

Биология внутренних вод, 2020, № 5, стр. 419-430
Особенности биоморфы Hydrilla verticillata (Hydrocharitaceae)
А. Н. Ефремов a, *, А. А. Пушкина a, Я. В. Болотова b
a Омский государственный педагогический университет
Омск, Россия
b Амурский филиал Ботанического сада-института, Дальневосточное отделение Российской академии наук
Благовещенск, Россия
* E-mail: stratiotes@yandex.ru
Поступила в редакцию 27.09.2019
После доработки 20.11.2019
Принята к публикации 10.12.2019
Аннотация
Изучены биоморфологические особенности Hydrilla verticillata (L. fil.) Royle в тропических и умеренных районах евразиатской части ареала. Побеговая система длительно нарастающая, симподиальная, состоящая из одно- или малолетних моноподиальных побегов главного и последующих порядков. Наиболее изменчивые признаки – количество листьев в узле; форма, длина и ширина листовой пластинки; характер ветвления побега; длина междоузлия. Hydrilla verticillata является многолетним (вегетативно малолетним) поликарпическим укореняющимся длиннопобеговым турионово-клубневым вегетативно-подвижным гидрофитом. Пластичность биоморфы у разных биотипов H. verticillata проявляется в продолжительности онтогенеза побегов и изменении соотношения образуемых специализированных вегетативных диаспор. Турионы и клубни представляют собой две альтернативные модели поддержания численности.
ВВЕДЕНИЕ
Hydrilla verticillata (L. fil.) Royle – погруженное, укореняющееся, одно- или двудомное, многолетнее (факультативно однолетнее) растение, переносящее неблагоприятные условия в форме вегетативных диаспор, реже – семян. Высокая экологическая пластичность и некоторые особенности биоморфы позволили сформировать обширный географический ареал (Cook, Lüönd, 1982; Efremov et al., 2019; Sousa, 2011). Содержание растворенных питательных веществ, активная реакция и минерализация воды, тип донных осадков, освещенность и скорость течения существенно не ограничивают рост и распространение H. verticillata (Baniszewski et al., 2016; Basiouny et al., 1978; Cook, Lüönd, 1982; Efremov et al., 2019; Steward et al., 1984; Sousa, 2011).
Ранее (Mitra, 1956; Mitra, 1966; Netherland, 1997; Yeo et al., 1984) была исследована морфология двух типов вегетативных диаспор, во многом определяющих особенности биоморфы данного вида. В последнее время контроль за инвазией H. verticillata вызвал интерес к изучению способа возобновления и морфогенеза (Benoit, 2011; Bianchini et al., 2010; Hofstra et al., 1999; Langeland, 1996; Netherland, 1997; True-Meadows et al., 2016).
Цель работы – исследовать пластичность биоморфы H. verticillata в евразиатской части ареала, уделив основное внимание морфологии вегетативных диаспор и побега.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Область исследования. Основной сбор материала проведен в 2008–2018 гг. в следующих географических районах: Восточная Европа (Саратовская обл., Россия), Западная Сибирь (Омская, Кемеровская, Новосибирская области, Россия; Северо-Казахстанская, Восточно-Казахстанская области, Казахстан), Средняя Сибирь (Иркутская обл., Россия), Дальний Восток (Хабаровский и Приморский края, Амурская обл., Россия; провинция Хэйлунцзян, Китай), Юго-Восточная тропическая Азия (Гуанси-Чжуанский автономный район, Китай; штат Керала, Индия; провинция Чанг Май, Тайланд). Использованы материалы авторской базы данных, включающей сведения о 949 образцах из гербариев BP, ERA, FI, IBCS, IBIW, LE, LISP, MW, NS, NSK, OMSK, P, PRC, QBG, RIG, TK, VLA, W, WAG, Z+ZT, и портала Global Biodiversity Information Facility, GBIF (https://www.gbif.org/). Акронимы коллекций приведены согласно Index Herbariorum (http://sweetgum.nybg.org/science/ih/).
Морфологические исследования. Основной объем морфологических исследований (морфология побега, определение половых типов цветков) проведен на образцах из различных частей ареала, охватывающих максимальный диапазон изменчивости. Морфолого-анатомические исследования специализированных диаспор и генеративных органов выполнены для образцов, полученных из Омской, Новосибирской и Амурской областей (Россия), Гуанси-Чжуанского автономного района (Китай) и штата Керала (Индия).
Методологической базой для биоморфологического исследования послужил сравнительно-морфологический подход (Серебрякова, 1972; Свириденко, 2000). Морфологический анализ проведен на живом и фиксированном материале (гербарном, фиксированном в 70%-ном растворе этанола) по традиционной схеме (O’Brien, McCully, 1981). Отдельные органы исследованы светлопольным методом с помощью микроскопа проходящего света “Микмед-2-1600” (Россия) и стереомикроскопа “Микромед МС 2 Zoom. 2 CR” (Китай). Срезы выполнены бритвой и с помощью ротационного микротома “МЗП-01 Технон” (Россия). Морфометрический анализ проведен с использованием Image-Pro Inside 9 с фотоснимка после его предварительной калибровки. Коэффициент формы, как базовый геометрический параметр, определен для проекции исследованных органов. В качестве фонового красителя использован метиленовый синий (O’Brien, McCully, 1981), для детализации гистологических комплексов − метод комбинированной окраски (Михальцов, 2012). Для измерений, проведенных на гербарном материале, определен коэффициент изменения линейных размеров листа при высушивании, который равнялся 0.7 ± 0.2.
Проведена статистическая обработка результатов (проверка на нормальность распределения, описательная статистика, t-тест) средствами Statistica 6.5. В качестве показателей описательной статистики даны средние значения и стандартные отклонения.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
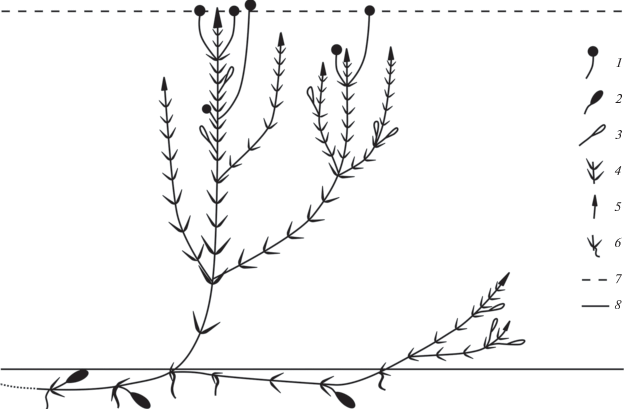
Система побегообразования. Побеговая система H. verticillata длительно нарастающая, симподиальная, состоящая из одно- или малолетних моноподиальных побегов главного и последующих порядков. В системе побегов H. verticillata (рис. 1) выделяются геотропные удлиненные побеги с адвентивными корнями и ортотропные, плагиотропные удлиненные, развивающиеся из узлов геотропных побегов, несущие побеги (n + 2)-порядка и репродуктивные структуры. Боковые побеги (n + 2)-порядка напоминают основной побег.
Рис. 1.
Схема биоморфы Hydrilla vertcillata: 1 – цветок, 2 – клубень, 3 – турион, 4 – листовая мутовка, 5 – апекс побега, 6 – адвентивный корень в узле, 7 – уровень воды, 8 – уровень грунта.
Основные результаты статистического анализа показателей вегетативного побега приведены на рис. 2 (гербарный материал). В средней части ортотропного побега междоузлия длиной 1.3 ± 0.7 см (рис. 2д), в основании – более удлиненные, до 10–20 см (рис. 1). Листорасположение мутовчатое, каждая мутовка состоит из 4.7 ± 1.5 сидячих листьев; чаще встречаются растения с четырьмя, пятью, реже – тремя, шестью листьями в узле (рис. 2е).
Рис. 2.
Основные морфологические параметры средней и верхней частей вегетативного побега (n = 618) Hydrilla vertcillata: а – длина листовой пластинки; б – ширина листовой пластинки; в – площадь листовой пластинки; г – коэффициент формы листовой пластинки; д – длина междоузлия; е – количество листьев в узле. Заштрихованные “контуры” показывают 25–75%-ные значения вероятного отклонения, белые “контуры” − размах без выбросов (квадраты соответствуют медиане, окружности – выбросам), “*” – крайние точки. Измерения проведены на гербарном материале.
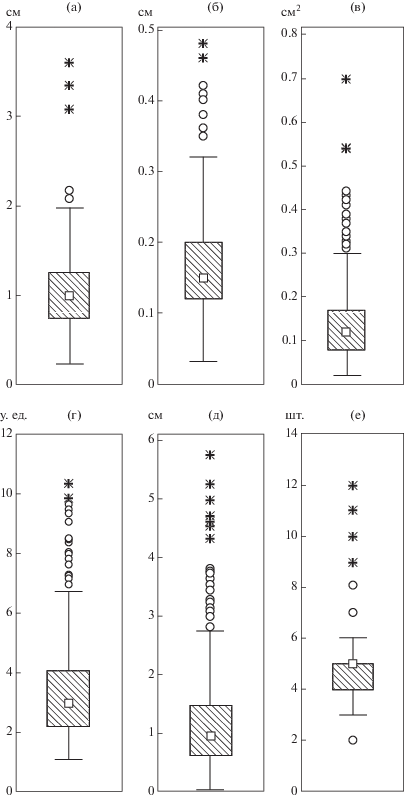
Морфологические параметры листовой пластинки H. verticillata в различных частях ареала существенно различаются (рис. 2а–2г, 3). У изученных образцов длина листовой пластинки в средней и верхней частях побега почти равна длине междоузлия и составляет 1.0 ± 0.4 см, ширина – 0.16 ± 0.07 см (рис. 2а, 2б); площадь листовой пластинки очень вариабельна (рис. 2в). Соотношение длины листовой пластинки к ширине у листьев в средней и верхней частях побега равно 5–7 : 1; коэффициент формы, характеризующей геометрию листа, – 3.3 ± 1.5 (рис. 2г).
Рис. 3.
Вариабельность числа и формы листовых пластинок средней и верхней частей побега Hydrilla vertcillata. a – Unknown place, Planten van de hortus botanicus Leiden, H.J. van Hattum, 15.VII.1965, WAG; б – USA, Indiana, city of Rochester, Lake Manitou, M.S. Alix, R. Scribailo, 26.IX.2006, GBIF; в – Rwanda, north end of Lac Bulera, near Kagogo/Idaho, R.H. Markham, 8.VI.1983, cultivated in Botanical garden of the University Zurich, C.D.K. Cook, 30.VIII.1983, Z+ZT; г – Java, in ditch growing wild in Bogor Botanic Garden, H. Wanner, 1978, cultivated in Botanical garden of the University Zurich, 26.II.1980, R. Lüönd, Z+ZT; д – India, Kerala state, Kottayam district, ca. 6 km east of Kottayam, C.D.K. Cook, 16.X.1979, Z+ZT; е – China, Hubei Province, near the city of Wuhan, north part of Futou Lake, about 30 km south of Wuhan, C.D.K. Cook, 18.VIII.1990, Z+ZT; ж – India, Bharatpur, S.K. Banerjee, H.Y. Mohan Ram, 25.XII.1956, WAG; з – USA, Texas, channels in Colorado River, east side of Austin, T. Wendt, M. Lyday, 22.X.2011, GBIF; и – Burundi, Ramonge, 73 km south of Bujumbura, Lake Tanganyika, E.A. Okrah, R.H. Markham, P. Ndayiragigii, 12–26.V.1990, GBIF; к – China, Yangsho County, Guilin Prefecture, Guangxi Zhuang Autonomus Region, Lijang River, A. Efremov, 02.I.2017, сборы авторов; л – Россия, Омская обл., Горьковский р-н, окрестности пос. Саратово, оз. Саратово, Б.Ф. Свириденко, 10.VIII.2006, LE; м – Afghanistan, Province Laghman, 2 km östlich Laghman (Charbagh) an der Strabe von Mitarlam nach Darunta, D. Podlrch, 23.X.1970, W.

В базальной части ортотропного побега и на столонах листья обычно яйцевидные или широкоовальные, с широко расставленными зубцами или почти цельнокрайние. Среднее отношение длины листовой пластинки к ширине у таких листьев 2–3 : 1, коэффициент формы 1–2.
Специализированные побеги. Для H. verticillata характерно образование специализированных укороченных вегетативных побегов: клубней и турионов (рис. 4). Клубни развиваются на погруженных в грунт геотропных побегах, с которыми связаны нефотосинтезирующими столонами, фотосинтезирующие турионы со столонами развиваются на ортотропных побегах аксиально или терминально (рис. 1).
Рис. 4.
Морфолого-анатомические особенности специализированных вегетативных диаспор Hydrilla vertcillata: a – молодой клубень со столоном; б – апикальная зона клубня, продольный срез, комбинированная окраска (видны листовые зачатки и внутрипазушные чешуи); в – клубень, общий вид; г – клубень, продольный срез; д – турион, общий вид; е – турион, продольный срез, окраска нейтральным красным (внутрипазушные чешуи темно окрашены).
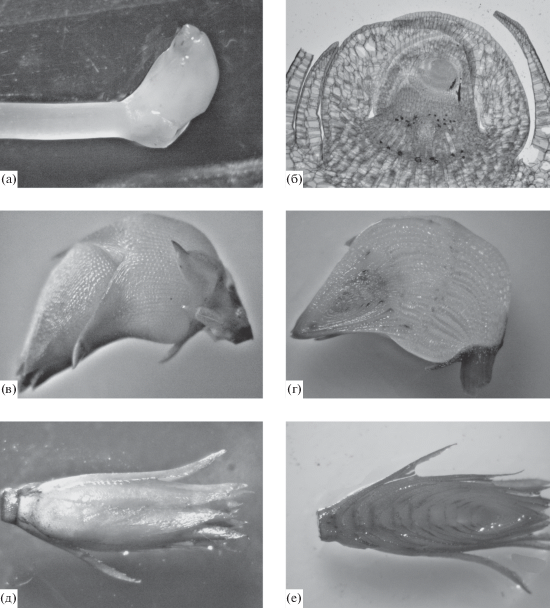
Столоны, как правило, длиной 0.2–1.5 см, до 2–5 см у клубней, хрупкие, с удлиненными 1–2 междоузлиями, несущими 2–4 укороченных чешуевидных листа (рис. 4а). В узле располагаются один, реже два клубня (туриона). Клубни по сравнению с турионами крупнее (отмечали Spencer, Ksander, 2001; Yeo et al., 1984), округло-эллиптические (коэффициент формы составляет 1.2 ± 0.1), с меньшим количеством метамеров, турионы овально-ланцетные, более вытянутые (коэффициент формы − 1.5 ± 0.3) (рис. 4а, 4в, 4д, табл. 1).
Таблица 1.
Морфометрическая характеристика специализированных вегетативных диаспор Hydrilla verticillata
| Показатель | Турионы | Клубни |
|---|---|---|
| Длина, см | 0.7 ± 0.3 | 0.8 ± 0.2 |
| Ширина, см | 0.3 ± 0.1 | 0.4 ± 0.1 |
| Коэффициент формы | 1.5 ± 0.3 | 1.2 ± 0.1 |
| Количество метамеров, шт. | 8 ± 3 | 5 ± 1 |
| Пропорция осевой части, %* | 41 ± 10 | 22 ± 4 |
| Пропорция листьев, %* | 59 ± 10 | 78 ± 4 |
| Доля базальной части, %** | 14 ± 5 | 39 ± 1 |
| Площадь тангентального среза, см2* | 0.09 ± 0.04 | 0.23 ± 0.09 |
Примечание. Приведены средние значения и стандартное отклонение для живого и фиксированного в этаноле материала (n = 68). Пропорция осевой части (листьев) определена как доля площади тангентального среза, приходящаяся на долю осевой части побега (листьев). * – статистически значимые различия при p ≤ 0.15. ** – при p ≤ 0.05.
У турионов преобладает пропорция ассимилирующих листьев, а у клубней – запасающей осевой части побега, паренхима которой несет многочисленные крахмальные зерна (табл. 1). Соотношение гистологических зон у специализированных побегов определяет стратегию возобновления.
Генеративное возобновление. Для оценки характера возобновления у гербарных образцов определена доля цветущих и плодоносящих растений, изучены особенности формирования соцветий. Основные результаты морфолого-анатомических исследований генеративных органов и их интерпретация приведены в разделе “Обсуждение результатов”.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Побеговая система. Вегетативный (вегетативно-генеративный) ортотропный побег H. verticillata состоит из 30–100 метамеров. Длина почти цилиндрического побега обычно составляет 1.0–1.5 м, но может достигать ≥2–3 м (Al-Mandeel, 2013; Cook, Lüönd, 1982). Такие длинные побеги могут образовывать плотные маты, достигающие 20% общей биомассы растения (Spencer, 2011). Ортотропные побеги ‒ умеренно разветвленные у основания (n + 1) и более обильно в средней части (n + 2–3 или n + 3) (60% исследованных случаев). Благодаря мезотонному ветвлению основная зона обогащения находится на уровне, позволяющем максимально эффективно использовать ресурсы хорошо прогреваемой и освещенной водной толщи, колебания уровня оказывают меньшее влияние.
Геотропные побеги ‒ укореняющиеся, состоят из 10–30 удлиненных метамеров 10–20 см длиной, слабо ветвящиеся. Они достигают 8.5 м длины (обычно 2–5 м), погружены в донные осадки на глубину до 20 см (Mitra, 1956), их скорость роста достигает 2 см/сут. (Cook, Lüönd, 1982). Такой тип побега обеспечивает закрепление в обводненных грунтах и захват новых территорий.
Листья сидячие в мутовках по 3–8(12) или супротивные (Cook, Lüönd, 1982; Symones, Sosef, 2015; Wang et al., 2016), что подтверждается результатами выполненного анализа (рис. 2а). В средней части побега листья линейные, ланцетные, редко широкоовальные (например, у основания ортотропных побегов), обычно 7–15(20) × × 1–4 мм, зеленые, с красновато-коричневыми пятнами и полосами (идиобласты) или без них, слабо полустеблеобъемлющие, острые, оканчивающиеся одним шипиком, голые (Cook, Lüönd, 1982; Haynes, 2001; Symones, Sosef, 2015; Wang et al., 2016). Однодомным растениям, выращенным ex situ, свойственны более короткие листья (12.0 × 2.1 мм) и более длинные междоузлия (16.1 × 1.5 мм), чем двудомным (14.0–16.1 × 0.8–1.5 и 11.0–12.0 × 0.8–2.1 мм соответственно) (Les et al., 1997). Средняя жилка отчетливая, часто несущая шипики на абаксиальной поверхности. Край листа мелкопильчато-зубчатый, с одним–тремя рядами гиалиновых клеток и одноклеточными шипиками (Cook, Lüönd, 1982; Haynes, 2001; Symones, Sosef, 2015; Wang et al., 2016). Морфология края листовой пластинки является важным и относительно стабильным диагностическим признаком, зубцы обычно хорошо видны невооруженным глазом (3а, 3б, 3г–3з, 3л), реже – расставлены и очень мелкие (3в, 3к, 3м). Погруженный лист коротко-линейной формы, свойственный H. verticillata, – анцестральный для Hydrocharitaceae (Chen et al., 2012).
Для H. verticillata, как и других Hydrocharitaceae, характерно наличие внутрипазушных чешуй (squamules intravaginales), которые у изученных образцов располагаются по 2(4) в узле и имеют размер 0.5 × 0.1–0.2 мм (рис. 4е, 5д, 5е). Внутрипазушные чешуи от узко-треугольных до ланцетных, полупрозрачные, нерегулярно окаймленные оранжево-коричневыми, (суб-)маргинальными, (одно-)двуклеточными выростами (Cook, Lüönd, 1982), наиболее длинными в апикальной зоне (рис. 5д, 5е).
Рис. 5.
Морфолого-анатомические особенности генеративных органов и внутрипазушных чешуй Hydrilla vertcillata: a – мужское соцветие; б – женский цветок в стадии бутонизации; в – женский цветок в фазе полного цветения; г – незрелый плод; д – внутрипазушные чешуи в основании листа туриона, окраска нейтральным красным; е – внутрипазушная чешуя, общий вид.

Корневая система. Корни H. verticillata неветвящиеся, 5–20 см длиной, диаметром до 2–3 мм, адвентивные, формирующиеся в узлах геотропных побегов (Cook, Lüönd, 1982). Кора представлена двумя слоями клеток колленхимы, внутренние слои – аэренхима с крахмалоносной колленхимой. Пояски Каспари не диагностируются. Ксилема центрального цилиндра с множеством лакун, имеются ситовидные трубки и клетки-спутницы (Al-Mandeel, 2013).
Неспециализированное вегетативное возобновление. Способность H. verticillata заселять новые места обитания во многом связана с существованием нескольких способов возобновления с помощью специализированных и неспециализированных структур. Hydrilla может возобновляться фрагментами ортотропных или плагиотропных побегов, может регенерировать даже из отдельных мутовок и расти до 2.5 см/сут. (Basiouny et al., 1978; Langeland et al., 1992; Netherland, 1997). Экспериментально подтвержденная скорость роста из фрагментов побега все же существенно ниже скорости роста из клубней (Bianchini et al., 2010).
Умецу и др. (Umetsu et al., 2012) показали, что неспециализированные вегетативные пропагулы демонстрируют две стратегии выживания: колонизация (способность развивать корни) и регенерация (способность развивать новые пропагулы, которые могут рассеиваться).
Специализированное вегетативное возобновление. Турионы (пазушные турионы, turions) и клубни (подземные турионы, терминальные турионы, стеблевые клубни, клубнелуковицы, swollen stem, subterranean turions, tubers) представляют различные стратегии выживания и расселения (Al-Mandeel, 2013; Langeland, 1996; Miller et al., 1993; Spencer, 2011; Van, Steward, 1990; Yeo et al., 1984). Вероятно, образование специальных вегетативных диаспор определяется условиями обитания, максимум образования клубней наблюдался в диапазонах глубины воды 1–2 м, турионов – 1.5–4 м (Hofstra et al., 1999; Van, Steward, 1990).
Подземные специализированные диаспоры H. verticillata (по тексту – клубни) можно идентифицировать как клубнелуковицы, имеющие утолщенный из-за сильного разрастания укороченных междоузлий стебель, окруженный сочными чешуями, и единственную апикальную меристему (Жмылев и др., 2013). Клубни от беловатого до почти черного цвета. Две трети клубня (базальные) вздутые и заполнены крахмалом, одна треть (терминальная) содержит апикальную меристему, изогнутую под углом 90° (рис. 4в, 4г). Клубни отделяются от родительского растения при разложении геотропных побегов (Yeo et al., 1984). Прорастая, специализированные диаспоры формируют первое длинное междоузлие с бледными, нефотосинтезирующими рудиментарными листьями (Netherland, 1997).
В США средняя масса клубней достигает 160–376 мг у двудомных и 179–202 мг у однодомных растений, масса турионов у однодомных растений – 36–77 мг (Netherland, 1997; Spencer, 2011). С использованием метода мезокосм установлено, что в условиях более прохладного климата у двудомных растений плотность клубней существенно ниже (823 экз./м2 против 2142 экз./м2 в условиях теплого климата), и они крупнее (0.113 г против 0.096 г) (Henry, 2017). Клубни однодомных растений H. verticillata in situ сохраняются >4 лет в донных осадках (в Новой Зеландии − до 8 лет), в лабораторных условиях их всхожесть >90% (Hofstra et al., 1999; Van, Steward, 1990). Таким образом, клубни обеспечивают возможность многолетнего роста в занятых в настоящий момент биотопах (Van, Steward, 1990) и отличаются большей устойчивостью к высыханию (Basiouny et al., 1978).
В отличие от клубней, образующихся в узлах геотропных побегов, турионы образуются в пазухах листьев или ветвей ортотропных побегов (Mitra, 1956; Mitra, 1966; Netherland, 1997; Yeo et al., 1984). Как и клубни, имеют утолщенные чешуевидные (с многослойной паренхимой и толстой кутикулой) фотосинтезирующие листья, защищающие спящую меристему (рис. 3б). Турионы формируют зону отделения и опускаются на дно, где прорастают или остаются плавающими в толще воды и погружаются на грунт поздней осенью (Lakshmanan, 1951; Mitra, 1966; Spencer, 2011; Yeo et al., 1984). Обычно они прорастают на следующий год (Langeland et al., 1992; Van, Steward, 1990). При этом, оптимально процесс проходит при температуре 15–35°С (Netherland, 1997). По сравнению с клубнями, турионы прорастают раньше и при более низких температурах (Spencer, Ksander, 2001; Sousa, 2011).
Турионы, в большей степени, чем клубни, обладают способностью рассеиваться водными потоками и имеют бóльшую вероятность закрепиться на незанятой территории (Miller et al., 1993). Такие специализированные надземные (axillary tuber) (Pilon et al., 2003) и подземные (клубни, tuber) (Лапиров, 1995; Pilon et al., 2003) побеги возобновления известны и у широко распространенного гидрофита Stuckenia pectinata (L.) Börner (Pilon et al., 2003).
Генеративное возобновление. Hydrilla verticillata часто обильно и продолжительно цветет, в тропиках в благоприятных условиях – круглогодично (Cook, Lüönd, 1982), однако обильное плодоношение наблюдается редко. Только 36% изученных образцов в гербарных коллекциях имели цветки, 2.5% – плоды. Это может быть связано со следующими причинами: половая сегрегация (у двудомных биотипов не всегда в пределах возможной зоны опыления находятся растения обоих полов); непропорциональное распределение растений различных половых типов, соотношение пестичных и тычиночных растений у изученных образцов 12 : 1; вероятное отсутствие плодоношения у анеу- и полипоидов (Cook, Lüönd, 1982); общая тенденция у гидрофильных растений к снижению доли генеративного размножения.
В переделах ареала растения различных половых типов встречаются неравномерно. Среди исследованных гербарных образцов 5.4% имели мужские цветки (вероятно, это число больше, поскольку мужские цветки хорошо сохраняются только в стадии бутонизации), 92.3% − женские, 2.3% растений были с мужскими и женскими цветками. По материалам натурных наблюдений на юге Западной Сибири преобладают тычиночные растения (данные авторов), на Дальнем Востоке – пестичные (Пробатова, Буч, 1981).
Соцветия развиваются в пазухах листьев средней и верхней частях ортотропных побегов, брактеи парные (интеркалярные фрондозные), сросшиеся в основании. Мужские цветки на короткой цветоножке, собраны обычно по 4–6 в узле, с шаровидным покрывалом диаметром 1.5 мм и с придатками на верхушке (рис. 5а). Женские цветки одиночные, реже парные, с гипантием длиной 10–50 мм. Околоцветник из трех белых лепестков, чередующихся с тремя беловатыми более крупными чашелистиками, сильно выпуклыми у мужских цветков (рис. 5б, 5в) (Cook, Lüönd, 1982; Haynes, 2001; Symones, Sosef, 2015; данные авторов).
Плоды у исследованных образцов ‒ длиной 9 ± 3 мм (до 15 мм) (Cook, Lüönd, 1982), длина зависит от количества семян), гладкие или c боковыми отростками (рис. 5г) (Cook, Lüönd, 1982; Wang et al., 2016; данные авторов). По мере развития нижняя часть плода утолщается и заполняется слизистым содержимым. Завязь ‒ нижняя, одногнездная с несколькими (до шести) семяпочками, плацентация свободная париетальная. Семена в количестве 1-3(6), коротко-цилиндрические, заостренные к обоим концам, каждое размером 5–6 × 0.8–1.0 мм; зародыш прямой, эндосперм крахмалистый (Пробатова, Буч, 1981).
Морфологические биотипы. Hydrilla verticillata отличается значительной морфолого-биологической изменчивостью, формирует два генетически разных биотипа (формы): двудомный и однодомный. Мадейра и др. (Madeira et al., 1997; 1999) обнаружили существенные генетические различия у исследованных образцов из разных частей мирового ареала. Отдельные изолированные популяции (например, однодомные растения в Японии), не демонстрируют высокой изменчивости (Nakamura et al., 1998). Центр генетического разнообразия H. verticillata находится в юго-восточной Азии (Zhu et al., 2015).
Однодомный биотип H. verticillata − травянистые многолетние (вегетативно-малолетние) растения с разрушающимися в зимний период побегами и возобновляемые посредством турионов и малолетних перезимовавших побегов (Benoit, 2011; True-Meadows et al., 2016). Рост наблюдается в течение всего года, менее интенсивно в зимние месяцы (Steward et al., 1984; Sutton et al., 1992; True-Meadows et al., 2016). Отмечено, что при более низких температурах турионы однодомного биотипа прорастают с большей скоростью (True-Meadows et al., 2016). Прорастание турионов обычно происходит до прорастания клубней (Spencer, Ksander, 2001). После периода интенсивного формирования клубней биомасса растений уменьшается, и они отрываются от субстрата (True-Meadows et al., 2016). Несмотря на то, что продукция клубней у однодомной H. verticillata на 50% выше, чем у двудомной, их средняя масса на 32% ниже (Sutton et al., 1992).
Двудомный биотип H. verticillata – более длительно вегетирующие растения, продуцирующее меньше турионов (Sutton et al., 1992; True-Meadows et al., 2016). Этот биотип зимует в вегетативной форме и формирует меньше клубней.
Считается, что однодомный биотип преобладает в тропических широтах, двудомный – в умеренных (Cook, Lüönd, 1982). Однако распределение двух биотипов в Северной Америке противоречит этому наблюдению (Langeland, 1996; Netherland, 1997; Steward et al., 1984). В обследованных биотопах Западной Сибири, Дальнего Востока и Индии преобладают двудомные растения. В культуре растения могут сформировать цветки одного пола в течение одного или двух сезонов, а затем – другого (Cook, Lüönd, 1982).
Следует отметить, что у растений H. verticillata в тропических широтах доля клубней выше, в умеренных широтах более многочисленны пазушные турионы. У Stuckenia pectinata, формирующей также специализированные вегетативные диаспоры, в северных популяциях с укороченным жизненным циклом биомасса клубней выше, в низких широтах жизненный цикл удлиняется и увеличивается общая биомасса растений (Pilon et al., 2003).
Своеобразный габитус имеют южноазиатские растения H. verticillata, обитающие в проточных водах: длинные слабоветвящиеся побеги (n + 1)-порядка, узкие и более длинные листья (рис. 3к), многочисленные клубни. Восточноафриканские растения, обитающие в щелочных водах, сильноветвящиеся, несут 10–25 ортотропных побегов n + 1 с укороченными междоузлиями и более короткими и широкими листьями (рис. 3и), присутствуют турионы, клубни формируются крайне редко (Pemberton, 1980; Symones, Sosef, 2015; P, QBG, Z+ZT).
В пределах ареала обнаруживаются растения с разным набором хромосом. Растения из Азии и Европы – диплоиды (2n = 2x = 16) или триплоиды (2n = 3x = 24) (Cook, Lüönd, 1982), изредка в США встречаются тетраплоиды (2n = 4x = 32) (Langeland, 1996). Двудомные и однодомные биотипы при скрещивании образуют семена в 71% случаев, из них 90% жизнеспособных (Steward, 1993).
У индийских популяций (2n = 24, 2n = 16) не выявлена устойчивая связь между морфологией побега и числом хромосом (Chaudhuri, Sharma, 1978). Однако, в большинстве случаев ди- и триплоидные расы имеют существенные морфологические различия. Так, диплоидные растения с более узкими листьями (0.7–1.7 см × 1.0–1.8 мм), с плодами, несущими 2–9 выростов и 2–6 семян, относят к разновидности H. verticillata var. verticillata. Триплоиды с листьями (0.7–1.3) см × (3–4) мм и гладкими плодами с одним–тремя семенами стерильны, без семязачатков, принадлежат к разновидности H. verticillata var. roxburghii Caspary, Jahrb. Wiss (Steward, 1993; Wang et al., 2016). Hydrilla lithuanica (Andrz. ex Besser) Dandy представляет собой стерильную триплодиную расу.
Выводы. Как и у большинства водных растений, морфология побега H. verticillata отличается пластичностью. Наиболее изменчивы ‒ количество листьев в узле, форма, длина и ширина листа, характер ветвления побега и длина междоузлий. Однодомные и двудомные биотипы H. verticillata демонстрируют отчетливые различия в сезонности формирования турионов. Биоморфа H. verticillata сходна с длиннопобеговыми биоморфами Elodea Michx., Egeria Planch. и Largosiphon Harv. Побеговая система ‒ длительно нарастающая, симподиальная, состоящая из малолетних моноподиальных побегов главного и подчиненных порядков. Соцветия диморфные, малоцветковые. Hydrilla verticillata – многолетний (вегетативно малолетний) поликарпический укореняющийся длиннопобеговый турионово-клубневой вегетативно-подвижный гидрофит. Пластичность биоморфы проявляется в продолжительности онтогенеза побегов однодомных и двудомных растений, изменении соотношения образуемых специализированных вегетативных диаспор (клубней и турионов). Специализированные вегетативные диаспоры представляют собой две альтернативные модели поддержания численности, характер их развития зависит от ряда факторов. Клубни увеличивают вероятность возобновления при действии неблагоприятных факторов, а турионы и неспециализированные побеги обеспечивают максимально быстрое закрепление и распространение. В стабильных условиях более эффективное возобновление и расселение растений происходит с помощью турионов, которые, вероятно, являются более поздним приобретением при переходе от геофитного предка к гидрофилии.
Список литературы
Жмылев П.Ю., Карпухина Е.А., Леднев С.А. 2013. К проблемам биоморфологии водных сосудистых растений // Вестник Тверского государственного университета. Биол. и экол. Т. 32. № 31. С. 137.
Лапиров А.Г. 1995. Рдест гребенчатый // Биологическая флора Московской области. М: Изд-во МГУ. Вып. 11. С. 37.
Михальцов А.Н. 2012. Модификация нового метода полихромной окраски тканей растения // Природные ресурсы, биоразнообразие и перспективы естественнонаучного образования. Омск: Изд-во ООО “Омскбланкиздат”. С. 57.
Пробатова Н.С., Буч Т.Г. 1981. Hydrilla verticillata (Hydrocharitaceae) на Советском Дальнем Востоке // Бот. журн. № 66(2). С. 208.
Серебрякова Т.И. 1972. Учение о жизненных формах растений // Итоги науки техн. Сер. Ботан. Т. 1. С. 84.
Свириденко Б.Ф. 2000. Флора и растительность водоемов Северного Казахстана. Омск: Изд-во ОмГПУ.
Al-Mandeel F.A. 2013. A new record of the invasive species Hydrilla verticillata (Linn. f.) Royal on the Iraqi rivers // Adv. Environ. Biol. V. 7(2). P. 384.
Baniszewski J., Cuda J.P., Gezan S.A. et al. 2016. Stem fragment regrowth of Hydrilla verticillata // J. Aquat. Plant Manag. V. 54. P. 53.
Basiouny F.M., Haller W.T., Garrard L.A. 1978. Survival of Hydrilla (Hydrilla verticillata) plants and propagules after removal from the aquatic habitat // Weed Science. V. 26(5). P. 502.
Benoit L.K. 2011. Cryptic speciation, genetic diversity and herbicide resistance in the invasive aquatic plant Hydrilla verticillata (L. f.) Royle (Hydrocharitaceae): Doctoral Dissertation, University of Connecticut. http://digitalcommons.uconn.edu/dissertations/AAI349-2149 (10 February 2016)
Bianchini Jr.I., Cunha-Santino M.B., Milan J.A.M. et al. 2010. Growth of Hydrilla verticillata (L.f.) Royle under controlled conditions // Hydrobiologia. V. 644. P. 301. https://doi.org/10.1007/s10750-010-0191-1
Chaudhuri J.B., Sharma A. 1978. Cytological studies on three aquatic members of Hydrocharitaceae in relation to their morphological and ecological characteristics // Cytologia. V. 43. P. 1.
Chen L.-Y., Chen J.-M., Gituru R.W., Wang Q.-F. 2012. Generic phylogeny, historical biogeography and character evolution of the cosmopolitan aquatic plant family Hydrocharitaceae // BMC Evol. Biol. V. 12(30). http://www.biomedcentral.com/1471-2148/12/30.
Cook C.D.K., Lüönd R. 1982. A revision of the genus Hydrilla (Hydrocharitaceae) // Aquat. Bot. V. 13. P. 485.
Efremov A.N., Sviridenko B.F., Bolotova Ya.V. et al. 2019. Coenocomplex and ecological features of Hydrilla verticillata (L. f.) Royle (Hydrocharitaceae) in Northern Eurasia // Inland Water Biology. V. 12(1). P. 57. https://doi.org/10.1134/S1995082919010061
Haynes R.R. 2001. Hydrocharitaceae // Flora of Thailand. V. 7(3). Bangkok: Royal Forest Department. P. 365.
Henry A.L. 2017. Monoecious Hydrilla (Hydrilla verticillata (L. f) Royle) growth and phenology in two dissimilar climates: a thesis of Master of Science. North Carolina: North Carolina State University.
Hofstra D.E., Clayton J.S. Champion P.D., Green J.D. 1999. Distribution and density of vegetative Hydrilla propagules in the sediments of two New Zealand lakes // J. Aquat. Plant Manag. V. 37. P. 41.
Lakshmanan C.A. 1951. Note on the occurrence of turions in Hydrilla verticillata Presl. // J. Bombay Nat. Hist. Soc. V. 49. P. 802.
Langeland K.A., Shilling D.G., Carter J.L. et al. 1992. Chromosome morphology and number in various populations of Hydrilla verticillata // Aquat. Bot. V. 42. P. 253.
Langeland K.A. 1996. Hydrilla verticillata (L. f.) Royle (Hydrocharitaceae), The Perfect Aquatic Weed // Castanea. V. 61. P. 293.
Les D.H., Mehrhoff L.J., Cleland M.A., Gabel J.D. 1997. Hydrilla verticillata (Hydrocharitaceae) in Connecticut // J. Aquat. Plant Manag. V. 35. P. 10.
Madeira P.T., Van T.K., Steward K.K., Schnell R.J. 1997. Random amplified polymorphic DNA analysis of the phenetic relationships among world-wide accessions of Hydrilla verticillata // Aquat. Bot. V. 59(3). P. 217.
Madeira P.T., Van T.K., Center T.D. 1999. Integration of five Southeast Asian accessions into the world-wide phenetic relationships of Hydrilla verticillata as elucidated by random amplified polymorphic DNA analysis // Aquat. Bot. V. 63(2). P. 161.
Miller J.L., Haller W.T., Glenn M.S. 1993. Turion production by dioecious Hydrilla in North Florida // J. Aquat. Plant Manag. V. 31. P. 101.
Mitra E. 1956. Notes on the germination of turions in Hydrilla verticillata Presl. // Curr. Sci. V. 25. P. 25.
Mitra E. 1966. On some aspects of the morphological and anatomical studies of turions of Hydrilla verticillata (Linn. f.) Royle // J. Asiatic Soc. V. 6. P. 17.
Nakamura T., Suzuki T., Kadono Y. 1998. A comparative study of isoenzyme patterns of Hydrilla verticillata (L. f.) Royle in Japan // J. Pl. Res. V. 111. P. 581.
Netherland M.D. 1997. Turion ecology of Hydrilla // J. Aquat. Plant Manag. V. 35. P. 1.
O’Brien T.P., McCully M.E. 1981. The study of plant structure: principles and selected methods. Melbourne: Termarcarphi Pty.
Pemberton R.W. 1980. Exploration for natural enemies of final report Hydrilla verticillata in eastern Africa. U.S. Department of Agriculture Agricultural Research Service, Southern Region Aquatic Plant Control Gainesville, Fla.
Pilon J., Santamaría L., Hootsmans M., van Vierssen W. 2003. Latitudinal variation in life-cycle characteristics of Potamogeton pectinatus L.: vegetative growth and asexual reproduction // Plant Ecology. V. 165(2). P. 247.
Spencer D.F., Ksander G.G. 2001. Field evaluation of degree-day based equations for predicting sprouting of Hydrilla (Hydrilla verticillata) turions and tubers // J. Freshwater Ecol. V. 16(3). https://doi.org/10.1080/02705060.2001.9663836
Spencer L. 2011. Hydrilla verticillata // Aquatic Invasion Ecology. Fish 423. http://depts.washington.edu/oldenlab/wordpress/wp-content/uploads/2013/03/HydrillaVerticillata_ Spencer.pdf (01 March 2019).
Steward K.K., Van T.K., Carter V., Pieterse A.H. 1984. Hydrilla invades Washington, D.C. and the Potomac // Am. J. Bot. V. 71. P. 162.
Steward K.K. 1993. Seed production in monoecious and dioecious populations of Hydrilla // Aquat. Bot. V. 46. P. 69.
Sousa W.T.Z. 2011. Hydrilla verticillata (Hydrocharitaceae), a recent invader threatening Brazil’s freshwater environments: a review of the extent of the problem // Hydrobiologia. V. 669. P. 1. https://doi.org/10.1007/s10750-011-0696-2
Sutton D.L., Van T.K., Portier K.M. 1992. Growth of dioecious and monoecious hydrilla from single tubers // J. Aquat. Plant Manag. V. 30. P. 15.
Symones J.-J., Sosef M.S.M. 2015. Hydrocharitaceae // Flore d’Afrique Centrale. Zurich.
True-Meadows S., Haug E.J., Richardson R.J. 2016. Monoecious Hydrilla – a review of the literature // J. Aquat. Plant Manag. V. 54. P. 1.
Umetsu C.A., Evangelista H.B.A., Thomaz S.M. 2012. Colonization, regeneration potential and growth rates of fragments of the exotic aquatic macrophyte Hydrilla verticillata // Aquat. Biol. V. 16. P. 197. https://doi.org/10.3354/ab00450
Van T.K., Steward K.K. 1990. Longevity of monoecious Hydrilla propagules // J. Aquat. Plant Manage. V. 28. P. 74.
Wang Q., Guo Y., Haynes R.R., Hellquist C.B. 2016. Hydrocharitaceae // Flora of China. http://www.efloras.org/florataxon.aspx?flora_id=2&taxon_id=10426 (01 March 2016).
Yeo R.R., Falk R.H., Thurston J.R. 1984. The morphology of Hydrilla (Hydrilla verticillata (L. f.) Royle) // J. Aquat Plant Manag. V. 22. P. 1.
Zhu J., Yu D., Xu X. 2015. The phylogeographic structure of Hydrilla verticillata (Hydrocharitaceae) in China and its implications for the biogeographic history of this worldwide-distributed submerged macrophytes // BMC Evol. Biol. V. 15(95). https://doi.org/10.1186/s12862-015-0381-6
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод