

Биология внутренних вод, 2019, № 4-1, стр. 3-13
Флора Bacillariophyta планктона тундровых экосистем в зоне освоения газоконденсатного месторождения (п-ов Ямал)
С. И. Генкал 1, *, М. И. Ярушина 2
1 Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
152742 Ярославская обл., Некоузский р-н, пос. Борок, Россия
2 Институт экологии растений и животных Уральского отделения Российской академии наук
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: genkal@ibiw.yaroslavl.ru
Поступила в редакцию 24.11.2017
После доработки 16.02.2018
Принята к публикации 29.05.2018
Аннотация
Изучение фитопланктона водотоков и водоемов из бассейна р. Мордыяха (п-ов Ямал) с помощью сканирующей электронной микроскопии позволило расширить таксономический спектр Bacillariophyta – выявлен 281 таксон видового и внутривидового рангов диатомовых водорослей из 63 родов. Зафиксировано 14 видов и разновидностей, новых для флоры России, 30 форм из 14 родов определены только до рода. Максимальное таксономическое разнообразие отмечено в оз. Пэбтавыто (116) и протоке между двумя озерами (125). Наиболее распространены в исследованных водных объектах Diatoma tenuis, Encyonema silesiacum, Fragilaria vaucheriae, Tabellaria flocculosa и Ulnaria ulna.
ВВЕДЕНИЕ
В последние десятилетия в разных регионах Российской Арктики резко возросли и продолжают увеличиваться антропогенные нагрузки на водные экосистемы. Полуостров Ямал – один из важнейших стратегических нефтяных и газоносных районов России. Промышленное освоение месторождений полуострова и прилегающих акваторий имеет принципиальное значение для обеспечения роста российской добычи нефти и газа. Именно с этим регионом связывают дальнейшее экономическое развитие страны.
В условиях повсеместных многолетних мерзлых пород интенсивное освоение природных ресурсов Крайнего Севера Западной Сибири приводит к негативным последствиям легко уязвимых арктических экосистем, что обусловливает актуальность всестороннего изучения их биоразнообразия и продуктивности.
Основное воздействие обустройства месторождений на водные экосистемы оказывается в период строительства. При сооружении траншей для переходов через водотоки образуются высокая мутность воды и наносы грунта, что приводит к гибели или угнетению гидробионтов, ухудшению условий воспроизводства.
Прямое и косвенное разрушительное воздействие на русла и пойменные водоемы рек п-ва Ямал оказало обустройство Бованенковского газоконденсатного месторождения (БГКМ). В бассейне р. Мордыяхa до начала 1990-х гг. при освоении БГКМ специфического влияния на водные экосистемы не обнаруживали. К середине 90-х гг. пойменные участки, особенно вблизи р. Сёяхa, в результате строительства стали интенсивно изменяться. Наиболее значимые изменения коснулись озер, часть которых поменяла свои границы и площадь [1]. Ряд озер и проток оказались засыпанными или отрезанными от реки. В 2000-е гг. усиливались специфические виды воздействия, неизбежные при обустройстве месторождения. Так, изменения гидрологического и гидрохимического режимов стали определяющими при формировании рыбных запасов р. Пелхатосё [3]. В пойменном участке междуречья р. Морыяхa и р. Сёяхa произошло исчезновение нескольких водоемов из-за отсыпки территории, превращения пойменных озер в хасыреи. Река Пелхатосё стала терять рыбохозяйственное значение. Большая часть поймы междуречья р. Мордыяхa и р. Сёяхa уже его утратила из-за размещения там объектов БГКМ. Причем альтернативы нет, так как только в пойме возможно обеспечение безопасности объектов. По-видимому, рыбохозяйственное значение этой части поймы будет утрачено.
С 1989 по 2009 гг. коллективом сотрудников лаборатории экологии рыб Института экологии растений и животных Уральского отделения РАН проведен экологический мониторинг за состоянием биоценозов водных экосистем на территории месторождений п-ова Ямал, в том числе в бассейнах рек Мордыяха и Сёяха в районах среднего и нижнего течений и на озерах их поймы в районах первоочередного освоения БГКМ [3, 7].
Первое упоминание о водорослях в бассейне р. Мордыяха дано в работе Н.В. Воронкова [4]. По материалам 1908 г., автором обнаружено семь родов водорослей из пяти семейств в некоторых безымянных озерах, расположенных вблизи слияния рек Сёяха и Мордыяха.
В 1989 и 1990 гг. в период с июня по сентябрь впервые было проведено полное флористическое обследование рек Мордыяха, Сёяха (Мутная), Юреяха и восьми небольших озер в их бассейне на территории БГКМ [9–11]. С использованием световой микроскопии был выявлен видовой состав водорослей планктона, была изучена сезонная динамика численности и биомассы фитопланктона водоемов различного типа. По материалам исследований был составлен первый сводный список водорослей [10]. Флора водорослей планктона отличалась обилием видов и включала 219 видовых и внутривидовых таксонов, относящихся к семи отделам. Наибольшим разнообразием во всех типах водоемов, и в альгофлоре в целом, характеризовались диатомовые водоросли. Всего в планктоне обследованных водных экосистем выявлено 106 видовых и внутривидовых таксонов диатомей, но с учетом современных номенклатурных преобразований – только 90 видов разновидностей и форм из пяти порядков 15 семейств и 34 родов. Из них только 7 таксонов составляли центрические водоросли. Наибольшим таксономическим разнообразием отличались семейства Naviculaceae (18) и Fragilariaceae (12). Видовая насыщенность остальных семейств не превышала 10. Среди родов ведущее положение занимали Gomphonema и Nitzschia (по 8 видов).
В 2005 и 2009 гг. мониторинговые исследования Института экологии растений и животных Уральского отделения РАН были продолжены в реках Мордыяха, Сёяха, Пензетарка, Ханголоваяха, в безымянной протоке и семи озерах в их пойме (рис. 1). По неопубликованным данным М.И. Ярушиной, в период летней межени этих лет зарегистрировано 112 видов (121 видовой и внутривидовой таксон) диатомовых водорослей. Ведущее положение по видовому обилию сохранили семейства Naviculaceae (33) и Fragilariaceae (19). Высокие позиции заняли семейства Achnanthaceae (11) и Cymbellaceae (10). Заметно изменилось таксономическое богатство родов Pinnularia (10), Fragilaria (9) и Eunotia (9).
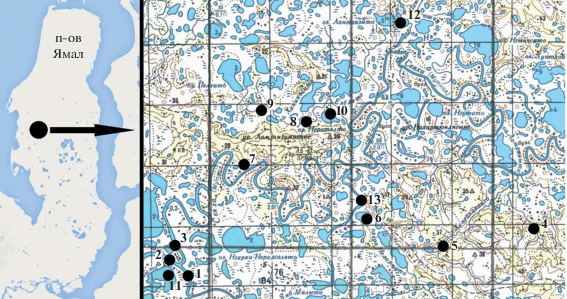
Рис. 1.
Карта-схема отбора альгологических проб в бассейне р. Мордыяха. Исследованные водные объекты ⚫: 1 – р. Мордыяха, выше устья р. Сёяха; 2 – р. Мордыяха, ниже устья р. Сёяха; 3 – р. Сёяха, устье; 4 – р. Пензетарка; 5 – р. Ханголоваяха; 6 – озеро без названия (у свалки); 7 – озеро без названия (полигон 8); 8 – озеро без названия (первое); 9 – озеро без названия; 10 – озеро Неротэлто; 11 – озеро Пэбтавыто; 12 – озеро без названия (верховье р. Пиметосё); 13 – протока без названия между двух безымянных озер.
В последние годы уделяется большое внимание исследованиям по изучению биоразнообразия диатомовых водорослей водных экосистем п-ова Ямал с использованием методов электронной микроскопии [5, 6].
Цель работы – изучение видового состава диатомовых водорослей планктона в водных объектах различного типа в бассейне р. Мордыяха (находящихся в прямой или косвенной зависимости от обустройства и добычи минерально-сырьевых ресурсов) с использованием методов электронной микроскопии.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материалом для электронно-микроскопических исследований диатомей послужили пробы фитопланктона, отобранные в июле 2005 г. и августе 2009 г. в четырех реках, протоке и семи озерах, расположенных в их пойме (рис. 1).
Для диатомового анализа в озерах Пэбтавыто и Неротэлто использовали интегральные пробы, которые содержали материал, собранный в данном водоеме за оба года.
Освобождение створок диатомей от органических веществ проводили методом холодного сжигания [2]. Препараты исследовали в сканирующем электронном микроскопе JSM-25S в Центре коллективного пользования электронной микроскопии Института биологии внутренних вод им. И.Д. Папанина РАН.
При идентификации водорослей использовали современные систематические сводки [11–26].
Река Мордыяха берет начало из оз. Ямбуто. При впадении в Карское море образует сложную дельту и на большом протяжении (60–70 км) подвержена приливно-отливным и сгонно-нагонным явлениям. Подъем уровней воды в половодье достигает абсолютных отметок 5 м, происходит залитие поймы. Длина реки 300 км, площадь бассейна 8530 км2. На территории месторождения находится участок реки в 52–87 км от ее устья. Самый большой приток – р. Сёяха, остальные 43 притока длиной <100 км (табл. 1).
Таблица 1.
Некоторые характеристики водных объектов на территории месторождения БГКМ
| Номер объекта | Название водного объекта | Координаты | Длина, км | Ширина, км |
| р. Мордыяха | 300 | |||
| 1 | выше устья р. Сёяха | 70°19′00″ c.ш., 68°13′00″ в.д. | 0.07–0.20 | |
| 2 | ниже устья р. Сёяха | 70°19′33″ c.ш., 68°10′54″ в.д. | – | 0.15–0.20 |
| 3 | р. Сёяха устье | 70°19′58″ c.ш., 68°11′45″ в.д. | 229 | 0.005–0.025 |
| Притоки р. Сёяха | ||||
| 4 | р. Пензетарка | 70°20′34″ c.ш., 68°49′48″ в.д. | 12 | 0.005–0.010 |
| 5 | р. Ханголоваяха | 70°20′07″ c.ш., 68°39′57″ в.д. | 15 | 0.003–0.012 |
| Озера безымянные (б/н) | ||||
| 6 | бассейн р. Ханголоваяха (у свалки) | 70°21′12″ c.ш., 68°31′34″ в.д. | 1.10 | 0.903 |
| 7 | правый берег р. Сёяхa (полигон 8) | 70°22′58″ c.ш., 68°18′34″ в.д. | 0.311 | 0.144 |
| 8 | левый берег р. Пелхатосё (первое) | 70°24′25″ c.ш., 68°25′00″ в.д. | 0.364 | 0.351 |
| 9 | правый берег р. Пелхатосё | 70°25′00″ c.ш., 68°20′24″ в.д. | 0.438 | 0.300 |
| 10 | оз. Неротэлто на правом берегу р. Пелхатосё | 70°24′36″ c.ш., 68°27′40″ в.д. | 1.60 | 0.756 |
| 11 | оз. Пэбтавыто, слияние рек Сёяхa и Мордыяха | 70°18′58″ c.ш., 68°10′54″ в.д. | 1.89 | 1.73 |
| 12 | оз. б/н, верховье р. Пиметосё | 70°28′02″ c.ш., 68°35′39″ в.д. | 0.436 | 0.283 |
| 13 | Протока б/н между двух озер в бассейне р. Ханголоваяха | 70°21′37″ c.ш., 68°31′17″ в.д. | 1.0 | 0.005–0.007 |
Примечание. Номера безымянных озер 6–9, 12 как на рис. 1.
Река Сёяха (Мутная) берет начало из оз. Нейто и впадает в р. Мордыяха с правого берега на 62 км от ее устья. Длина реки 229 км, площадь водосбора 3550 км2. Нижний участок реки длиной 80 км протекает по территории месторождения.
Реки Перзетарка и Ханголоваяха – малые левобережные притоки р. Сёяха (табл. 1).
Пойма в районах нижних течений рек с множеством озер. Озера термокарстового происхождения, образовавшиеся в результате протаивания мерзлотных грунтов и ледовых линз. Площади наиболее крупных обследованных озер (Пэбтавыто и Неротэлто) ≤3.5 км2. Все озера мелководны, их глубины 1.3–6 м (табл. 1). Вследствие сильных ветров и небольших глубин, температурное и газовое перемешивание воды интенсивное. Грунт дна больших озер плотный. Фрагменты отмерших макрофитов волнобоем выбрасываются за пределы уреза воды. Прозрачность воды в реках и озерах низкая – от 0.05 до 0.5 м. Питание рек и озер – атмосферное, сток – поверхностный, коэффициент стока – 0.8. По гидрохимическому составу вода в реках и озерах пресная, слабоминерализованная. По нашим данным, сумма ионов в реках Сёяха и Мордыяха 50.5–76.7 мг/л, но в устье р. Мордыяха может достигать 6.1 г/л. Активная реакция среды в реках и озерах слабокислая – 6–6.6.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Выявлен 281 таксон диатомовых водорослей из 63 родов: Achnanthes distincta Messikommer (8, здесь и далее (даны номера водных объектов согласно табл. 1 и рис. 1), A. biasolettiana Grunow (11), A. ingratiformis Lange-Bertalot (13), A. polaris Oestrup (10), Achnanthidium bioretii (Germain) Monnuer, Lange-Bertalot et Ector (13), A. daonense (Lange- Bertalot) Lange-Bertalot, Monnuer et Ector (8), A. helveticum (Hustedt) Monnuer, Lange-Bertalot et Ector (8, 10, 13), A. kranzii (Lange-Bertalot) Round et Bukhtiyarova (13), A. minutissimum (Kützing) Czarnecki (11), A. jackii Rabenhorst (8), A. subatomoides (Hustedt) Monnuer, Lange-Bertalot et Ector (7, 13), Amphora copulata (Kützing) Schoemann et Archibald (7), A. cf. neglectiformis Levkov et Edlund (13), A. sp. 1 (7), A. sp. 2 (12), A. sp. 3 (13), Asterionella formosa Hassal (6–8, 10, 13), Aulacoseira ambigua (Grunow) Simonsen (7), A. islandica (O. Müller) Simonsen (2, 7, 8, 10, 11), A. sp. (11), A. subarctica (O. Müller) Haworth (1, 3, 5, 8–10, 12), Boreozonacola hustedtii Lange-Bertalot, Kulikovskiy et Witkowski (7, 8, 11), Caloneis bacillum (Grunow) Cleve (7, 11), C. silicula (Ehrenberg) Cleve (7, 8, 11, 13), Cavinula cocconeiformis (Gregory) D.G. Mann et Stickle (11, 13), C. pseudoscutiformis (Hustedt) D.G. Mann et Stickle (7, 13), C. sp. (11), Chamaepinnularia circumborealis Lange-Bertalot (4), Ch. gandrupii (Petersen) Lange-Bertalot et Krammer (4, 5, 7, 8, 11), Ch. krookii (Grunow) Lange-Bertalot et Krammer (8), Cocconeis placentula var. lineata (Ehrenberg) Van Heurck (11), Cosmioneis cf. pusilla (W. Smith) D.G. Mann et Stickle (12), Craticula cuspidata (Kützing) D.G. Mann (11), Ctenophora pulchella (Ralfs ex Kützing) D.M. Williams et Round (6), Cyclostephanos dubius (Fricke) Round (5, 10), Cyclotella meneghiniana Kützing (6, 11, 13), C. ocellata Pantoscek emend. Genkal et Popovskaya (12), Cymatopleura elliptica var. hibernica (W. Smith) Van Heurck (6), Cymbella lanceolata (Agardh) Agardh (13), C. proxima Reimer (11), C. sp. (11), C. subcistula Krammer (11), Cymbopleura naviculiformis var. laticapitata Krammer (8, 12), C. peranglica Krammer (7, 8, 11, 13), C. subcuspidata (Krammer) Krammer (11, 13), Diatoma ehrenbergii Kützing (5, 8, 10), D. moniliformis Kützing (13), D. tenuis Agardh (1–3, 7, 8, 10, 12, 13), D. vulgaris Bory (4, 7–9, 13), Diploneis elliptica (Kützing) Cleve (11, 13), D. marginestriata Hustedt (7, 8, 13), D. sp. 1 (6), D. sp. 2 (13), Discostella pseudostelligera (Hustedt) Houk et Klee (10, 13), Encyonema elginense (Krammer) D.G. Mann (11), E. hebridiforme Krammer (11), E. hophense Krammer (11), E. incurvatum Krammer (11), E. cf. kamtschaticum Krammer (11), E. latens (Krasske) D.G. Mann (8, 13), E. minutum (Hilse) D.G. Mann (11), E. obscurum var. alpina Krammer (8), E. paucistriatum (Cleve-Euler) D.G. Mann (8, 12, 13), E. perelginense Krammer (11), E. silesiacum (Bleisch) D.G. Mann (5–8, 10–13), E. sp. 1 (11), E. sp. 2 (8), E. cf. stigmoideum Krammer (11), Encyonopsis aequalis (W. Smith) Krammer (13), Entomoneis ornata (Bailey) Reimer (11–13), Epithemia adnata (Kützing) Brébisson (11), Eucocconeis laevis (Oestrup) Lange-Bertalot (10), Eunotia bidens Ehrenberg (11), E. bilunaris (Ehrenberg) Schaarschmidt (5, 6, 8, 10, 11, 13), E. ewa Lange-Bertalot et Witkowski (11), E. exsecta (Cleve-Euler) Nörpel-Schempp et Lange-Bertalot (13), E. fennica (Hustedt) Lange-Bertalot (8, 13), E. incisadistans Lange-Bertalot et E. Sienkiewicz (7, 11, 13), E. intermedia (Krasske) Nörpel-Schempp et Lange-Bertalot (8, 11, 13), E. julma Lange-Bertalot (12), E. michaelis Metzeltin, Witkowski et Lange-Bertalot (7, 8, 11, 12), E. minor (Kützing) Grunow (6, 8, 11, 13), E. neocompacta var. vixcompacta Lange-Bertalot (12), E. paludosa Grunow (11), E. pectinalis (Kützing) Rabenhorst (12), E. praerupta Ehrenberg (12), E. pseudogroenlandica Lange-Bertalot et Tagliaventi (8), E. rhomboidea Hustedt (11), E. scandiorussica Kylikovskiy, Lange-Bertalot, Genkal et Witkowski (5, 8, 11–13), E. septentrionalis Oestrup (8, 13), Fallacia losevae Lange-Bertalot, Genkal et Vekhov (8, 10, 11, 13), Fragilaria austriaca (Grunow) Lange-Bertalot (5, 8, 13), F. binodis Ehrenberg (13), F. capucina Desmazieres (13), Fragilaria construens (Ehrenberg) Grunow (7, 13), F. cyclopum (Britschy) Lange-Bertalot (8, 13), F. delicatissima (W. Smith) Lange-Bertalot (5, 13), F. exigua Grunow (11, 13), F. famelica (Kützing) Lange-Bertalot (6, 8, 11, 13), F. gracilis Oestrup (5, 8, 11, 12), F. mesolepta Rabenhorst (11), F. radians (Kützing) Lange-Bertalot (8, 12), F. sp. (11), Fragilaria tenera (W. Smith) Lange-Bertalot (5, 8), F. vaucheriae (Kützing) Petersen (2, 3, 5, 7–12, 13), Fragilariforma quadrata (Hustedt) Kharitonov (8), F. virescens (Ralfs) D.M. Williams et Round (8, 12), Frustulia lange-bertalottii Metzeltin (7), F. vulgaris (Thwaites) De Toni (5), Geissleria similis (Krasske) Lange-Bertalot et Metzeltin (1, 6, 10, 11, 13), Genkalia digituloides (Lange-Bertalot) Lange-Bertalot et Kulikovskiy (11, 13), Gomphonema acidoclinatum Lange-Bertalot et Reichardt (11), G. angustatum (Kützing) Rabenhorst (11), G. angustum Agardh (5), G. brebissonii Kützing (7, 11, 12), G. clavatulum Reichardt (10, 13), G. extentum Reichardt et Lange-Bertalot (6), G. hebridense Gregory (7, 11, 12), G. interpositum Reichardt (8, 11), G. micropus Kützing (6, 8, 13), G. parvulum (Kützing) Kützing (11, 12), G. sarcophagus Gregory (10), G. sp. 1 (10), G. sp. 2 (13), G. sp. 3 (8), G. truncatum Ehrenberg (7, 11, 12), G. utae Lange-Bertalot et Reichardt (13), G. wiltschkorum Lange-Bertalot (8), Gomphoshpenia stoermeri Kociolek et Thomas (8, 10, 13), Gyrosigma spenceri (Quekett) Griffith et Henfrey (7, 11, 13), Hannaea arcus (Ehrenberg) Patrick (10), Hantzschia amphyioxys (Ehrenberg) Grunow (6, 7, 11–13), Hippodonta capitata (Ehrenberg) Lange-Bertalot, Metzeltin et Witkowski (1, 6, 8, 10, 12, 13), H. coxiae Lange-Bertalot (6), H. hungarica (Grunow) Lange-Bertalot, Metzeltin et Witkowski (7), H. pseudopinnularia Lange-Bertalot (11), Humidophila schmassmannii (Hustedt) Buczkό et Wojtal (13), (?) H. sp. (13), Hydropetra balfouriana (Grunow ex Cleve) Krammer et Lange-Bertalot (8), Karayevia suchlandtii (Hustedt) Bukhtiyarova (13), Luticola aequalis Levkov, Metzeltin et Pavlov (7), L. cf. mutica (Kützing) D.G. Mann (5), L. ventricosa (Kützing) D.G. Mann (12), Melosira varians Agardh (12, 13), Navicula antonii Lange-Bertalot (4), N. cf. bahusiensis (Grunow) Grunow (13), N. cincta Ehrenberg (1, 4, 5, 8, 12, 13), N. cryptocephala Kützing (7, 8, 10–13), N. doehleri Lange-Bertalot (10), N. exilis Kützing (4, 10, 11), N. gregaria Donkin (4, 6), N. hanseatica Lange-Bertalot et Stachura (11–13), N. kohlmaieri Lange-Bertalot (4, 6, 11, 12, 13), N. margalithii Lange-Bertalot (7, 11, 13), N. menisculus Schumann (6, 7, 12), N. moskalii Metzeltin, Witkowski et Lange-Bertalot (6), N. oligotraphenta Lange-Bertalot et Hofmann (11, 13), N. platystoma Ehrenberg (7), N. pseudolanceolata Lange-Bertalot (13), N. radiosa Kützing (4, 6, 7, 11–13), N. rhynchotella Lange-Bertalot (5, 7, 8, 11, 12, 13), N. slesvicensis Grunow (4, 11), N. sp. 1 (5), N. sp. 2 (7), N. sp. 3 (11), N. sp. 4 (11), N. sp. 5 (11), N. venerablis Hohn et Hellerman (13), Neidium affine (Ehrenberg) Pfitzer (7, 8, 10, 11, 13), N. ampliatum (Ehrenberg) Krammer (7, 13), N. bisulcatum (Lagerstedt) Cleve (4, 7, 8, 12), N. hitchcockii (Ehrenberg) Cleve (13), N. iridis (Ehrenberg) Cleve (13), N. productum (W. Smith) Cleve (11–13), Nitzschia acidoclinata Lange-Bertalot (8), N. alpina Hustedt emend. Lange-Bertalot (4, 8), N. bacillarieformis Hustedt (7), N. capitellata Hustedt (11, 13), N. clausii Hantzsch (4, 5), N. commutatoides Lange-Bertalot (13), N. dissipata (Kützing) Grunow var. dissipata (5), N. dissipata var. media (Hantzsch) Grunow (11), N. draveillensis Coste et Ricard (13), N. gracilis Hantzsch (6, 7, 11, 13), N. intermedia Hantzsch ex Cleve et Grunow (5, 8, 11, 13), N. lacunarum Hustedt (13), N. linearis var. subtilis (Grunow) Hustedt (11), N. nana Grunow (7, 11), N. palea (Kützing) W. Smith (8, 10, 13), N. paleaeformis Hustedt (13), N. palustris Hustedt (5, 11, 13), N. perminuta (Grunow) Peragallo (13), N. recta Hantzsch ex Rabenhorst (8, 13), N. sigma (Kützing) W. Smith (4), N. sp. 1 (11), N. sp. 2 (13), N. sp. 3 (13), N. strelnikovae Lange-Bertalot, Genkal et Vekhov (5, 10, 12, 13), N. terrestris (Petersen) Hustedt (7), N. tubicola Grunow (7, 8), N. vermicularis (Kützing) Hantzsch (7, 11, 13), Nupela impexiformis (Lange-Bertalot) Lange-Bertalot (13), Parlibellus crucicula (W. Smith) Witkowski, Lange-Bertalot et Metzeltin (6), Pinnularia anglica Krammer (7, 13), P. biceps Gregory (11), P. birnirkiana Patrick et Freese (7), P. borealis var. islandica Krammer (12), P. borealis var. sublinearis Krammer (7), P. borealis var. thuringiaca (Rabenhorst) Krammer (13), P. brebissonii (Kützing) Rabenhorst var. brebissonii (11, 13), P. brebissonii var. acuta Cleve-Euler (7), P. divergens W. Smith (11), P. genkalii Krammer et Lange-Bertalot (7, 11), P. grunowii Krammer (8, 13), P. isostauron (Grunow) Cleve (7), P. lutarae Krammer (7), P. macilenta Ehrenberg (6, 7, 11, 12), P. mesolepta (Ehrenberg) W. Smith (11), P. microstauron (Ehrenberg) Cleve var. microstauron (8, 11), P. microstauron var. nonfasciata Krammer (13), P. nodosa var. percapitata Krammer (6), P. nodosa var. robusta (Foged) Krammer (10, 13), P. nodosiformis Krammer (8, 11), P. cf. notabilis Krammer (12), P. obscuriformis Krammer (7), P. cf. peracuminata Krammer (7), P. cf. polyonca (Brébisson) W. Smith (11), P. sp. 1 (12), P. sp. 2 (13), P. sp. 3 (7), P. sp. 4 (12), P. subcommutata Krammer (7), P. subrhombica var. angusta Krammer (7), P. tirolensis var. julma Krammer et Metzeltin (7), P. undula (Schumann) Krammer (11), P. viridiformis Krammer (7, 11), Placoneis constans (Hustedt) Cox (7, 11, 13), P. elginensis (Gregory) Cox (13), P. cf. pseudanglica (Lange-Bertalot) Cox (11), Planothidium haynaldii (Schaarschmidt) Lange-Bertalot (6, 13), Psammothidium rechtense (Leclerq) Lange-Bertalot (11, 13), P. rossii (Hustedt) Bukhtiyarova et Round (7), P. ventralis (Krasske) Bukhtiyarova et Round (7, 13), Rossithidium petersenii (Hustedt) Round et Bukhtiyarova (7, 8, 11–13), R. pusillum (Grunow) Round et Bukhtiyarova (7, 8, 10–13), (?) R. sp. (11), Sellaphora absoluta (Hustedt) C.E. Wentzel, Ector, Van de Vijver, Compére et D.G. Mann (13), S. bacillum (Ehrenberg) D.G. Mann (6), S. disjuncta (Hustedt) D.G. Mann (8, 11, 13), S. elorantana Lange-Bertalot (5, 6, 8, 11, 13), S. laevissima (Kützing) D.G. Mann (4, 7, 8, 10–13), S. mutatoides Lange-Bertalot et Metzeltin (6, 10, 13), S. parapupula Lange-Bertalot (11, 13), S. pupula (Kützing) Mereschkowsky (7, 8, 10, 11, 13), S. sp. 1 (7), S. sp. 2 (13), S. sp. 3 (7), S. stauroneiodes (Lange-Bertalot) J. Veseda et J.R. Johansen (13), S. vitabunda (Hustedt) D.G. Mann (7, 13), Stauroneis anceps Ehrenberg (7, 8, 11–13), S. gracilis Ehrenberg (11), S. phoenicenteron (Nitzsch) Ehrenberg (7, 11, 13), S. prominula (Grunow) Hustedt (10), S. smithii Grunow (7, 11, 13), S. sp. 1 (13), S. sp. 2 (11), Staurosira lapponica (Grunow) Lange-Bertalot (13), Stephanodiscus hantzschii Grunow (10, 12), S. makarovae Genkal (6), S. minutulus (Kützing) Cleve et Möller (6, 12), S. neoastraea Håkansson et Hickel emend. Carter, Scheffler et Augsten (10), Surirella angusta Kützing (4, 5, 7, 8, 12, 13), S. helvetica Brun (7, 13), S. linearis W. Smith var. linearis (11, 13), S. minuta Brébisson (2, 4, 10–13), Tabellaria flocculosa (Roth) Kützing (2, 4, 5, 6, 8–13), Tryblionella debilis Arnott ex O′Meara (4), T. littoralis (Grunow) D.G. Mann (11), T. salinarum (Grunow) Pelletan (7, 10, 11, 13), Ulnaria acus (Kützing) Aboal (1, 5, 13), U. ulna (Nitzsch) P. Compére (4–6, 8, 10–13).
Ниже приводятся краткие диагнозы, синонимика и распространение с иллюстрациями редких (*) и новых видов для флоры России, включая 30 форм, определенных только до рода.
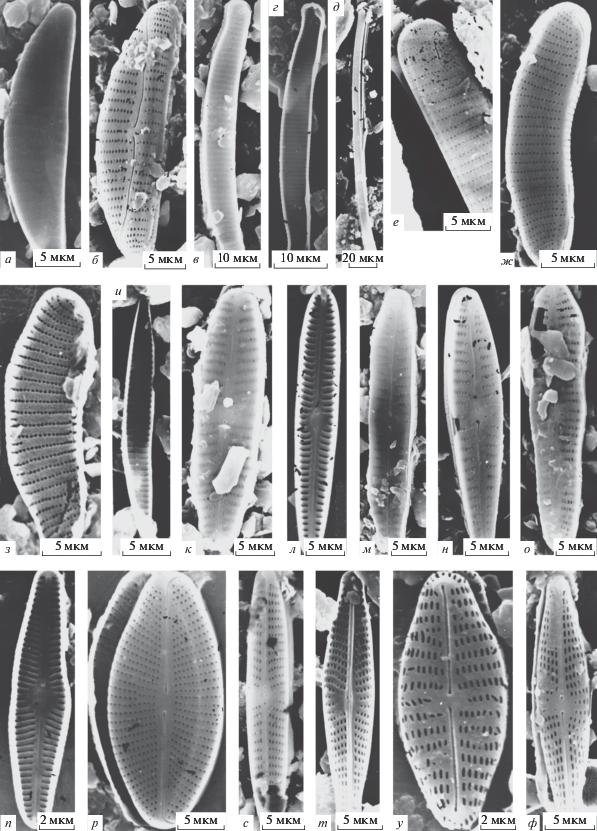
Achnanthes distincta (рис. 2а). (Syn. A. hirta Carter.). Створки длиной 13.6–18.6 мкм, шириной 6.4–7.8 мкм, штрихов 19–20 в 10 мкм. Северное полушарие, олиготрофные воды [20].
Рис. 2.
Электронные микрофотографии створок (СЭМ): a – Achnanthes distincta, б – Amphora cf. neglectiformis, в – A. sp. 1, г – A. sp. 2, д – A. sp. 3, е – Aulacoseira sp., ж – Cavinula sp., з – Cymbella sp., и – Diploneis sp. 1, к – D. sp. 2, л – Encyonema hebridiforme, м – E. hophense, н – E. cf. kamtschaticum, о – E. sp. 1; а, е–л, н, о – створки с наружной поверхности, б, в, м – створки с внутренней поверхности, г, д – панцири.

Amphora cf. neglectiformis (рис. 2б). Створка длиной 18.6 мкм, шириной 3.9 мкм, штрихов 18 в 10 мкм.
A. sp. 1 (рис. 2в). Створка длиной 40 мкм, шириной 13 мкм.
A. sp. 2 (рис. 2г). Створка длиной 33 мкм.
A. sp. 3 (рис. 2д). Створка длиной 21.4 мкм, шириной 7.1 мкм, штрихов 14 в 10 мкм.
Aulacoseira sp. (рис. 2е). Диаметр 20 мкм, высота 25.7 мкм, рядов ареол 7 в 10 мкм, ареол в ряду 5 в 10 мкм.
Cavinula sp. (рис. 2ж). Створка длиной 27.8 мкм, шириной 13.3 мкм, штрихов 15 в 10 мкм.
Cymbella sp. (рис. 2з). Створка длиной 16.4 мкм, шириной 4.1 мкм, штрихов 8 в 10 мкм.
Diploneis sp. 1 (рис. 2и). Створка длиной 23.6 мкм, шириной 9.3 мкм, штрихов 18 в 10 мкм.
D. sp. 2 (рис. 2к). Створка длиной 26.4 мкм, шириной 14.4 мкм, штрихов 11 в 10 мкм.
Encyonema hebridiforme (рис. 2л). Створки длиной 27.8 мкм, шириной 9.2–10 мкм, штрихов 8–20 в 10 мкм, ареол в ряду 22–26 в 10 мкм.
E. hophense (рис. 2м). Створка длиной 22.8 мкм, шириной 7.1 мкм, штрихов 9 в 10 мкм, ареол в ряду 30 в 10 мкм.
*E. cf. kamtschaticum (рис. 2н). Створка длиной 66.6 мкм, шириной 20 мкм, штрихов 6 в 10 мкм, ареол в ряду 15 в 10 мкм.
E. sp. 1 (рис. 2о). Створка длиной 32 мкм, шириной 9.4 мкм, штрихов 8 в 10 мкм, ареол в ряду 24 в 10 мкм.
E. sp. 2 (рис. 3а). Створка длиной 27.8 мкм, шириной 6.4 мкм, штрихов 9 в 10 мкм, ареол в ряду 30 в 10 мкм.
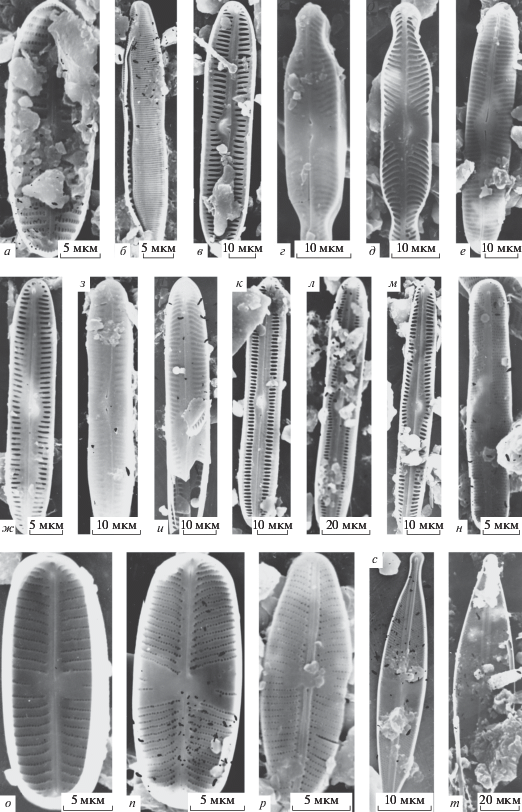
Рис. 3.
Электронные микрофотографии створок (СЭМ): а – Encyonema sp. 2, б – E. cf. stigmoideum, в, г – Eunotia fennica, д, е – E. julma, ж, з – E. michaelis, и – Fragilaria sp., к – Gomphonema sp. 1, л – G. sp. 2, м – G. sp. 3, н – G. utae, о – G. cf. wiltschkorum, п – (?) Humidophila sp., р – Navicula cf. bahusiensis, с – N. sp. 1, т – N. sp. 2, у – N. sp. 3, ф – N. sp. 4; а–в, д–ж, к, м–о, р, с, у, ф – створки с наружной поверхности, г, з, и, л, п, т – створки с внутренней поверхности.
E. cf. stigmoideum (рис. 3б). Створка длиной 32.2 мкм, шириной 8.3 мкм, штрихов 9 в 10 мкм, ареол в ряду 35 в 10 мкм.
Eunotia fennica (рис. 3в–3г). Створки длиной 38.6–42.8 мкм, шириной 4.3–5.0 мкм, штрихов 14 в 10 мкм.
E. julma (рис. 3д–3е). Створка длиной 18.6 мкм, шириной 5.2 мкм, штрихов 14 в 10 мкм. Финляндия [22].
*E. michaelis (рис. 3ж–3з). Створки длиной 22.8–24.4 мкм, шириной 5.7–7.8 мкм, штрихов 12–16 в 10 мкм. Медвежий остров, европейская часть России [22].
Fragilaria sp. (рис. 3и). Створка длиной 27.8 мкм, шириной 2.8 мкм, штрихов 16 в 10 мкм.
Gomphonema sp. 1 (рис. 3к). Створка длиной 34 мкм, шириной 7.8 мкм, штрихов 8 в 10 мкм.
G. sp. 2 (рис. 3л). Створка длиной 20.7 мкм, шириной 3.2 мкм, штрихов 20 в 10 мкм.
G. sp. 3 (рис. 3м). Створка длиной 38.9 мкм, шириной 6.7 мкм, штрихов 8 в 10 мкм.
G. utae (рис. 3н). Створка длиной 33 мкм, шириной 6.7 мкм, штрихов 8 в 10 мкм.
G. cf. wiltschkorum (рис. 3о). Створка длиной 20.7 мкм, шириной 4.3 мкм, штрихов 17 в 10 мкм.
?Humidophila sp. (рис. 3п). Створка длиной 15 мкм, шириной 3.4 мкм, штрихов 28 в 10 мкм.
Navicula cf. bahusiensis (рис. 3р) (Syn. N. minuscula var. bahusiensis Grunow, N. bremeyerii var. rostrata Hustedt, N. koenigii Brockmann). Створки длиной 13.6 мкм, шириной 5.9–6.4 мкм, штрихов 22–23 в 10 мкм. Европа [17].
N. sp. 1 (рис. 3с). Створка длиной 33.3 мкм, шириной 5 мкм, штрихов 10 в 10 мкм, линеол 35 в 10 мкм.
N. sp. 2 (рис. 3т). Створка длиной 28.9 мкм, шириной 6.1 мкм, штрихов 11 в 10 мкм, линеол 30 в 10 мкм.
N. sp. 3 (рис. 3у). Створки длиной 14.5 мкм, шириной 5.4–6.4 мкм, штрихов 12 в 10 мкм, линеол 35–40 в 10 мкм.
N. sp. 4 (рис. 3ф). Створка длиной 25.7 мкм, шириной 6.4 мкм, штрихов 11 в 10 мкм, линеол 25 в 10 мкм.
N. sp. 5 (рис. 4а). Створка длиной 31 мкм, шириной 11 мкм, штрихов 9 в 10 мкм, линеол 30 в 10 мкм.
Рис. 4.
Электронные микрофотографии створок (СЭМ): а – Navicula sp. 5, б – Nitzschia lacunarum, в – Pinnularia genkalii, г, д – P. latarae, е, ж – P. peracuminata, з – P. sp. 1, и – P. sp. 2, к – P. sp. 3, л – P. sp. 4, м – P. subrhombica var. angusta, н – (?) Rossithidium sp., о – (?) Sellaphora sp. 1, п – S. sp. 2, р – S. sp. 3, с – Stauroneis sp. 1, т – S. sp. 2, а, в, д, ж, к–п, с, т – створки с внутренней поверхности, б, г, е, з, и, р – с наружной поверхности.
Nitzschia lacunarum (рис. 4б). Створка длиной 42.8 мкм, шириной 5 мкм, фибул 12 в 10 мкм, штрихов 12 в 10 мкм. Вероятно, космополит [18].
*Pinnularia genkalii (рис. 4в). Створки длиной 100–109 мкм, шириной 19–19.5 мкм, штрихов 6 в 10 мкм. Остров Вайгач [14].
P. latarea (рис. 4г, 4д). Створки длиной 45.7–55.5 мкм, шириной 10–11 мкм, штрихов 8–10 в 10 мкм. Германия [14].
P. cf. peracuminata (рис. 4е, 4ж). Створки длиной 75.5–80 мкм, шириной 14.4 мкм, штрихов 8 в 10 мкм. Германия [14].
P. sp. 1 (рис. 4з). Створка длиной 53.3 мкм, шириной 8.9 мкм, штрихов 8 в 10 мкм.
P. sp. 2 (рис. 4и). Створка длиной 93 мкм, шириной 13.3 мкм, штрихов 8 в 10 мкм.
P. sp. 3 (рис. 4к). Створка длиной 88 мкм, шириной 13.2 мкм, штрихов 9 в 10 мкм.
P. sp. 4 (рис. 4л). Створка длиной 113 мкм, шириной 16 мкм, штрихов 9 в 10 мкм.
P. subrhombica var. angusta Krammer (рис. 4м). Створка длиной 84.4 мкм, шириной 11 мкм, штрихов 9 в 10 мкм. Германия [14].
(?) Rossithidium sp. (рис. 4н). Створка длиной 38.6 мкм, шириной 6.5 мкм, штрихов 25 в 10 мкм.
(?) Sellaphora sp. 1 (рис. 4о). Створка длиной 26.7 мкм, шириной 9.4 мкм, штрихов 9 в 10 мкм.
S. sp. 2 (рис. 4п). Створка длиной 24.3 мкм, шириной 10 мкм, штрихов 17 в 10 мкм.
S. sp. 3 (рис. 4р). Створка длиной 22 мкм, шириной 8.5 мкм, штрихов 19 в 10 мкм.
Stauroneis sp. 1 (рис. 4с). Створка длиной 52.8 мкм, шириной 9.3 мкм, штрихов 25 в 10 мкм.
S. sp. 2 (рис. 4т). Створка длиной 153 мкм, шириной 38 мкм, штрихов 16 в 10 мкм.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Согласно единственным опубликованным данным [10], в 1990-е гг. в бассейне р. Мордыяхa выявлен 91 таксон Bacillariophyta из 5 порядков, 15 семейств, 34 родов. Наибольшим таксономическим разнообразием отличались семейства Naviculaceae (18) и Fragilariaceae (12). Видовая насыщенность остальных семейств не превышала 10. Среди родов ведущее положение заняли Gomphonema и Nitzschia (по 8 видов) [10]. По неопубликованным данным М.И. Ярушиной, в летний период 2005 и 2009 гг. зарегистрирован 121 таксон диатомовых водорослей видового и внутривидового рангов. Ведущее положение по видовому разнообразию сохранили семейства Naviculaceae (33) и Fragilariaceae (19). Наибольшее таксономическое богатство было в родах Pinnularia (10), Fragilaria (9) и Eunotia (9). По данным электронной микроскопии, выявлено значительно бóльшее число таксонов диатомовых водорослей видового и внутривидового (281) и родового (63) рангов. Таксономическая насыщенность родов была сходная: Pinnulara – 33, Nitzschia – 27, Navicula s. str. – 24, Eunotia – 18. Высокое видовое разнообразие отмечено в озерах 8 (71), 7 (72), Пэбтавыто (116) и протоке между двумя озерами (125). В водных экосистемах западного Ямала имеет место аналогичная картина: Pinnularia (30), Navicula (35), Nitzschia (17), Eunotia (16) [5]. По нашим данным, максимальная частота встречаемости во всех исследованных озерах и реках отмечена для Diatoma tenuis, Encyonema silesiacum, Fragilaria vaucheriae, Tabellaria flocculosa и Ulnaria ulna.
Выводы. В результате первого электронно-микроскопического изучения диатомовых водорослей водотоков и водоемов бассейна р. Мордыяха (п-ов Ямал) зафиксирован 281 таксон видового и внутривидового рангов из 63 родов, в том числе 14 видов и разновидностей, новых для флоры России, 30 форм из родов Amphora, Aulacoseira, Cavinula, Cymbella, Diploneis, Encyonema, Fragilaria, Gomphonema, Humidophila, Navicula, Pinnularia, Rossithidium, Sellaphora, Stauroneis определены только до рода. Максимальное таксономическое разнообразие отмечено в оз. Пэбтавыто (116) и протоке между двумя озерами (125). Наиболее распространены в бассейне р. Мордыяха представители бесшовных диатомовых водорослей.
Список литературы
Аэрокосмический мониторинг природной среды полуострова Ямал // Газовая пром-сть. 2003. № 7. С. 72–76.
Балонов И.М. Подготовка водорослей к электронной микроскопии // Методика изучения биогеоценозов внутренних водоемов. М.: Наука, 1975. С. 87–90.
Богданов В.Д., Богданова Е.Н., Мельниченко И.П. и др. Проблемы охраны биоресурсов при обустройстве Бованенковского газоконденсатного месторождения // Экономика региона. 2012. № 4(32). С. 68–79.
Воронков Н.В. Планктон водоемов полуострова Ямал // Ежегодник Зоол. музея Импер. АН. СПб., 1911. Т. 16. № 2. С. 180–214.
Генкал С.И., Ярушина М.И. Bacillariophyta водных экосистем арктических тундр западного Ямала (бассейн р. Харасавэйяха) // Альгология. 2014. № 2. С. 195–208.
Генкал С.И., Ярушина М.И. К морфологии, таксономии и распространению редкого вида Sellaphora hustedtii (Krasske) Lange-Bertalot et Werum (Bacillariophyta) из водоемов Гыданского полуострова // Новости системат. низш. раст. 2014. Т. 48. С. 57–65.
Мониторинг биоты полуострова Ямал в связи с развитием объектов добычи и транспортировки газа. Екатеринбург: Изд-во УРЦ “Аэрокосмоэкология”, 1997. Ч. 2. 191 с.
Ямало-Гыданская область (физико-географическая характеристика). Л.: Гидрометеоиздат, 1977. 309 с.
Ярушина М.И. Фитопланктон водоемов п-ва Ямал // Биологические ресурсы Белого моря и внутренних водоемов европейского Севера: Тез. докл. Сыктывкар, 1990. С. 41.
Ярушина М.И. Фитопланктон // Биология гидробионтов экосистемы р. Мордыяхи. Свердловск, 1991. С. 25–45. Рукопись деп. в ВИНИТИ. 06.06.1991, № 2367-В. С. 25–45.
Ярушина М.И. Фитопланктон водоемов бассейна р. Мордыяхи // Современное состояние растительного и животного мира полуострова Ямал. Екатеринбург: Уральск. издат. фирма, 1995. С. 37–40.
Krammer K. Die cymbelloiden Diatomeen. Teil 1: Allgemeines und Encyonema part. // Bibl. Diatomologica. 1997. Bd 36. 382 s.
Krammer K. Die cymbelloiden Diatomeen. Teil 2: Encyonema part., Encyonopsis und Cymbellopsis // Bibl. Diatomologica. 1997. Bd 37. 469 s.
Krammer K. Diatoms of Europe. V. 1: Pinnularia. Ruggell: A.R.G.Gantner Verlag K.G. 2000. 703 p.
Krammer K. Diatoms of Europe. V. 3: Cymbella. Ruggell: A.R.G.Gantner Verlag K.G. 2002. 584 p.
Krammer K. Diatoms of Europe. V. 4. Cymbopleura, Delicata, Navicymbula, Gomphocymbellopsis, Afrocymbella. Ruggell: A.R.G.Gantner Verlag K.G. 2003. 530 p.
Krammer K., Lange-Bertalot H. Teil 1: Naviculaceae // Die Süsswasserflora von Mitteleuropa. Stuttgart: Gustav Fischer Verlag, 1986. Bd 2/1. 876 s.
Krammer K., Lange-Bertalot H. Bacillariophyceae. Teil. 2: Epithemiaceae, Bacillariaceae, Surirellaceae // Die Süsswasserflora von Mitteleuropa. Stuttgart: Gustav Fischer Verlag, 1988. Bd 2/2. 536 s.
Krammer K., Lange-Bertalot H. Bacillariophyceae. Teil 3: Centrales, Fragilariaceae, Eunotiaceae // Die Süsswasserflora von Mitteleuropa. Stuttgart; N.Y.: Gustav Fischer Verlag, 1991. 576 s.
Krammer K., Lange-Bertalot H. Bacillariophyceae. Teil 4: Achnanthaceae, Kritische Erganzungen zu Navicula (Lineolatae) und Gomphonema // Die Süsswasserflora von Mitteleuropa. Stuttgart: Gustav Fischer Verlag, 1991. Bd 2/4. 437 s.
Lange-Bertalot H. Diatoms of Europe. V. 2. Navicula sensu stricto, 10 genera separated from Navicula sensu lato Frustulia. Ruggell: A.R.G.Gantner Verlag K.G. 2001. 526 p.
Lange-Bertalot H., Bak M., Witkowski A. Diatoms of Europe. V. 6. Eunotia and some related genera. Ruggell: A.R.G. Gantner Verlag K.G. 2011. 747 p.
Lange-Bertalot H., Moser G. Brachysira-Monographie der Gattung. Wichtige indicator-species für das gewässer-monitoring und Naviculadicta nov. gen. ein lösungsvorschlag zu dem problem Navicula sensu lato onhe Navicula sensu stricto // Bibl. Diatomologica. Berlin; Stuttgart, 1994. Bd 29. 212 s.
Levkov Z. Diatoms of Europe. V. 5: Amphora sensu lato. 2009. 916 p.
Levkov Z., Metzeltin D., Pavlov A. Luticola, Luticolopsis // Diatoms of Europe. 2013. V. 7. 697 p.
Reichardt E. Zur revision der gattung Gomphonema // Iconographia Diatomologica. 1999. V. 8. S. 1–203.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод