

Биология моря, 2023, T. 49, № 1, стр. 37-44
Оценка пространственной неоднородности условий культивирования тихоокеанской устрицы Magallana gigas (Thunberg, 1793) с помощью модели управления марифермами FARM в бухте Воевода (Японское море)
С. В. Катрасов 1, *, А. Н. Бугаец 1, В. В. Жариков 1
1 Тихоокеанский институт географии ДВО РАН
690041 Владивосток, Россия
* E-mail: sergey_katrasov@mail.ru
Поступила в редакцию 13.05.2022
После доработки 06.07.2022
Принята к публикации 06.10.2022
- EDN: LRWITV
- DOI: 10.31857/S0134347523010059
Аннотация
В настоящем исследовании с помощью модели управления ресурсами аквакультуры на фермах FARM (Farm Aquaculture Resource Management) рассчитана потенциальная продуктивность плантаций устрицы Magallana gigas (Thunberg, 1793) (Bivalvia: Ostreidae) в б. Воевода (Японское море, зал. Петра Великого, о-в Русский). При расчете продуктивности садковых и придонных плантаций M. gigas использовали полученные ранее результаты численного моделирования динамики гидрологических параметров бухты с помощью открытого программного комплекса Delft3D-Flow с учетом притока пресной воды в бухту в 1990–2019 гг. Для демонстрации неоднородности условий выращивания M. gigas были выбраны три участка размером 100 × 100 м, расположенные в бухтах Круглая и Мелководная, а также в центральной части б. Воевода. Для каждого участка выполнено моделирование 28 периодов выращивания, продолжительностью 22 мес. каждый. Выбраны максимальные значения урожайности устрицы на второй год выращивания, выраженные в тоннах общей сырой массы, и представлены гистограммы распределения для каждого района. Показано, что по гидрологическим и биологическим показателям условия выращивания даже в небольшой по размерам б. Воевода неоднородны. Установлено, что вариабельность продуктивности M. gigas связана преимущественно с крайней неравномерностью перераспределения первичной продукции в б. Воевода под воздействием гидродинамических факторов.
Успешное функционирование хозяйств марикультуры возможно только при комплексном учете условий культивирования, что позволяет уже на стадии проектирования обосновать как выбор технологии выращивания гидробионтов, так и местоположение плантаций (Гаврилова, Кучерявенко, 2011; Гаврилова, 2012; Гаврилова, Кондратьева, 2015; Гаврилова, Ким, 2016). Продуктивность культивирования гидробионтов определяется физическими, химическими и биологическими параметрами окружающей среды. Прежде всего это концентрация растворенного кислорода и питательных веществ в водных массах, переносимых локальными морскими течениями. Выводы о пригодности акватории для выращивания того или иного вида, как правило, делают на основе данных сети мониторинга Росгидромета, плотность которой в прибрежной зоне зал. Петра Великого недостаточна для обоснования широкомасштабного развития марикультуры в данном районе (Гаврилова, Ким, 2016).
Методики оценки потенциала акваторий для развития марикультуры, основанные на режимной гидрометеорологической информации и средних для акваторий значениях океанографических показателей (Гайко, 2017), содержат допущения, ограничивающие объективный анализ возможности устойчивого развития марикультуры и эффективного использования ресурсов прибрежных зон. Поскольку условия выращивания могут различаться даже в небольших бухтах, подобные расчеты не дают полного представления о пространственной дифференциации продуктивных возможностей акватории, а также о локальных рисках культивирования гидробионтов.
Дополнительные натурные исследования позволяют сделать выводы о текущем состоянии среды, планктона и бентоса, однако возможности подобных съемок достаточно ограничены, а из-за кратковременности они не могут улучшить точность статистических характеристик гидродинамического и биогеохимического режима акваторий. Привлечение данных дистанционного зондирования и использование различных геостатистических моделей могут повысить точность фиксации границ распространения параметров в определенный момент времени. Однако для комплексного анализа динамики основных факторов, определяющих устойчивость культивирования и возможность развития ферм с учетом рисков, обусловленных флуктуациями параметров среды, подобной информации во многих случаях недостаточно.
Перспективы гидробиологических исследований в области марикультуры, как и получение новых экспериментальных данных, связаны с развитием математических моделей, в которых были бы учтены все имеющиеся сведения по каждой акватории и конкретному объекту выращивания.
В современной стратегии комплексного управления хозяйствами марикультуры используется понятие приемной емкости (carrying capacity) (Nobre et al., 2005; McKindsey et al., 2006), которая включает четыре иерархические категории: физическую, продукционную, экологическую и социальную. Из них в настоящей работе рассмотрены первые две. Физическая емкость – географически выделенная часть акватории, подходящая по абиотическим свойствам (гидродинамический и гидрохимический режимы) для определенных объектов культивирования и типов аквакультуры. Она может быть установлена с помощью комбинации гидродинамических моделей, обеспечивающих гидрологическую информацию о районе размещения ферм. Продукционная емкость – максимально возможное количество продукции при культивировании конкретных видов. К настоящему времени накоплен большой опыт в области биогеохимического моделирования параметров, определяющих продуктивность хозяйств марикультуры. Обобщение, анализ, а также визуализация результатов гидродинамического и биогеохимического моделирования проводятся с помощью географических информационных систем (ГИС).
В связи с этим актуальной задачей является зонирование акваторий на основе данных комплексных исследований с привлечением результатов гидродинамического и биогеохимического моделирования, позволяющее объективно учесть условия культивирования гидробионтов. В локальном масштабе участков марикультуры несомненный интерес представляет оценка пространственной неоднородности условий культивирования, обеспечивающая мотивированный выбор конкретных мест донного выращивания и размещения установок с учетом эффективности производства и рисков, возникающих в результате воздействия неблагоприятных гидрометеорологических факторов. Данное исследование посвящено методическим и практическим подходам к решению обозначенных выше задач.
МАТЕРИАЛ И МЕТОДИКА
Б. Воевода, площадь которой составляет около 4 км2, расположена в юго-западной части о-ва Русский (зал. Петра Великого, Японское море) (рис. 1). Ее акватория включает две бухты второго порядка – б. Круглую (1.6 км2) и б. Мелководную (2.4 км2). Климат в районе исследований умеренный, муссонный. Зимой северные ветра обусловливают холодную ясную погоду, летом преобладают южные ветра. Преимущественным направлением ветров является северное (37%). По данным метеостанции “Владивосток” (31960) Приморского УГМС за период моделирования (1989–2019 гг.) средняя скорость ветра составляла 5.8 м/с, максимальная – 36.0 м/с. Среднегодовое количество осадков составляет 848 мм; среднемесячная относительная влажность воздуха варьирует от 58 до 91%; минимальная, максимальная и средняя температура воздуха характеризуются величинами –25.2, 32.8 и 9.2°С соответственно.
Рис. 1.
Карта-схема района исследований, рельеф дна б. Воевода, выбранные участки выращивания устрицы (I, II, III).
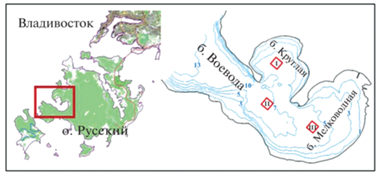
Водообмен в б. Воевода определяется циклонической циркуляцией течений, средняя величина прилива составляет 0.2 м с максимальной амплитудой 0.43 м (Барабанщиков и др., 2015). Рельеф дна большей части б. Мелководная очень пологий, без резких перепадов. Подводные береговые склоны подразделяются на равнины и участки с более широкой амплитудой рельефа и контрастными геоморфологическими формами. На входе в бухту находятся небольшие по площади валунно-глыбовые развалы с хорошо выраженными склонами (Барабанщиков и др., 2015).
Площадь водосбора б. Воевода, составляющая 23.59 км2 (17.18 км2 – р. Русская), покрыта густым широколиственным лесом. Почвенный покров представлен буроземами (Иванов, 1976), гидрологический режим которых характеризуется свободным внутрипочвенным дренажем, что обусловлено легкосуглинистым механическим составом поверхностных горизонтов и повышенной скелетностью (до 80% от объема почвенной массы) нижней части профиля. Содержание скелета уже на глубине 11–20 см достигает 50% от объема почвенной массы и нарастает вниз по профилю, переходя в грубообломочные элюво-делювиальные отложения. Минимальный измеренный расход р. Русская составляет 0.25 м3/с. Максимальный модуль стока, определенный по результатам физического моделирования, составляет 840 л с/км2, что соответствует расходу воды 14.4 м3/с (Катрасов и др., 2021а).
В мористой и средней частях бухты осуществляется свободный обмен с морскими водами. В бухтах Мелководная и Круглая под влиянием прилива и батиметрии формируется циклоническая циркуляция. Режим солености определяется водным балансом, который зависит от притока вод Амурского залива и внутреннего водообмена в бухте, а также от притока пресных вод с водосбора и от осадков, выпадающих непосредственно на акваторию. Во время сильных дождей в период циклонов и выхода тайфунов граница влияния стока р. Русская со значениями солености 20–24‰ в среднем и придонном слоях достигает входа в б. Круглая и распространяется до центральной части б. Воевода (Барабанщиков и др., 2015).
Физическая емкость культивирования Magallana gigas для б. Воевода со стороны входа в эту бухту ограничена воздействием морских вод Амурского залива с соленостью на 4–6‰ выше оптимального диапазона (23–28‰) для выращивания этого вида (Кучерявенко, Жук, 2011). Граница выращивания M. gigas расположена по линии 80%-ной обеспеченности соленостью 28‰. В кутовой части б. Воевода сток р. Русская, объем которого за катастрофические паводки 1990 г. (0.00424 км3 12.07–20.07) и 2016 г. (0.003 км3 29.08–12.09) вполне сопоставим с объемом б. Мелководная (0.00469 км3), может вызывать сильное опреснение. Однако тихоокеанская устрица способна выдерживать подобное краткосрочное воздействие, и границы культивирования в этом районе проведены по изобатам 1 м для донного и 5 м для садкового выращивания (Катрасов и др., 2021б).
Модель управления ресурсами аквакультуры на фермах FARM (Farm Aquaculture Resource Management, www.farmscale.org) (Ferreira et al., 2007) предназначена для решения задач, связанных с размещением плантаций, выбором культивируемых видов, экологической оценкой воздействия хозяйств на окружающую среду и с экономической оптимизацией марикультуры. В рамках FARM для определения продукции и оценки эвтрофикации применяется комбинация физических, биогеохимических моделей и моделей роста моллюсков. Основные особенности FARM связаны с использованием временных рядов исходных данных, позволяющих рассчитать урожайность гидробионтов с учетом геометрического размера марифермы, плотности культивирования, параметров среды и локальных трофических условий выращивания.
Программный комплекс включает в себя экофизиологические модели жизнедеятельности для семи видов двустворчатых моллюсков, в том числе для тихоокеанской устрицы M. gigas (Hawkins, Bayne, 1985, 1992; Solidoro, Pastres et al., 2000; Hawkins et al., 2002a, 2002b; Bricker et al., 2003; Nunes et al., 2003; Rueda et al., 2005; McCausland et al., 2006; McKindsey et al., 2006; Ferreira et al., 2008; Brigolin et al., 2009). Входные данные представлены временными рядами температуры воды, солености, концентраций хлорофилла-а (chlorophyll-a, Chl-a), растворенного кислорода (dissolved oxygen, DO), концентрации взвешенных твердых частиц (total particulate matter, TPM) и твердых частиц органического вещества (particulate organic matter, POM). Данные задаются с суточным или большим временным разрешением (в последнем случае производится линейная интерполяция в суточные интервалы). Элиминация гидробионтов, связанная с не учитываемыми в модели факторами, задается как параметр.
Общий подход, используемый в FARM для моделирования изменения характеристик водной массы под воздействием аквакультуры, описывается системой уравнений для каждого заданного биогеохимического параметра, которая в общем случае может быть выражена в виде квазилинейного дифференциального уравнения 1-го порядка с частными производными (1):
(1)
$\frac{{\partial C}}{{\partial t}} = \,\,~ - u\frac{{\partial C}}{{\partial x}} - w\frac{{\partial C}}{{\partial z}} + ~f~\left( {C,~\sum\nolimits_{i = 1}^{i = m} {{{n}_{i}}{{\gamma }_{i}}} } \right),$Первый член уравнения (1) описывает усредненный одномерный горизонтальный поток, основанный на текущей скорости течения и пространственных размерах фермы. Размеры фермы могут быть определены как ряд смежных секций, что позволяет проводить анализ различных вариантов выращивания гидробионтов. Второй член уравнения (1) описывает падение взвешенных частиц. На каждом шаге рассчитывается динамическая вязкость среды на основе температуры и солености воды с использованием уравнения Стокса, а также определяется скорость оседания и отложения взвешенных частиц с учетом их гидравлической крупности. Третий член в уравнении (1) описывает изменение концентрации ресурсов C (Chl-а, POM и TPM) на каждом расчетном шаге моделирования в результате поглощения (Chl-a, POM, TPM) и выделения (POM, TPM) вещества с учетом численности и размера моллюсков.
Модель предназначена для временных интервалов, характерных для циклов выращивания (от нескольких месяцев до 2–3 лет). Временной шаг моделирования подбирается автоматически на основе физических размеров секции марифермы и условий критерия Куранта–Фридрихса–Леви. Результатом модельных расчетов являются временные ряды концентраций хлорофилла-а, растворенного кислорода, взвешенных твердых частиц, взвешенного органического вещества и биомассы гидробионтов на каждый день цикла культивирования.
РЕЗУЛЬТАТЫ
Для демонстрации неоднородности условий выращивания в б. Воевода были выбраны три участка размером 100 × 100 м, расположенные в бухтах Круглая и Мелководная, а также в центральной части б. Воевода в районе входа в б. Круглая (рис. 1). При расчете продуктивности садковых и придонных плантаций Magallana gigas на этих участках были использованы полученные ранее результаты численного моделирования динамики гидрологических параметров бухты с помощью открытого программного комплекса Delft3D-Flow и с учетом притока пресной воды в акваторию бухты в 1990–2019 гг. Горизонтальное разрешение расчетной сетки гидродинамической модели составляло 20 м, вертикальное разрешение – 3 σ-слоя, границы которых установлены в 10% от поверхности и дна бухты (Катрасов и др., 2021а). В качестве исходных данных использовали временные ряды температуры воды, солености, концентраций Chl-a, DO, TPM и POM суточного разрешения, полученные при обработке выходных данных модели DELWAQ (DELft WAter Quality) (Катрасов и др., 2021б). Плотность посадки спата, товарные характеристики и период выращивания были заданы в соответствии с региональными рекомендациями (Кучерявенко, Жук, 2011). Смертность гидробионтов, связанная с не учитываемыми моделью FARM факторами, задана равной 10% за период культивирования. Для расчета продуктивности донного выращивания использовали данные придонного, а для расчета продуктивности садкового выращивания – среднего σ-слоя модели Delft3D Flow.
Использованные в работе данные 29-летнего биогеохимического моделирования (Катрасов и др., 2021б; Бугаец и др., 2022) позволили с помощью FARM выполнить расчет для 28 рекомендованных региональной биотехнологией культивирования 22-месячных периодов выращивания устрицы на выбранных участках (рис. 1). Для каждого периода выращивания были выбраны максимальные значения урожайности устрицы, выраженные в тоннах общей сырой массы (total fresh weight, TFW) (см. рис. 2). На основе анализа представленных графиков можно заключить, что по сравнению с внутренними бухтами район II, расположенный в центральной части б. Воевода, характеризуется более жесткими гидрологическими условиями для выращивания тихоокеанской устрицы. Лишь в отдельные годы (1991–1992, 1997 и 2003) за 22-месячный период культивирования здесь удалось бы вырастить урожай более 5 т/га при садковом культивировании и от 0.5 до 2.5 т/га на пастбищных плантациях. В остальные годы значения модельной продуктивности составили от 0 (2016 г.) до 486 (в среднем 66) кг/га.
Рис. 2.
График продуктивности (т/га) садкового и донного культивирования устрицы Magallana gigas для трех выделенных районов в б. Воевода по данным моделирования с помощью FARM.

Гистограммы распределения для каждого из районов (рис. 3) характеризуют вариабельность значений TFW для всех расчетных периодов моделирования. Для района II, расположенного в б. Воевода, в 70% процентах случаев модельная продуктивность не превышала 0.3 и 2.0 т/га для донного и садкового культивирования соответственно. Для участков, расположенных в бухтах Круглая и Мелководная, около 60% значений модельной продуктивности для донного выращивания находилось в диапазоне 42–50 т/га. При садковом выращивании в 50% случаев продуктивность района I (б. Круглая) изменялась от 78 до 95 т/га. Продуктивность района III (б. Мелководная) в 80% случаев превышала 50 т/га, при этом значения TWF были достаточно равномерно распределены в указанном диапазоне.

ОБСУЖДЕНИЕ
Основными источниками первичной продукции в районе исследования являются заросли морской травы Zostera marina и фитопланктон. Заросли зостеры расположены преимущественно во внутренних бухтах. Перераспределение первичной продукции по акватории при воздействии гидродинамических факторов крайне неравномерное. Во внутренних бухтах наблюдается ее избыток (в среднем 6–8 мгC/м3 POM и 1.5 мг/м3Chl-a), а в более мористых участках – дефицит (в среднем 2–3 мгC/м3 POM и около 0.5 мг/м3Chl-a). Низкая урожайность Magallana gigas в районе II (рис. 1) связана с пониженным количеством доступной для гидробионтов пищи, что вызвано сочетанием этих факторов.
Гистограммы POM и Chl-a для каждого из рассматриваемых участков, построенные за период моделирования (рис. 4) для районов, расположенных во внутренних бухтах, достаточно хорошо описываются нормальным распределением. Параметры нормального распределения для района I (б. Круглая) и III (б. Мелководная) различаются незначительно: среднее и дисперсия для POM составляют в первом районе 6.1 и 1.97, во втором – 6.2 и 1.76 соответственно. Для распределения Chl-a в первом районе среднее значение составляет 1.18, дисперсия – 0.37, во втором районе – 1.2 и 0.33 соответственно. Во втором районе, расположенном в центральной части б. Воевода, гистограммы Chl-a и POM могут быть описаны логнормальным распределением, которое имеет выраженную положительную асимметрию, среднее и дисперсия для POM составляют соответственно 3.1 и 5.0, для Chl-a – 0.61 и 0.2. Количество первичной продукции здесь в основном находится в диапазоне низких значений, и потенциальные возможности расположенных в данном районе участков культивирования значительно ниже.
Рис. 4.
Гистограмма распределения POM мгC/м3 (а, б, в) и Chl-a мг/м3 (г, д, е) для районов культивирования: I – б. Круглая (а, г), II – центральная часть б. Воевода (б, д), III – б. Мелководная (в, е).

Данные, представленные на рис. 4, наглядно демонстрируют, что условия культивирования по критерию обеспеченности устриц пищей в районах I и II, расположенных во внутренних бухтах, намного лучше, чем в мористой части б. Воевода. В то же время, несмотря на оптимальные концентрации трофических ресурсов во внутренних бухтах, гидрологический режим здесь более жесткий, с резкими перепадами солености. В б. Мелководная соленость во время выхода тайфунов и активного циклогенеза на короткое время (до 5 сут) снижается практически до полного распреснения. Катастрофические осадки, выпадающие на водосбор и акваторию бухты, могут привести к полной гибели урожая. Водосбор б. Круглая намного меньше такового б. Мелководная. Воздействие пресных вод здесь не так резко выражено, условия культивирования более стабильны, соответственно, смертность гидробионтов намного ниже, чем в районе II, расположенном в зоне воздействия паводкового стока р. Русская.
Таким образом, можно сделать вывод, что условия выращивания даже в небольшой по размерам б. Воевода по гидрологическим и биологическим показателям настолько неоднородны, что для каждого объекта и способа культивирования необходимо проводить специальные расчеты и районирование акватории. При садковом культивировании продуктивность устрицы ожидаемо выше, чем при пастбищном, однако продуктивность садкового культивирования гораздо в большей степени зависит от гидрометеорологических условий. Потери урожайности устрицы при садковом выращивании могут быть намного более ощутимыми, чем при выращивании на донных плантациях. В условиях минимального неблагоприятного воздействия пресных вод урожайность в бухтах Круглая и Мелководная практически одинакова как для донного, так и для садкового культивирования.
ЗАКЛЮЧЕНИЕ
В настоящем исследовании на основе результатов гидродинамического и биогеохимического моделирования с использованием модели FARM определены параметры и выполнена оценка пространственной неоднородности условий культивирования устрицы Magallana gigas в б. Воевода. Анализ пространственной динамики абиотических факторов показал, что возможность выращивания устрицы на локальных участках бухты определяется главным образом квазипериодическими изменениями солености. При этом продуктивность культивирования зависит от трофических условий, связанных с производством органического вещества и его транспортом по акватории под влиянием гидродинамических факторов. На основе данных моделирования показано, что по направлению к открытой части бухты количество пищевых ресурсов для выращивания гидробионтов уменьшается, в то же время гидрологические условия, прежде всего режим солености, стабилизируются. Таким образом, рациональный выбор места и условий является ключевым для успешного культивирования устрицы.
Список литературы
Барабанщиков Ю.А., Тищенко П.Я., Семкин П.Ю. и др. Сезонные гидролого-гидрохимические исследования бухты Воевода (Амурский залив, Японское море) // Изв. ТИНРО. 2015. Т. 180. С. 161–178.
Бугаец А.Н., Катрасов С.В., Жариков В.В. и др. Вероятностно-статистическая оценка потенциальной продуктивности марикультуры (на примере бухты Воевода, юг Приморского края) // Докл. Рос. акад. наук. Науки о Земле. 2022. Т. 503. № 1. С. 104–107.
Гаврилова Г.С. Приемная емкость аквакультурной зоны залива Петра Великого (Японское море): Автореф. дис. … докт. биол. наук. Владивосток: ТИНРО-Центр. 2012. 37 с.
Гаврилова Г.С., Ким Л.Н. Эффективность культивирования приморского гребешка (Mizuhopecten yessoensis) в Уссурийском заливе (Японское море) // Изв. ТИНРО. 2016. Т. 185. С. 240–250.
Гаврилова Г.С., Кондратьева Е.С. Результаты хозяйственной деятельности и проблемы развития марикультуры залива Посьета (Японское море) в 2000–2015 гг. // Изв. ТИНРО. 2018. Т. 195. С. 229–243.
Гаврилова Г.С., А.В. Кучерявенко. Продуктивность плантаций двустворчатых моллюсков в Приморье. Владивосток: ТИНРО-Центр. 2011. 113 с.
Гайко Л.А. Современные подходы к прогнозированию урожайности гидробионтов в хозяйствах марикультуры с учетом климатических факторов // Науч. тр. Дальрыбвтуза. 2017. Т. 43. № 4. С. 5–11.
Иванов Г.И. Почвообразование на юге Дальнего Востока. М.: Наука. 1976. 200 с.
Катрасов С.В., Бугаец А.Н., Жариков В.В. и др. Определение районов размещения плантаций марикультуры на основе результатов гидродинамического моделирования // Океанология. 2021а. Т. 61. № 3. С. 433–443.
Катрасов С.В., Бугаец А.Н., Жариков В.В. и др. Оценка продуктивности плантаций двустворчатых моллюсков на основе результатов моделирования // Океанология. 2021б. Т. 61. № 5. С. 759–768.
Кучерявенко А.В., Жук А.П. Инструкция по технологии культивирования тихоокеанской устрицы. Владивосток: ТИНРО-Центр. 2011. 27 с.
Bricker S.B., Ferreira J.G., Simas T. An integrated metho-dology for assessment of estuarine trophic status // Ecol. Model. 2003. V. 169. № 1. P. 39–60.
Brigolin D., Dal Maschio G., Rampazzo F. et al. An individual-based population dynamic model for estimating biomass yield and nutrient fluxes through an off-shore mussel (Mytilus galloprovincialis) farm // Estuar. Coast. Shelf Sci. 2009. V. 82. № 3. P. 365–376.
Ferreira J.G., Hawkins A.J.S., Bricker S.B. Management of productivity, environmental effects and profitability of shellfish aquaculture – the Farm Aquaculture Resource Management (FARM) model // Aquaculture. 2007. V. 264. P. 160–174.
Ferreira J.G., Hawkins A.J.S., Monteiro P. et al. Integrated Assessment of Ecosystem-scale Carrying Capacity in Shellfish Growing Areas // Aquaculture. 2008. V. 275. № 1–4. P. 138–151.
Hawkins A.J.S., Bayne B.L. Seasonal variation in the relative utilization of carbon and nitrogen by the mussel Mytilus edulis: budgets, conversion efficiencies and maintenance requirements // Mar. Ecol. Prog. Ser. 1985. V. 25. P. 181–188.
Hawkins A.J.S., Bayne B.L. Physiological processes, and the regulation of production // The Mussel Mytilus: Ecology, Physiology, Genetics and Culture. Amsterdam: Elsevier. 1992. 590 p.
Hawkins A.J.S., Duarte P., Fang J.G. et al. A functional model of responsive suspension-feeding and growth in bivalve shellfish, configured and validated for the scallop Chlamys farreri during culture in China // J. Exp. Mar. Biol. Ecol. 2002. V. 281. P. 13–40.
Hawkins A.J.S., Pascoe P.L., Parry H. A generic model structure for the dynamic simulation of feeding, meta-bolism and growth in suspension-feeding bivalve shellfish (ShellSIM): calibrated and validated for both Mytilus edulis and Crassostrea gigas cultured at contrasting sites throughout Europe // J. Exp. Mar. Biol. Ecol. 2002. V. 281. № 1–2. P. 13–40.
McCausland W.D., Mente E., Pierce, G.J., Theodossiou I. A simulation model of sustainability of coastal communities: aquaculture, fishing, environment and labour markets // Ecol. Model. 2006. V. 193. № 3–4. P. 271–294.
McKindsey C.W., Thetmeyer H., Landry T. et al. Review of recent carrying capacity models for bivalve culture and recommendations for research and management // Aquaculture. 2006. V. 261. № 2. P. 451–462.
Nobre A.M., Ferreira J.G., Newton A. et al. Management of coastal eutrophication: integration of field data, ecosystem-scale simulations and screening models // J. Mar. Syst. 2005. V. 56. № 3/4. P. 375–390.
Nunes J.P., Ferreira J.G., Gazeau F. et al. A model for sustainable management of shellfish polyculture in coastal bays // Aquaculture. 2003. V. 219. № 1–4. P. 257–277.
Rueda J.L., Smaal A.C., Scholten H. A growth model of the cockle (Cerastodermaedule L.) tested in the Oosterschelde estuary (The Netherlands) // J. Sea Res. 2005. V. 54. P. 276–298.
Solidoro C., Pastres R., MelakuCanu D. et al. Modelling the growth of Tapes phillipinarum in Northern Adriatic lagoons // Mar. Ecol. Prog. Ser. 2000. V. 199. P. 137–148.
Дополнительные материалы отсутствуют.