

Биология внутренних вод, 2021, № 2, стр. 160-170
Цветение рясковых (Lemnaceae S.F. Gray) в Сибири: биоэкологическая характеристика
О. А. Капитонова a, b, *, С. А. Николаенко c
a Тобольская комплексная научная станция Уральского отделения Российской академии наук
Тобольск, Россия
b Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл, Россия
c Институт проблем освоения Севера Федерального исследовательского центра Тюменский научный центр Сибирского отделения Российской академии наук
Тюмень, Россия
* E-mail: kapoa.tkns@gmail.com
Поступила в редакцию 04.03.2020
После доработки 26.05.2020
Принята к публикации 12.11.2020
Аннотация
На территории Сибири известно произрастание пяти видов сем. Lemnaceae: Lemna gibba, L. minor, L. trisulca, L. turionifera, Spirodela polyrhiza. Дана характеристика их распространения в регионе и биоэкологических особенностей, приведены факты цветения и плодоношения. Показано, что генеративное размножение рясковых – нередкое явление и характерно для всех видов семейства, известных на территории региона. Наиболее часто цветение наблюдается у Lеmna trisulca и L. turionifera, единичные случаи известны для Spirodela polyrhiza. Почти все выявленные в Сибири популяции Lemna gibba содержали цветущие и плодоносящие растения. L. minor имеет ограниченное распространение в Сибири, случаи ее цветения редки. Генеративное размножение рясковых зафиксировано лишь в пресноводных водоемах (соленость 0.1–0.7‰) глубиной 0.1–0.7 м, c температурой поверхностного слоя воды +20–+33 °С, рН 5.96–9.30, не затененных древесно-кустарниковой и высокотравной растительностью.
ВВЕДЕНИЕ
Lemnaceae S.F. Gray – полностью гидрофильное семейство, объединяющее в своем составе погруженные в воду или свободноплавающие на ее поверхности травянистые растения (Landolt, 1986). Согласно современным представлениям, семейство содержит 38 видов, объединенных в пять родов, что подтверждено данными молекулярно-генетических исследований (Les et al., 2002). На территории России известно произрастание 10 видов рясковых, объединенных в три рода (Черепанов, 1995; Лисицына, Папченков, 2000; Щербаков, 2010). В пределах Сибири выявлено пять видов этого семейства: Lemna gibba L., L. minor L., L. trisulca L., L. turionifera Landolt, Spirodela polyrhiza (L.) Schleid. (Власова, 2005; Капитонова, 2019).
Представители семейства являются самыми маленькими в мире цветковыми растениями, достигшими в результате гидрофильной эволюции крайней степени редукции всех своих органов. Их считают неотеническими формами, произошедшими от предков современного тропического рода Pistia (Ростовцев, 1905; Тахтаджян, 1966), который имеет монофилетическое происхождение с Lemnaceae и представляет собой сестринскую группу по отношению к этому семейству (Stockey et al., 1997). Установленное близкое родство рясковых с ароидными (Rothwell et al., 2004; Cabrera et al., 2008) дает основание рассматривать их как подсемейство Lemnoideae в составе сем. Araceae Juss. (Les et al., 2002), что реализовано в рамках последнего варианта системы классификации цветковых растений APG IV (Chase et al., 2016).
Известно, что ряски способны чрезвычайно быстро размножаться вегетативно за счет дочерних фрондов (листецов). Так, время удвоения по сухому веществу для ряски малой составляет 5–6 сут, по количеству листецов 2–3 cут (Лукина, 1977). В то же время сведений о генеративном способе размножения рясковых очень мало. Считалось, что цветение рясковых – явление крайне редкое (Иванова, 1970; Иконников, 1979; Цвелев, 1996; Azer, 2013) и такие случаи специально регистрировали (Иванова, 1970; Жмылёв и др., 1995). По мере накопления данных об этом (Матвеев, 1977; Лукина, 1984; Landolt, 1986; Капитонова, 2000) стало очевидным, что генеративное размножение рясок – “явление обычное, хотя и не ежегодное в пределах одной популяции” (Кузнецов, Заиченко, 1979, с. 28). Однако, если для территории Европы, в том числе европейской части России, случаи цветения этих растений уже описаны, то для азиатской части нашей страны это явление все еще относится к категории малоизученных.
Первые сведения о цветении Lemnaceae на территории Сибири, вероятно, следует отнести к данным П.Н. Крылова (1929), который указывал на один наблюдаемый им случай цветения L. minor и один случай цветения L. trisulсa, однако в обоих случаях без указания местонахождения. В отношении Spirodela polyrhiza автор отмечает, что “c цветами не наблюдалось” (Крылов, 1929, с. 542). В работе В.Ф. Тарасевич (1990) имеются сведения об найденном ею в гербарии Главного ботанического сада им. Н.В. Цицина РАН образце Lemna gibba, собранном П.П. Грищенко в устье р. Тавдa (Тюменская обл.) в августе 1985 г. и содержащем цветущие экземпляры ряски горбатой. Н.И. Науменко (2008) отмечает неоднократное и обильное цветение L. minor и L. trisulca в озерах Курганской обл. и Северного Казахстана, однако без указания конкретных местонахождений. В июне 1998 г. им найдена “цветущая L. turionifera на юге Тюменской обл.” (Науменко, 2008, с. 234). В результате специально проведенных в 2012 г. исследований распространения L. turionifera и L. gibba на территории Ханты-Мансийского автономного округа – Югры (ХМАО-Югра) Н.Л. Панкова (2013) обнаружила в одном из водоемов цветущие растения ряски горбатой, однако вновь без указания конкретного места.
Таким образом, к настоящему времени о цветении рясковых на территории Сибири имеются немногочисленные и в основном не конкретизированные сведения. Изучение этого явления в указанном регионе не имело систематического характера, нет никаких сведений о плодоношении видов Lemnaceae на этой территории, не выяснены факторы, способствующие генеративному размножению рясковых.
Цель работы – выявить и изучить случаи цветения и плодоношения представителей сем. Lemnaceae на территории Сибири и выяснить эколого-фитоценотические условия этих явлений.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Основным материалом для исследования послужили сборы рясок и геоботанические описания авторов на территории Тюменской обл. (включая автономные округа) за 2016–2019 гг. Также изучены гербарные коллекции, охватывающие районы Восточной и Западной Сибири, Северного Казахстана, Средней Азии (TK, TMN, ряд частных гербариев). Всего изучено свыше 175 гербарных образцов рясковых.
При проведении полевых исследований в местах сбора образцов рясок геоботанические описания проводили по общепринятым методам (Миркин и др., 2002). В пределах описываемого фитоценоза измеряли ряд физико-химических параметров биотопа: температуру и рН воды с помощью портативного рН-метра ИТ-1101 в комплекте с электродом ЭКС-10609/7-К80.12, концентрацию растворенных солей – солемером ST 20S, глубину воды – деревянной рейкой с делениями через 5 см. В полевых условиях ряски предварительно просматривали под лупой при увеличении ×10.
В лабораторных условиях свежие сборы рясок, а также гербарные образцы изучали с помощью микроскопа МСП-1 (вариант 22). При обнаружении цветущих или плодоносящих растений определяли расположение генеративных органов относительно продольной оси фронда (в левом или правом почечном (боковом) кармашке), подсчитывали количество семян в плоде. Для четырех изученных образцов расположение генеративных органов нами не зафиксировано, отмечен лишь факт цветения или плодоношения растений; эти образцы в цитируемых гербарных этикетках отмечены знаком вопроса. Еще один образец со знаком вопроса – сбор П.П. Грищенко, процитированный в работе В.Ф. Тарасевич (1990) без уточнения расположения генеративных органов рясок. Цветущие и плодоносящие экземпляры рясок фотографировали с помощью цифровой камеры Levenhuk М800 PLUS.
Гербарный материал хранится в Институте проблем освоения Севера Тюменского научного центра СО РАН (TMN) и гербарии Тобольской комплексной научной станции Уральского отделения РАН.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Из всего количества изученных гербарных образцов рясковых в 41 из них (примерно в каждом четвертом образце) обнаружены цветущие и плодоносящие растения разных видов рясок, причем на одном гербарном листе могло присутствовать сразу несколько видов сем. Lemnaceae, находящихся в состоянии бутонизации, цветения или плодоношения. В это число не входят сведения из литературных источников, в которых отсутствуют точные данные о местонахождении цветущих растений рясковых. Ниже приведен перечень образцов с данными о их местонахождении, полученными с этикеток сборов (также см. Сокращения).
1. Lemna minor L. Около Аркатьевой. 21.07.1886 г. П.Н. Крылов; ТК (2 гербарных листа), переопределено на L. turionifera Landolt, цв. (?).
2. L. minor. Ишимск. окр. Около дер. Афонькиной, 55.5° с.ш. и 39° в.д., болото. 21.07.1927 г. П. Крылов и Л. Сергиевская; ТК, переопределено на L. turionifera, цв. (?).
3. L. minor. Бурятская АССР, Селенгинский р-н, окр. оз. Щучье, в болотце. 02.08.1958. Л.П. Сергиевская и З.Г. Шункова”; ТК, переопределен на L. turionifera, цв. (?).
4. L. gibba L. Тюменская обл., устье р. Тавда, 19.08.1985, П.П. Грищенко; MHA (Тарасевич, 1990), цв. (?).
5. L. turionifera + L. trisulca L. Тюменская обл., Исетский р-н, окр. с. Ботники, болото, 06.06.1998. Хозяинова Н.В., Глазунов В.А., Науменко Н.И.; TMN, L. turionifera: цв. (л., пр.), м., L. trisulca: цв. (л.), ед.
6. L. turionifera. Новосибирская обл., Карасукский р-н, оз. Кротово, 23.08.2015. Максимова А.Ю., опр. Капитонова О.А.; гербарий А.Ю. Максимовой, пл. (?).
7. L. turionifera. Тюменская обл., Уватский р-н, 6 км к ЮЗ от с. Горнослинкино, 58.7217° с.ш., 68.6981° в.д., мелководье р. Иртыш, 16.06.2016. Капитонова О.А.; ТКНС, цв. (л.), м.
8. L. turionifera + L. trisulca. Тюменская обл., г. Тобольск, 58.1946° с.ш., 68.2331° в.д., водоем возле паромной переправы, 05.07.2016. Капитонова О.А.; ТКНС, L. turionifera: цв. (л., пр.), пл. (л., пр.), м., L. trisulca: цв. (л., пр.), пл. (л., пр.), об., ОПП 100%.
9. L. gibba. Тюменская обл., Уватский р-н, окр. с. Горнослинкино, 58.7720° с.ш., 68.7642° в.д., мелководье р. Иртыш, 05.07.2017. Капитонова О.А.; ТКНС, цв. (л., пр.), пл. (л., пр.), об.
10. L. gibba. ХМАО-Югра, Ханты-Мансийский р-н, правый берег р. Иртыш, карьер, 60.6397° с.ш., 69.8930° в.д., 08.07.2016. Николаенко С.А.; TMN, пл. (л., пр.), м.
11. L. gibba. ХМАО-Югра, Октябрьский р-н, пос. Мал. Атлым, левый берег р. Тошманки, 62.2144° с.ш., 67.0027° в.д., 14.07.2016. Николаенко С.А.; TMN, цв. (л., пр.), пл. (л., пр.), об.; t 24°С, D 0.3–0.5 м.
12. L. turionifera. ХМАО-Югра, Октябрьский р-н, о-в напротив пос. Мал. Атлым, пойменное озеро, 62.2036° с.ш., 66.8227° в.д., 13.07.2016. Николаенко С.А.; TMN, цв. (л., пр.), пл. (л., пр.), м.; t 26 °С, D 0.1–0.4.
13. L. minor. Тюменская обл., Упоровский р-н, 2.5 км к СВ от д. Черная, 56.3524° с.ш., 66.3670° в.д., торфяник, 01.08.2017. Капитонова О.А.; ТКНС, цв. (л.), ед., ОПП 60%.
14. Spirodela polyrhiza (L.) Schleid. Тюменская обл., окр. г. Тюмень, оз. Кыртыма, 56.9936° с.ш., 65.8329° в.д., 08.08.2017. Николаенко С.А.; TMN, цв. (л., пр.), об.; t 30°C, D 0.2–0.7 м.
15. L. turionifera. Тюменская обл., Вагайский р-н, в 0.5 км к югу от пос. Первомайский, 57.3951° с.ш., 69.0564° в.д., пруд, 14.08.2017. Капитонова О.А.; ТКНС, цв. (л., пр.), ед., ОПП 90%.
16. L. trisulca. Тюменская обл., Сладковский р-н, в 0.5 км к югу от дер. Таволжан, 55.3361° с.ш., 70.1517° в.д., обводненная выемка, 20.06.2018. Капитонова О.А.; ТКНС, цв. (л., пр.), об.; рН 8.36, t 22°C, S 0.3‰, D 0.3 м, ОПП 30%.
17. L. trisulca. Тюменская обл., Бердюжский р-н, в 4 км к ЮВ от с. Зарослое, 55.4444° с.ш., 68.6412° в.д., придорожный водоем, 11.07.2018. Капитонова О.А.; ТКНС, цв. (л., пр.), пл. (л., пр.), об. ОПП 100%.
18. L. trisulca. Тюменская обл., Казанский р-н, в 3.5 км к СЗ от д. Новоалександровка, 55.4147° с.ш., 68.8158° в.д., восточный берег оз. Сиверга, протока, 13.07.2018. Капитонова О.А.; ТКНС, цв. (пр.), м.; рН 8.14, t 23°C, S 0.7‰, D 0.25 м, ОПП 95%.
19. L. turionifera + L. trisulca. Тюменская обл., г. Тобольск, 58.1611° с.ш., 68.2597° в.д., 17.07.2018. Капитонова О.А.; ТКНС, L. turionifera: цв. (л., пр.), м., L. trisulca: цв. (л., пр.), м.; рН 6.72, t 33°C, S 0.1‰, D 0.4 м, ОПП 100%.
20. L. minor + L. turionifera + L. trisulca. Тюменская обл., г. Тобольск, 58.1620° с. ш., 68.2596° в. д., 17.07.2018. Капитонова О.А.; ТКНС, L. minor: цв. (л., пр.), об.; L. turionifera: цв. (л., пр.), пл. (л.), м.; L. trisulca: цв. (л.), пл. (пр.), м.; рН 5.96, t 31°C, S 0.2‰, D 0.3 м, ОПП 100%.
21. L. minor + L. turionifera + L. trisulca + S. polyrhiza. Тюменская обл., г. Тобольск, 58.1626° с.ш., 68.2601° в.д., 17.07.2018. Капитонова О.А.; ТКНС, L. minor: цв. (л., пр.), м.; L. turionifera: цв. (л., пр.), пл. (л.), м.; L. trisulca: цв. (л.), пл. (л., пр.), м., S. polyrhiza: цв. (л., пр.), пл. (л., пр.), ед.; рН 5.86, t 31°C, S 0.1‰, D 0.5 м, ОПП 100%.
22. L. minor + L. turionifera + L. trisulca + S. polyrhiza. Тюменская обл., г. Тобольск, 58.1770° с.ш., 68.2210° в.д., 18.07.2018. Капитонова О.А.; ТКНС, L. minor: цв. (пр.), м.; L. turionifera: цв. (пр.), м.; L. trisulca: цв. (пр.), м., S. polyrhiza: цв. (л.), ед.; рН 6.24, t 32°C, S 0.2‰, D 0.2 м, ОПП 100%.
23. L. turionifera. Тюменская обл., Тобольский р-н, в 8 км к северу от с. Абалак, 58.2084° с.ш., 68.5861° в.д., пруд, 19.07.2018. Капитонов В.И., Капитонова О.А.; ТКНС, цв. (л., пр.), м.
24. L. minor + L. turionifera + S. polyrhiza. Тюменская обл., Тобольский р-н, дер. Савина, 58.2090° с.ш., 68.2164° в.д., 21.07.2018. Капитонова О.А.; ТКНС, L. minor: цв. (л., пр.), м.; L. turionifera: цв. (л., пр.), м.; S. polyrhiza: цв. (пр.), ед.; рН 8.15, t 24°C, S 0.2‰, D 0.5 м, ОПП 95%.
25. L. minor + L. turionifera + L. trisulca. Тюменская обл., Тобольский р-н, дер. Савина, 58.2145° с.ш., 68.2150° в.д., 21.07.2018. Капитонова О.А.; ТКНС, L. minor: цв. (л., пр.), м.; L. turionifera: цв. (пр.), м.; L. trisulca: цв. (пр.), пл. (л.), м.; рН 6.18, t 23°C, S 0.2‰, D 0.5 м, ОПП 100%.
26. L. minor L. Тюменская обл., Тобольский р-н, дер. Савина, старица, 21.07.2018. Капитонова О.А.; ТКНС, цв. (пр.), м.; рН 9.30, t 25°C, S 0.2‰, D 0.3 м, ОПП 70%.
27. L. turionifera + L. trisulca. Тюменская обл., г. Тобольск, 58.1830° с.ш., 68.2243° в.д., старица, 24.07.2018. Капитонова О.А.; ТКНС, L. turionifera: цв. (л., пр.), м.; L. trisulca: цв. (л., пр.), пл. (л., пр.), м.; D 0.3 м, ОПП 100%.
28. L. minor + L. trisulca. Тюменская обл., г. Тобольск, 58.1650° с.ш., 68.2369° в.д., озеро, 24.07.2018. Капитонова О.А.; ТКНС, L. minor: цв. (л., пр.), м.; L. trisulca: цв. (л., пр.), пл. (л., пр.), м.; рН 6.34, t 25°C, S 0.3‰, D 0.3 м, ОПП 100%.
29. L. turionifera + L. trisulca. Тюменская обл., с. Уват, 59.1493° с.ш., 68.9093° в.д., старица, 01.08.2018. Капитонова О.А.; ТКНС, L. turionifera: цв. (л., пр.), м.; L. trisulca: цв. (л., пр.), пл. (л., пр.), м, ОПП 100%.
30. L. trisulca. Тюменская обл., Сладковский р-н, в 0.5 км к югу от дер. Таволжан, 55.3361° с.ш., 70.1517° в.д., копань, 08.08.2018. Капитонова О.А.; ТКНС, пл. (л., пр.), ед.; рН 8.46, t 27°C, S 0.3‰, D 0.2 м, ОПП 95%.
31. L. trisulca. Тюменская обл., Сладковский р-н, в 0.2 км к северу от дер. Михайловка, 55.2691° с.ш., 70.0874° в.д., пруд, 08.08.2018. Капитонова О.А.; ТКНС, пл. (л., пр.), ед.; рН 8.48, t 24°C, S 0.4‰, D 0.1 м, ОПП 100%.
32. L. trisulca. Тюменская обл., Ярковский р-н, водоем на берегу оз. Тамырлы, 57.4825° с.ш., 67.4604° в.д., 17.06.2019. Капитонов В.И., Капитонова О.А.; ТКНС, цв. (пр.), пл. (пр.), м.
33. L. trisulca. Тюменская обл., Вагайский р-н, в 600 м к северу от дер. Доронина, 58.0285° с.ш., 69.2827° в.д., 27.06.2019. Капитонова О.А.; ТКНС, цв. (пр.), пл. (пр.), м., ОПП 75%.
34. L. trisulca. Тюменская обл., Вагайский р-н, в 2.5 км к востоку от дер. Бегишево, 58.0477° с.ш., 69.1432° в.д., 27.06.2019. Капитонов В.И., Капитонова О.А.; ТКНС, цв. (пр.), пл. (пр.), м.
35. L. turionifera + L. trisulca. Тюменская обл., Тобольский р-н, в 0.4 км к югу от дер. Байгара, 58.0236° с.ш., 68.8439° в.д., 03.07.2019. Капитонова О.А.; ТКНС, L. turionifera: цв. (л., пр.), м.; L. trisulca: цв. (л., пр.), пл. (л., пр.), м.; t 20°C, S 0.2‰, D 0.4 м, ОПП 95%.
36. L. turionifera + L. trisulca. Тюменская обл., Тобольский р-н, в 0.3 км к югу от дер. Байгара, 58.0245° с.ш., 68.8442° в.д., 03.07.2019. Капитонова О.А.; ТКНС, L. turionifera: цв. (л., пр.), м.; L. trisulca: цв. (л., пр.), пл. (л., пр.), об.; t 20 °C, S 0.2‰, D 0.2 м, ОПП 100%.
37. L. turionifera + L. trisulca. Тюменская обл., Тобольский р-н, в 1.1 км к ЮЗ от дер. Байгара, 58.0216° с.ш., 68.8277° в.д., 03.07.2019. Капитонова О.А.; ТКНС, L. turionifera: цв. (л., пр.), м.; L. trisulca: цв. (л., пр.), пл. (л., пр.); ОПП 90%.
38. L. turionifera. Тюменская обл., г. Тобольск, мелиоративный канал, 19.07.2019. Капитонова О.А.; ТКНС, цв. (л.), ед.; ОПП 60%.
39. L. trisulca. Тюменская обл., Тобольский р-н, окр. пос. Прииртышский, озеро, 21.07.2019. Капитонова О.А.; ТКНС, цв. (л., пр.), пл. (л., пр.), м.; ОПП 90%.
40. L. turionifera. Тюменская обл., Ярковский р-н, окр. с. Аксарина, 57.7534° с.ш., 67.2687° в.д., копань, 23.07.2019. Николаенко С.А.; TMN, цв. (л., пр.), об.; t 24°C, D 0.1–0.4 м.
41. L. turionifera + S. polyrhiza. Тюменская обл., Уватский р-н, окр. с. Тугалово, 59.7657° с.ш., 69.7208° в.д., 06.08.2019. Николаенко С.А.; TMN, L. turionifera: цв. (л.), м.; t 23°C, D 0.4–0.6 м.
Наибольшее число образцов с цветущими растениями выявлено для L. turionifera и L. trisulca, плодоносящих – для L. trisulca (табл. 1). В целом, выявлено 84 случая генеративного размножения всех пяти видов рясковых, известных в Сибири, образцов с плодоносящими растениями оказалось в 2 раза меньше, чем с цветущими. В некоторых гербарных образцах зафиксировано цветение сразу нескольких видов рясковых. Так, в трех пойменных водоемах г. Тобольск (образцы под № 20, 24 и 25) в состоянии генеративного размножения отмечено сразу три вида этого семейства, в двух образцах (№ 21 и 22), также собранных в пойме р. Иртыш в пределах г. Тобольск, – четыре вида; еще девять образцов содержали по два вида, находящихся в состоянии цветения или плодоношения.
Таблица 1.
Число гербарных образцов с цветущими и плодоносящими растениями сем. Lemnaceae и локализация их генеративных органов
| Вид | Генеративные органы и их локализация | Всего случаев цветения и плодоношения | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| цветки | плоды | ||||||||||
| л. | пр. | л., пр. | ? | всего | л. | пр. | л., пр. | ? | всего | ||
| Lemna minor | 1 | 2 | 5 | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 8 |
| L. turionifera | 3 | 2 | 15 | 3 | 23 | 2 | 0 | 2 | 1 | 5 | 28 |
| L. gibba | 0 | 0 | 2 | 1 | 3 | 0 | 0 | 3 | 0 | 3 | 6 |
| L. trisulca | 3 | 6 | 11 | 0 | 20 | 1 | 4 | 12 | 0 | 17 | 37 |
| Spirodela polyrhiza | 1 | 1 | 2 | 0 | 4 | 0 | 0 | 1 | 0 | 1 | 5 |
| Всего | 8 | 11 | 35 | 4 | 58 | 3 | 4 | 18 | 1 | 26 | 84 |
Цветки или плоды рясковых обнаружены и в левых, и в правых кармашках фрондов. Для многих образцов в расположении генеративных органов не выявлено какой-либо закономерности – с равной вероятностью они были сосредоточены в обоих кармашках. Часто в одном сборе обнаруживали растения и в цветущем, и в плодоносящем состоянии.
Генеративное размножение рясковых выявлено в южных лесостепных, подтаежных и в северных таежных районах Сибири (рис. 1). Наиболее северный пункт, где было зафиксировано цветение рясковых, – окрестности пос. Мал. Атлым (Октябрьский р-н, ХМАО-Югра), чуть севернее 62° с.ш., где обнаружены цветущие растения L. turionifera и L. gibba (Глазунов, Николаенко, 2018). Окрестности оз. Щучье в Селенгинском р-не Бурятии (51.4° с.ш., 106.5° в.д.), где Л.П. Сергиевской и З.Г. Шунковой собрана цветущая L. turionifera (гербарий ТК), являются наиболее восточным пунктом, где обнаружены цветущие рясковые.
Рис. 1.
Карта-схема находок цветущих и плодоносящих видов Lemnaceae на территории Сибири: ☆ – Lеmna minor, ⚪ – L. gibba, △ – L. trisulca, ◇ – L. turionifera, ◻ – Spirodela polyrhiza; незакрашенные значки – цветущие образцы, закрашенные – плодоносящие образцы, наполовину закрашенные – цветущие и плодоносящие.

Значения физико-химических параметров экотопов в местах произрастания цветущих рясковых варьировали в широких пределах. Глубина воды в таких водоемах, как правило, была 0.1–0.7 м и хорошо прогревалась в летний период. Значения рН находились в пределах 5.96–9.30. Температура воды под цветущими растениями (в момент отбора проб) колебалась от 20 до 33°С. Цветение рясковых всегда наблюдали на открытых участках водоемов, не затененных прибрежной древесно-кустарниковой или высокотравной растительностью. Все цветущие и плодоносящие растения произрастали в пресных водах (концентрация растворенных солей была ≤0.7‰). Так, 20 июня и 8 августа 2018 г. в обводненной копани с пресной водой (соленость 0.3‰), находящейся на берегу оз. Соленое (Сладковский р-н, окр. дер. Таволжан), мы наблюдали цветение L. trisulca, однако в 200 м от копани, в оз. Соленое с соленостью воды 6.1‰, этот вид, имея такое же высокое проективное покрытие (95–100%), не цвeл.
В основном цветение рясковых наблюдали в составе маловидовых сообществ водных растений с доминированием разных видов рясок и их высоким обилием. Кроме них в составе фитоценозов присутствовали еще один–шесть видов гидрофитов с проективным покрытием ≤25%, но в ряде случаев достигающим 40–60%, иногда 80% (сообщества с доминированием Stratiotes aloides L. и Utricularia vulgaris L.).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Явление цветения и плодоношения рясковых издавна привлекало к себе внимание исследователей, и к настоящему времени генеративная сфера этих растений хорошо изучена. Согласно мнению многих авторов, она представлена крайне редуцированным соцветием ароидных, которое состоит из двух мужских и одного женского цветков, развивающихся в боковых кармашках фронда (Ростовцев, 1905; Иванова, 1970; Landolt, 1986). Соцветие окружено пленчатой спатой – редуцированным покрывалом ароидных. Плод может содержать от одного до нескольких семян, на поверхности которых отчетливо видны 10–18 продольных ребер (Landolt, 1986; Жмылёв и др., 1995). Считается, что соцветия развиваются лишь в одном из боковых кармашков фронда, чаще всего в левом (Леонова, 1982), реже – в обоих кармашках одновременно (Жмылёв и др., 1995), а в расположении цветоносных кармашков может наблюдаться как зеркальная, так и перевернутая симметрия (Иванова, 1970).
Изученные нами образцы показали, что в пределах одной популяции соцветия с равной вероятностью формировались как в левом, так и в правом кармашке, при этом не удалось выявить какую-либо закономерность в локализации генеративных органов рясковых. У всех рясок к моменту цветения фронды становились значительно мельче, особенно это было заметно у Lеmna trisulca. У L. gibba нами зафиксированы случаи одновременного развития в одном кармашке и дочернего фронда, и соцветия, причем дочерний фронд находился в состоянии цветения, в то время как в самом материнском растении развивался плод в том же самом кармашке.
По мнению Питерса (2013), цветение Lemnaceae – стресс-индуцированное явление, однако до сих пор не установлены экологические условия, которые вызывают переход растений к генеративному размножению (Жмылёв и др., 1995). Условия, при которых мы наблюдали цветение рясок, показывали широкие колебания значений измеряемых абиотических факторов. Предположительно, стимулирующие цветение рясковых факторы – повышенные температура воды и освещенность, но не до экстремально высоких значений, к биотическим факторам можно отнести переуплотнение популяции.
Согласно нашим наблюдениям, произрастающие в Сибири виды рясковых обладают не только видоспецифическими характеристиками морфологического строения и географического распространения, но и имеют некоторые свойственные им особенности генеративного размножения.
Lеmna minor считается гемикосмополитным плюризональным видом, широко распространенным в Сибири (Ковтонюк, 1987). Растения этого вида имеют обратнояйцевидные, немного асимметричные фронды зеленого или светло-зеленого (салатового) цвета, с обеих сторон плоские, сверху немного килеватые, с немногими бугорками вдоль киля, причем апикальный бугорок отчетливо крупнее остальных. Длина фронда составляет 1–8(10) мм, ширина 0.6–5 мм (Landolt, 1986; Wolff, Landolt, 1994), однако, в природных условиях они обычно значительно мельче: длина чаще всего не превышает 2–6(7) мм.
Ряску малую долгое время считали в Сибири единственным из плавающих на поверхности воды видов этого рода даже после того, как обнаружено произрастание в регионе L. turionifera (Волобаев, 1992), обитающей в континентальных районах Северной Америки и Евразии (Landolt, 1975, 1986). Просмотр гербарного материала по Сибири (TK) показал, что многие сборы, определенные как L. minor, на самом деле относятся к L. turionifera. Материалы собственных исследований также свидетельствуют в пользу широкого распространения в Сибири L. turionifera, тогда как L. minor встречается гораздо реже. Этот вид более теневынослив и чаще, чем L. turionifera, отмечается в условиях затенения древесно-кустарниковой и высокотравной растительностью.
Для Lеmna minor характерна протогиния, отмеченная и другими авторами (Леонова, 1982; Жмылёв и др., 1995): сначала из кармашка появляется пестичный цветок, затем один из тычиночных, через некоторое время – второй тычиночный цветок (рис. 2а). За все время наблюдений мы обнаружили лишь восемь случаев цветения растений этого вида и ни одного случая плодоношения (табл. 1). По-видимому, это связано не только с редкостью данного вида на территории Сибири, но и с условиями, в которых он обитает в регионе: затененностью и часто относительно низкой температурой воды (обводненные участки залесенных болот, заболоченные леса). Цветение L. minor можно наблюдать лишь в более благоприятных условиях, где она чаще всего произрастает в совместных сообществах с другими рясковыми, имея при этом невысокое обилие.
Рис. 2.
Генеративные органы рясковых: а – соцветие L. minor в правом почечном кармашке (стенка кармашка разорвана и отогнута), б – соцветие L. turionifera в левом почечном кармашке, в – плодоносящие растения L. gibba, плоды расположены в правых кармашках, г – односемянный плод L. gibba в левом кармашке (семя вынуто из плода). 1 – рыльце пестичного цветка, 2 – тычинка, 3 – апикальный бугорок, 4 – плод, 5 – семя.
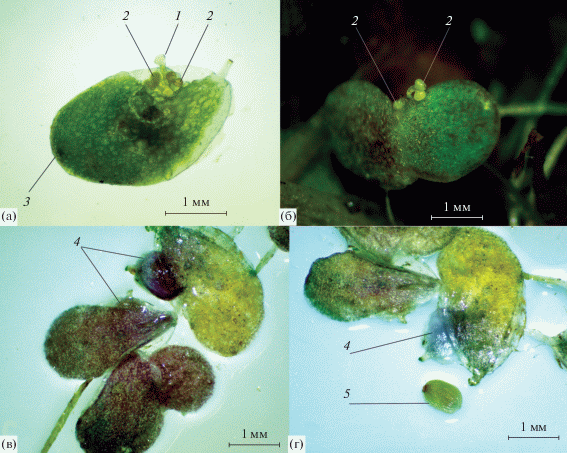
L. turionifera по своим морфологическим характеристикам очень похожа на L. minor, однако имеется ряд существенных отличий. Фронды L. turionifera обычно небольшие (2–4 мм длины и 0.8–3.5 мм ширины), слегка толстоватые, часто округлые; если более вытянутые, то цвет у них темно-зеленый, оливковый, обычно с красной пигментацией, более интенсивной в базальной части нижней стороны фронда. На верхней поверхности фронда вдоль срединного гребня имеется отчетливый ряд бугорков, причем они не отличаются друг от друга по размерам. К осени у растений этого вида формируются турионы – мелкие бурого или оливкового цвета круглые бескорневые зимующие фронды (Landolt, 1975, 1986). В Сибири этот вид распространен почти повсеместно, за исключением арктических широт. Цветущие растения L. turionifera (рис. 2б). встречались чаще, чем плодоносящие: всего отмечено 23 случая цветения и пять случаев плодоношения. Как и для L. minor, для него характерна протогиния. Плоды односемянные.
Lemna gibba имеет округлые или обратнояйцевидные цельнокрайние, снизу шарообразно выпуклые фронды 3–7 мм длиной, 2.5–5 мм шириной, желто-зеленого цвета с крупными воздухоносными полостями, хорошо заметными с нижней стороны. Также отличительный признак вида – наличие красной пигментации, сосредоточенной в основном с нижней стороны по периферии фронда (а не в базальной части, как у L. turionifera) (Landolt, 1986). На территории Сибири распространение вида пока ограничено Западной Сибирью, где он отмечен в Курганской (Науменко, 2008) и Тюменской областях (Тарасевич, 1990; Глазунов и др., 2017; Капитонова, 2018), ХМАО-Югре (Панкова, 2013, 2014). Полученные данные говорят о расширении в последнее время ареала вида в Западной Сибири, как в северном, так и восточном направлениях.
Почти во всех местах находок L. gibba в Сибири она находилась в состоянии генеративного размножения (рис. 2в, 2г), что, вероятно, является подтверждением предположения о цветении рясок в стрессовых условиях (Pieterse, 2013). В настоящее время наиболее северное нахождение L. gibba – пос. Мал. Атлым (ХМАО-Югра), здесь отмечено обильное цветение и плодоношение вида. В период цветения фронды L. gibba обычно окрашиваются в яркий красно-фиолетовый цвет, причем не только с нижней, но с верхней стороны. Возможно, у растений этого вида существует связь между наступлением генеративного периода онтогенеза и выработкой ими антоциана для привлечения насекомых-опылителей, что может считаться подтверждением энтомофилии рясковых (Иванова, 1970). Также это, вероятно, связано с более интенсивным поглощением тепла для перехода к генеративной стадии развития, что особенно важно в экстремальных условиях существования у северной границы ареала этого макротермного вида. Соцветия и плоды L. gibba отмечены и в левых, и в правых кармашках. Для растений характерна протогиния (Иванова, 1970); плоды чаще всего содержали два семени, редко – три–четыре.
L. trisulca имеет существенные отличия от других рясок в морфологическом строении и экологии, что дает основание некоторым авторам рассматривать ее в рамках особого рода – Staurogeton trisulcus (L.) Schur (Цвелёв, 2000; Ковтонюк, 2012). Ряска трехдольная имеет зубчатые по краю фронды, самые крупные по сравнению с таковыми остальных представителей семейства в рассматриваемом регионе. Их длина вместе со стипой (ножкой, соединяющей отдельные фронды) достигает 5–20 мм, ширина – 2.5–5 мм. В отличие от остальных рясок, у растений L. trisulca щель почечного кармашка не совпадает с краем фронда. Растения полностью погружены в воду и всплывают на поверхность лишь во время цветения.
L. trisulca широко распространена по всей Сибири и лишь в арктических широтах встречается крайне редко (Толмачёв, 1963; Ковтонюк, 1987). Это наиболее северный вид семейства, недавно найденный в зоне северных гипоарктических тундр на п-ве Ямал (70° с.ш.) (Зарубина, 2016). Наиболее северные места произрастания вида в Сибири – на п-ве Таймыр (72° с.ш.) (Поспелова, Поспелов, 2006, 2016, 2020).
Цветение L. trisulca представляет собой любопытное явление в связи с происходящими в растении морфологическими изменениями при наступлении генеративной стадии. Цветущие фронды ряски трехдольной значительно мельче вегетирующих, однако форма их не меняется. Они хорошо отличаются от других видов рясок не только своей формой, но и цветом: у L. trisulca фронды зеленого или ярко-зеленого цвета, тогда как у L. minor они светло-зеленые, у L. turionifera – темно-зеленые, оливковые. К признакам отличия особей L. trisulca от других видов рясок также относятся зубчатый край фронда, хорошо заметный при небольшом увеличении, и отсутствие бугорков на адаксиальной поверхности фронда. Кроме того, у цветущей ряски трехдольной фронды немного изогнуты, “кудрявятся” на поверхности воды, что, возможно, связано с необходимостью приподнять цветки над водой.
Этот вид оказался наиболее часто цветущим из всех произрастающих в Сибири рясковых. Соцветия могут закладываться в обоих кармашках. Как и для других рясок, для него характерна протогиния. Плод ~1 мм в поперечнике, с широким крылом в верхней части, содержит единственное семя (рис. 3).
Рис. 3.
Генеративные органы L. trisulca: а – “перевернутая” симметрия в локализации генеративных органов, расположенных в правых кармашках, б – расположение генеративных органов в левых кармашках, в – выход из почечного кармашка пестичного цветка (видно рыльце пестика), г – выход из почечного кармашка одного из тычиночных цветков (вид снизу), д – сформировавшиеся плоды в правых кармашках материнского и дочернего фрондов, e – плод с одним семенем (извлечено из плода). 1 – рыльце пестичного цветка, 2 – тычинка, 3 – плод, 4 – семя.
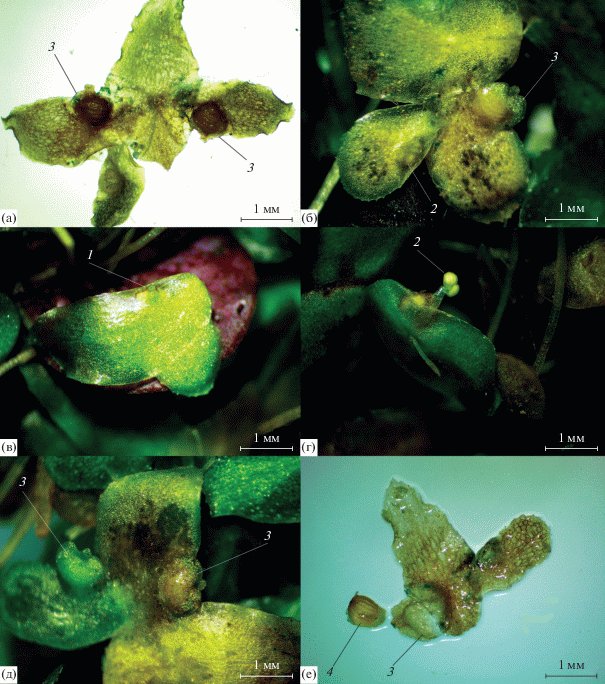
Spirodela polyrhiza имеет плавающие на поверхности воды округлые, плотные, плоские, цельнокрайние фронды 3–8 мм длиной и 3–6 мм шириной (рис. 4). Верхняя сторона фронда зеленая, с хорошо различимыми дугообразными жилками, нижняя – красно-фиолетовая, с пучком корней. К осени в кармашках образуются округлые турионы (рис. 2б) в диаметре 2–3 мм (Иконников, 1979). В Сибири вид распространен в умеренных широтах, произрастает в пресной воде богатой биогенами.
Рис. 4.
Spirodela polyrhiza: a – цветущее растение (край левого бокового кармашка отогнут), б – фронды с турионами. 1 – рыльце пестичного цветка, 2 – тычинка, 3 – турион.

Цветение S. polyrhiza можно отнести к редким явлениям: нам известно лишь четыре случая цветения вида в рассматриваемом регионе. Соцветия формировались как в левых (рис. 4а), так и в правых кармашках. Удалось обнаружить растения с формирующимися плодами. По-видимому, генеративное размножение многокоренника происходит значительно реже по сравнению с видами рода Lemna. Наши наблюдения на востоке европейской части России дали те же результаты: за все время, начиная с 90-х годов прошлого столетия, зафиксирован лишь один случай цветения многокоренника (Капитонова, 2001).
Выводы. Выявлено, что все пять видов рясковых, известных на территории Сибири, реализуют генеративный способ размножения. Наиболее северное местонахождение цветущих рясковых известно из окрестностей пос. Мал. Атлым (ХМАО-Югра, 62° с.ш.), наиболее восточное – в окрестностях оз. Щучье (Селенгинский р-н, Бурятия, 51.4° с.ш., 106.5° в.д.). Для широко распространенных в Сибири Lеmna turionifera и L. trisulca цветение и плодоношение можно считать обычным явлением, особенно характерным для переуплотненных популяций. Часты случаи генеративного размножения у L. gibba, в последнее время расширяющей свой ареал в северном и восточном направлениях. Более редкие случаи цветения зафиксированы у L. minor, распространение которой в регионе ограничено южными и западными районами. У широко распространенного в Сибири Spirodela polyrhiza генеративное размножение наблюдается крайне редко: выявлено четыре случая цветения и один случай плодоношения.
Случаи генеративного размножения рясковых зафиксированы лишь в пресноводных водоемах (соленость 0.1–0.7‰) с глубиной воды 0.1–0.7 м, температурой поверхностного слоя воды +20– +33°С, рН 5.96–9.30, не затененных прибрежной древесно-кустарниковой и высокотравной растительностью.
Список литературы
Власова Н.В. 2005. Семейство Lemnaceae // Конспект флоры Сибири: Сосудистые растения. Новосибирск: Наука. С. 318.
Волобаев П.А. 1992. Дополнения к флоре гидрофильных растений Сибири // Бот. журн. Т. 77. № 5. С. 62.
Глазунов В.А., Науменко Н.И., Хозяинова Н.В. 2017. Определитель сосудистых растений Тюменской области. Тюмень: ООО “РГ “Проспект”.
Глазунов В.А., Николаенко С.А. 2018. Новые данные о северных границах распространения некоторых видов растений в Западной Сибири // Труды XIV съезда РБО и конференции “Ботаника в современном мире”. Т. 1. Махачкала: АЛЕФ. С. 123.
Жмылёв П.Ю., Кривохарченко И.С., Щербаков А.В. 1995. Семейство рясковые // Биологическая флора Московской области. Вып. 10. Москва: Изд-во МГУ. С. 20.
Зарубина Е.Ю. 2016. Флора разнотипных водоемов Бованенковского нефтегазоконденсатного месторождения (полуостров Ямал) // Проблемы ботаники Южной Сибири и Монголии: Cб. науч. ст. по материалам XV международ. науч.-практ. конф. Барнаул: Концепт. С. 78.
Иванова И.Е. 1970. Некоторые особенности цветения и опыления рясок (Lemnaceae S. Gray) // Бот. журн. Т. 55. № 5. С. 649.
Иконников С.С. 1979. Семейство Lemnaceae S.F. Gray – Рясковые // Флора европейской части СССР. Т. 4. Ленинград: Наука. С. 317.
Капитонова О.А. 2000. Некоторые результаты изучения рясковых (Lemnaceae S. Gray) в Удмуртии // Вестн. Удмуртского ун-та. Сер. Биология. № 5. С. 3.
Капитонова О.А. 2001. К анализу флоры высших водных растений Удмуртии // Вестн. Удмуртского ун-та. № 7. С. 92.
Капитонова О.А. 2018. Ряска горбатая (Lemna gibba, Lemnaceae) – чужеродный вид во флоре Западной Сибири // Проблемы ботаники Южной Сибири и Монголии: Сб. науч. ст. по материалам XVII Международ. науч.-практ. конф. Барнаул: Изд-во АлтГУ. С. 83.
Капитонова О.А. 2019. Материалы к биологии и экологии рясковых (Lemnaceae) Сибири // Проблемы ботаники Южной Сибири и Монголии: Сб. науч. ст. по материалам XVIII международ. науч.-практ. конф. Барнаул: Изд-во АлтГУ. С. 127. https://doi.org/10.14258/pbssm.20190242019
Ковтонюк Н.К. 1987. Семейство Lemnaceae – Рясковые // Флора Сибири. Araceae – Orchidaceae. Новосибирск: Наука. С. 15.
Ковтонюк Н.К. 2012. Семейство Lemnaceae Martinov // Конспект флоры Азиатской России: Сосудистые растения. Новосибирск: Изд-во СО РАН. С. 439.
Крылов П.Н. 1929. Флора Западной Сибири. Руководство к определению западносибирских растений. Вып. III. Cyperaceae – Orchidaceae. Томск: Издание Томского отделения РБО. С. 374.
Кузнецов Л.А., Заиченко О.Б. 1979. Три вопроса о ряске малой (Lemna minor L.) // Систематика, анатомия и экология растений Европейской части СССР: Сб. науч. тр. Ленинград: Ленингр. гос. пед. ин-т. С. 18.
Леонова Т.Г. 1982. Семейство рясковые (Lemnaceae) // Жизнь растений. Т. 6. Москва: Просвещение. С. 493.
Лисицына Л.И., Папченков В.Г. 2000. Флора водоемов России: определитель сосудистых растений. Москва: Наука.
Лукина Г.А. 1977. Биология рясковых // I Всесоюзная конф. по высшим водным и прибрежно-водным растениям: Тез. докл. Борок: Ин-т биол. внутр. вод. С. 21.
Лукина Г.А. 1984. Рясковые в природных и лабораторных условиях // Бот. журн. Т. 69. № 1. С. 81.
Матвеев В.И. 1977. О цветении рясок (Lemnaceae) в водоемах Заволжья // Бот. журн. Т. 62. № 10. С. 1498.
Миркин Б.М., Наумова Л.Г., Мулдашев А.А. 2002. Высшие растения: краткий курс систематики с основами науки о растительности. Москва: Логос.
Науменко Н.И. 2008. Флора и растительность Южного Зауралья. Курган: Изд-во Курганского ун-та.
Панкова Н.Л. 2013. Ряски Lemna turionifera и Lemna gibba в Ханты-Мансийском автономном округе – Югре // Биоразнообразие экосистем Крайнего Севера: инвентаризация, мониторинг, охрана: Матер. Всерос. конф. (Сыктывкар, 3–7 июня 2013 г.) [Электронный ресурс]. Сыктывкар: Ин-т биологии Коми НЦ УрО РАН. С. 239. Режим доступа: http://ib.komisc.ru/add/conf/tundra
Панкова Н.Л. 2014. Новые находки водных растений на территории Ханты-Мансийского автономного округа – Югры // Turczaninowia. Т. 17. Вып. 1. С. 66.
Печенюк Е.В. 1985. О цветении рясковых (Lemnaceae) в Хоперском государственном заповеднике // Бот. журн. Т. 70. № 8. С. 1066.
Поспелова Е.Б., Поспелов И.Н. 2006. Прибрежно-водные сосудистые растения во флоре полуострова Таймыр // Бот. журн. Т. 91. № 10. С. 1441.
Поспелова Е.Б., Поспелов И.Н. 2016. Изменения во флоре окрестностей с. Хатанга, Таймырский заповедник, за длительный период // Nature Conservation Research. Заповедная наука. Вып. 1(2). С. 65. https://doi.org/10.24189/ncr.2016.017
Поспелова Е.Б., Поспелов И.Н. 2020. Флора Таймыра: Информационно-справочная система [Электронный ресурс]. Режим доступа: http://byrranga.ru/lemnaceae/lemna_trisulka/index.htm (дата обращения: 04.03.2020).
Ростовцев С.И. 1905. Биолого-морфологический очерк рясок. Москва: Типо-литогр. В. Рихтер.
Тарасевич В.Ф. 1990. О положении семейства Lemnaceae в системе цветковых растений по палинологическим данным // Бот. журн. Т. 75. № 7. С. 959.
Тахтаджян А.Л. 1966. Система и филогения цветковых растений. Москва: Наука.
Толмачёв А.И. 1963. Арктическая флора СССР. Критический обзор сосудистых растений, встречающихся в арктических районах СССР. Вып. 4. Москва: Изд-во АН СССР.
Цвелёв Н.Н. 1996. Рясковые – Lemnaceae S.F. Gray // Сосудистые растения советского Дальнего Востока. Т. 8. Санкт-Петербург: Наука. С. 364.
Цвелёв Н.Н. 2000. Определитель сосудистых растений Северо-Западной России (Ленинградская, Псковская и Новгородская области). Санкт-Петербург: Изд-во СПХФА.
Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). Русское издание. Санкт-Петербург: Мир и семья.
Щербаков А.В. 2010. Сосудистая водная флора Орловской области. Москва: Товарищество научных изданий КМК.
Azer S.A. 2013. Taxonomic revision of genus Lemna L. (Lemnaceae Gray) in Egypt // Ann. Agric. Sci. V. 58. Iss. 2. P. 257. https://doi.org/10.1016/j.aoas.2013.07.015
Cabrera L.I., Salazar G.A., Chase M.W. et al. 2008. Phylogenetic relationships of aroids and duckweeds (Araceae) inferred from coding and noncoding plastid DNA // American J. of Botany. V. 95. Is. 9. P. 1153. https://doi.org/10.3732/ajb.0800073
Chase M.W., Christenhusz M.J.M., Fay M.F. et al. 2016. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV // Bot. J. Linn. Soc. V. 181. Is. 1. P. 1. https://doi.org/10.1111/boj.12385
Landolt E. 1975. Morphological differentiation and geographical distribution of the Lemna gibba–Lemna minor group // Aquat. Bot. V. 1. P. 345.
Landolt E. 1986. The family of Lemnaceae – a monographic study // Veröff. Geobot. Inst. Zürich: Stiftung Rübel. H. 71. 572 S.
Les D.H., Crawford D.J., Landolt E. et al. 2002. Phylogeny and Systematics of Lemnaceae, the Duckweed Family // Syst. Bot. V. 27. Is. 2. P. 221. https://doi.org/10.1043/0363-6445-27.2.221
Pieterse A.H. 2013. Is flowering in Lemnaceae stress-induced? A review // Aquat. Bot. V. 104. P. 1. https://doi.org/10.1016/j.aquabot.2012.08.002
Rothwell G.W., Van Atta M.R., Ballard Jr. et al. 2004. Molecular phylogenetic relationships among Lemnaceae and Araceae using the chloroplast trnL-trnF intergenic spacer // Mol. Phylogenet. Evol. V. 30. Iss. 2. P. 378. https://doi.org/10.1016/S1055-7903(03)00205-7
Stockey R.A., Hoffman G.L., Rothwell G.W. 1997. The Fossil Monocot Limnobiophyllum scutatum: Resolving the Phylogeny of Lemnaceae // Am. J. Bot. V. 84. Iss. 3. P. 355.
Wolff P., Landolt E. 1994. Spread of Lemna turionifera (Lemnaceae), the red duckweed, in Poland // Fragm. Flor. Geobot. V. 39. Iss. 2. P. 439.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод