

Биология моря, 2023, T. 49, № 6, стр. 375-385
Структура зоопланктонных сообществ губы Сухое Море (юго-восток Белого моря)
Е. И. Собко 1, *, С. И. Климов 1, Н. В. Неверова 1, О. Ю. Морева 1
1 Федеральный исследовательский центр комплексного изучения Арктики им. академика Н.П. Лаверова УрО РАН
163000 Архангельск, Россия
* E-mail: elfisina@yandex.ru
Поступила в редакцию 04.02.2023
После доработки 16.05.2023
Принята к публикации 08.06.2023
- EDN: FRMNKA
- DOI: 10.31857/S0134347523060086
Аннотация
В работе представлены исследования зоопланктонных сообществ, выполненные в июле−августе 2018 г. в губе Сухое Море (юго-восточная часть Двинского залива Белого моря). Изучены видовой состав, количественные характеристики и пространственное распределение зоопланктона. Зоопланктонные сообщества представлены 34 видами гидробионтов. Доминирующей группой во всем исследуемом районе были копеподы. Зоопланктонные сообщества характеризовались высокими количественными показателями. Существенный вклад в формирование общей численности зооценоза вносили ювенильные стадии копепод. На полной воде численность зоопланктона составляла 37 тыс. экз./м3, биомасса – 0.6 г/м3(сырой массы), на малой – соответственно 210 тыс. экз./м3 и 1.7 г/м3(сырой массы). Структура, обилие и пространственное распределение зоопланктона в губе обусловлены морфометрическими особенностями водоема (мелководностью), влиянием комплекса факторов внешней среды (температуры и солености), а также приливо-отливными явлениями.
В последние годы значительно возрос интерес к изучению прибрежных морских экосистем. Прибрежные зоны морей, в том числе и устьевые области рек, являются пограничными районами между наземными и морскими экосистемами, что делает их уникальными и в то же время уязвимыми (Бузолева и др., 2014). В них происходит сложное взаимодействие природных процессов и антропогенного воздействия, определяющих состояние и функционирование прибрежных экосистем. Морские прибрежные местообитания занимают особое место среди природных экосистем и играют важную роль в поддержании экологического равновесия и сохранении видового разнообразия морей и океанов. Большинство прибрежных экосистем характеризуются высокой продуктивностью и служат идеальным местом для развития молоди многих морских организмов (Сафьянов, 1987; Курилов, 2000; Примаков и др., 2009; Селифонова, 2012).
Белое море – одно из наиболее изученных морей России (Дерюгин, 1928; Перцова, 1962, 1970, 1983; Прыгункова, 1987; Перцова, Кособокова, 2002; Примаков, 2004), однако уровень изученности отдельных его районов не одинаков.
В настоящей работе представлены результаты исследования зоопланктона в губе Сухое Море – мелководном лагунообразном заливе, расположенном в юго-восточной части Двинского залива. Гидрографические и гидрологические исследования в этом районе имеют многовековую историю (Рейнеке, 1883; Ляхницкий, 1917; Мискевич и др., 2018), но гидробиологические работы в губе Сухое Море до настоящего времени не проводились.
Цели данной работы – изучить видовой состав, количественные характеристики и пространственное распределение зоопланктона, а также влияние факторов среды (солености и температуры) и приливо-отливных циклов на структурную организацию зооценозов в прибрежных водах мелководной губы Сухое Море.
МАТЕРИАЛ И МЕТОДИКА
Характеристика района исследований
Губа Сухое море – вытянутый в направлении с юга на север мелководный лагунообразный залив, площадь которого 106 км2, а общая протяженность – 28.3 км. Губа отделена от Двинского залива Никольской косой, о-вами Голая Кошка и Мудьюгский. С востока залив омывает материковую часть беломорского побережья (Зимний берег), прилегающую к дельте р. Северная Двина. На широте мыса Заячий (ст. 21) губа сужается, разделяясь на северную (ст. 1, 2, 3, 8 и 9) и южную (ст. 11, 4а, 4, 5 и 6) части (рис. 1).
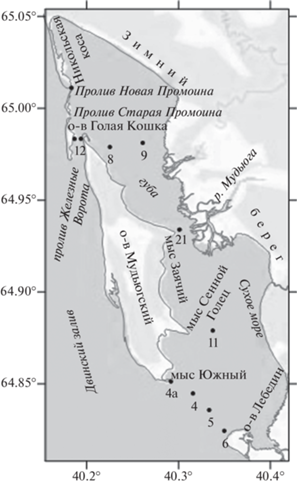
Отличительная особенность губы Сухое Море – ее мелководность. Северная часть залива самая мелководная; на створе мыса Сенной Голец и в прол. Новая Промоина отмечены более глубокие места (до 3−4 м). Глубины, превышающие 5-метровую отметку, находятся в проливе между о-вами Лебедин и Мудьюгский (южная часть залива), а также в прол. Железные Ворота (до 8−9 м) и на прилегающих к нему участках. На малой воде преобладающие глубины в северной части залива во время отлива не превышают 0.5−0.8 м (Гидрология устьевой области…, 1965; Михайлов, 1998; Мискевич и др., 2018).
Гидрологический режим губы Сухое Море подвержен значительной изменчивости из-за мелководного и сложного рельефа района. В северной части залива основной водообмен за приливный цикл происходит через прол. Железные Ворота, в южной – через желоб у о-ва Лебедин. Течения в заливе носят реверсивный (обратный) характер. В течение приливного цикла преобладает поступление воды из южной части залива в северную.
Южная часть залива (ст. 4, 4а, 5, 6 и 11) находится под влиянием пресных вод р. Северная Двина. В зоне смешения морских и речных вод наблюдался четко выраженный градиент солености (по горизонтали и по вертикали) (рис. 2а). Вертикальное распределение солености обусловлено выносом пресных вод у поверхности и компенсационным затоком морских вод у дна. Кратковременная стратификация вод наблюдалась только на полной воде (рис. 2б).
Рис. 2.
Распределение солености на разрезе мыс Южный – о-в Лебедин 24−30 июля 2018 г. на полной (а) и малой (б) воде.
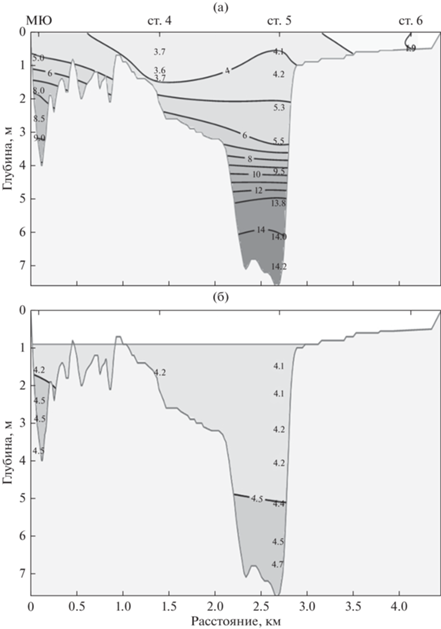
Благодаря хорошему перемешиванию вод и морфометрическим особенностям района, в северной части губы Сухое Море (ст. 1, 2, 3, 8 и 9) градиент солености формируется по горизонтали. Соленость вод в северной части губы в 2−3 раза выше, чем в южной. Средние значения минимумов и максимумов по станциям составляли в южной части залива 3.8 и 8.1 psu, в северной – 11.5 и 18.5 psu.
Исследования зоопланктонных сообществ в губе Сухое Море проводили с 24 июля по 14 августа 2018 г. В районах станций эхолотом Humminbird GPS-Fishfinder 363 произвели предварительные промеры глубин разрезов. Температуру, соленость, кислород и удельную электропроводность воды, а также скорость и направление течений измеряли зондом Seaguard RCM SW фирмы AANDERAA.
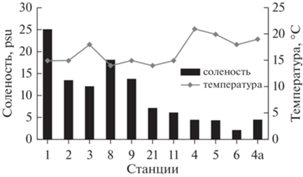
В период исследований температура воды в поверхностном горизонте на станциях разреза мыс Южный – о-в Лебедин была 18−21°C, в северной и центральной частях залива − 14−18°C (рис. 3).
Рис. 3.
Соленость и температура воды в губе Сухое Море по станциям в поверхностном горизонте (полная вода).
Пробы зоопланктона отбирали с катера на 11 станциях (рис. 1) в период малой (МВ) и полной воды (ПВ). Всего собрали и обработали 35 проб. На мелководье пробы отбирали с поверхности ведром (50 л), а затем процеживали через планктонную сеть. На глубоководных станциях отбор проб от дна до поверхности осуществляли планктонной сетью Джеди (диаметр входного отверстия – 18 см, размер ячеи 74 мкм), дополнительно проводили отбор по слоям через 2−3 м. В зоне смешения морских и пресных вод на станциях, где был четко выражен градиент солености (ст. 5 и 4а в южной части залива), пробы отбирали от поверхности до начала галоклина и от галоклина до дна, а также через весь слой от дна до поверхности. Образцы фиксировали 4% нейтрализованным формалином. Пробы зоопланктона отбирали и обрабатывали в соответствии со стандартными гидробиологическими методами (Инструкции по сбору…, 1971; Руководство…, 1977; Руководство…, 1980; Методические рекомендации…, 1982). Для определения видовой структуры зоопланктона использовали соответствующие определители (Бродский, 1950; Определитель…, 2010). В зависимости от отношения гидробионтов к изменению солености и способности адаптироваться к данным условиям выделяли пресноводные, солоноватоводные и морские группы организмов (Хлебович, 1962, 1974).
При анализе проб определяли видовой состав и выделяли доминантные комплексы, а также подсчитывали численность (экз./м3) и биомассу (г/м3) организмов. Сырую биомассу зоопланктона рассчитывали, используя таблицы стандартных весов и номограмм (Перцова, 1967; Численко, 1981).
Поскольку ряды значений короткие и распределение данных не соответствует нормальному закону, для статистического описания применяли квартильные характеристики и непараметрические методы сравнения данных и изучения связи (соответственно U-критерий Манна–Уитни, а также ранговый коэффициент корреляции Спирмена – rS). Для оценки сходства зоопланктонных сообществ использовали кластерный анализ (метод взвешенного попарного среднего – Weighted pair-group average) по нормированным значениям. Данные обрабатывали с использованием Microsoft Excel 2010 и Statistica 10.
Для анализа структуры зоопланктонных сообществ рассчитывали индекс видового разнообразия Шеннона (по численности). При расчете использовали натуральный логарифм (Мэгарран, 1992).
РЕЗУЛЬТАТЫ
Видовой состав и количественные характеристики планктонного сообщества
В планктонном сообществе залива было обнаружено 34 вида беспозвоночных, 15 из которых относятся к пресноводной фауне. Веслоногие ракообразные (Copepoda) были представлены 12 видами, ветвистоусые (Cladocera) – шестью и коловратки (Rotifera) – 10 видами. В пробах были обнаружены представители класса Hydrozoa и морские инфузории семейства Tintinnoidea, а также личинки Cirripedia и Polychaeta.
Копеподы преобладали в сообществах зоопланктона всего исследуемого района. На некоторых участках они составляли 90−100% от общей численности зоопланктона, а относительная биомасса варьировала в пределах 16–99% от суммарных значений. Существенный вклад в формирование общей численности зоопланктона вносили ювенильные стадии копепод (науплии). Их доля составляла 16–88% от численности веслоногих ракообразных и 13–86% от общего количества зоопланктона. Науплии веслоногих ракообразных доминировали почти на всей акватории, за исключением глубоководных станций 1 и 3 в северной части губы, где происходил контакт с водами Двинского залива (табл. 1). Наибольшая численность науплиев Copepoda (24−86% от общего количества зоопланктона) была отмечена на хорошо прогреваемых мелководьях, расположенных в северо-восточной части губы, а также на станциях в центральной и южной частях залива. Доля копеподитных стадий I−IV в зоопланктонных сообществах была представлена незначительно (около 7% от общей численности зоопланктона).
Таблица 1.
Показатели видового разнообразия и количественного обилия зоопланктона в губе Сухое Море в июле–августе 2018 г.
| № станции | N, тыс. экз./м3 | B, г/м3 | n | Индекс Шеннона | Доминанты по численности | Доминанты по биомассе |
|---|---|---|---|---|---|---|
| Северная часть | ||||||
| 1 | 37 | 0.6 | 9 | 2.7 | Microsetella norvegica, Oithona similis, науплиусы Copepoda | Microsetella norvegica, Oithona similis, Calanus glacialis |
| 2 | 28 | 0.6 | 12 | 2.9 | Podon leuckartii, науплиусы Copepoda, науплиусы Cirripedia | Podon leuckartii, Eurytemora affinis, науплиусы Cirripedia |
| 3 | 19 | 0.3 | 9 | 2.0 | Keratella cruciformis, науплиусы Cirripedia, P. leuckartii | P. leuckartii, науплиусы Cirripedia, Calanus glacialis |
| 8 | 160 | 1.2 | 13 | 2.8 | Науплиусы Copepoda, K. cruciformis, M. norvegica | M. norvegica, Acartia longiremis, Podon leuckartii |
| 9 | 136 | 0.7 | 12 | 2.9 | Науплиусы Copepoda, Metridia longa | Metridia longa, M. norvegica |
| Центральная часть | ||||||
| 21 | 339 | 2.9 | 5 | 0.9 | Науплиусы Copepoda, A. longiremis | A. longiremis, M. norvegica |
| Южная часть | ||||||
| 11 | 52 | 0.7 | 6 | 0.8 | M. norvegica, науплиусы Copepoda | M. norvegica, A. longiremis |
| 6 | 115 | 0.2 | 7 | 2.5 | Науплиусы Copepoda, Ceriodaphnia reticulate | Ceriodaphnia reticulate, науплиусы Copepoda |
| 5 | 34 | 0.2 | 18 | 3.8 | Науплиусы Copepoda, Cyclops strenuus, Daphnia galeata | Cyclops strenuus, Eurytemora affinis, Daphnia galeata |
| 4 | 18 | 0.1 | 18 | 3.8 | D. galeata, C. strenuus, науплиусы Copepoda, Polyarthra dolichoptera | C. strenuus, M. norvegica, Asplanchna priodonta |
| 4а | 22 | 0.1 | 8 | 2.9 | Науплиусы Copepoda, Filinia longiseta, C. strenuus, Eurytemorahirundoides | E. affinis, C. strenuus, Eurytemorahirundoides, P. leuckartii |
Примечание. Здесь и в табл. 2: N – численность, B – биомасса, n – количество видов.
Субдоминантами по численности были копеподы Microsetella norvegica (Boeck, 1865), Oithona similis (Claus, 1866), Acartia longiremis (Lilljeborg, 1853) и Metridia longa (Lubbock, 1854), а также науплиусы Cirripedia; на устьевом взморье – пресноводные виды коловраток и кладоцер. По биомассе во всех районах доминировали копеподы.
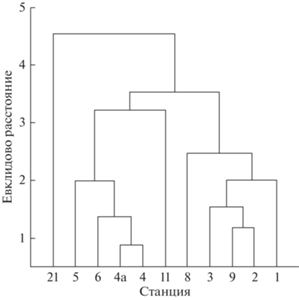
Сравнение сходства зоопланктонных сообществ на разных участках по количеству и видовому составу позволило выделить в акватории залива три части – южную (ст. 5, 6, 4, 4а и 11), центральную (ст. 21) и северную (ст. 1, 2, 3, 8 и 9) (табл. 1, рис. 4).
Рис. 4.
Дендрограмма сходства разных районов губы Сухое Море по численности, биомассе и биоразнообразию зоопланктонных сообществ.
В южной части залива, находящейся под доминирующим влиянием стока пресных вод, формирование морского сообщества затруднено. На этом участке устьевого взморья мы обнаружили пресноводные эвригалинные виды Cladocera и Rotifera (до 44% от общего числа видов), и отметили рост численности солоноватоводных форм, представленных родом Eurytemora. Здесь присутствовало максимальное количество видов, среднее значение индекса Шеннона составляло 3.36. Значения численности и биомассы зоопланктона в южной части залива в период полной воды варьировали в пределах 18–116 тыс. экз./м3 и 0.1–0.8 г/м3 соответственно.
В центральной части губы доминирующий комплекс образовали два вида – A. longiremis и M. norvegica. Среднее значение индекса Шеннона для данного района составляло 0.85. На полной воде здесь зарегистрировали максимальные (338 тыс. экз./м3) значения численности зоопланктона, а его биомасса составляла 2.9 г/м3.
В северной части залива, где соленость вод достигала 12−25 psu, сформировались благоприятные условия для морских видов зоопланктона. В этом районе доминирующий комплекс был представлен морскими и солоноватоводными видами A. longiremis, Calanus glacialis (Jaschnov, 1955), M. norvegica и O. similis (Copepoda), Podon leuckartii (G.O. Sars, 1862) (Cladocera), а также личинками усоногих ракообразных. Кроме того, здесь были отмечены многочисленные морские инфузории семейства Tintinnoidea. Среднее значение индекса Шеннона для этого района составляло 2.65. Численность и биомасса зоопланктона в кутовой части губы (ПВ) изменялись в пределах 19–160 тыс. экз./м3 и 0.3– 1.2 г/м3 соответственно.
Количественные показатели зоопланктона в акватории губы Сухое Море варьировали в широких пределах. Численность и биомасса в период полной воды изменялись в пределах 18–339 тыс. экз./м3 и 0.1–2.9 г/м3 соответственно, при медианных значениях 37 тыс. экз./м3 и 0.6 г/м3 (рис. 5, 6а, 6б). На всех станциях, за исключением ст. 1, 8, 9, 11 и 21, отмечали возрастание численности и биомассы зоопланктона с увеличением глубины. В период малой воды численность и биомасса зоопланктона изменялись в пределах 10–478 тыс. экз./м3 и 0.1–7.5 г/м3 соответственно, при медианах 210 тыс. экз./м3 и 1.7 г/м3 (рис. 5, 6в, 6г).
Рис. 5.
Статистические квартильные характеристики численности (а) и биомассы (б) зоопланктона на полной и малой воде.
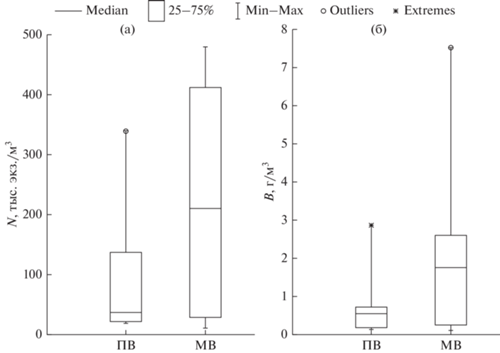
Рис. 6.
Распределение численности (а, в) и биомассы (б, г) зоопланктона в период полной (а, б) и малой (в, г) воды в губе Сухое Море (июль–август 2018 г.).
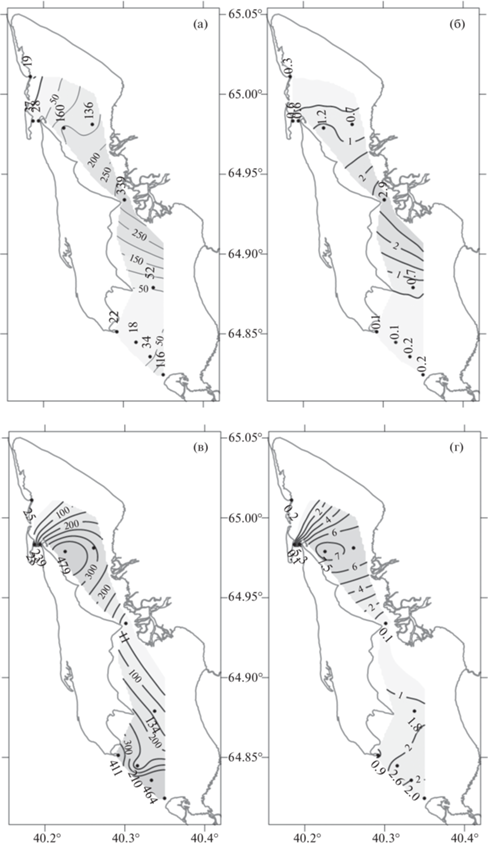
На полной воде зона повышенного обилия зоопланктона образовывалась у мыса Заячий, в центральной части залива (ст. 21). В период малой воды повышенные значения численности и биомассы зоопланктона отмечены в южной (ст. 5 и 4а) и северной частях залива (ст. 8) (рис. 6).
Пресноводные виды преобладали при солености ниже 5 psu (южная оконечность губы). При солености 2–4 psu было отмечено максимальное количество видов и большое разнообразие зоопланктона. С повышением солености эти формы постепенно заменялись эвригалинными солоноватоводными видами. В диапазоне солености от 6 до 8 psu (зона критической солености) наблюдали минимальное количество видов и малое биоразнообразие гидробионтов. При солености выше 12 psu в зоопланктонном сообществе присутствовали морские виды. Доля морских видов в зооценозе составляла 55% от общего числа при солености 25 psu (рис. 7).
Рис. 7.
Распределение экологических групп зоопланктона в губе Сухое Море в зависимости от солености.
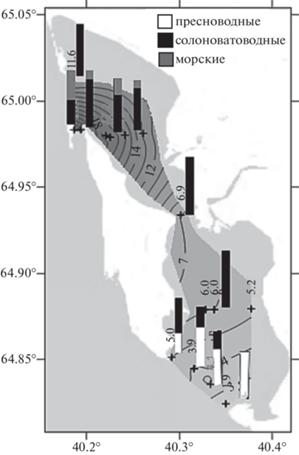
На устьевом взморье у о-ва Лебедин в зоне смешения морских и речных вод (южная оконечность губы, ст. 5 и 4а) наблюдался четко выраженный градиент солености (рис. 2), структурирующий зоопланктонное сообщество на данном участке по вертикали и горизонтали. В поверхностном горизонте преобладали пресноводные виды, выносимые в акваторию залива водами р. Северная Двина, в придонном горизонте – солоноватоводные и морские виды (рис. 7). Для оценки влияния факторов среды на зоопланктонные сообщества использовали непараметрический коэффициент ранговой корреляции Спирмена (табл. 2). Согласно полученным результатам, температура воды (значимая отрицательная связь с численностью и биомассой) и соленость (значимая положительная связь с биомассой) – это основные факторы, влиявшие на количественные показатели зоопланктона. Общее число видов имело умеренную положительную связь с соленостью, значимую при уровне p < 0.2.
ОБСУЖДЕНИЕ
Полученные результаты показали, что в исследованном районе Белого моря сформировался тепловодный бореально-космополитический зоопланктонный комплекс, возникший благодаря мелководности губы Сухое Море и хорошему прогреву поверхностных вод в летний период. Зоопланктонное сообщество губы, представленное 34 видами, в основном неритическими, характеризовалось высокими количественными показателями, по численности и биомассе в нем доминировали копеподы. Существенный вклад в формирование общей численности зоопланктона вносили науплии копепод.
Численность и биомасса планктона в заливе на полной воде составляли 37 тыс. экз./м3 и 0.6 г/м3 (медианные значения). Максимальные значения наблюдались в северо-восточной и центральной частях губы. Численность и биомасса планктона на малой воде достигали 210 тыс. экз./м3 и 1.7 г/м3 (медианные значения). Эти результаты близки к литературным данным, приведенным ранее для других районов Белого моря: Никольская губа – 32.1 тыс. экз./м3 и 0.2 г/м3, губа Кереть – 50.8 тыс. экз./м3 и 0.4 г/м3 и губа Чупа – 13.7 тыс. экз./м3 и 0.2 г/м3 (Примаков, 2004; Примаков и др., 2009).
Закономерности количественного распределения зоопланктона в акватории Сухого Моря связаны с особенностями циркуляции вод. В северной части залива основной водообмен происходит через прол. Железные Ворота. Приливные течения движутся в северо-восточном направлении, далее поток разворачивается на юг. В южной части губы приливные течения поворачивают сначала на восток, а затем на север с частичным затоком вод в северную часть. Поворот течений приводит к образованию в южной части круговоротов – циклоническому с адвекцией соленой воды на приливе и антициклоническому при отливе с захватом пресной воды. На полной воде течения из северной и южной части несут зоопланктон к перешейку у мыса Заячий (ст. 21), образуя зону его повышенного обилия в центральной части залива.
На отливе течения движутся в обратном направлении. В период малой воды повышенные значения численности и биомассы зоопланктона отмечали в южной (ст. 5 и 4а) и северной частях залива (ст. 8). На ст. 8 отлив наступал на 25 мин позже. К тому же этот район расположен в зоне зарастания, и макрофиты выступали в роли барьера (Мосеев и др., 2018), препятствуя оттоку зоопланктона вглубь залива на малой воде, что влияло на количественные значения зоопланктона на этом участке. Повышенные значения численности и биомассы зоопланктона на станциях устьевого взморья могут быть связаны с высокими количественными показателями гидробионтов в придонных слоях на приливе.
Известно, что соленость воды – это лимитирующий фактор, влияющий на зооценозы (Хлебович, 1974, 1986; Сафьянов, 1987). В период наших исследований на устьевом взморье у о-ва Лебедин в зоне смешения морских и речных вод на полной воде градиент солености структурировал зоопланктонное сообщество по вертикали. В северной и центральной частях Сухого Моря, благодаря хорошему перемешиванию вод и морфометрическим особенностям района, градиент солености формировался по горизонтали. Он обусловливал изменение видовой структуры и обилия зоопланктона в заливе. Структура зоопланктонных сообществ в зависимости от солености вод менялась с юга на север. Наблюдалось постепенное замещение пресноводных форм солоноватоводными и морскими. Наибольшее количество видов зарегистрировано на границе соприкосновения разных сообществ.
Губу Сухое Море можно рассматривать как экотон, переходную зону между пресноводным сообществом р. Северная Двина и морским сообществом Двинского залива Белого моря. Большое видовое разнообразие на границе сообществ можно объяснить явлением “краевого эффекта” (Green, 1968; Беклемишев, 1969; Attrill, 2002; Дроздов и др., 2005).
Влияние приливо-отливной гидродинамики на зоопланктонное сообщество губы Сухое Море проявлялось в увеличении плотности зоопланктона в малую воду по сравнению с полной водой и уменьшении численности зоопланктона в период смены вод. Максимальные значения численности и биомассы зоопланктона соответствовали фазе малой воды приливо-отливного цикла. Гидрологический режим обусловливал мозаичность распределения биомассы и численности зоопланктона в заливе. Высокие значения количественных характеристик зоопланктона на полной воде отмечали в центральной части залива, что, вероятно, связано с наличием зарослей тростника обыкновенного Phragmites australis (см. Мосеев и др., 2018). Максимальные значения численности и биомассы зоопланктона на малой воде были отмечены в южной части Сухого Моря на устьевом взморье.
Таким образом, структура, обилие и пространственное распределение зоопланктона в губе Сухое Море обусловлены влиянием сложного комплекса факторов внешней среды – температурой, соленостью, динамикой приливо-отливных явлений, а также мелководностью водоема.
Список литературы
Беклемишев К.В. Экология и биогеография пелагиали. М.: Наука. 1969. 291 с.
Бродский К.А. Веслоногие рачки (Calanoida) дальневосточных морей СССР и Полярного бассейна. М.; Л.: Изд-во АН СССР. 1950. 441 с.
Бузолева Л.С., Богатыренко Е.А., Голозубова Ю.С., Ким А.В. Влияние антропогенного загрязнения на качество прибрежных вод рекреационных зон Приморского края // Фундамент. исслед. 2014. № 11. С. 2423–2425.
Гидрология устьевой области Северной Двины. М.: Гидрометеоиздат. 1965. 376 с.
Дерюгин К.М. Фауна Белого моря и условия ее существования // Исследование морей СССР. Л.: Гос. гидрол. ин-т. 1928. Вып. 7–8. 511 с.
Дроздов В.В., Смирнов Н.П., Гасанова Э.Г. Влияние солености воды на видовой состав биоценозов и формирование экотонов в Балтийском море // Уч. записки Росс. гос. ун-та. 2005. № 1. С. 109–136.
Инструкции по сбору и обработке планктона. М.: Изд-во ВНИРО. 1971. 82 с.
Курилов А.В. Инфузории прибрежных биотопов Одесского залива // Экология моря. 2000. Вып. 52. С. 9–11.
Ляхницкий В.Е. Изыскания в устьях р. Северной Двины, произведенные в 1915–1916 гг. для составления проекта аванпорта у г. Архангельска. Атлас чертежей. Тр. Отдела торговых портов. Петроград: Типолитография Г. де-Кельш. 1917. Вып. XLIX.
Методические рекомендации по сбору и обработке материалов при гидробиологических исследованиях на пресноводных водоемах. Зоопланктон и его продукция. Л.: ГосНИОРХ. 1982. 34 с.
Мискевич И.В., Мосеев Д.С., Брызгалов В. В. Комплексные экспедиционные исследования северной части Сухого Моря в Двинском заливе Белого моря. Архангельск: СОЛТЕК. 2018. 74 с.
Михайлов В.Н. Гидрология устьев рек. М.: Изд-во МГУ. 1998. 176 с.
Мосеев Д.С., Кряучунас В.В., Игловский С.А. и др. Состояние экосистемы губы Сухое Море на юго-востоке Двинского залива Белого моря // Экологическая, промышленная и энергетическая безопасность. 2018. С. 832–837.
Мэгарран Э. Экологическое разнообразие и его измерение. М.: Мир. 1992. 184 с.
Определитель зоопланктона и зообентоса пресных вод Европейской России. Зоопланктон. М.: Тов-во науч. изд. КМК. 2010. Т. 1. 495 с.
Перцова Н.М. Состав и динамика биомассы зоопланктона пролива Великая Салма Белого моря // Биология Белого моря. М.: Изд-во МГУ. 1962. Т. 1. С. 35–50.
Перцова Н.М. Средние веса и размеры массовых видов зоопланктона Белого моря // Океанология. 1967. Т. 7. № 2. С. 305–313.
Перцова Н.М., Сахарова М.И. Особенности развития зоопланктона в прибрежных районах Кандалакшского залива (Великая Салма) в 1966−1967 гг. // Биология Белого моря. М.: Изд-во МГУ. 1970 Т. 3. С. 22–33.
Перцова Н.М. Зоопланктон Горла Белого моря и Мезенского залива // Экология и физиология животных и растений Белого моря. М.: Изд-во МГУ, 1983. Т. 6. С. 17–25.
Перцова Н.М., Кособокова К.Н. Межгодовые изменения биомассы и распределения зоопланктона в Кандалакшском заливе Белого моря // Океанология. 2002. Т. 42. № 2. С. 240–248.
Примаков И.М. Структура планктонного сообщества в устьевой части губы Чупа: опыт многомерного анализа // Морские и пресноводные биосистемы севера Карелии. Тр. БиНИИ СПбГУ. Вып. 51. 2004. С 138–152.
Примаков И.М., Иванова Н.А., Ласовецкая О.А., Чернова Е.Н. Исследования морского зоопланктона в Керетской губе // Вестн. СПб гос. ун-та. 2009. Сер. 3. Вып. 3. С. 135–144.
Прыгункова Р.В. Пространственно-временные изменения структуры и количества зоопланктона Кандалакшского залива Белого моря в летний период // Тр. ЗИН АН СССР. 1987. Т. 172. С. 68–82.
Рейнеке М.Ф. Гидрографическое описание Северного берега России. Белое море. СПб.: Мор. тип. Ч. 1. 1883. 514 с.
Руководство по гидробиологическим работам в океанах и морях. Л.: Гидрометеоиздат. 1977. 724 с.
Руководство по методам биологического анализа морской воды и донных отложений. Л.: Гидрометеоиздат. 1980. 192 с.
Сафьянов Г.А. Эстуарии. М.: Мысль. 1987. 189 с.
Селифонова Ж.П. Таксономический состав и сезонная динамика меропланктона прибрежных вод северо-восточного шельфа Черного моря // Биол. моря. 2012. Т. 38. № 1. С. 3–10.
Хлебович В.В. Особенности состава водной фауны в зависимости от солености среды // Журн. общ. биол. 1962. Т. 23. № 2. С. 90–97.
Хлебович В.В. Критическая соленость биологических процессов. Л.: Наука. 1974. 236 с.
Хлебович В.В. К биологической типологии эстуариев Советского Союза // Тр. ЗИН АН СССР. Л.: Наука. 1986. Т. 141. С. 5–16.
Численко Л.Л. Структура фауны и флоры в связи с размерами организмов. М.: Изд-во МГУ. 1981. 208 с.
Attrill M.J., Rundle S.D. Ecotone or ecocline ecological boundaries in estuaries // Estuarine, Coastal Shelf Sci. 2002. V. 55. P. 929–936. https://doi.org/10.1006/ecss.2002.1036
Green J. The biology of estuarine animals. Seattle: Univ. of Washington Press. 1968. 401 p.
Дополнительные материалы отсутствуют.