих видов угреобразных Anguilliformes имеется удлиненная передняя часть нейрокраниума, которая представляет собой результат слияния ethmoideum, vomer и praemaxillare. Клыки на praemaxilloethmovomer больше, чем на maxillare, поэтому praemaxilloethmovomer играет главную роль в укусе добычи (Eagderi, 2010). У рыбы-меча кости вторичной верхней челюсти включены в состав конусообразного рострума нейрокраниума. Поэтому длина “функциональной” верхней челюсти, составленной из vomer, mesethmoideum, frontale, maxillare и praemaxillare, значительно превышает величину нижней челюсти (Cancino, Burgos, 2009).
Некоторые представители Teleostei обладают “комплексной” верхней челюстью, включающей элементы первичной, вторичной верхних челюстей и ростральный отдел нейрокраниума. Например, у самцов пеликановой ножетелки maxillare и praemaxillare имеют весьма маленькие размеры по сравнению с длиной dentale и не вносят какой-либо существенный вклад в захват и удержание добычи (рис. 5б). Поэтому “функциональная” верхняя челюсть формируется протяженным передним отделом суспензориума (хрящ palatinum и entopterygoideum), преорбитальной частью нейрокраниума и миниатюрными костями вторичной верхней челюсти (Albert, Crampton, 2009). Отмечено, что у малоротой макропинны Macropinna microstoma maxillare настолько слабо соединено с нейрокраниумом, что не может играть роли в удерживании добычи (Chapman, 1942). Поэтому в процессе схватывания корма функцию “верхней челюсти” исполняет передняя часть черепной коробки – vomer, работающий вместе с прочно фиксированными относительно него palatinum и pterygoideum.
ЗАКЛЮЧЕНИЕ
Ведущим направлением эволюции вторичной верхней челюсти Teleostei является увеличение сложности ее строения, взаимосвязанное с формированием подвижности как целого, так и отдельных составляющих. На основе анализа литературных данных составлена качественная систематизация вариантов кинетизма вторичной верхней челюсти у Teleostei на дефинитивной стадии развития с указанием механизмов, лежащих в основе выделяемых способов подвижности. Показано, что все возможные перемещения maxillare и praemaxillare относительно нейрокраниума происходят преимущественно в сагиттальной и/или поперечной плоскостях. В сагиттальной плоскости описаны четыре варианта подвижности (a−d) двух указанных костей. Предыдущие работы подобной направленности в основном были сосредоточены на описании лишь одного способа перемещения вторичной верхней челюсти – протракции praemaxillare, другим видам ее мобильности уделено небольшое внимание.
В настоящей работе приведены примеры того, что внешне сходное устройство аппарата питания в основе может иметь разную конструкцию вторичной верхней челюсти. У ряда представителей Teleostei praemaxillare, способное к сравнительно значительной по амплитуде протракции, обладает хорошо развитым восходящим отростком, крупными менисками, опосредующими контакт кости с соседними элементами черепа, и большим ростральным хрящом. Для многих таксонов характерно наличие нескольких разных способов выдвижения praemaxillare, которые могут присутствовать в одном аппарате питания. На основании недавно полученных данных об особенностях устройства вторичной верхней челюсти у цихлид (Conith et al., 2019) нами указан приоритет lig. intermaxillare в выдвижении praemaxillare взамен ведущей роли lig. palato-palatinum в механизме, предложенном Лиемом (Liem, 1978). Уточнен вариант кинетизма вторичной верхней челюсти у эустомиаса. Приведен не указанный в предшествующих работах описываемой тематики пример внутренней подвижности praemaxillare, сочетающей смещения данной кости в сагиттальной и поперечной плоскостях, имеющейся у представителей хоботнорыловых. Отмечено, что тенденция к преимущественному использованию укуса и/или тарана тела в процессе схватывания добычи у Teleostei может приводить к уменьшению подвижности вторичной верхней челюсти. Отсутствие кинетизма костей вторичной верхней челюсти, встречающееся у представителей группы, по-видимому, приобретено ими вторично. Для определенных форм Teleostei, имеющих специализированное устройство аппарата питания, роль maxillare и praemaxillare в начальном схватывании добычи снижается в пользу задействования в этом процессе других отделов черепа – суспензориума и/или нейрокраниума.
Приведенный обзор и систематизация морфофункциональных особенностей вторичной верхней челюсти Teleostei способствуют более глубокому пониманию направлений эволюции данной группы рыб. Выполненное сравнение анатомии аппарата питания разных таксонов может быть востребовано в палеонтологических исследованиях для оценки и воссоздания вероятных связей структур черепа и особенностей экологии ископаемых форм Teleostei. Анализ широкого спектра структурных адаптаций челюстей рыб полезен для решения таксономических и филогенетических проблем у костистых рыб.
Список литературы
Воскобойникова О.С. Эволюционные преобразования висцерального скелета и вопросы филогении нототениевых рыб (Nototheniidae) // Тр. ЗИН АН СССР. Морфология и распространение рыб Южного океана. 1986. Т. 153. С. 46–66.
Воскобойникова О.С., Назаркин М.В. Остеология южноокеанского кинжалозуба Anotopterus vorax и положение рода Anotopterus в отряде Aulopifomes // Вопр. ихтиологии. 2017. Т. 57. № 1. С. 4–14.
Гуртовой Н.Н., Матвеев Б.С., Дзержинский Ф.Я. Практическая зоотомия позвоночных (низшие хордовые, бесчелюстные, рыбы). Учеб. пособ. для биол. спец. ун-тов. М.: Высшая школа. 1976. 351 с.
Дзержинский Ф.Я. Сравнительная анатомия позвоночных животных. М.: Аспект Пресс. 2005. 300 с.
Дзержинский Ф.Я., Васильев Б.Д., Малахов В.В. Зоология позвоночных // Учебник для студентов учрежд. высш. проф. образования. М.: Академия. 2013. 464 с.
Еремеева Е.Ф. Строение и развитие ротового аппарата леща, воблы и сазана // Морфологические особенности, определяющие питание леща, воблы и сазана на всех стадиях развития. М.: Изд-во АН СССР. 1948. С. 54–143.
Иорданский Н.Н. Биомеханика и функции челюстных мышц змей (Boidae и Colubridae) // Зоол. журн. 1980. Т. 59. № 9. С. 1343–1355.
Иорданский Н.Н. Кинетизм черепа ящериц (Lacertilia): происхождение, биомеханика, эволюция // Зоол. журн. 2011. Т. 90. № 8. С. 976–986.
Криволуцкий Д.А. Понятие жизненная форма в экологии животных // Экология. 1971. № 3. С. 19–25.
Решетников Ю.С. Особенности ихтиофауны в районе биостанции на р. Укаяли // Сб. науч. тр. “Экология и культивирование амазонских рыб” / Ин-т эвол. морфол. и экол. животных им. А.А. Северцова РАН. М.: Наука. 1993. С. 4−29.
Сиделева В.Г., Механикова И.В. Пищевая специализация и эволюция керчаковых рыб (Cottoidei) озера Байкал // Тр. ЗИН АН СССР. Экология и морфология рыб. 1990. Т. 222. С. 144–161.
Шарова И.Х. Жизненные формы и значение конвергенций и параллелизмов в их классификации // Журн. общ. биол. 1973. Т. 34. № 4. С. 563–570.
Albert J.S., Crampton W.G.R. A new species of electric knifefish, genus Compsaraia (Gymnotiformes: Apteronotidae) from the Amazon River, with extreme sexual dimorphism in snout and jaw length // Syst. Biodiversity. 2009. V. 7. № 1. P. 81–92.
Alexander R.McN. The functions and the mechanisms of the protrusible upper jaws of some acanthopterygian fish // J. Zool. 1967a. V. 151. P. 43−64.
Alexander R.McN. Mechanisms of the jaws of some atheriniform fish // J. Zool. 1967b. V. 151. P. 233−255.
Alexander R.McN. Functional design in fishes. London: Anchor Press. 1967c. 160 p.
Anker G.C. The morphology of joints and ligaments of a generalized Haplochromis species: H. elegans Trewavas 1933 (Teleostei, Cichlidae) // Neth. J. Zool. 1986. V. 37. № 3−4. P. 395–425.
Baldwin C.C., Johnson G.D. Interrelationships of Aulopiformes // In “Interrelationships of fishes”. Stiassny M.L.J., Parenti L.R., Johnson G.D. Eds. Acad. Press. San Diego. 1996. P. 355–404.
Barel C.D.N., van Oijen M.J.P., Witte F., Witte-Maas E.L. An introduction to the taxonomy and morphology of the haplochromine Cichlidae from Lake Victoria // Neth. J. Zool. 1977. V. 27. № 4. P. 333−389.
Berry F.H. Aspects of the development of the upper jaw bones in teleosts // Copeia. 1964. V. 1964. № 2. P. 375–384.
Bertelsen E., Struhsaker P.J. The ceratioid fishes of the genus Thaumatichthys: osteology, relationships, distribution and biology // Galathea Rep. 1977. V. 14. P. 7–40.
Branch G.M. The feeding mechanism of Syngnathus acus Linnaeus // Zool. Afr. 1966. V. 2. № 1. P. 69–89.
Britz R., Kottelat M. Descriptive osteology of the family Chaudhuriidae (Teleostei, Synbranchiformes, Mastacembeloidei), with a discussion of its relationships // Amer. Mus. Novit. 2003. № 3418. P. 1–62.
Cancino C., Burgos F. Consideraciones ecomorfologicas del mecanismo de captura de presas en Xiphias gladius (Linnaeus, 1758) // Lat. Am. J. Aquat. Res. 2009. V. 37. № 1. P. 17–28.
Chapman W.McL. The osteology and relationship of the bathypelagic fish Macropinna microstoma, with notes on its visceral anatomy // Ann. Mag. Nat. Hist. 1942. Ser. 11. V. 9. P. 272–304.
Cloutier R., Arratia G. Early diversification of actinopterygians // Recent advances in the origin and early radiation of vertebrates. Arratia G., Wilson M.V.H., Cloutier R., Eds. 2004. P. 217–270.
Conith M.R., Conith A.J., Albertson R.C. Evolution of a soft-tissue foraging adaptation in African cichlids: roles for novelty, convergence, and constraint // Evolution. 2019. V. 73. № 10. P. 2072−2084.
Datovo A., Vari R.P. The adductor mandibulae muscle complex in lower teleostean fishes (Osteichthyes: Actinopterygii): comparative anatomy, synonymy, and phylogenetic implications // J. Linn. Soc. London, Zool. 2014. V. 171. P. 554–622.
Delsman H.C. Fishes with protrusible mouths // Treubia. 1925. V. 6. № 2. P. 98–106.
Diogo R., Vandewalle P. Catfishes as a case study for discussions on general evolution: the importance of functional uncouplings in morphological macroevolution // Eur. J. Morphol. 2003. V. 41. № 3/4. P. 139–148.
Eagderi S. Structural diversity in the cranial musculoskeletal system in Anguilliformes: an evolutionary-morphological study // Cand. Sci. (Biol.) Dissertation, Gent, Belgium: Universiteit Gent. 2010. P. 1–157.
Ferguson A.R. Feeding performance of king mackerel, Scomberomorus cavalla // Extended Abstract of Cand. Sci. (Biol.) Dissertation, Tampa, FL, USA: University of South Florida. 2014. P. 1–32.
Ferry-Graham L.A., Wainwright P.C., Bellwood D.R. Prey capture in long-jawed butterflyfishes (Chaetodontidae): the functional basis of novel feeding habits // J. Exp. Mar. Biol. Ecol. 2001a. V. 256. P. 167–184.
Ferry-Graham L.A., Wainwright P.C., Hulsey C.D., Bellwood D.R. Evolution and mechanics of long jaws in butterflyfishes (Family Chaetodontidae) // J. Morph. 2001b. V. 248. P. 120–143.
Ferry-Graham L.A., Hernandez L.P., Gibb A.C., Pace C. Unusual kinematics and jaw morphology associated with piscivory in the poeciliid, Belonesox belizanus // Zoology (Jena). 2010. V. 113. P. 140–147.
Field J.G. Contributions to the functional morphology of fishes. Part II. The feeding mechanism of the angler-fish, Lophius piscatorius Linnaeus // Zool. Afr. 1966. V. 2. № 1. P. 45–67.
Fink W.L. Phylogenetic interrelationships of the stomiid fishes (Teleostei: Stomiiformes) // Misc. Publ. − Mus. Zool. Univ. Mich. 1985. № 171. P. 1–127.
Fink W.L., Weitzman S.H. Relationships of the stomiiform fishes (Teleostei), with a description of Diplophos // Bull. Mus. Comp. Zool. 1982. V. 150. № 2. P. 31–93.
Gardiner B.G. A revision of certain actinopterygian and coelacanth fishes, chiefly from the Lower Lias // Bull. Br. Mus. (Nat. Hist.) Geol. 1960. V. 4. № 7. P. 242–382.
Gardiner B.G. The significance of the preoperculum in actinopterygian evolution // J. Linn. Soc. London, Zool. 1967. V. 47. № 311. P. 197–209.
Gibb A.C. Do flatfish feed like other fishes? A comparative study of percomorph prey-capture kinematics // J. Exp. Biol. 1997. V. 200. P. 2841–2859.
Gosline W.A. Comments on the classification of the percoid fishes // Pac. Sci. 1966. V. 20. № 4. P. 409–418.
Gosline W.A. Considerations regarding the phylogeny of cypriniform fishes, with special reference to structures associated with feeding // Copeia. 1973. V. 1973. № 4. P. 761–776.
Gosline W.A. The relationships of the Mastacembelid and Synbranchid fishes // Jpn. J. Ichthyol. 1983. V. 29. № 4. P. 323−328.
Gosline W.A. Jaw structures and movements in higher teleostean fishes // Jpn. J. Ichthyol. 1987. V. 34. P. 21–32.
Gosline W.A. Two patterns of differentiation in the jaw musculature of teleostean fishes // J. Zool. 1989. V. 218. P. 649–661.
Gosline W.A. Structures associated with feeding in three broad-mouthed, benthic fish groups // Environ. Biol. Fishes. 1996. V. 47. P. 399–405.
Grande T., Poyato-Ariza F.J. Reassessment and comparative morphology of the gonorynchiform head skeleton // Gonorynchiformes and Ostariophysan relationships. A Comprehensive Review. Ser. Teleost. Fish Biol. Grande T., Poyato-Ariza F.J., Diogo R., Eds. 2010. P. 1–39.
Greenfield D.W., Winterbottom R., Collette B.B. Review of the toadfish genera (Teleostei: Batrachoididae) // Proc. Calif. Acad. Sci. 2008. Ser. 4. V. 59. № 15. P. 665–710.
Gromova E.S., Makhotin V.V. Maxillary apparatus in the feeding of the silver carp Hypophthalmichthys molitrix (Cyprinidae) // J. Ichthyol. 2018. V. 58. № 6. P. 857 –877.
Hartel K.E., Stiassny M.L.J. The identification of larval Parasudis (Teleistei, Chlorophthalmidae); with notes on the anatomy and relationships of aulopiform fishes // Breviora Geoi. Asturica. 1986. № 487. P. 1–23.
Hernandez L.P., Gibb A.C., Ferry-Graham L.A. Trophic apparatus in cyprinodontiform fishes: functional specializations for picking and scraping behaviors // J. Morph. 2009. V. 270. P. 645–661.
Hilton E.J. Redescription of Orthosternarchus tamandua (Boulenger, 1898) (Gymnotiformes, Apteronotidae), with reviews of its ecology, electric organ discharges, external morphology, osteology, and phylogenetic affinities // Proc. Acad. Nat. Sci. Philadelphia. 2007. V. 156. P. 1–25.
Hongjamrassilp W., Summers A.P., Hastings P.A. Heterochrony in fringeheads (Neoclinus) and amplification of an extraordinary aggressive display in the Sarcastic fringehead (Teleostei: Blenniiformes) // J. Morph. 2018. V. 1. P. 1–10.
Ishida M. Phylogeny of the suborder Scorpaenoidei (Pisces: Scorpaeniformes) // Bull. Nansei Natl. Fish. Res. Inst. 1994. № 27. P. 1–112.
Janovetz J. Functional morphology of feeding in the scale-eating specialist Catoprion mento // J. Exp. Biol. 2005. V. 208. P. 4757–4768.
Johnson G.D., Baldwin C.C., Okiyama M., Tominaga Y. Osteology and relationships of Pseudotrichonotus altivelis (Teleostei: Aulopiformes: Pseudotrichonotidae) // Ichthyol. Res. 1996. V. 43. P. 17–45.
Kirckhoff H. Funktionell-anatomische Untersuchung des Visceralaooarates von Clupea harengus L. // Zool. Jahrb. Abt. Anat. Ontog. Tiere. 1958. V. 76. № 4. P. 462−539.
Konstantinidis P., Harris M.P. Same but different: ontogeny and evolution of the musculus adductor mandibulae in the Tetraodontiformes // J. Exp. Zool. 2011. V. 316. № 1. P. 10–20.
Konstantinidis P., Johnson G.D. Osteology of the telescopefishes of the genus Gigantura (Brauer, 1901), Teleostei: Aulopiformes // J. Linn. Soc. London, Zool. 2016. V. 179. № 2. C. 338–353.
Lauder G.V. Patterns of evolution in the feeding mechanism of actinopterygian fishes // Amer. Zool. 1982. V. 22. P. 275–285.
Lauder G.V. Aquatic feeding in lower vertebrates // In “Functional vertebrate morphology”. Hildebrand M., Bramble D.M., Liem K.F., Wake D.B., Eds. Cambridge, MA: Harvard Univ. Press. 1985. P. 210–229.
Lesiuk T.P., Lindsey C.C. Morphological peculiarities in the neck-bending amazonian characoid fish Rhuphiodon vulpinus // Can. J. Zool. 1978. V. 56. № 4. P. 991–997.
Liem K.F. A morphological study of Luciocephalus pulcher, with notes on gular elements in other recent teleosts // J. Morph. 1967. V. 121. P. 103−133.
Liem K.F. Modulatory multiplicity in the functional repertoire of the feeding mechanism in cichlid fishes // J. Morph. 1978. V. 158. P. 323−360.
Lundberg J.G., Fernandes C.C., Albert J.S., Garcia M. Magosternarchus, a new genus with two new species of electric fishes (Gymnotiformes: Apteronotidae) from the Amazon River basin, South America // Copeia. 1996. V. 1996. № 3. P. 657–670.
Michel K.B., Aerts P., Gibb A.C., Wassenbergh S. Functional morphology and kinematics of terrestrial feeding in the largescale foureyes (Anableps anableps) // J. Exp. Biol. 2015. V. 218. P. 2951–2960.
Motta P.J. Functional morphology of the head of the inertial suction feeding butterflyfish, Chaetodon miliaris (Perciformes, Chaetodontidae) // J. Morph. 1982. V. 174. P. 283–312.
Motta P.J. Mechanics and functions of jaw protrusion in teleost fishes: a review // Copeia. 1984. V. 1984. P. 1–18.
Motta P.J. Functional morphology of the feeding apparatus of ten species of Pacific butterflyfishes, (Perciformes, Chaetodontidae): an ecomorphological approach // Environ. Biol. Fishes. 1988. V. 22. № 1. P. 39–67.
Muller M., Osse J.W.M. Hydrodynamics of suction feeding in fish // Trans. Zool. Soc. Lond. 1984. V. 37. P. 51–135.
Nel P.C., Swanepoel J.H. The development of the nasal capsule of the silver carp Hypophthalmichthys molitrix (Valenciennes) // Zool. Afr. 1984. V. 19. № 4. P. 309–313.
Nelson J.S. Fishes of the world // New York, London: John Wiley & Sons, Inc. 1976. P. 1−416.
Nelson J.S. Fishes of the world // New Jersey: John Wiley & Sons, Inc. 2006. Nielsen J.G., Bertelsen E., Jespersen A. The Biology of Eurypharynx pelecanoides (Pisces, Eurypharyngidae) // Acta Zool. (Stockholm). 1989. V. 70. № 3. P. 187–197.
Olney J.E., Johnson G.D., Baldwin C.C. Phylogeny of lampridiform fishes // Bull. Mar. Sci. 1993. V. 52. № 1. P. 137–169.
Osse J.W.M. Functional morphology of the head of the perch (Perca fluviatilis L.): an electromyographic study // Neth. J. Zool. 1969. V. 19. P. 289−392.
Parmentier E., Chardon M., Poulicek M. et al. Morphology of the buccal apparatus and related structures in four species of Carapidae // Aust. J. Zool. 1998. V. 46. P. 391–404.
Parmentier E., Castro-Aguirre J.L., Vandewalle P. Morphological comparison of the buccal apparatus in two bivalve commensal Teleostei, Encheliophis dubius and Onuxodon fowleri (Ophidiiformes, Carapidae) // Zoomorphology. 2000. V. 120. P. 29–37.
Pietsch T.W. The feeding mechanism of Stylephorus chordatus (Teleostei: Lampridiformes): functional and ecological implications // Copeia. 1978. V. 1978. № 2. P. 255–262.
Pietsch T.W., Orr J.W. Phylogenetic relationships of deep-sea anglerfishes of the suborder Ceratioidei (Teleostei: Lophiiformes) based on morphology // Copeia. 2007. V. 2007. № 1. P. 1–34.
Regan C.T. The classification of the teleostean fishes of the order Ostariophysi. I. Cyprinoidea // Ann. Mag. Nat. Hist. 1911. Ser. 8. V. 8. № 43. P. 13–32.
Roberts T.R. Osteology and relationships of characoid fishes, particularly the genera Hepsetus, Salminus, Hoplias, Ctenolucius and Acestrorhynchus // Proc. Calif. Acad. Sci. 1969. ser. 4. V. 36. № 15. P. 391−500.
Rosen D.E. Telostean interrelationships, morphological function and evolutionary inference // Am. Zool. 1982. V. 22. P. 261−273.
Rosen D.E., Patterson C. The structure and relationships of the paracanthopterygian fishes // Bull. Am. Mus. Nat. Hist. 1969. V. 141. № 3. P. 361–474.
Sanford C.P.J., Lauder G.V. Kinematics of the tongue-bite apparatus in osteoglossomorph fishes // J. Exp. Biol. 1990. V. 154. P. 137−162.
Sasaki K., Tanaka Y., Takata Y. Cranial morphology of Ateleopus japonicus (Ateleopodidae: Ateleopodiformes), with a discussion on metamorphic mouth migration and lampridiform affinities // Ichthyol. Res. 2006. V. 53. P. 254–263.
Schaefer S.A., Lauder G.V. Historical transformation of functional design: evolutionary morphology of feeding mechanisms in loricarioid catfishes // Syst. Zool. 1986. V. 35. № 4. P. 489–508.
Schaeffer B., Rosen D.E. Major adaptive levels in the evolution of the actinopterygian feeding mechanism // Am. Zool. 1961. V. 1. P. 187−204.
Schnell N.K., Johnson G.D. Evolution of a functional head joint in deep-sea fishes (Stomiidae) // PloS One. 2017. V. 12. № 2. P. 1–11.
Sidlauskas B.L., Vari R.P. Phylogenetic relationships within the South American fish family Anostomidae (Teleostei, Ostariophysi, Characiformes) // J. Linn. Soc. London, Zool. 2008. V. 154. P. 70–210.
Smith H.M., Radcliffe L. Description of a new family of pediculate fishes from Celebes // Proc. U.S. Natn. Mus. 1912. V. 42. P. 579−581.
Staab K.L., Ferry L.A., Hernandez L.P. Comparative kinematics of cypriniform premaxillary protrusion // Zoology (Jena). 2012. V. 115. P. 65–77.
Sufi S.M. Revision of the Oriental fishes of the family Mastacembelidae // Bull. Raff. Mus. Singap. 1956. V. 27. P. 93–146.
Tchernavin V.V. Further notes on the structure of the bony fishes of the order Lyomeri (Eurypharynx) // J. Linn. Soc. London, Zool. 1947. V. 41. № 280. P. 377–393.
Theisen B. On the cranial morphology of Ipnops murrayi Gunther, 1878 with special reference to the relations between the eyes and the skull // Galathea Rep. 1966. V. 8. P. 7–18.
Tracy M. Functional anatomy of the Macrouridae (Teleostei, Gadiformes) // Cand. Sci. (Biol.) Dissertation, Cambridge, MA, USA: Massach. Inst. Technol., and Falmouth, MA, USA. Woods Hole Ocean. Inst. 1976. P. 1–311.
Travers R.A. A review of the Mastacembeloidei, a suborder of synbranchiform teleost fishes. Part 1. Anatomical descriptions // Bull. Br. Mus. (Nat. Hist.) Zool. 1984. V. 46. № 1. P. 1–133.
Tyler J.C. Osteology, phylogeny, and higher classification of the fishes of the order Plectognathi (Tetraodontiformes) // United States. Nat. Mar. Fish. Serv. (U.S.), Circ. U.S., Natl. Oceanic Atmos. Adm. 1980. V. 434. P. 1–422.
Vandewalle P., Lambert V., Parmentier E. Particularities of the bucco-pharyngeal apparatus in Zenarchopterus kampeni (Pisces: Hemiramphidae) and their probable significance in feeding // Belg. J. Zool. 2002. V. 132. № 2. P. 125–132.
Vari R.P. Anatomy, relationships and classification of the families Citharinidae and Distichodontidae (Pisces, Characoidea) // Bull. Br. Mus. Nat. Hist. Zool. 1979. V. 36. № 2. P. 261–344.
Vigliotta T.R. A phylogenetic study of the African catfish family Mochokidae (Osteichthyes, Ostariophysi, Siluriformes), with a key to genera // Proc. Acad. Nat. Sci. Philadelphia. 2008. V. 157. P. 73−136.
Vilasri V. Comparative anatomy and phylogenetic systematics of the family Uranoscopidae (Actinopterygii: Perciformes) // Mem. Fac. Fish. Sci., Hokkaido Univ. 2013. V. 55. № 1−2. P. 1–106.
Volkov A.G., Murphy V.A., Clemmons J.I. et al. Energetics and forces of the Dionaea muscipula trap closing // J. Plant Physiol. 2012. V. 169. № 1. P. 55–64.
Waltzek T.B., Wainwright P.C. Functional morphology of extreme jaw protrusion in neotropical cichlids // J. Morph. 2003. V. 257. P. 96–106.
Wassenbergh S., Roos G., Aerts P., Herrel A., Adriaens D. Why the long face? A comparative study of feeding kinematics of two pipefish with different snout lengths // J. Fish Biol. 2011. V. 78. № 6. P. 1786–1798.
Weitzman S.H. Osteology and evolutionary relationships of the Sternoptychidae with a new classification of stomiatoid families // Bull. Am. Mus. Nat. Hist. 1974. V. 153. Art. 3. P. 327–478.
Westneat M.W. Feeding, function and phylogeny: analysis of historical biomechanics in labrid fishes using comparative methods // Syst. Biol. 1995. V. 44. № 3. P. 361–383.
Westneat M.W. Skull biomechanics and suction feeding in fishes // Fish physiology: fish biomechanics. Lauder G.V., Shadwick R.E. Eds. Chapt. 2. Fish Physiol. Series. 2006. V. 23. P. 29–75.
Westneat M.W., Wainwright P.C. The feeding mechanism of the sling-jaw wrasse Epibulus insidiator (Labridae: Teleostei): evolution of a novel functional system // J. Morph. 1989. V. 202. P. 129–150.
Дополнительные материалы отсутствуют.
Инструменты
Биология моря