

Биология внутренних вод, 2021, № 1, стр. 24-34
Биогенные элементы и их роль в развитии фитопланктона водохранилищ Верхней Волги
Н. М. Минеева a, *, И. Э. Степанова a, И. В. Семадени a
a Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
Ярославская обл., Некоузский р-н, пос. Борок, Россия
* E-mail: mineeva@ibiw.ru
Поступила в редакцию 20.11.2019
После доработки 24.12.2019
Принята к публикации 28.01.2020
Аннотация
Исследовано содержание Хл а и биогенные элементы в водохранилищах Верхней Волги в летний период 2015–2018 гг. При типичном для летнего максимума фитопланктона содержании Хл а (от 19.9 ± 3.9 до 48.0 ± 12.0 мкг/л) водохранилища относятся к водоемам эвтрофного типа, только в прохладном 2017 г. Рыбинское водохранилище характеризовалось как мезотрофное (10.3 ± 4.7 мкг/л). Содержание биогенных элементов (БЭ) (0.71 ± 0.06–1.54 ± 0.07 мг/л Nобщ и 51 ± 12–106 ± 3 мкг/л Pобщ) также отражает эвтрофный статус водохранилищ Верхней Волги. Средние величины Nобщ/Pобщ (11–16) указывают на отсутствие биогенного лимитирования фитопланктона, лишь в Рыбинском (2015, 2016 гг.) и Иваньковском (2015 г.) водохранилищах отмечен дефицит БЭ. В большинстве случаев между содержанием Хл а и БЭ получены невысокие коэффициенты корреляции (r <0.7), свидетельствующие о сложном и многокомпонентном влиянии БЭ на фитопланктон. Множественный корреляционный анализ подтверждает высокую значимость гидрологических условий для развития автотрофного планктона водохранилищ Верхней Волги.
ВВЕДЕНИЕ
Водохранилища Верхней Волги относятся к объектам многолетних экологических исследований, успешное проведение которых обеспечивается использованием интегральных количественных биологических характеристик, обладающих индикаторными свойствами. Фотосинтетические пигменты служат таким индикатором для фитопланктона – основного продуцента автохтонного органического вещества в экосистеме волжских водохранилищ, играющего ключевую роль в трансформации вещества и энергии. К основным факторам развития и функционирования фитопланктона относится обеспеченность клеток элементами минерального питания. Изучение влияния БЭ на развитие водорослей стало центральной темой лимнологии в последней четверти ХХ в. в связи с усилившимся вниманием к эвтрофированию водоемов. Многочисленные исследования на разнотипных озерах позволили получить эмпирические связи содержания Хл а (биомассы фитопланктона) с концентрациями фосфора и азота. Ставшие классическими работы этого направления (Eutrophication…, 1982; Vollenweider, 1979 и др.) широко цитируются в современных публикациях (Минеева, 2004; Palsson, Graneli, 2004; Søndergaard et al., 2017 и др.). Содержание БЭ служит фактором, определяющим трофию водоема, и вместе с содержанием Хл а относится к маркерам трофического статуса (см. сводку: Китаев, 2007). Новая волна интереса к влиянию БЭ на развитие водорослей возникла при анализе изменений, происходящих в водных экосистемах в условиях глобального потепления, которое проявляется в увеличении приземной температуры воздуха и температуры воды водоемов (Второй…, 2014). Само повышение температуры считается эвтрофирующим фактором, способствующим росту внутренней фосфорной нагрузки, а также более обильной и продолжительной вегетации цианопрокариот (Jeppesen et al., 2005). С потеплением связывают изменения структуры сообществ и скорости метаболизма планктона, изменение режима стратификации и потока питательных веществ, расширение бескислородной зоны (Hallstan et al., 2013; Lewandowska et al., 2014; Xiao et al., 2018). Вариации трофического статуса под влиянием глобальных и региональных факторов выявлены для водоемов, на которых проводят многолетние наблюдения: Нарочанские озера (Zhukova, 2013), Рыбинское водохранилище (Минеева, 2016; Структура…, 2018). Изучение фотосинтетических пигментов и содержания БЭ в воде водохранилищ Верхней Волги ведется на протяжении нескольких десятилетий (Пырина, 2000; Экологические…, 2001; Минеева, 2004, 2016; Stepanova, Bikbulatova, 2016; Степанова, 2018; Структура…, 2018; Mineeva, 2019). К настоящему времени накоплены обширные материалы, которые дают представление о состоянии и развитии их экосистем и демонстрируют стабильный трофический статус Иваньковского водохранилища, повышение трофии Угличского и ее циклические изменения в Рыбинском. Однако исследования связи продуктивности фитопланктона с БЭ единичны (Минеева, Разгулин, 1995; Минеева, 2004; Степанова и др., 2012; Структура…, 2018). Данное обстоятельство определило цель настоящей работы как анализ взаимосвязи между содержанием Хл а и БЭ в крупных перемешиваемых водохранилищах Верхней Волги в современных условиях глобальных климатических изменений.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
В работе использованы материалы полевых наблюдений на 26 станциях Иваньковского, Угличского и Рыбинского водохранилищ в августе 2015−2018 гг. (рис. 1). Хл а определяли в интегральных (0 м–дно) пробах воды флуоресцентным методом (Гольд и др., 1986), который позволяет оценивать суммарное количество Хл (ƩХл) по его содержанию у основных представителей пресноводного фитопланктона – синезеленых (цианопрокариот), диатомовых и зеленых водорослей (ХлCyan, ХлBac, ХлChl соответственно). Анализ БЭ выполняли в пробах воды из верхнего 0.5-метрового слоя. Определение неорганических форм азота и фосфора проводили стандартными методами (Семенов, 1977), Nобщ и Pобщ – после предварительного окисления органических фракций персульфатом калия соответственно до нитратов и ортофосфатов (Бикбулатов, 1974; Гапеева и др., 1984). При статистической обработке данных – расчете средних показателей, их погрешностей, Cv, r и R2, уравнений регрессии, проведения факторного анализа и построения графиков использовали стандартные программные пакеты для персонального компьютера.
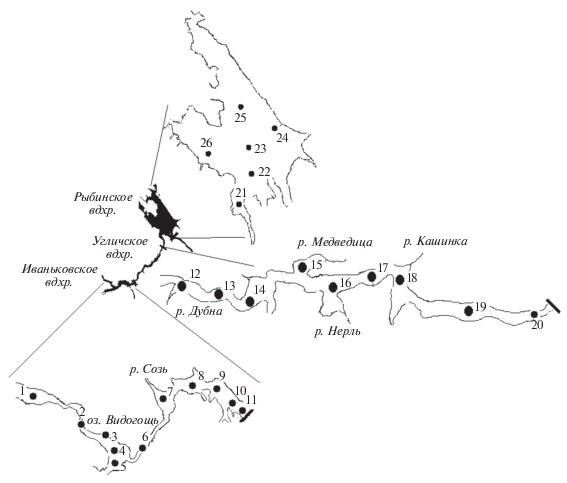
Водохранилища Верхней Волги – крупные относительно мелководные водоемы, расположенные между 56°51′ и 58°22′ с.ш., 35°55′ и 38°25′ в.д. в лесной зоне в подзонах от хвойно-широколиственных лесов до южной тайги, представляют собой головную часть волжского каскада. Русловые Иваньковское и Угличское водохранилища характеризуются сезонным регулированием стока и высокой интенсивностью водообмена (10.6 и 10.1 год–1), озеровидное Рыбинское – многолетним регулированием стока и замедленным водообменном (1.9 год–1) (Экологические…, 2001). Водохранилища различаются по морфометрическим и гидрологическим параметрам (табл. 1).
Таблица 1.
Морфометрические характеристики водохранилищ Верхней Волги (hср – средняя глубина, S1 – площадь акватории, S2 – площадь водосборного бассейна по: (Экологические…, 2001)) и абиотические условия в период исследования
| Год | Температура, °С | Прозрачность, м | Цветность, град | Nобщ, мг/л | Pобщ, мкг/л | Nобщ/Pобщ, мг/мг |
|---|---|---|---|---|---|---|
| Иваньковское вдхр. (hср 3.4 м, S1 327 км2, S2 41 000 км2) | ||||||
| 2015 | 21.5 ± 1.4 (12) | 1.1 ± 0.02 (17) | 39 ± 4 (34) | 1.35 ± 0.31 (68) | 76 ± 7 (27) | 18 ± 4 |
| 2016 | 22.8 ± 1.4 (11) | 0.8 ± 0.02 (35) | 60 ± 6 (33) | 1.33 ± 0.08 (17) | 95 ± 5 (16) | 14 ± 1 |
| 2017 | 22.3 ± 0.6 (22) | 0.9 ± 0.02 (21) | 70 ± 4 (18) | 1.11 ± 0.12 (33) | 90 ± 10 (34) | 14 ± 2 |
| 2018 | 22.6 ± 0.2 (23) | 0.9 ± 0.03 (33) | 55 ± 8 (46) | 1.54 ± 0.07 (17) | 95 ± 6 (23) | 16 ± 1 |
| Угличское вдхр. (hср 5.0 м, S1 249 км2, S2 60 022 км2) | ||||||
| 2015 | 20.9 ± 0.3 (4) | 0.8 ± 0.02 (18) | 43 ± 1 (6) | 1.15 ± 0.07 (16) | 92 ± 5 (17) | 13 ± 1 |
| 2016 | 23.7 ± 0.2 (2) | 0.8 ± 0.01 (13) | 43 ± 1 (6) | 1.26 ± 0.04 (8) | 106 ± 3 (7) | 12 ± 1 |
| 2017 | 21.7 ± 0.2 (2) | 0.9 ± 0.01 (8) | 69 ± 3 (13) | 1.11 ± 0.12 (29) | 85 ± 3 (10) | 13 ± 2 |
| 2018 | 23.1 ± 0.1 (2) | 0.9 ± 0.02 (18) | 48 ± 3 (14) | 1.50 ± 0.10 (19) | 90 ± 6 (19) | 17 ± 2 |
| Рыбинское вдхр. (hср 5.6 м, S1 4550 км2, S2 150 500 км2) | ||||||
| 2015 | 16.6 ± 0.4 (5) | 0.8 ± 0.01 (5) | 43 ± 2 (10) | 1.08 ± 0.06 (12) | 146 ± 9 (14) | 8 ± 1 |
| 2016 | 20.2 ± 0.2 (1) | 0.8 ± 0.02 (11) | 46 ± 4 (17) | 1.15 ± 0.09 (17) | 51 ± 12 (50) | 26 ± 5 |
| 2017 | 17.7 ± 0.2 (2) | 1.1 ± 0.02 (9) | 77 ± 4 (11) | 0.71 ± 0.06 (20) | 71 ± 9 (28) | 11 ± 2 |
| 2018 | 19.9 ± 0.1 (1) | 1.1 ± 0.01 (7) | 57 ± 2 (9) | 0.82 ± 0.07 (18) | 68 ± 9 (28) | 13 ± 3 |
Примечание. Nобщ – общий азот; Pобщ – общий фосфор. Здесь и в табл. 2 даны средние величины со стандартной ошибкой, в скобках коэффициент вариации, %.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Годы наблюдения, которые в многолетнем ряду характеризовались как теплые, различались региональными погодными условиями с количеством осадков от ~97% нормы в 2015 г. до 135% нормы в 2017 г. В августе 2015 и 2018 гг. преобладали ясные и малооблачные дни, в 2016 и 2017 гг. – пасмурная погода (Доклад…, 2016, 2017, 2018, 2019). Уровень двух верхних водохранилищах менялся незначительно, а в Рыбинском водохранилище был минимальным в 2015 г., высоким в 2016 и 2017 гг. и более низким в 2018 г. (соответственно 100.32, 101.15, 101.63 и 100.93 м БС в среднем за май-октябрь)11. Средняя за эти месяцы температура воздуха в районе Рыбинского водохранилища в 2017 г. (12.1°С) была значительно ниже по сравнению с тремя остальными годами (13.5–14.6°С)22. Однако в августе 2017 г. прогрев водной толщи достиг обычных значений, а более низкая температура отмечена в 2015 г. В целом температура воды была выше средних многолетних показателей для августа (Экологические…, 2001) Прозрачность и цветность воды характеризовались типичными для водохранилищ величинами, лишь в дождливом 2017 г. цветность была высокой (табл. 1). Коэффициенты вариации не превышали 23% для температуры, 35% для прозрачности воды и 46% для цветности. Эти показатели наиболее изменчивы в Иваньковском водохранилище.
Концентрации БЭ в водохранилищах Верхней Волги изменялись в близких пределах. Среднее содержание N-N${\text{O}}_{3}^{ - }$ составляло 0.01–0.14 мг/л, было минимальным в 2015 г. и лишь в Рыбинском водохранилище в 2018 г. увеличивалась до 0.30 мг/л. Пониженное количество Nобщ повсеместно отмечено в 2017 г., самое высокое – в Иваньковском и Угличском водохранилищах в 2018 г. (табл. 1). В двух верхних водохранилищах ~90% значений равнялись 1–2 мг/л, в Рыбинском в одинаковых пропорциях представлены величины 0.5–1 и 1–2 мг/л (рис. 2). В отдельные периоды по акватории каждого водохранилища концентрации Nобщ различались не более чем вдвое. Лишь за счет максимальной величины 4.1 мг/л в Волжском плесе Иваньковского водохранилища в 2015 г. и минимальной 0.59 мг/л в устье р. Дубна (Угличское водохранилище в 2017 г.) различия возрастали. Среднее содержание Nобщ, которое незначительно менялось в двух верхних водохранилищах, снижалось от Иваньковского водохранилища к Рыбинскому, причем в 2017 и 2018 гг. существенно, в 1.6–1.9 раза (табл. 1). Средняя концентрация P-P${\text{O}}_{4}^{{3 - }}$ в Иваньковском и Угличском водохранилищах увеличивалась от 27 мкг/л в 2015 г. до 40–63 мкг/л в остальные годы, а в Рыбинском во все сроки составляла 30–40 мкг/л. Предельные концентрации Pобщ в каждом водохранилище в основном различались в 1.2–2.5 раза. Лишь в двух случаях (2016 г. в Рыбинском и 2017 г. в Иваньковском) при низкой (~30 мкг/л) концентрации на одной из станций разница увеличивалась до 3–5-кратной. Более 60% величин Pобщ в Иваньковском и Угличском и 45% в Рыбинском водохранилище ограничены диапазоном 50–100 мкг/л, треть величин превышает 100 мкг/л. В Рыбинском водохранилище часто встречаются и более низкие значения (рис. 2). Средняя концентрация Pобщ в Иваньковском и Угличском водохранилищах во все года изменялась от 80 до 110 мкг/л, в Рыбинском она увеличивалась в 2015 г. и снижалась в августе остальных лет, причем в 2016 г. существенно (табл. 1). Отношение Nобщ/Pобщ в каждом водохранилище колебалось от минимальных величин 5–8 (по массе) до максимальных 30–57. В Иваньковском и Угличском водохранилищах преобладали величины 10–15, в Рыбинском равными пропорциями представлен весь их диапазон от <10 до >15. Среднее отношение Nобщ/Pобщ в основном менялось в пределах 11–18, а в Рыбинском водохранилище снижалось до 8 в 2015 г. и возрастало до 25 в 2016 г. (табл. 1). Судя по коэффициентам вариации (16–29%), изменчивость концентраций Nобщ, Pобщ и их соотношения в водохранилищах Верхней Волги невелика. Лишь для Pобщ в Рыбинском водохранилище, а также Nобщ/Pобщ в Рыбинском и Иваньковском водохранилищах она увеличивается до умеренной (Cv 50–66%).
Рис. 2.
Содержание Хл а (а, в, д, ж) и БЭ (б, г, е, з) в Иваньковском (ст. 1–11), Угличском (ст. 12–20) и Рыбинском (ст. 21–26) водохранилищах в 2015 (а, б) , 2016 (в, г), 2017 (д, е), 2018 (ж, з) годах. 1 – ХлCyan, 2 – ХлBac, 3 – ХлChl, 4 – Nобщ, 5 – Робщ.
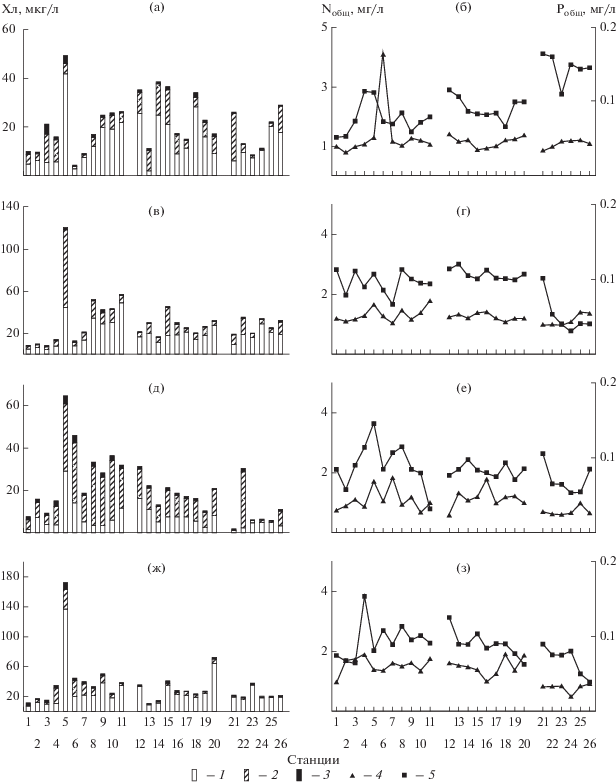
Содержание Хл а в Иваньковском водохранилище в разные годы варьировало от минимальных 7–9 мкг/л до максимальных 49–172 мкг/л, в Угличском водохранилище от 10–16 до 31–72 мкг/л, в Рыбинском водохранилище – от 2 до 30 мкг/л в 2017 г. и от 9–19 до 29–37 мкг/л в остальные годы (рис. 2). Распределение фитопланктона (Хл a) по акватории водохранилищ в большинстве случаев характеризовалось умеренной степенью неоднородности (Cv < 70%), лишь в Иваньковском (2016, 2018 гг.) и в Рыбинском (2017 г.) водохранилищах оно было более дискретным (Cv= 87–100%). Во всех водохранилищах прослеживались межгодовые различия Хл а, среднее содержание которого менялось от минимальных 10.3–19.9 мкг/л до максимальных 27.7–48.0 мкг/л. Самая низкая величина, полученная в 2015 г. в Иваньковском водохранилище и в 2017 г. в двух других, достоверно отличалась от показателей остальных лет (t = 1.9–6.3). Различия были незначимы для величин, полученных в 2016 и 2018 гг. в Угличском водохранилище, а также в 2015 и 2018 гг. в Рыбинском. В суммарном фонде Хл а в основном преобладал ХлCyan (рис. 2, табл. 2), средний вклад которого составлял 53–92% в Рыбинском водохранилище, 40–85% в Угличском и 28–67% в Иваньковском. Минимальные величины повсеместно отмечены летом прохладного 2017 г. Средняя доля Хлbac изменялась в пределах 6–40, 10–55 и 25–60% соответственно при максимальной в августе 2017 г. Доля ХлChl составляла 1–6% ƩХл в Рыбинском и Угличском водохранилищах и увеличивается до 9–10% в Иваньковском.
Таблица 2.
Содержание суммарного Хл а и хлорофилла основных отделов водорослей в водохранилищах Верхней Волги
| Водохранилище | Год | Суммарный Хл а, мкг/л |
ХлCyan | ХлBac | ХлChl | |||
|---|---|---|---|---|---|---|---|---|
| мкг/л | % | мкг/л | % | мкг/л | % | |||
| Иваньковское | 2015 | 19.9 ± 3.9 (69) | 15.9 ± 4.6 | 59.0 ± 7.4 | 7.6 ± 3.4 | 27.3 ± 5.2 | 4.7 ± 3.8 | 9.3 ± 3.4 |
| 2016 | 32.2 ± 8.8 (95) | 19.7 ± 4.5 | 51.8 ± 6.9 | 17.2 ± 7.3 | 30.0 ± 4.1 | 5.3 ± 4.5 | 6.9 ± 3.1 | |
| 2017 | 28.1 ± 4.7 (58) | 11.6 ± 4.1 | 27.6 ± 3.6 | 18.2 ± 2.9 | 59.6 ± 5.5 | 4.7 ± 2.8 | 10.1 ± 2.3 | |
| 2018 | 48.0 ± 12.0 (87) | 34.1 ± 10.2 | 66.8 ± 5.0 | 10.9 ± 2.4 | 25.1 ± 4.8 | 2.9 ± 0.5 | 9.3 ± 2.5 | |
| Угличское | 2015 | 25.3 ± 4.1 (42) | 16.6 ± 3.5 | 61.8 ± 7.2 | 8.0 ± 1.4 | 34.9 ± 6.9 | 0.8 ± 0.1 | 3.4 ± 0.6 |
| 2016 | 27.6 ± 3.3 (30) | 18.2 ± 1.7 | 68.3 ± 5.1 | 8.6 ± 2.7 | 29.0 ± 4.9 | 0.8 ± 0.2 | 2.8 ± 0.5 | |
| 2017 | 19.0 ± 2.3 (32) | 7.9 ± 1.5 | 39.9 ± 3.4 | 10.2 ± 0.9 | 55.3 ± 3.2 | 0.9 ± 0.1 | 4.8 ± 0.6 | |
| 2018 | 30.3 ± 7.0 (60) | 26.5 ± 6.4 | 85.1 ± 3.0 | 2.6 ± 0.6 | 9.5 ± 2.1 | 1.5 ± 0.3 | 6.1 ± 1.2 | |
| Рыбинское | 2015 | 18.4 ± 3.9 (46) | 12.1 ± 2.6 | 72.0 ± 11.6 | 6.2 ± 3.4 | 27.4 ± 11.5 | 0.1 ± 0.1 | 0.6 ± 0.2 |
| 2016 | 27.7 ± 3.2 (25) | 19.0 ± 2.9 | 68.0 ± 7.7 | 8.1 ± 2.2 | 29.7 ± 7.4 | 0.7 ± 0.2 | 2.3 ± 0.5 | |
| 2017 | 10.3 ± 4.7 (100) | 3.5 ± 0.7 | 53.4 ± 13.7 | 6.2 ± 4.7 | 40.0 ± 13.7 | 0.6 ± 0.2 | 6.6 ± 0.9 | |
| 2018 | 22.6 ± 3.2 (31) | 21.1 ± 3.3 | 92.4 ± 2.7 | 1.3 ± 0.4 | 6.4 ± 2.4 | 0.3 ± 0.0 | 1.6 ± 0.3 | |
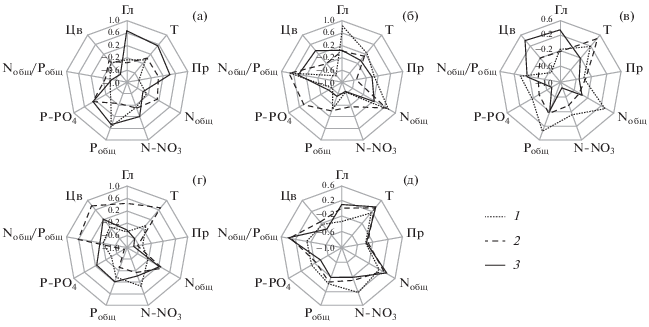
Теснота и направленность связи содержания Хл а и БЭ менялась в разные годы (рис. 3). Парный корреляционный анализ показывает, что чаще всего Хл а достоверно коррелировал с Pобщ и P-P${\text{O}}_{4}^{{3 - }}$ (в 9 случаях из 12), Nобщ, N-N${\text{O}}_{3}^{ - }$, и отношением Nобщ/Pобщ (по 6 случаев). Наиболее тесно (r > 0.70) в Иваньковском водохранилище он связан с цветностью, Nобщ и отношением Nобщ/Pобщ (2016 г.), в Угличском – с Nобщ/Pобщ (2016 г.), P-P${\text{O}}_{4}^{{3 - }}$ (2017 г.) и цветностью (2018 г.), в Рыбинском – с содержанием N-N${\text{O}}_{3}^{ - }$, P-P${\text{O}}_{4}^{{3 - }}$ (2017 г.) и прозрачностью (2018 г.). По совокупности четырехлетних данных в каждом водохранилище выявлена лишь умеренная связь содержания Хл а с абиотическими факторами, включая БЭ (r = 0.30–0.70), а корреляционный граф выглядит сходным (рис. 3д). Во всех водохранилищах фитопланктон оказывает отрицательное влияние на прозрачность воды. В Рыбинском и Угличском водохранилищах содержание Хл а прямо зависит от температуры и отношения Nобщ/Pобщ, в Рыбинском – от концентрации Nобщ, в Иваньковском отмечена обратная зависимость от глубины станции.
Рис. 3.
Коэффициенты корреляции Хл а с факторами среды в Иваньковском (1), Угличском (2) и Рыбинском (3) водохранилищах в 2015–2018 гг. (а–г, соответственно) и за весь период наблюдения (д). (Гл – глубина станций, Т – температура воды, Пр – прозрачность, Цв – цветность).
Множественный корреляционный анализ показывает, что влияние БЭ объясняет невысокую долю вариации суммарного Хл а: 14% в Иваньковском водохранилище, 34% в Угличском и 25% в Рыбинском, а также соответственно 21, 12 и 46% вариации ХлCyan, 13, 27 и 23% вариации ХлBac. Значимые коэффициенты регрессии (t > 1.96) получены только для Nобщ, Pобщ и их соотношения в Угличском водохранилище. При введении в регрессионную модель температуры, прозрачности, цветности воды и глубины станции коэффициенты детерминации увеличиваются. С совокупностью всех рассматриваемых переменных связан 48% вариации суммарного Хл а в Иваньковском водохранилище, 57% в Угличском и 59% в Рыбинском, а также соответственно 54, 63 и 69% вариации ХлCyan, 30, 26 и 52% вариации ХлBac. Коэффициенты регрессии значимы для температуры и прозрачности воды в Угличском водохранилище, прозрачности, цветности, содержания минеральных форм азота и фосфора в Иваньковском.
Если рассматривать три водохранилища как единую систему, то, используя многомерную статистику, можно выделить три ведущих фактора, которые объединяют разные характеристики водохранилищ и включают >80% общей дисперсии. Величины факторных нагрузок >0.70 свидетельствуют о том, что с фактором 1 тесно связаны показатели открытости, площади водоемов и водосборных бассейна, показатели удельного водосбора и коэффициенты условного водообмена. Для ХлBac в этот перечень добавляется прозрачность и Nобщ. Фактор 2 отражает влияние Pобщ и отношения Nобщ/Pобщ. С фактором 3 тесно коррелируют цветность, содержание суммарного Хл а и ХлCyan, а для всего автотрофного планктона и цианопрокариот – Nобщ (табл. 3).
Таблица 3.
Факторные нагрузки (абсолютные величины) содержания хлорофилла и абиотических параметров водохранилищ Верхней Волги по данным факторного анализа
| Показатель | Суммарный Хл а | ХлCyan | ХлBac | ||||||
|---|---|---|---|---|---|---|---|---|---|
| фактор 1 | фактор 2 | фактор 3 | фактор 1 | фактор 2 | фактор 3 | фактор 1 | фактор 2 | фактор 3 | |
| Хлорофилл | 0.39 | 0.22 | 0.72 | 0.21 | 0.21 | 0.83 | 0.57 | 0.04 | 0.53 |
| S1 | 0.98 | 0.00 | 0.19 | 0.99 | 0.02 | 0.13 | 0.98 | 0.01 | 0.03 |
| S2 | 0.95 | 0.03 | 0.22 | 0.97 | 0.00 | 0.15 | 0.98 | 0.03 | 0.08 |
| S1/hср | 0.98 | 0.01 | 0.18 | 0.99 | 0.02 | 0.13 | 0.98 | 0.01 | 0.02 |
| S2/S1 | 0.85 | 0.09 | 0.06 | 0.84 | 0.08 | 0.04 | 0.78 | 0.04 | 0.10 |
| Квод | 0.97 | 0.01 | 0.20 | 0.98 | 0.01 | 0.14 | 0.98 | 0.02 | 0.05 |
| Глубина станции | 0.57 | 0.35 | 0.57 | 0.59 | 0.40 | 0.51 | 0.70 | 0.41 | 0.21 |
| Температура воды | 0.66 | 0.63 | 0.20 | 0.67 | 0.61 | 0.21 | 0.69 | 0.62 | 0.02 |
| Прозрачность | 0.51 | 0.31 | 0.61 | 0.56 | 0.38 | 0.46 | 0.72 | 0.35 | 0.19 |
| Цветность | 0.10 | 0.00 | 0.73 | 0.05 | 0.06 | 0.78 | 0.12 | 0.02 | 0.94 |
| Nобщ | 0.58 | 0.15 | 0.74 | 0.62 | 0.07 | 0.72 | 0.78 | 0.10 | 0.50 |
| Pобщ | 0.08 | 0.90 | 0.37 | 0.11 | 0.93 | 0.26 | 0.20 | 0.92 | 0.23 |
| Nобщ/Pобщ | 0.07 | 0.91 | 0.32 | 0.03 | 0.87 | 0.38 | 0.05 | 0.88 | 0.30 |
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Водохранилища Верхней Волги расположены в пределах одной географической зоны, что определяет сходство климатических условиях и природные особенностей водосборного бассейна. Исходя из этого, можно предполагать, что существенное влияние на динамику фитопланктона и режим БЭ оказывают региональные факторы и внутриводоемные процессы, которые, вероятно, должны различаться вследствие различной антропогенной нагрузки, морфометрических и гидрологических особенностей водохранилищ, а также в зависимости от погодных условий в годы исследования. Судя по невысоким коэффициентам вариации температуры, прозрачности, цветности воды и содержания БЭ, в периоды наблюдения фитопланктон водохранилищ развивался при стабильных термических и гидрооптических и гидрохимических условиях. Межгодовые изменения температуры обусловлены региональными климатическими особенностями. Повышенная цветность в 2017 г. связана с максимальным за годы наблюдения летним количеством осадков, составившим 135% нормы (Доклад…, 2018) и повлекшим вынос с водосборной площади окрашенных органических веществ.
Содержание Хл а, которое меняется в широком диапазоне, в целом типично для летнего максимума фитопланктона Верхней Волги (Минеева, 2016; Экологические…, 2001; Mineeva, 2019). Неравномерное распределение по акватории водоемов обусловлено повышенными концентрациями пигмента на локальных участках: в Иваньковском водохранилище – в Шошинском плесе и нижней части; в Угличском водохранилище – на среднем участке, принимающем воды притоков – рек Медведица, Нерль, Кашинка и/или в верхней части, куда поступают воды из Иваньковского водохранилища; в Рыбинском водохранилище в штилевую погоду – на центральных станциях Главного плеса. По среднему содержанию Хл а водохранилища относятся к водоемам эвтрофного типа и только Рыбинское водохранилище в прохладном 2017 г. характеризовалось как мезотрофное.
В годы с разными температурными условиями отмечены существенные различия таксономического состава фитопланктона, косвенное представление о котором дают данные по содержанию Хл а основных отделов водорослей. При смешанном составе альгоценозов, типичном для волжских водохранилищ, в летнем планктоне обычно преобладают цианопрокариоты. В последние годы в условиях глобального потепления наблюдается увеличение их обилия и продолжительности вегетации в Рыбинском водохранилище (Корнева, 2015; Минеева, 2016). Хлорофилл этой группы водорослей преобладал в фонде Хл а в 2015, 2016 и 2018 гг., уступив лидирующее положение ХлBac в прохладном 2017 г.
Содержание основных БЭ в водохранилищах Верхней Волги, как и в предыдущие годы (Экологические…, 2001; Структура…, 2018), характеризуется высокими величинами, которые типичны для эвтрофных водоемов. Концентрации минеральных форм азота и фосфора превосходят порог, необходимый для развития водорослей (Reynolds, 2012). При небольшой вариабельности общих и минеральных форм БЭ в отдельные сроки наблюдения, фитопланктон на всей акватории водохранилищ в достаточной степени должен быть обеспечен минеральным питанием.
При анализе связи между содержанием Хл а и БЭ в полимиктических водоемах используют эмпирические уравнения, рассчитанные для летних показателей в исходном или логарифмическом виде (см. сводку: Китаев, 2007) и отражающие стохастические связи между основными параметрами качества воды. Однако универсальные зависимости, которые, как правило, получены при крупномасштабных обобщениях (Eutrophication…, 1982), в конкретных случаях могут не проявляться. Последнее обусловлено спецификой водоемов, определяющим влиянием других факторов, наличием “шумов” и помех, вероятной сменой лимитирующего БЭ в течение сезона, сезонной сукцессией фитопланктона (Жукова, 2001). Такие связи менее выражены и редко значимы для водохранилищ – переходных от речного к озерному типу экосистем, которые, как и реки (Vannote et al., 1980), не испытывают дефицита БЭ.
Характер зависимости Хл а от содержания БЭ может меняться в течение вегетационного сезона (Hao et al., 2014). Согласно предыдущим исследованиям на Рыбинском водохранилище, связь между ними прослеживалась лишь в отдельные периоды наблюдения и чаще всего была лишь умеренной (Минеева, Разгулин, 1995; Степанова и др., 2012; Структура…, 2018). В настоящей работе для водохранилищ Верхней Волги также получены невысокие коэффициенты корреляции между содержанием Хл а и БЭ. После логарифмического преобразования данных теснота связи лишь незначительно увеличилась в Иваньковском водохранилище, стала ниже в Угличском и не изменилась в Рыбинском (во всех случаях R2 ≤ 0.1).
Характер связи Хл а с содержанием БЭ менялся в разные годы. Отрицательная корреляция, свидетельствующая о потреблении минеральных форм азота и фосфора, выявлена в 2016 и 2017 гг. в Рыбинском и Иваньковском водохранилищах, минерального фосфора – в 2018 г. в Угличском. Предположительный дефицит фосфора (положительная корреляция с Хл а) отмечен в 2015 г. в Рыбинском и Иваньковском водохранилищах, дефицит азота – в 2016 г. в Угличском и Иваньковском. В период максимального прогрева водной толщи содержание Хл а слабо зависит от температуры воды. Фитопланктон оказывает умеренное отрицательное влияние на формирование подводного светового режима всех водохранилищ, где взвешенное вещество содержит планктонную и терригенную компоненты.
Влияние БЭ на содержание Хл а чаще всего обсуждают на фоне отношения Nобщ/Pобщ, которое считают показателем обеспеченности фитопланктона минеральным питанием. Низкие величины Nобщ/Pобщ свидетельствуют о дефиците азота, высокие – о дефиците фосфора, промежуточные, близкие к соотношению элементов в клетках, – об отсутствии биогенного лимитирования. В литературе для этих ситуаций приводят различные пограничные значения Nобщ/Pобщ (Downing, 1992; Guildford, Hecky, 2000; Søndergaard et al., 2017), при анализе настоящих данных за основу взяты Nобщ/Pобщ <10 и >15 (по массе) (Sakamoto, 1966). Средние для водохранилищ величины Nобщ/Pобщ указывают на отсутствие лимитирования фитопланктона Верхней Волги БЭ, что согласуется с высокими концентрациями последних. Лишь в Рыбинском водохранилище в августе 2015 г. отмечен возможный дефицит азота и в 2016 г. – дефицит фосфора, который также выявлен в 2015 г. в Иваньковском водохранилище.
Отсутствие тесной статистической зависимости Хл а от БЭ свидетельствует о сложном и многокомпонентном характере биогенного контроля за развитием водорослей. Маскировать прямое влияние БЭ может различная потребность в минеральном питании видов, входящих в состав альгоценозов (Suttle, Harrison, 1988); совместное влияние (ко-лимитирование) азота и фосфора (Müller, Mitrovic, 2015; Pearl et al., 2011); конкуренция за БЭ между водорослями и бактериями (Brett et al., 1999); изменяющийся в течение вегетационного сезона пресс растительноядного зоопланктона (Kalff, 2002). Доступность фосфора снижается в окрашенных водах в присутствии гуминовых веществ, связывающих фосфат-ионы (Palsson, Graneli, 2004), а также в присутствии железа (Tang et al., 2019), содержание которого в природных водах увеличивается с увеличением цветности (Китаев, 2007). При высоком содержании БЭ и/или при низкой прозрачности воды рост водорослей может быть лимитирован не БЭ, а светом (Burson et al., 2018; Kalff, 2002). Причиной слабой связи между Хл а и БЭ может быть небольшой диапазон их концентраций, в пределах которого зависимость не прослеживается (Huszar et al., 2006). Все перечисленные аспекты присущи водохранилищам Верхней Волги.
Общий запас БЭ в водоеме формируется за счет притока с водосборной территории (внешняя нагрузка) и разнонаправленных внутриводоемных процессов, включающих поступление из донных осадков (внутренняя нагрузка), рециклинг и др. Роль биотического рециклинга БЭ в водохранилищах Верхней Волги, вероятно, сопоставима. Внешним источником БЭ во всех трех водохранилищах служит речной сток – основная составляющая водного баланса (Экологические…, 2001). Показателем внешней биогенной нагрузки может быть коэффициент удельного водосбора (S2/S1), величина которого в Угличском водохранилище (241) в 1.9 раза выше, чем в Иваньковском, в 7.3 раза выше, чем в Рыбинском. По аналогии с Нарочанскими озерами (Zhukova, 2013), можно предположить, что Угличское водохранилище испытывает максимальную внешнюю нагрузку, повлекшую повышение его трофического статуса в последние годы (Mineeva, 2019). Внутренняя нагрузка определяется выделением БЭ тонкодисперсными донными отложениями, площадь которых занимает 34% общей в Иваньковском водохранилище, 26% в Угличскоми и 28% в Рыбинском (Законнов и др., 2018; Степанова, Литвинов, 2019). В Рыбинском водохранилище взмучивание седиментов, обеспечивающее дополнительно поступление БЭ, происходит при частом ветровом перемешивании мелководной открытой акватории. Следовательно, погодные условия (ветровое воздействие) могут быть одним из факторов, влияющих на развитие фитопланктона и, соответственно, – вариации трофического статуса водохранилища. Показатель открытости S1/hср для Рыбинского водохранилища равен 812 и в 9–16 раз выше, чем в двух русловых водохранилищах, в которых, по-видимому, фитопланктон развивается при более стабильной внутренней нагрузке.
Теснота и направленность связи Хл а с параметрами среды меняется в зависимости от конкретных условий в период наблюдения. В 2015 г. при минимальном количестве осадков и самом низком уровне получены лишь невысокие коэффициенты корреляции Хл а со всеми факторами, в 2016 г. – тесная связь с отношением Nобщ/Pобщ, в наиболее влажном 2017 г. при высоком уровне воды – с минеральными формами азота и фосфора, в 2018 г. – с цветностью и прозрачностью. Множественный корреляционный анализ подтверждает высокую значимость гидрологических показателей как для развития всего автотрофного планктона водохранилищ Верхней Волги, так и его основных представителей – диатомовых водорослей и цианопрокариот. Тот факт, что по совокупности четырехлетних данных в каждом водохранилище выявлена лишь умеренная связь содержания Хл а с абиотическими показателями, включая БЭ, свидетельствует о существенном влиянии на развитие фитопланктона климата, гидрологических условий, динамики вод, интенсивности водообмена, особенностей водосборной территории. Последнее, в частности, подтверждается результатами факторного анализа.
Выводы. В летний период 2015–2018 гг. содержание Хл а было типичным для летнего максимума фитопланктона Верхней Волги. По среднему содержанию Хл а водохранилища относятся к водоемам эвтрофного типа и только Рыбинское водохранилище в прохладном 2017 г. характеризовалось как мезотрофное. ХлCyan преобладал в фонде Хл а в 2015, 2016 и 2018 гг., уступив лидирующее положение ХлBac в прохладном 2017 г. Фитопланктон водохранилищ развивался при стабильных термических и гидрооптических условиях.
Содержание основных БЭ также отражают эвтрофный статус водохранилищ. Концентрации минеральных форм азота и фосфора превосходят порог, необходимый для развития водорослей. При небольшой вариабельности общих и минеральных форм БЭ в отдельные сроки наблюдения, фитопланктон на всей акватории водохранилищ в достаточной степени обеспечен минеральным питанием. Средние величины Nобщ/Pобщ указывают на отсутствие биогенного лимитирования фитопланктона, лишь в Рыбинском водохранилище в августе 2015 г. отмечен возможный дефицит азота и в 2016 г. – дефицит фосфора, который также выявлен в 2015 г. в Иваньковском водохранилище.
Невысокие коэффициенты корреляции между содержанием Хл а и БЭ свидетельствуют о сложном и многокомпонентном характере их влияния на водоросли. Множественный корреляционный анализ подтверждает значимость гидрологических показателей для развития автотрофного планктона водохранилищ Верхней Волги.
Список литературы
Бикбулатов Э.С. 1974. О методе определения общего фосфора в природных водах // Гидрохимические материалы. Т. 60 С. 167.
Второй оценочный доклад Росгидромета об изменениях климата и их последствиях на территории Российской Федерации. Общее резюме. 2014. Москва: Росгидромет.
Гапеева М.В., Разгулин С.М., Скопинцев Б.А. 1984. Ампульный персульфатный метод определения общего азота в природных водах // Гидрохимические материалы. Т. 87. С. 67.
Гольд В.М., Гаевский Н.А. Шатров И.Ю. и др. 1986. Опыт использования флуоресценции для дифференциальной оценки содержания хлорофилла у планктонных водорослей // Гидробиол. журн. Т. 22. № 3. С. 80.
Доклад об особенностях климата на территории Российской Федерации за 2015 год. 2016. Москва: Росгидромет.
Доклад об особенностях климата на территории Российской Федерации за 2016 год. 2017. Москва: Росгидромет.
Доклад об особенностях климата на территории Российской Федерации за 2017 год. 2018. Москва: Росгидромет.
Доклад об особенностях климата на территории Российской Федерации за 2018 год. 2019. Москва: Росгидромет.
Жукова Т.В. 2001. Потоки фосфора и азота в пограничном слое “дно–вода” и их роль в функционировании полимиктических озер (на примере Нарочанских озер). Автореф. дис. … докт. биол. наук. Минск.
Законнов В.В., Законнова А.В., Цветков А.И., Шерышева Н.Г. 2018. Гидродинамические процессы и их роль в формировании донных осадков водохранилищ Волжско-Камского каскада // Труды ИБВВ РАН. Вып. 81(84). С. 35.
Китаев С.П. 2007. Основы лимнологии для гидробиологов и ихтиологов. Петрозаводск: КарНЦ РАН.
Корнева Л.Г. 2015. Фитопланктон водохранилищ бассейна Волги. Кострома: Костромской печатный дом.
Минеева Н.М. 2004. Растительные пигменты в воде волжских водохранилищ. Москва: Наука.
Минеева Н.М. 2016. Сезонная и межгодовая динамика хлорофилла в планктоне Рыбинского водохранилища по данным флуоресцентной диагностики // Систематика, морфология и экология водных растений. Труды Института биологии внутренних вод им. И.Д. Папанина РАН. Вып. 76(79). Ярославль: Филигрань. С. 75.
Минеева Н.М., Разгулин С.М. 1995. О влиянии биогенных элементов на содержание хлорофилла в Рыбинском водохранилище // Водные ресурсы. Т. 22. № 2. С. 218.
Пырина И.Л. 2000. Многолетняя динамика и цикличность межгодовых колебаний содержания хлорофилла в Рыбинском водохранилище // Озерные экосистемы: биологические процессы, антропогенная трансформация, качество воды. Матер. международ. науч. конф. Минск: Белорусский гос. ун-т. С. 375.
Семенов А.Д. 1977. Практическое руководство по химическому анализу вод суши. Ленинград: Гидрометеоиздат.
Степанова И.Э. 2018. Гидрохимические показатели воды Угличского и Иваньковского водохранилищ // Экологическая химия. Т. 27. № 3. С. 156.
Степанова И.Э., Литвинов А.С. 2019. Водный и химический балансы в Рыбинском водохранилище // Экологическая химия. Т. 28. № 5. С. 280.
Степанова И.Э., Пырина И.Л., Бикбулатов Э.С., Бикбулатова Е.М. 2012. Анализ связи развития фитопланктона с содержанием биогенных элементов в Рыбинском водохранилище // Органическое вещество и биогенные элементы во внутренних водоемах и морских водах. Петрозаводск: КарНЦ. С. 129.
Структура и функционирование экосистемы Рыбинского водохранилища в начале XXI века. 2018. Москва: РАН.
Экологические проблемы Верхней Волги. 2001. Ярославль: Ярослав. гос. техн. ун-т.
Brett M.T., Lubnow F.S., Villar-Argaiz M. et al. 1999. Nutrient control of bacterioplankton and phytoplankton dynamics // Aquatic Ecology V. 33. № 2. P. 135. https://doi.org/10.1023/A:1009998915078
Burson A., Stomp M., Greenwell E. et al. 2018. Competition for nutrients and light: testing advances in resource competition with a natural phytoplankton community // Ecology. V. 99. № 5. P. 1108. https://doi.org/10.1002/ecy.2187
Downing J.A. 1992. The nitrogen: phosphorus relationship in lakes // Limnol., Oceanogr. V. 37. № 5. P. 936. https://doi.org/10.4319/lo.1992.37.5.0936
Eutrophication of waters. Monitoring, assessment and control. 1982. Paris: OECD.
Guildford S.J., Hecky R.E. 2000. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: Is there a common relationship? // Limnol., Oceanogr. V. 45. № 6. P. 1213. https://doi.org/10.4319/lo.2000.45.6.1213
Hallstan S., Trigal C., Johansson K.S.L., Johnson R.K. 2013. The impact of climate on the geographical distribution of phytoplankton species in boreal lakes // Oecologia. V. 173. № 4. P. 1625. https://doi.org/10.1007/s00442-013-2708-6
Hao Y., Yu R., Zhang U., Yanget H. 2014. The relationship of chlorophyll-a, total nitrogen and total phosphorus in Wuliangsuhai Lake // Advanced Materials Research. V. 955–959. P. 1393. https://doi.org/10.1186/s13750-017-0097-8
Huszar V.L.M., Caraco N.F., Roland F., Cole J. 2006. Nutrient-chlorophyll relationships in tropical–subtropical lakes: do temperate models fit? // Biogeochemistry. V. 79. Issue 1–2. P. 239. https://doi.org/10.1007/s10533-006-9007-9
Jeppesen E., Sondergaard M., Jensen J.P. et al. 2005. Lake responses to reduced nutrient loading – an analysis of contemporary long-term data from 35 case studies // Freshwater Biol. V. 50. № 9. P. 1747. https://doi.org/10.1111/j.1365-2427.2005.01415.x
Kaiser H.F. 1960. The application of electronic computers to factor analysis // Educational and Psychological Measurement. V. 20. № 1. P. 141. https://doi.org/10.1177/001316446002000116
Kalff J. 2002. Limnology. Inland Water Ecosystems. Upper Saddle River, N. J. : Prentice Hall.
Lewandowska A.M., Boyce D.J., Hofmann M. et al. 2014. Effect of sea surface warming on marine plankton // Ecology Letters. V. 17. № 5. P. 614. https://doi.org/10.1111/ele.12265
Mineeva N.M. 2019. Content of photosynthetic pigments in the Upper Volga Reservoirs (2005–2016) // Inland Water Biology. V. 12. № 2. P. 161. https://doi.org/10.1134/S199508291902010X
Müller S., Mitrovic S.M. 2015. Phytoplankton co-limitation by nitrogen and phosphorus in a shallow reservoir: progressing from the phosphorus limitation paradigm // Hydrobiologia. V. 744. P. 255. https://doi.org/10.1007/s10750-014-2082-3
Palsson C., Graneli W. 2004. Nutrient limitation of autotrophic and mixotrophic phytoplankton in a temperate and tropical humic lake gradient // J. Plankton Research. V. 26. № 9. P. 1005. https://doi.org/10.1093/plankt/fbh089
Paerl H.W., Xu H., McCarthy M.J. et al. 2011. Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): the need for a dual nutrient (N and P) management strategy // Water Research. V. 45. № 5. P. 1973. https://doi.org/10.1016/j.watres.2010.09.018
Pridmore R.D., Vant W.N., Rutherford J.C. 1985. Chlorophyll-nutrient relationships in North Island lakes (New Zealand) // Hydrobiologia. V. 121. № 2. P. 181. https://doi.org/10.1007/BF00008722
Reynolds C.S. 2012. Environmental requirements and habitat preferences of phytoplankton: chance and certainty in species selection // Botanica Marina. V. 55. № 1. P. 1. https://doi.org/10.1515/bot.2011.121
Sakamoto M. 1966. Primary production by phytoplankton community in some Japanese lakes and its dependence on lake depth // Arch. Hydrobiol. V. 62. № 1. P. 1. https://doi.org/10.1371/journal.pone.0095757
Søndergaard M., Lauridsen T.L., Johansson L.S., Jeppesen E. 2017. Nitrogen or phosphorus limitation in lakes and its impact on phytoplankton biomass and submerged macrophyte cover // Hydrobiologia. V. 795. P. 35. https://doi.org/10.1007/s10750-017-3110-x
Stepanova I.E., Bikbulatova E.M. 2016. Impotance of the ratios of the forms of nutrients for assessment of the modern state of Rybinsk reservoir // Byology Bulletin. V. 43. № 10. P. 1401.https://doi.org/10.1134/S1062359016100198
Suttle C.A., Harrison P.J. 1988. Ammonium and phosphate uptake rates, N : P supply ratios, and evidence for N and P limitation in some oligotrophic lakes // Limnol., Oceanogr. V. 33. № 2. P. 186. https://doi.org/10.4319/lo.1988.33.2.0186
Tang Q., Peng L., Yang Y. et al. 2019. Total phosphorus-precipitation and chlorophyll a – phosphorus relationship of lakes and reservoirs mediated by soil iron at regional scale // Water Research. V. 154. P. 136. https://doi.org/10.1016/j.watres.2019.01.038
Vannote R.L., Minshall G.W., Cummins K.W. et al. 1980. The river continuum concept // Can. J. Fish. Auqat. Sci. V. 37. № 1. P. 130. https://doi.org/10.1139/f80-017
Vollenweider R.A. 1979. Das Nachrstoffbelastungskonzept als Grundlage fur den externen in den Eutrophierungsprozess stehender Gewasser und Talsperren // Z. Wasser und Abwasser Forsch. Bd 12. № 2. S. 46.
Xiao W., Liu X., Irwin A.J. et al. 2018. Warming and eutrophication combine to restructure diatoms and dinoflagellates // Water Research. V. 128. № 1. P. 206. https://doi.org/10.1016/j.watres.2017.10.051
Zhukova T.V. 2013. Long-term dynamics of phosphorus in the Narochanskie Lakes and factors determining it // Water Resourses. V. 40. № 5. P. 510. https://doi.org/10.1134/S0097807813050072
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод