

Биология внутренних вод, 2019, № 1, стр. 56-64
Влияние усиления интенсивности любительского лова на рыбное население водотока зоны хозяйственного освоения
Э. И. Бознак 1, *, А. Б. Захаров 2, В. Г. Терещенко 3
1 Сыктывкарский государственный университет им. Питирима Сорокина
167001, Республика Коми, Сыктывкар, Октябрьский проспект, д. 55, Россия
2 Институт биологии Коми научного центра Уральского отделения Российской академии наук
167982, Республика Коми, Сыктывкар, ГСП-2, ул. Коммунистическая, д. 28, Россия
3 Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
152742 Ярославская обл., Некоузский р-н, пос. Борок, Россия
* E-mail: boznak06@rambler.ru
Поступила в редакцию 10.02.2017
После доработки 20.10.2017
Принята к публикации 05.09.2017
Аннотация
На примере одного из Тиманских водотоков (р. Вымь), расположенного в зоне влияния Среднетиманского бокситового рудника, показано, что на первом этапе функционирования предприятия существенное увеличение численности рыбаков-любителей привело к снижению разнообразия рыбного населения верхнего течения р. Вымь и уменьшению численности популяций хозяйственно ценных видов рыб, которые из длинноцикловых стали среднецикловыми.
ВВЕДЕНИЕ
Хозяйственное освоение территории Севера России, связанное с разведкой и добычей минеральных ископаемых, неизбежно приводит к увеличению антропогенной нагрузки на водные объекты, вовлекаемые в сферу деятельности человека. На первом этапе освоения месторождений может и не происходить заметных изменений в гидрохимическом режиме водоемов. Однако развитие системы транспортных коммуникаций, крайне неэффективное регулирование любительского рыболовства и слабый контроль над выловом рыбы вызывают существенное возрастание нагрузки рыбаков-любителей на популяции ценных видов рыб. Дальнейшая разработка месторождений часто приводит к прямому нарушению самих водных объектов и обитающих в них сообществ гидробионтов.
Для прогнозирования возможных изменений в рыбном населении осваиваемых территорий Севера России удобным модельным объектом может служить одна из рек Среднего Тимана (р. Вымь). Несмотря на интенсивные лесозаготовки на водосборе реки во второй половине ХХ в. и промышленную разработку Средне-Тиманского бокситового месторождения с 1998 г., основные гидрохимические и гидробиологические показатели отражают довольно стабильное и благополучное состояние экосистемы этого водотока [7]. Заметные нарушения среды обитания гидробионтов отмечены лишь на локальных участках, непосредственно примыкающих к транспортным коммуникациям или объектам рудника. Таким образом, на протяжении длительного времени основные абиотические показатели остаются благоприятными для обитания холодолюбивых оксифильных видов (атлантического лосося, европейского хариуса и обыкновенного сига), доминирующих в рыбном населении верхнего течения р. Вымь, и сопоставимы с таковыми уральских притоков р. Печоры. В настоящее время число участков, арендованных для любительского рыболовства на водосборе водотока, превышает 10 (по сравнению с одной базой отдыха в конце ХХ в).
В данном регионе популяции сига и европейского хариуса достаточно обособлены. Хариус совершает сезонные перемещения, однако в пределах нерестовых притоков бассейна он формирует репродуктивно изолированные группировки с незначительным обменом мигрантами [18]. Еще более замкнутая группировка – жилая форма сига, образующая самостоятельную устойчивую популяцию [15, 22, 23], что позволяет рассматривать популяции ценных видов рыб и все рыбное население реки в качестве самостоятельной динамической системы.
Цель работы – анализ влияния увеличения пресса любительского рыболовства на состояние рыбной части сообщества и популяций ценных промысловых видов рыб р. Вымь в связи с хозяйственным освоением территории ее водосбора.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Работа выполнена на основе анализа материалов, собранных в ходе мониторинга за состоянием рыбного населения верхнего течения р. Вымь (крупный тиманский водоток, относящийся к бассейну р. Северная Двина), осуществляемого Институтом биологии Коми научного центра Уральского отделения РАН.
Рыб отлавливали в летне-осенний период 2002–2016 гг. ставными жаберными сетями ячеей 20–40 мм. В течение всего периода наблюдений соблюдали единообразие применяемых методов сбора (конструкция и места постановки сетей, режим проверки и т.д.) и обработки материала. Возраст рыб определяли по чешуе, собранной из первого-второго чешуйного ряда над боковой линией, позади от вертикали, проведенной от заднего края спинного плавника по стандартной методике [19]. Для уменьшения погрешности материал обрабатывал один оператор.
Относительную численность рыб рассчитывали как величину улова на промысловое усилие, т.е. количество особей, отловленных в стандартную сеть длиной 30 м в течение 24 ч (экз./(30 м сети · сут)) за 6–8 сут в период с 10 по 30 августа соответствующего года.
При выделении видов, доминирующих в рыбном населении, использовали логарифмическую шкалу. Доминантом считали вид, доля которого по численности в улове была >10%, супердоминантом, если его доля превышала 50%.
Изменения среды обитания и различные воздействия приводят к структурным и функциональным перестройкам в сообществах животных. При этом структурные параметры более чувствительны, поскольку в стрессовом состоянии сообщество, прежде всего, изменяет свою структуру, обеспечивая сохранение внешних функций [2].
Количественное описание видовой структуры рыбного населения было основано на индексе разнообразия контрольных уловов по формуле Шеннона [20]:
где H ' – разнообразие структуры рыбного населения (бит), рi – доля вида в улове по численности, N ‒ число видов.Выбор данного индекса обусловлен предсказуемостью его поведения при изменении структуры сообщества организмов (возрастает при увеличении количества видов и выравненности их обилия) и хорошей обоснованностью с математической точки зрения (наряду с индексами Симпсона, межвидовых встреч, Шелдона и показателя Животовского) [9, 25]. Кроме того, проведенная ранее оценка влияния недоучета малочисленных и неучитываемых в уловах видов, составляющих до половины списка обитающих в водоеме рыб, показала, что происходящая при этом потеря информации приводит к относительной погрешности индекса разнообразия Шеннона ≤15%. Все это позволяет нам по динамике разнообразия сетных уловов судить и о таковой всего рыбного населения.
Для ответа на вопросы находится ли сообщество (или отдельные его компоненты) в равновесном состоянии, и когда произошел переход системы из одного состояния в другое, анализировали “динамический фазовый портрет” значимого параметра системы [5, 13]. Методика построения фазовых портретов реальных биологических систем подробно описана ранее [16, 29]. Преимущество динамического фазового портрета в том, что он позволяет рассматривать не только динамику важного параметра сообщества или популяций, но и скорость его изменения. Такой подход показал свою эффективность при изучении как отдельных популяций [26, 28], так и сообществ гидробионтов [6, 10, 24 и др.].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В настоящее время в водоемах бассейна р. Вымь зарегистрировано 20 видов рыб (без рыбообразных), относящихся к 10 семействам. В верхнем течении реки и ее притоках видовое богатство рыбного населения снижается до 10 видов (табл. 1), причем окунь, ерш и плотва обитают, главным образом, в озерах бассейна. Состав сетных уловов на русловых участках верховий водотока, как правило, представлен лишь пятью видами рыб (табл. 2), из них в доминирующий комплекс входят супердоминант европейский хариус (>55% уловов) и жилая форма сига-пыжьяна (>10%). Щука входила в состав доминантов только в 2007 г. (13.6%), однако за период 2002–2016 гг. наблюдается постепенное снижение ее уловов (с 6.3 до 0.7% общего улова). Регулярно в небольшом количестве встречается атлантический лосось, доля которого, как и налима, обычно ≤2%. Елец, широко распространенный в среднем и нижнем течении р. Вымь, не характерен для верхней части водотока. Впервые здесь он отмечен летом 2006 г., однако в последующие годы не обнаружен. Столь же нерегулярно в уловах присутствует и окунь, предпочитающий участки реки с замедленным течением и развитой высшей водной растительностью.
Таблица 1.
Видовой состав рыбного населения р. Вымь
| Вид | Бассейн реки | Верхнее течение |
|---|---|---|
| Cем. Acipenseridae – осетровые | ||
| Acipenser ruthenus Linnaeus, 1758 – стерлядь | + | – |
| Сем. Cyprinidae – карповые | ||
| Abramis brama (Linnaeus, 1758) – лещ | + | – |
| Alburnus alburnus (Linnaeus, 1758) – уклейка | + | – |
| Carassius carassius (Linnaeus, 1758) – золотой карась | + | – |
| Gobio gobio (Linnaeus, 1758) – пескарь | + | – |
| Leuciscus cephalus (Linnaeus, 1758) – голавль | + | – |
| L. idus (Linnaeus, 1758) – язь | + | – |
| L. leuciscus (Linnaeus, 1758) – елец | + | + |
| Phoxinus phoxinus (Linnaeus, 1758) – обыкновенный гольян | + | + |
| Rutilus rutilus (Linnaeus, 1758) – плотва | + | + |
| Сем. Balitoridaes – балиторовые | ||
| Barbatula barbatula (Linnaeus, 1758) – усатый голец | + | – |
| Сем. Esocidae – щуковидные | ||
| Esox lucius Linnaeus, 1758 – обыкновенная щука | + | + |
| Сем. Coregonidae – сиговые | ||
| Coregonus lavaretus pidschian (Gmelin, 1872) – сиг-пыжьян | + | + |
| Stenodus leucichthys leucichthys (Pallas, 1773) – нельма | + | – |
| Сем. Thymmalida | ||
| Thymallus thymallus (Linnaeus, 1758) – европейский хариус | + | + |
| Сем. Salmonidae – лососевые | ||
| Salmo salar Linnaeus, 1758 – атлантический лосось | + | + |
| Сем. Lotidae – налимовые | ||
| Lota lota (Linnaeus, 1758) – налим | + | + |
| Сем. Cottidae – керчаковые, рогатковые | ||
| Cottus gobio Linnaeus, 1758 – обыкновенный подкаменщик | + | + |
| Сем. Percidae – окуневые | ||
| Perca fluviatilis Linnaeus, 1758 – речной окунь | + | + |
| Gymnocephalus cernuus (Linnaeus, 1758) – обыкновенный ерш | + | – |
Таблица 2.
Видовой состав сетных уловов рыб на русловых участках верховий р. Вымь
| Годы | Обилие вида, % общей численности | ||||||
|---|---|---|---|---|---|---|---|
| Сиг | Хариус | Щука | Налим | Окунь | Семга | Елец* | |
| 2002–2005 | 17.0 | 75.5 | 4.7 | 0.9 | 2.1 | – | – |
| 2006–2009 | 12.9 | 76.8 | 5.7 | 1.0 | – | – | 3.7 |
| 2010–2015 | 15.9 | 82.1 | 1.8 | – | – | 0.6 | – |
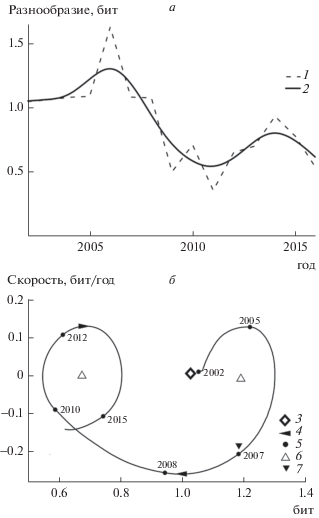
Такие сообщества, в состав которых входит супердоминант и небольшое число остальных видов, характеризуются низким уровнем видового разнообразия. В течение всего периода наблюдений разнообразие контрольных уловов в верхнем течении р. Вымь ≤1.6 бит (рис. 1а). Динамический фазовый портрет наглядно демонстрирует переход рыбного населения из равновесного состояния, соответствующего разнообразию уловов 1.2 бит, в состояние с разнообразием 0.7 бит (рис. 1б). С 2002 по 2006 г. разнообразие рыбного населения возрастало, в дальнейшем этот показатель снижался. В 2006 г. скорость изменения разнообразия стала нулевой, а траектория движения системы в период 2002–2007 гг. приобрела вид эллипса, т.е. рыбная часть сообщества находилась в равновесном состоянии, соответствующем разнообразию уловов 1.2 бит (особая точка типа “центры”). Но уже в 2007 г. в ее функционировании отмечена критическая точка – смена типа траектории движения системы. Она приобретает вид вогнутой дуги, характерной для формирующихся систем [1]. Далее рыбное население перешло в новое равновесное состояние, соответствующее разнообразию уловов 0.7 бит, в котором и находится с 2011 г.
Рис. 1.
Динамика разнообразия (а) и динамический фазовый портрет (б) уловов рыб верхнего течения р. Вымь: 1 – исходные данные; 2 – сглаженные данные; 3 – исходное состояние системы; 4 – направление ее перемещения; 5 – состояние системы в год, отмеченный рядом; 6 – устойчивое состояние системы; 7 – критическая точка в функционировании.
Численность популяции европейского хариуса верхнего течения р. Вымь колебалась с периодом 4 года. Также отмечена тенденция снижения общего уровня численности и амплитуды ее колебаний. Если в 2005–2007 гг. амплитуда колебаний численности была 2 экз./(30 м сети · сут), то в 2009–2016 гг. она не превышала 1 экз./(30 м сети · сут) (рис. 2а). На этом возможности обычного анализа оказываются исчерпанными.
Рис. 2.
Динамика уловов хариуса (а) и их динамический фазовый портрет (б). Обозначения, как на рис. 1.
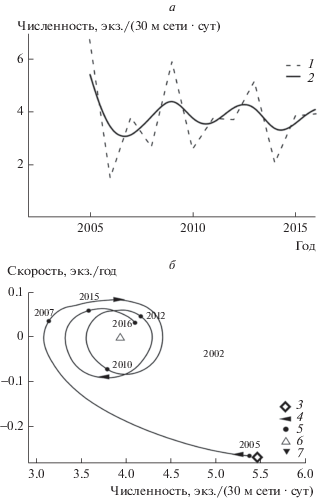
Анализ динамического фазового портрета оказался значительно более информативным. В 2005 г. популяция хариуса находилась в состоянии, соответствующем относительной численности 5.5 экз./(30 м сети · сут). В это время скорость изменения численности была отрицательной, причем максимальной (по модулю) за весь период наблюдений, что характерно для формирующейся системы, переходящей в состояние с меньшим уровнем анализируемого параметра (рис. 2б). К 2007 г. снижение численности замедлилось до 1 экз./(30 м сети · сут), а траектория системы приобрела вид закручивающейся спирали, т.е. популяция хариуса перешла в стационарное состояние, соответствующее относительной численности 4 экз./(30 м сети · сут) (особая точка типа “устойчивый фокус”), в котором она находилась по 2016 г.
Период колебаний численности популяции сига-пыжьяна, как и у европейского хариуса, был 4 года (рис. 3а). Как и у хариуса, наблюдалось снижение численности, однако амплитуда ее колебаний не превышала 0.6 экз./(30 м сети · сут) (рис. 3а).
Рис. 3.
Динамика уловов сига-пыжьяна (а) и их динамический фазовый портрет (б). Обозначения, как на рис. 1.
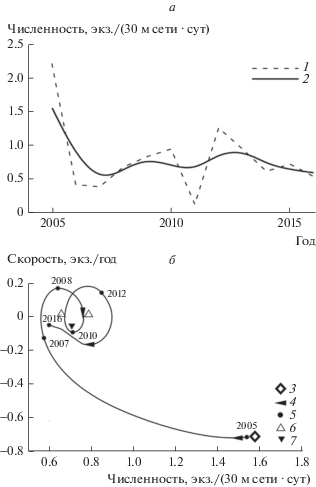
Анализ динамического фазового портрета показал, что популяция сига в 2005 г. находилась в состоянии (5.5 экз./(30 м сети · сут)), характеризующимся максимальной по модулю скоростью изменения численности за весь период наблюдений (рис. 3б). К 2007 г. скорость изменения численности сига снизилась в 5 раз, а траектория движения популяции на динамическом фазовом портрете приняла вид закручивающейся спирали. Следовательно, в этот период происходила стабилизация популяции сига в состоянии, соответствующем относительной численности 0.6 экз./(30 м сети · сут) (особая точка “устойчивый фокус”). Далее (в 2010 г.) в функционировании популяции наблюдалась критическая точка, миновав которую сиг перешел в следующее состояниe равновесия, соответствующее численности 0.7 экз./(30 м сети · сут). Причиной такого перехода могло послужить перераспределение нагрузки любительского рыболовства с жилой формы сига на хариуса.
Таким образом, в 2005–2007 гг. популяции европейского хариуса и сига-пыжьяна верхнего течения р. Вымь переходили в состояние с меньшей численностью. Популяция сига-пыжьяна после 2010 г. находится в состоянии с большим уровнем численности, соответствующим уловам 0.7 экз./(30 м сети · сут).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
На примере рыбного населения одного из водотоков Среднего Тимана, расположенного в зоне влияния Среднетиманского бокситового рудника, показано, что первый этап освоения месторождения бокситов, не повлияв на состояние водотока, привел к существенному увеличению численности рыбаков-любителей. В результате рыбное население верхнего течения р. Вымь перешло в состояние с меньшим по уровню разнообразия уловов. Сходная неспецифическая адаптивная реакция рыбной части сообщества отмечена нами при эвтрофировании озер [17], усилении их теплового [27] и токсического загрязнения [10].
Примененный в данной работе метод позволяет выявить равновесные уровни разнообразия, т.е. зоны устойчивого функционирования сообщества или его части, а также степень отклонения от этих зон. При этом поведение рыбного населения в зонах равновесия, описанное в терминах и формулах метода динамического фазового портрета, хорошо согласуется с общими закономерностями поведения биологических систем. Известно, что при отсутствии значимых воздействий наличие отрицательных обратных связей приводит к стабилизации системы, а выход ее из равновесного состояния свидетельствует об усилении степени воздействия [21].
Трактовка полученных нами результатов согласуется с описанием А.Ф. Алимовым реакции экологических систем на усиление воздействия внешних факторов: “…При усилении воздействия… или при наличии антропогенных нагрузок, к которым данное сообщество не адаптировано, его структура и функции изменяются, и система переходит в другое состояние, с новыми значениями структурных и функциональных характеристик” [2, с. 105–106].
Показано, что популяции доминирующих в водотоке хозяйственно ценных видов рыб при усилении промысловой нагрузки рыбаков-любителей перешли в состояние с меньшим уровнем численности. Анализ динамических фазовых портретов популяций позволяет несколько по-иному взглянуть на изменение их структурно-функциональных характеристик. Во-первых, удается разделить весь анализируемый временной интервал на две части: период перехода системы из одного состояния в другое и период нахождения популяции в равновесном состоянии, что избавляет от необходимости выявления общей тенденции для всего периода наблюдений. Иными словами, изменения численности хариуса и сига-пыжьяна, происходившие после 2007 г., можно рассматривать как периодические колебания вблизи стационарного состояния. Во-вторых, появляется возможность оценить время выхода популяций из предыдущих равновесных состояний.
Применение метода динамического фазового портрета к анализу состояния популяции рыб не сводится только к визуализации происходящих изменений, но и основано на разработках теории динамических систем. Выше отмечалось, что на фазовом портрете динамика системы, далеко отстоящей от состояния равновесия, описывается параболой. Это дает возможность выявить год, когда популяция покинула прежнее состояние. Парабола – симметричная фигура. В 2005 г. скорости изменения численности популяции хариуса, и сига-пыжьяна (по модулю) были максимальными, а в равновесное состояние они пришли к 2007 г. Отсюда можно сделать вывод, что обе популяции покинули свои прежние равновесные состояния в начале 2000-х годов, т.е. после начала работы рудника.
Полученные результаты позволяют корректно описать изменения возрастной структуры популяций, произошедшие за период наблюдений. C помощью имеющихся материалов можно проанализировать возрастную структуру популяций хариуса и сига в период их нахождения в двух последовательных равновесных состояниях: предыдущем (1996–2002 гг.) и в настоящее время (2010–2015 гг.). При переходе в новое состояние с низким уровнем численности в популяциях обоих видов рыб наблюдается сокращение числа возрастных групп, уменьшение максимального возраста и омоложение модального возрастного класса рыб, входящих в контрольные сетные уловы (рис. 4). Добавим, что изменения возрастной структуры (рис. 4) эквивалентны перестройке группировки хариуса с длинноциклового на среднецикловый тип популяции. Подобные изменения структуры промысловых популяций рыб в результате интенсивного вылова хорошо известны [8, 11 и др.].
Рис. 4.
Возрастная структура популяции хариуса (a, б) и сига-пыжьяна (в, г) в равновесных состояниях системы.
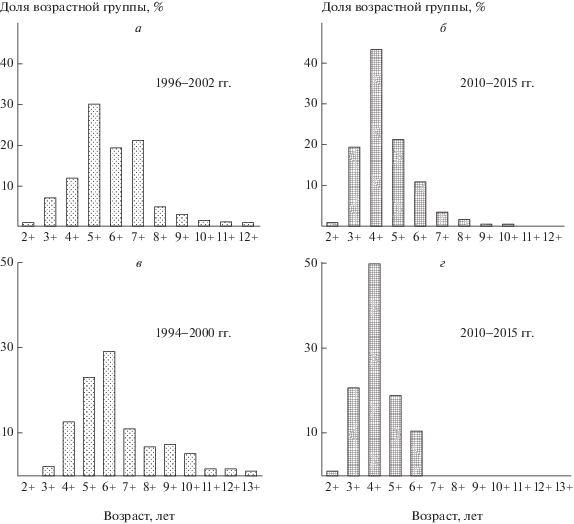
Следует также отметить, что снижение численности видов доминирующего комплекса происходит неравномерно. Если в 2005 г. хариус по численности превосходил сига в 3 раза, то в новом равновесном состоянии их соотношение в уловах различается более чем в 4 раза. В итоге уменьшение выравненности обилий основных промысловых видов рыб (при сохранении их общего числа) привело к снижению уровня разнообразия контрольных уловов.
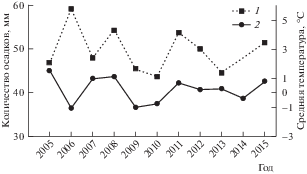
Подобная перестройка рыбного населения может быть вызвана рядом факторов как природного, так и антропогенного характера. Однако заметных нарушений среды обитания рыб в верхнем течении р. Вымь не обнаружено: состояние бентоса остается близким к естественному [7], а рыбы, обитающие в данном районе, характеризуются высоким линейным ростом. Отсутствует достоверная связь (при уровне значимости p < 0.05) между численностью основных промысловых видов рыб, среднегодовой температурой воздуха и количеством осадков (рис. 5). Таким образом, основной причиной наблюдаемых изменений может быть неконтролируемый и, по сути, незаконный лов рыбы. Действительно, в группировках основных промысловых рыб верхнего течения р. Вымь наблюдаются все основные признаки перелова: устойчивое сокращение размерно-массовых параметров, омоложение возрастной структуры контрольных уловов, у хариуса отмечено ускорение линейного роста и полового созревания [7].
Рис. 5.
Многолетняя динамика средней температуры воздуха (1) и количества осадков (2) на территории Среднего Тимана (по данным www.pogodaiklimat.ru/monitor.php?id=23606).
Резкое возрастание антропогенного пресса в последние десятилетия обусловлено, с одной стороны, развитием транспортных коммуникаций на территории Центрального Тимана и повышением технической вооруженности рыболовов. С другой стороны, огромную роль играют правовой нигилизм местного населения и низкая эффективность работы органов, обеспечивающих охрану и рациональное использование природных ресурсов.
К сожалению, детально описать изменение промысловой нагрузки рыбаков-любителей не представляется возможным. Известно, что до 1995 г. верховье р. Вымь оставалось крайне труднодоступным из-за отсутствия автомобильных дорог. Промышленная разработка Средне-Тиманского бокситового месторождения (с 1998 г.) сопровождалась строительством автодороги вдоль трассы (позднее и железной дороги), связавшей территорию месторождения с населенными пунктами. В результате этого значительный участок бассейна верхнего течения р. Вымь оказался доступным для рыбаков, деятельность которых почти не контролировалась. К началу 2000-х гг. в районе устья р. Вежаю построена первая охотничье-рыболовная база. Через 15 лет в бассейне р. Вымь уже >10 участков использовали для организации любительского спортивного рыболовства. Не снижается интенсивность и нелегального вылова. Во время лишь одного рейда в апреле 2014 г. сотрудниками Двинско-Печорского территориального управления Росрыболовства выявлено более 30 нарушений и изъято свыше 200 кг незаконно выловленной рыбы (в основном хариуса и сига).
Таким образом, изменения популяций хариуса и сига верхнего течения р. Вымь, по-видимому, вызваны неконтролируемым рыболовством. В настоящее время наблюдается стабилизация численности популяций этих видов, т.е. интенсивность их изъятия рыбаками-любителями пока компенсируется воспроизводительным потенциалом популяций. Однако сохранение существующего уровня вылова (или его усиление) может привести к резкому падению численности популяций хариуса и сига-пыжьяна, и, как следствие дестабилизации всего рыбного населения данного водотока.
Выводы. Видовой состав и структура рыбного населения верхнего участка р. Вымь сохраняют облик, характерный для тиманских рек полугорного типа. С 2002 по 2016 г. период наблюдений произошел переход рыбного населения водотока в новое равновесное состояние, соответствующее более низкому уровню разнообразия. Популяции европейского хариуса и сига, доминирующих в рыбном населении водотока, перешли в состояние с меньшим уровнем численности. Изменения возрастной структуры за анализируемый период соответствуют перестройкам с длинноциклового на среднецикловый тип популяции. Основная причина перехода рыбной части сообщества водотока в состояние с новыми структурными характеристиками – усиление воздействия рыбаков-любителей.
Список литературы
Айламазян А.К., Стась Е.В. Информатика и теория развития. М.: Наука, 1989. 174 с.
Алимов А.Ф. Элементы теории функционирования водных экосистем. СПб.: Наука, 2001. 147 с.
Атлас пресноводных рыб России. М.: Наука, 2002. 379 с.
Атлас пресноводных рыб России. М.: Наука, 2003. 253 с.
Волькенштейн М.В. Общая биофизика. М.: Наука, 1987. 592 с.
Доровских Г.Н., Терещенко В.Г., Степанов В.Г. Сезонная динамика видовой структуры компонентного сообщества паразитов ерша из реки Вычегда // Паразитология. 2016. Т. 50. № 1. С. 58–68.
Захаров А.Б., Черезова М.И. Ихтиофауна малых водотоков в районе разработки бокситовых месторождений Тимана // Разнообразие и пространственно-экологическая организация животного населения европейского Северо-Востока. Сыктывкар: Коми науч. центр УрО РАН, 2008. С. 54–80.
Никольский Г.В. Теория динамики стада рыб как биологическая основа рациональной эксплуатации и воспроизводства рыбных ресурсов. М.: Пищ. пром-сть, 1974. 447 с.
Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 287 с.
Решетников Ю.С., Терещенко В.Г., Лукин А.А. Динамика рыбной части сообщества в изменяющихся условиях среды обитания (на примере озера Имандра) // Рыб. хоз-во. 2011. № 6. С. 48–52.
Рикер У.Е. Методы оценки и интерпретация биологических показателей популяций рыб. М.: Пищ. пром-сть, 1979. 408 с.
Рыбы в заповедниках России. Т. 1. М.: Товарищество науч. изданий КМК, 2010. 627 с.
Свиржев Ю.М., Логофет Д.О. Устойчивость биологических сообществ. М.: Наука, 1978. 352 с.
Сидоров Г.П. Ихтиофауна // Тиманский кряж. В 2 т. Т. 1: История, география, жизнь: монография. Ухта: Ухт. гос. тех. ун-т, 2008. С. 244–251.
Сидоров Г.П., Решетников Ю.С. Лососеобразные рыбы водоемов европейского Северо-Востока. М.: Товарищество науч. изданий КМК, 2014. 346 с.
Терещенко В.Г., Вербицкий В.Г. Метод фазовых портретов для анализа динамики структуры сообществ гидробионтов // Биология внутр. вод. 1997. № 1. С. 23–31.
Терещенко В.Г., Стерлигова О.П., Павлов В.Т., Ильмаст Н.В. Многолетняя динамика структурных и системных характеристик рыбного населения эвтрофируемого Сямозера // Биология внутр. вод. 2004. № 3. С. 93–102.
Титов С.Ф., Студенов И.И. Генетическая дифференциация европейского хариуса (Thymallus thymallus L.) из рек восточной части ареала (бассейны Баренцева и Белого морей) // Лососевидные рыбы Восточной Фенноскандии. Петрозаводск, 2005. С. 178–190.
Чугунова Н.И. Руководство по изучению возраста и роста рыб. М.: Изд-во АН СССР, 1959. 164 с.
Шеннон К. Работы по теории информации и кибернетике: Пер. с англ. М.: Иностр. литература, 1963. 830 с.
Шмальгаузен И.И. Интеграция биологических систем и их саморегуляция // Кибернетические вопросы биологии. Новосибирск: Наука, 1968. С. 157–183.
Шубин П.Н., Ефимцева Э.А., Челпанова Т.И., Шубин Ю.П. Аллозимная изменчивость лососевидных рыб Европейского Севера. Сыктывкар: Коми науч. центр УрО РАН, 2000. 100 с.
Шубина В.Н. Бентос лососевых рек Урала и Тимана. СПб.: Наука, 2006. 416 с.
Mineeva N.M., Tereshchenko V.G. Application of the phase portrait method for analysis of seasonal dynamics in phytoplankton productivity in a large plain reservoir // Inland Water Biol., 2013, V. 6, № 1, pp. 70–79. doi 10.1134/S1995082913010112
Routledge R.D. Diversity indices: which ones are admissible? // J. Theor. Biol. 1979. V. 76. № 4. P. 503−515.
Tereschenko V.G., Buzevich I.Yu., Khrystenko D.S., Tereschenko L.I. Specific growth rate of kilka Clupeonella cultriventris (Nordmann, 1840) populations in Dneprodzerzhinsk and Kremenchug Reservoirs at different phases of its naturalization // Inland Water Biol. 2015. V. 8. № 3. P. 301–308. doi 10.1134/S1995082915030141
Tereshchenko V.G., Kapusta A., Wilkonska H., Strelnikova A.P. Long-term changes in 0+ fish assemblages in the littoral zone of heated lakes. 1. Using phase diagrams to evaluated changes in ichthyofauna // Archives of Polish Fisheries. 2007. V. 15. Fasc. 4. P. 415–430.
Tereshchenko V.G., Khrystenko D.S., Kotovska G.O., Tereshchenko L.I. Characteristics of stone moroko population dynamics at different stages of naturalization in lake- and steam-type Dnieper Reservoirs // Rus. J. Ecol. 2016. V. 47. № 7. P. 364–370.
Verbitsky V.B., Tereshchenko V.G. Structural phase diagrams of animal communities in assessment freshwater ecosystem conditions // Hydrobiologia. 1996. V. 322. P. 277–282.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод