

Биология внутренних вод, 2021, № 4, стр. 392-404
Многолетняя динамика зоопланктона Камского и Воткинского водохранилищ
Е. М. Целищева a, В. И. Лазарева b, *
a Пермский филиал Всероссийского научно-исследовательского института рыбного хозяйства и океанографии
Пермь, Россия
b Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
Некоузский р-н, Ярославская обл., пос. Борок, Россия
* E-mail: lazareva_v57@mail.ru
Поступила в редакцию 28.05.2020
После доработки 01.12.2020
Принята к публикации 28.12.2020
Аннотация
В течение шести лет (2014–2019 гг.) исследован летний (июль–август) зоопланктон Камского и Воткинского водохранилищ на р. Каме. В двух водоемах обнаружено 106 видов ракообразных и коловраток, среди которых 10 вселенцев, преимущественно южного происхождения. Установлено, что 70% вселенцев в обоих водохранилищах зарегистрированы с 2012 по 2016 гг. Обычными видами стали кладоцера Diaphanosoma orghidani и понто-каспийские копеподы Eurytemora caspica и Heterocope caspia, они встречаются ежегодно в >30% проб и локально формируют до 30% численности ракообразных. Выявлено увеличение численности (до 24–30% количества ракообразных) термофильной копеподы Thermocyclops crassus, которая ранее была немногочисленна. В среднем за период 2014–2019 гг. в обоих водохранилищах численность зоопланктона в прибрежье была выше (>150 тыс. экз./м3), чем в пелагиали (в 1.7–2.0 раза ниже). Высокие показатели летней биомассы зоопланктона наблюдались (1.1–1.7 г/м3) по всей акватории водохранилищ. Уровень биомассы в 2014–2019 гг. оказался в 2.5–3.0 раза выше отмеченного в период 1950–1960 гг.
ВВЕДЕНИЕ
Река Кама – самый большой (>2000 км) и многоводный приток р. Волги, среднемноголетний расход воды в нижнем течении >4000 м3/с (Волга…, 1978). Каскад камских водохранилищ создан в 1954–1979 гг., зарегулированный участок реки расположен ниже устья р. Вишеры и включает три водохранилища: Камское, Воткинское и Нижнекамское (Эдельштейн, 1998). Трофический статус всех Камских водохранилищ по содержанию хлорофилла а в планктоне определен как эвтрофный; по сумме хлорофилла и феопигментов в донных осадках Камское водохранилище – эвтрофное, Воткинское – мезотрофное (Беляева и др., 2018).
Зоопланктон Камского и Воткинского водохранилищ изучают с 1956 и 1965 гг. соответственно (Уломский, 1961; Удалова, 1968; Серкина, 1971, 1975; Поскрякова, 1977; Кортунова, Зуева, 1979; Кортунова, Серкина, 1980; Кортунова, 1983; Кортунова, Галанова, 1988; Костицын и др., 2011; Крайнев, Кузнецова, 2013; Преснова, Хулапова, 2015; Селеткова, 2015). Мониторинг состояния сообщества проводит Пермский филиал Всероссийского научно-исследовательского института рыбного хозяйства и океанографии (ПермНИРО). Показано, что обилие зоопланктона р. Камы отличается большими колебаниями год от года (Кортунова, 1983; Кортунова, Галанова, 1988; Селеткова, 2015).
В 2016 г. в Камских водохранилищах выявлена большая группа видов-вселенцев, среди которых понто-каспийские солоновато-водные ракообразные (Лазарева, 2020). Особенности структуры и динамики обилия сообщества с учетом влияния вселенцев в последние годы не рассматривали.
Цель работы – анализ структуры, динамики численности и биомассы летнего зоопланктона Камского и Воткинского водохранилищ за период 2014–2019 гг., а также выяснить особенности расселения вселенцев.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Пелагический и литоральный зоопланктон собирали на глубине 5–13 и 1–3 м соответственно, на трех участках обоих водохранилищ – верхнем (I), центральном (II) и приплотинном (III) – в июле–августе 2014–2019 гг. по стандартной схеме разрезов (створов) и точек отбора проб ПермНИРО (рис. 1). На двух водохранилищах было установлено 18 разрезов. Ракообразных и коловраток учитывали в тотальных пробах зоопланктона, которые отбирали сетью Джеди с диаметром входного отверстия 12 см и ситом с ячеей 100 мкм. Сборы фиксировали 4%-ным формалином и просматривали в лаборатории под микроскопами ЛОМО МСП-2 и БИОЛАМ 70 (ЛОМО-Микросистемы, г. Санкт-Петербург).
Рис. 1.
Схема и районирование Камского и Воткинского водохранилищ. Участки водохранилищ: I – верхний, II – центральный, III – приплотинный.
В пробах определяли численность и биомассу каждого обнаруженного вида, рассчитывали общие значения этих показателей для четырех крупных таксономических групп (Cladocera, Cyclopoida, Calanoida, Rotifera) и всего сообщества. К доминантам относили виды с обилием ≥10% общей численности ракообразных или коловраток.
Сходство структуры доминантных комплексов определяли по индексу Чекановского–Съеренсена для количественных данных (ICzS) (Песенко, 1982):
ICzS = Σminpi,
где pi – доля вида в общей численности зоопланктона.
Для Камского и Воткинского водохранилищ за период с начала изучения по 2019 г. (фактически за все время существования водоемов) проанализировано изменение биомассы зоопланктона с привлечением данных литературных источников и архивных материалов ПермНИРО.
Рассчитаны среднее число видов в пробе, численность, биомасса и ошибка среднего.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Видовое богатство и доминанты. В 2014–2019 гг. в Камском водохранилище обнаружено 100 видов, в Воткинском – 69, в обоих водоемах – 106 видов. Большую часть из них (40–45%) составляли Rotifera, на долю Cladocera приходилось 35–40%, Copepoda – <20%. Видовая плотность (число видов в пробе) была высокой во все годы наблюдений. В пробах из пелагиали Камского водохранилища обнаруживали в среднем 21 ± 3 вида и Воткинского – 18 ± 3 вида, в прибрежье обоих водохранилищ – несколько меньше видов (по 17 ± 2 вида). В период 2012–2014 гг. были выявлены не указанные в обзорном списке (Лазарева, 2020) семь видов коловраток Keratella serrulata (Ehrenberg, 1838), Notholca acuminata (Ehrenberg, 1832), Euchlanis lyra Hudson, 1886, Mytilina mucronata (O.F. Muller, 1773), Synchaeta oblonga Ehrenberg, 1831, S. stylata Wierzejski, 1893, Filinia brachiata (Rousselet, 1901) – и два вида ракообразных Alonella nana (Baird, 1843) и Ilyocryptus agilis Kurz, 1878. Большинство из указанных видов найдены в Камском водохранилище. В Воткинском регистрировали четыре вида: E. lyra, Notholca acuminata, Synchaeta oblonga и Alonella nana.
Состав массовых видов коловраток верхнего участка Камского водохранилища заметно отличался от такового двух других участков (рис. 2а). Здесь часто доминировали Asplanchna priodonta Gosse, 1850 (10–60% Nrot), Brachionus angularis Gosse, 1851 (25–50% Nrot) и Synchaeta pectinata Ehrenberg, 1832 (30–40% Nrot). В центральном и приплотинном участках основной вклад в Nrot вносили Kellicottia longispina (Kellicott, 1879) (20–90%), Euchlanis dilatata lucksiana (Hauer, 1939) (10–90%), Keratella quadrata (O.F. Müller, 1786) (10–20%) и Polyarthra major Bruckhardt, 1900 (10–20%).
Рис. 2.
Соотношение (%) численности основных доминантов в группах коловраток (а, в) и ракообразных (б, г) пелагиали (П) и литорали (Л) Камского (а, б) и Воткинского (в, г) водохранилищ в 2014–2019 гг. (среднее за шесть лет). Участки: I – верхний, II – центральный, III – приплотинный. Rotifera: 1 – Euchlanis dilatata lucksiana, 2 – Polyarthra major, 3 – Keratella quadrata, 4 – Kellicottia longispina, 5 – Synchaeta pectinata, 6 – Brachionus angularis, 7 – Asplanchna priodonta. Crustacea: 8 – Thermocyclops crassus, 9 – Mesocyclops leuckarti, 10 – Bosmina longirostris, 11 – Daphnia galeata, 12 – Bosmina cf. crassicornis.
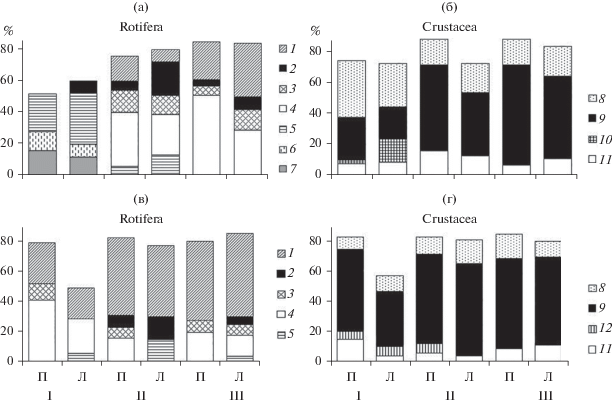
Обилие ракообразных по всей акватории водохранилища формировали в основном три вида (рис. 2б). Это копеподы Mesocyclops leuckarti (Claus, 1857) (10–70% Ncr) и Thermocyclops crassus (Fischer, 1853) (10–64% Ncr), а также кладоцера Daphnia galeata Sars, 1864 (10–60% Ncr). В литорали верхнего участка 10–23% Ncr приходилось на долю Bosmina (s. str) longirostris (O.F. Müller, 1785). В целом, пелагические и прибрежные сообщества каждого из участков характеризовались сходным соотношением главных доминантов (рис. 2а, 2б).
Вzoo на всей акватории водохранилища образовывала в основном крупная (длина тела до 2.0 мм) кладоцера Daphnia galeata. В течение шести лет наблюдений ее вклад в биомассу варьировал в пределах 10–90%, в среднем 55 ± 15% в пелагиали и 40 ± 10% в литорали (рис. 3а). В верхнем участке водоема большое значение имели коловратки рода Asplanchna (в среднем до 20%, в отдельные годы до 50% Вzoo).
Рис. 3.
Соотношение (%) биомассы основных доминантов зоопланктона пелагиали (П) и литорали (Л) Камского (а) и Воткинского (б) водохранилищ в 2014–2019 гг. (среднее за шесть лет). Участки: I – верхний, II – центральный, III – приплотинный. 1 – Thermocyclops crassus, 2 – Mesocyclops leuckarti, 3 – Diaphanosoma orghidani, 4 – Daphnia galeata, 5 – Asplanchna herriсki, 6 – A. priodonta, 7 – Heterocope caspia, 8 – Eurytemora caspica.
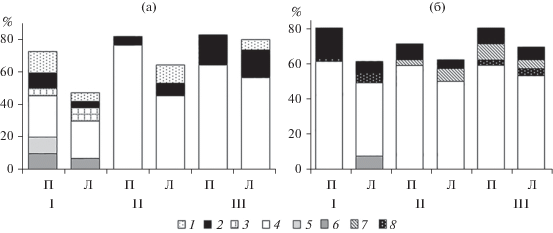
Структура доминантных комплексов зоопланктона различных участков водохранилища характеризовалась сравнительно низким уровнем сходства (в основном <50%). Это связано со “вспышками” численности (до 15–40% Nrot и 10–30% Ncr) в отдельные годы видов, в другое время малочисленных (например, Keratella cochlearis (Gosse, 1851), Ploesoma truncatum (Levander, 1894), Chydorus sphaericus (O.F. Müller, 1785), Bosmina (Eubosmina) cf. crassicornis (Lilljeborg, 1887)), а также нетипичных для биотопа (в пелагиали Brachionus quadridentatus Hermann, 1783, в литорали Eurytemora caspica Sukhikh et Alekseev, 2013). Наиболее высокие значения индекса сходства структуры сообщества отмечали между центральным и приплотинным участками, в пелагиали они достигали 60–70%, в литорали 65–75%. Сходство структуры зоопланктона в течение ряда лет с 2014 по 2019 гг. не превышало 60% в пелагиали и 40% в литорали. Наиболее близкой (50–60%) она была в годы (2014 и 2019, 2015, 2017 и 2019 гг.) со слабым летним прогревом воды (температура 17 ± 2°C), наибольшие различия (сходство <35%) отмечены в 2016 и 2018 гг. с высоким прогревом воды (22 ± 2°C).
Повсюду в пелагиали Воткинского водохранилища почти ежегодно были многочисленны коловратки Euchlanis dilatata lucksiana (15–95% Nrot) и Kellicottia longispina (10–80% Nrot) (рис. 2в). Среди ракообразных преобладали копеподы Mesocyclops leuckarti (30–80% Ncr) и Thermocyclops crassus (10–35% Ncr), которые в сумме формировали до 75% Ncr (рис. 2г). В отдельные годы (2014–2015 гг.) массовым видом была Daphnia galeata (25–26%).
Таксоценоз ракообразных прибрежной зоны этого водохранилища фактически не отличался от пелагического (рис. 2г). В литорали центрального участка наряду с обычными для всей акватории видами коловраток сравнительно многочисленными были Polyarthra major (до 35% Nrot) и Synchaeta pectinata (до 30% Nrot) (рис. 2в).
На всей акватории водохранилища почти всю биомассу зоопланктона (до 85%) образовывала кладоцера Daphnia galeata (рис. 3б). В пелагиали 40% Вzoo достигал вклад Mesocyclops leuckarti. В общем, соотношение биомассы основных доминантов в пелагиали и прибрежье водоема существенно не различалось.
Структура зоопланктона различных участков водохранилища была сравнительно однородной и характеризовалась высоким уровнем сходства (>50%). Наибольшие значения (60–70%) индекса сходства отмечали между комплексами доминантов центрального и приплотинного участков. В отдельные годы (2018 и 2019 гг.) сходство структуры зоопланктона пелагиали достигало 80%, тогда как литоральные сообщества во все 6 лет наблюдений сильно различались (сходство <45%). Здесь, как и в Камском водохранилище, локально в отдельные годы наблюдали массовое развитие (60–70% Nrot и 13–60% Ncr) обычно малочисленных видов (например, Brachionus angularis, Asplanchna priodonta, Bosmina longirostris, Eurytemora caspica, Heterocope caspia Sars, 1897). Сходство структуры зоопланктона Воткинского водохранилища год от года не превышало 60%, наиболее близкой (54–60%) она была в 2014, 2015, 2016 и 2018 гг., максимальные различия (сходство <35%) отмечены между годами 2014 и 2019, 2016 и 2019 гг. Во внутрикаскадном Воткинском водохранилище не обнаружено четкой связи структуры сообщества с термическим режимом вод.
Значение вселенцев в сообществе. В зоопланктоне исследованных водохранилищ зарегистрированы 9–10 видов-вселенцев, впервые обнаруженных в основном в 2010-х гг. (табл. 1). Большинство из них (кроме Kellicottia bostoniensis (Rousselet, 1908)) представлены южными теплолюбивыми видами. Три из них (кладоцера Cercopagis pengoi (Ostroumov, 1891), копеподы Heterocope caspia и Eurytemora caspica) – солоноватоводные понто-каспийские формы. Из 10 вселенцев обычными для обоих водохранилищ (встречаются ежегодно в >30% проб) стали 3 вида: кладоцера Diaphanosoma orghidani Negrea, 1982 и копеподы Eurytemora caspica и Heterocope caspia.
Таблица 1.
Встречаемость и численность видов-вселенцев в Камском и Воткинском водохранилищах в 2014–2019 гг.
| Вид | Год первой находки |
Источ-ник | Период наблюдений | |||||
|---|---|---|---|---|---|---|---|---|
| 2014 | 2015 | 2016* | 2017 | 2018 | 2019 | |||
| Камское вдхр. | ||||||||
| Pompholyx sulcata | 1980-е | 1 | $\frac{{14}}{{0.9}}$ | $\frac{8}{{0.01}}$ | $\frac{{46 - 90}}{{12.0}}$ | $\frac{{33}}{{11.0}}$ | $\frac{{26}}{{6.0}}$ | $\frac{{17}}{{0.4}}$ |
| Brachionus diversi-cornis (Daday, 1883) | 2002 | 3 | $\frac{{14}}{{0.5}}$ | – | $\frac{{13 - 40}}{{0.8}}$ | – | – | – |
| Brachionus budapes-tinensis Daday, 1885 | 2003 | 3 | $\frac{2}{{0.5}}$ | – | $\frac{{25}}{{0.8}}$ | $\frac{6}{{0.6}}$ | – | – |
| Kellicottia bostoniensis | 2012 | 4 | $\frac{2}{{0.1}}$ | $\frac{{69}}{{0.6}}$ | – | $\frac{{17}}{{1.6}}$ | – | $\frac{{28}}{{1.1}}$ |
| Eurytemora capsica | 2012 | 1 | $\frac{{62}}{{2.0}}$ | $\frac{{31}}{{16.0}}$ | $\frac{{46 - 75}}{{0.8}}$ | $\frac{{83}}{{7.5}}$ | $\frac{{37}}{{0.8}}$ | $\frac{{56}}{{0.5}}$ |
| Conochiloides coenobasis | 2013 | 1 | $\frac{{10}}{{7.1}}$ | – | $\frac{{25}}{{21.0}}$ | – | $\frac{8}{{10.0}}$ | $\frac{6}{{1.8}}$ |
| Asplanchna henrietta | 2016 | 2 | – | – | $\frac{{25}}{{1.6}}$ | – | – | – |
| Diaphanosoma orghidani | 2016 | 2 | – | – | $\frac{{52 - 60}}{{21.0}}$ | $\frac{{94}}{{8.4}}$ | $\frac{{81}}{{31.0}}$ | $\frac{{67}}{{3.5}}$ |
| Cercopagis pengoi | 2016 | 2 | – | – | $\frac{{40}}{{0.05}}$ | $\frac{8}{{0.04}}$ | $\frac{4}{{0.05}}$ | $\frac{6}{{0.01}}$ |
| Heterocope caspia | 2016 | 2 | $\frac{{74}}{{5.9}}$ | – | $\frac{{79 - 90}}{{5.0}}$ | $\frac{{72}}{{3.3}}$ | $\frac{{63}}{{1.9}}$ | $\frac{{72}}{{0.5}}$ |
| Воткинское вдхр. | ||||||||
| Pompholyx sulcata | 1980-е | 1 | – | $\frac{7}{{3.4}}$ | $\frac{{27 - 100}}{{5.0}}$ | $\frac{{17}}{{0.8}}$ | – | $\frac{6}{{0.2}}$ |
| Brachionus diversicornis | 2002 | 1 | $\frac{{22}}{{2.4}}$ | $\frac{{44}}{{4.1}}$ | $\frac{{27 - 40}}{9}$ | – | $\frac{4}{{1.8}}$ | – |
| Brachionus budapestinensis | 2003 | 1 | – | – | $\frac{{9 - 40}}{{3.0}}$ | – | – | – |
| Eurytemora caspica | 2012 | 1 | $\frac{{100}}{{2.8}}$ | $\frac{{100}}{{4.9}}$ | $\frac{{73 - 100}}{{3.0}}$ | $\frac{{67}}{{4.6}}$ | $\frac{{22}}{{1.1}}$ | $\frac{{78}}{{1.3}}$ |
| Conochiloides coenobasis | 2016 | 2 | – | – | $\frac{{70}}{{10}}$ | – | $\frac{4}{{1.8}}$ | – |
| Asplanchna henrietta | 2016 | 2 | – | – | $\frac{{60}}{{14}}$ | – | – | – |
| Diaphanosoma orghidani | 2016 | 2 | – | – | $\frac{{91 - 100}}{{3.0}}$ | $\frac{{50}}{{1.4}}$ | $\frac{{63}}{{0.2}}$ | $\frac{{17}}{{0.01}}$ |
| Cercopagis pengoi | 2016 | 2 | – | – | $\frac{{15}}{{0.02}}$ | – | – | – |
| Heterocope caspia | 2016 | 2 | $\frac{{56}}{{1.7}}$ | $\frac{{85}}{{4.5}}$ | $\frac{{82 - 100}}{{19.0}}$ | $\frac{{83}}{{6.6}}$ | $\frac{{81}}{{28.3}}$ | $\frac{{72}}{{1.9}}$ |
Примечание. Над чертой – встречаемость, %, под чертой – максимальная численность, тыс. экз./м3, “–” – вид отсутствовал в пробах. Eurytemora caspica до 2016 г. определяли как E. affinis (Poppe, 1880). Источник: 1 – Архив ПермНИРО, 2 – (Лазарева, 2020), 3 – (Селеткова, 2015), 4 – (Крайнев и др., 2018). Для 2016 г. дополнительно использованы данные источника 2. * – дополнительно приведены данные работы (Лазарева, 2020).
Максимальная численность большинства южных коловраток не превышала 10 тыс. экз./м3, лишь некоторые виды локально в отдельные годы доминировали в зоопланктоне водохранилищ. Так, в августе 2016 г. в верхнем участке Камского водохранилища Conochiloides coenobasis Skorikov, 1914 достигал >20 тыс. экз./м3 (22% Nrot), а Pompholyx sulcata Hudson, 1885 – 12 тыс. экз./м3 (24% Nrot). В 2016 г. в центральной части Воткинского водохранилища вблизи устья р. Очер Asplanchna henrietta Langhaus, 1906 формировала 14 тыс. экз./м3 (11% Nrot).
Существенно бо́льшим был вклад южных ракообразных в численность Ncr, а крупных видов также в Вzoo. В 2015 г. в прибрежной зоне копепода Eurytemora caspica формировала высокую численность (до 16 тыс. экз./м3) в центральном и приплотинном участках Камского водохранилища (25–30% Ncr), а в 2019 г. – на всей акватории Воткинского (13–17% Ncr). Вклад этого вида в Вzoo достигал 25–40% в литорали Камского и 10–23% Воткинского водохранилищ. В пелагиали последнего E. caspica образовывала 14–16% Вzoo.
Еще два вида вносили заметный вклад только в Вzoo. В 2016 и 2018 гг. в Верхнем участке Камского водохранилища обилие Diaphanosoma orghidani достигало 20–30 тыс. экз./м3 или 13–17% Вzoo в пелагиали и 15–18% в литорали. В центральном участке водоема вид доминировал только в литорали (до 14% Вzoo в 2019 г.). В 2019 г. в Воткинском водохранилище копепода Heterocope caspia формировала 12–40% Вzoo в литорали и до 55% в пелагиали приплотинного участка.
Численность и биомасса. В пелагиали Камского водохранилища летом 2014–2019 гг. общее количество зоопланктона варьировало в пределах 30–190, в среднем 110 ± 27 тыс. экз./м3 и 0.4–1.9, в среднем 1.2 ± 0.2 г/м3 (табл. 2). Наибольший вклад в численность сообщества (>50%) вносили Cyclopoida (>20%) и Rotifera (30%), биомассу формировали в основном Cladocera (>70%). В центральном и приплотинном участках соотношение основных таксонов зоопланктона было стабильным все 6 лет наблюдений, тогда как в Верхнем участке сильно варьировало год от года (рис. 4). Здесь наблюдали наибольший (до 60%) вклад коловраток в численность сообщества. Количество меропланктона, представленного велигерами моллюсков рода Dreissena, во все годы было очень низким (в среднем 1.0 ± 0.5 тыс. экз./м3) при биомассе <0.01 г/м3.
Таблица 2.
Численность (тыс. экз./м3) и биомасса (г/м3) зоопланктона Камского водохранилища в 2014–2019 гг.
| Таксон | 2014 г. | 2015 г. | 2016 г. | 2017 г. | 2018 г. | 2019 г. |
|---|---|---|---|---|---|---|
| Пелагиаль | ||||||
| Cladocera | $\frac{{18 \pm 5}}{{1.10 \pm 0.34}}$ | $\frac{{2 \pm 1}}{{0.29 \pm 0.08}}$ | $\frac{{30 \pm 8}}{{1.25 \pm 0.24}}$ | $\frac{{18 \pm 4}}{{0.71 \pm 0.16}}$ | $\frac{{29 \pm 11}}{{1.54 \pm 0.30}}$ | $\frac{{7 \pm 2}}{{0.48 \pm 0.13}}$ |
| Cyclopoida | $\frac{{38 \pm 8}}{{0.22 \pm 0.05}}$ | $\frac{{21 \pm 7}}{{0.04 \pm 0.01}}$ | $\frac{{106 \pm 6}}{{0.28 \pm 0.03}}$ | $\frac{{65 \pm 9}}{{0.23 \pm 0.05}}$ | $\frac{{75 \pm 23}}{{0.20 \pm 0.13}}$ | $\frac{{33 \pm 4}}{{0.13 \pm 0.05}}$ |
| Calanoida | $\frac{{1.0 \pm 0.2}}{{0.03 \pm 0.01}}$ | $\frac{{1.0 \pm 0.5}}{{0.01 \pm 0.001}}$ | $\frac{{1.0 \pm 0.5}}{{0.02 \pm 0.01}}$ | $\frac{{1.0 \pm 0.1}}{{0.03 \pm 0.01}}$ | $\frac{{1.0 \pm 0.6}}{{0.01 \pm 0.001}}$ | $\frac{{1.0 \pm 0.5}}{{0.02 \pm 0.01}}$ |
| Rotifera | $\frac{{17 \pm 3}}{{0.03 \pm 0.01}}$ | $\frac{{6 \pm 2}}{{0.01 \pm 0.001}}$ | $\frac{{46 \pm 13}}{{0.24 \pm 0.10}}$ | $\frac{{29 \pm 6}}{{0.04 \pm 0.01}}$ | $\frac{{80 \pm 2}}{{0.14 \pm 0.01}}$ | $\frac{{22 \pm 8}}{{0.25 \pm 0.13}}$ |
| Nzoo/Вzoo | $\frac{{76 \pm 9}}{{1.38 \pm 0.37}}$ | $\frac{{31 \pm 8}}{{0.35 \pm 0.09}}$ | $\frac{{184 \pm 20}}{{1.79 \pm 0.16}}$ | $\frac{{114 \pm 8}}{{1.01 \pm 0.20}}$ | $\frac{{191 \pm 30}}{{1.90 \pm 0.41}}$ | $\frac{{64 \pm 7}}{{0.88 \pm 0.04}}$ |
| Dreissena veliger | $\frac{{1.0 \pm 0.2}}{{ < 0.01}}$ | $\frac{{1.0 \pm 0.5}}{{ < 0.01}}$ | $\frac{{1.0 \pm 0.5}}{{ < 0.01}}$ | $\frac{{1.0 \pm 0.2}}{{ < 0.01}}$ | $\frac{{6 \pm 1}}{{0.01 \pm 0.001}}$ | $\frac{{1.0 \pm 0.5}}{{ < 0.01}}$ |
| Литораль | ||||||
| Cladocera | $\frac{{54 \pm 5}}{{2.64 \pm 0.67}}$ | $\frac{{3 \pm 1}}{{0.73 \pm 0.39}}$ | $\frac{{45 \pm 9}}{{0.81 \pm 0.27}}$ | $\frac{{48 \pm 11}}{{1.22 \pm 0.30}}$ | $\frac{{42 \pm 14}}{{1.76 \pm 0.25}}$ | $\frac{{5 \pm 1}}{{0.29 \pm 0.05}}$ |
| Cyclopoida | $\frac{{44 \pm 6}}{{0.34 \pm 0.08}}$ | $\frac{{48 \pm 24}}{{0.07 \pm 0.03}}$ | $\frac{{175 \pm 51}}{{0.51 \pm 0.18}}$ | $\frac{{137 \pm 23}}{{0.58 \pm 0.13}}$ | $\frac{{111 \pm 17}}{{0.32 \pm 0.08}}$ | $\frac{{14 \pm 3}}{{0.03 \pm 0.001}}$ |
| Calanoida | $\frac{{1.0 \pm 0.2}}{{0.03 \pm 0.01}}$ | $\frac{{17 \pm 8}}{{0.3 \pm 0.14}}$ | $\frac{{1.0 \pm 0.2}}{{0.01 \pm 0.001}}$ | $\frac{{3 \pm 1}}{{0.09 \pm 0.02}}$ | $\frac{{2.0 \pm 0.4}}{{0.05 \pm 0.02}}$ | $\frac{{1.0 \pm 0.1}}{{0.02 \pm 0.01}}$ |
| Rotifera | $\frac{{36 \pm 10}}{{0.05 \pm 0.01}}$ | $\frac{{22 \pm 6}}{{0.02 \pm 0.001}}$ | $\frac{{61 \pm 15}}{{0.11 \pm 0.03}}$ | $\frac{{44 \pm 2}}{{0.08 \pm 0.01}}$ | $\frac{{165 \pm 50}}{{0.21 \pm 0.08}}$ | $\frac{{7 \pm 1}}{{0.03 \pm 0.01}}$ |
| Nzoo/Вzoo | $\frac{{139 \pm 5}}{{3.07 \pm 0.69}}$ | $\frac{{90 \pm 36}}{{1.13 \pm 0.56}}$ | $\frac{{283 \pm 67}}{{1.44 \pm 0.47}}$ | $\frac{{233 \pm 30}}{{1.97 \pm 0.41}}$ | $\frac{{323 \pm 71}}{{2.33 \pm 0.38}}$ | $\frac{{28 \pm 3}}{{0.36 \pm 0.06}}$ |
| Dreissena veliger | $\frac{{4 \pm 1}}{{ < 0.01}}$ | 0 | $\frac{{1.0 \pm 0.5}}{{ < 0.01}}$ | $\frac{{1.0 \pm 0.3}}{{ < 0.01}}$ | $\frac{{3.0 \pm 1.0}}{{ < 0.01}}$ | $\frac{{1.0 \pm 0.2}}{{ < 0.01}}$ |
Примечание. Здесь и в табл. 3 над чертой – численность, под чертой – биомасса; Nzoo и Вzoo – суммарные численность и биомасса Cladocera, Cyclopoida, Calanoida и Rotifera.
Рис. 4.
Соотношение (%) крупных таксономических групп зоопланктона на разных участках (I – верхний, II – центральный, III – приплотинный) Камского и Воткинского водохранилищ в 2014–2019 гг.
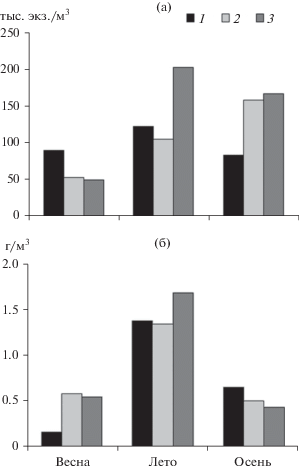
Сезонный ход развития пелагического зоопланктона исследован в Камском водохранилище в 2016 г., он характеризовался увеличением численности от весны (май) к лету (август) и незначительным ее снижением осенью (сентябрь–октябрь) (рис. 5а). В Центральном участке водоема осенью количество зоопланктона даже немного возрастало. Напротив, сезонная динамика биомассы отличалась четко выраженным летним максимумом, характерным для всех участков водохранилища (рис. 5б). В Верхнем его участке весной наблюдали минимальную по сравнению с другими участками биомассу, а осенью – максимальную.
Рис. 5.
Сезонный ход численности (а) и биомассы (б) зоопланктона Камского водохранилища в 2016 г. 1 – верхний участок (I), 2 – центральный (II), 3 – приплотинный (III).
В пелагиали Воткинского водохранилища летом 2014–2019 гг. численность зоопланктона варьировала в пределах 13–107, в среднем 76 ± ± 15 тыс. экз./м3, биомасса составляла 0.1–1.4, в среднем 1.0 ± 0.2 г/м3 (табл. 3). Наибольший вклад в численность сообщества вносили копеподы (~60%) и коловратки (>25%), биомассу формировали в основном кладоцеры (~80%). Соотношение численности основных таксонов зоопланктона год от года во всех трех участках водоема было нестабильным, варьировал не только вклад коловраток, но и доля копепод (рис. 4). Вариации структуры биомассы определялись преимущественно колебаниями обилия Cyclopoida. Количество велигеров моллюсков рода Dreissena в Воткинском водохранилище (в среднем 4.0 ± 0.5 тыс. экз./м3) было в 4 раза выше по сравнению с Камским, однако их биомасса не превышала 0.01 г/м3.
Таблица 3.
Численность (тыс. экз./м3) и биомасса (г/м3) зоопланктона Воткинского водохранилища в 2014–2019 гг.
| Таксон | 2014 г. | 2015 г. | 2016 г. | 2017 г. | 2018 г. | 2019 г. |
|---|---|---|---|---|---|---|
| Пелагиаль | ||||||
| Cladocera | $\frac{{10 \pm 3}}{{0.78 \pm 0.23}}$ | $\frac{{13 \pm 6}}{{1.21 \pm 0.42}}$ | $\frac{{4 \pm 1}}{{0.45 \pm 0.16}}$ | $\frac{{14 \pm 3}}{{1.26 \pm 0.45}}$ | $\frac{{5 \pm 2}}{{0.74 \pm 0.26}}$ | $\frac{{1.0 \pm 0.5}}{{0.06 \pm 0.02}}$ |
| Cyclopoida | $\frac{{33 \pm 9}}{{0.12 \pm 0.03}}$ | $\frac{{36 \pm 15}}{{0.11 \pm 0.04}}$ | $\frac{{79 \pm 12}}{{0.28 \pm 0.02}}$ | $\frac{{36 \pm 14}}{{0.10 \pm 0.02}}$ | $\frac{{67 \pm 12}}{{0.15 \pm 0.02}}$ | $\frac{{4 \pm 1}}{{0.02 \pm 0.01}}$ |
| Calanoida | $\frac{{1.0 \pm 0.1}}{{0.02 \pm 0.001}}$ | $\frac{{2.0 \pm 0.5}}{{0.06 \pm 0.02}}$ | $\frac{{2 \pm 0.5}}{{0.04 \pm 0.01}}$ | $\frac{{3 \pm 1}}{{0.06 \pm 0.01}}$ | $\frac{{2 \pm 0.5}}{{0.05 \pm 0.02}}$ | $\frac{{1 \pm 0.5}}{{0.02 \pm 0.01}}$ |
| Rotifera | $\frac{{12 \pm 5}}{{0.01 \pm 0.001}}$ | $\frac{{34 \pm 8}}{{0.04 \pm 0.01}}$ | $\frac{{20 \pm 4}}{{0.03 \pm 0.01}}$ | $\frac{{20 \pm 6}}{{0.02 \pm 0.01}}$ | $\frac{{26 \pm 10}}{{0.02 \pm 0.01}}$ | $\frac{{6 \pm 1}}{{0.01 \pm 0.001}}$ |
| Nzoo/Вzoo | $\frac{{60 \pm 18}}{{0.93 \pm 0.26}}$ | $\frac{{95 \pm 32}}{{1.42 \pm 0.49}}$ | $\frac{{107 \pm 18}}{{0.80 \pm 0.19}}$ | $\frac{{74 \pm 24}}{{1.44 \pm 0.49}}$ | $\frac{{105 \pm 25}}{{0.98 \pm 0.30}}$ | $\frac{{13 \pm 2}}{{0.11 \pm 0.03}}$ |
| Dreissena veliger | $\frac{{5 \pm 2}}{{0.01 \pm 0.001}}$ | $\frac{{9 \pm 4}}{{0.01 \pm 0.001}}$ | $\frac{{2 \pm 1}}{{0.01 \pm 0.001}}$ | $\frac{{1.0 \pm 0.5}}{{0.01 \pm 0.001}}$ | $\frac{{5 \pm 2}}{{0.01 \pm 0.001}}$ | $\frac{{1.0 \pm 0.5}}{{0.01 \pm 0.001}}$ |
| Литораль | ||||||
| Cladocera | $\frac{{29 \pm 15}}{{2.97 \pm 1.51}}$ | $\frac{{32 \pm 13}}{{1.29 \pm 0.64}}$ | $\frac{{39 \pm 13}}{{1.02 \pm 0.15}}$ | $\frac{{36 \pm 9}}{{1.43 \pm 0.40}}$ | $\frac{{10 \pm 3}}{{0.43 \pm 0.09}}$ | $\frac{{1.0 \pm 0.1}}{{0.02 \pm 0.01}}$ |
| Cyclopoida | $\frac{{73 \pm 26}}{{0.24 \pm 0.1}}$ | $\frac{{46 \pm 17}}{{0.10 \pm 0.03}}$ | $\frac{{113 \pm 18}}{{0.42 \pm 0.11}}$ | $\frac{{55 \pm 15}}{{0.11 \pm 0.03}}$ | $\frac{{68 \pm 16}}{{0.14 \pm 0.04}}$ | $\frac{{3.0 \pm 0.3}}{{0.01 \pm 0.001}}$ |
| Calanoida | $\frac{{5 \pm 2}}{{0.12 \pm 0.05}}$ | $\frac{{36 \pm 19}}{{0.13 \pm 0.05}}$ | $\frac{{2.0 \pm 0.5}}{{0.05 \pm 0.01}}$ | $\frac{{3 \pm 1}}{{0.08 \pm 0.04}}$ | $\frac{{3 \pm 1}}{{0.05 \pm 0.02}}$ | $\frac{{2.0 \pm 0.3}}{{0.04 \pm 0.01}}$ |
| Rotifera | $\frac{{64 \pm 17}}{{0.04 \pm 0.03}}$ | $\frac{{80 \pm 17}}{{0.12 \pm 0.03}}$ | $\frac{{107 \pm 44}}{{0.11 \pm 0.05}}$ | $\frac{{36 \pm 10}}{{0.06 \pm 0.01}}$ | $\frac{{50 \pm 20}}{{0.05 \pm 0.02}}$ | $\frac{{11 \pm 5}}{{0.01 \pm 0.01}}$ |
| Nzoo/Вzoo | $\frac{{177 \pm 60}}{{3.38 \pm 0.72}}$ | $\frac{{200 \pm 61}}{{1.65 \pm 0.72}}$ | $\frac{{262 \pm 39}}{{1.61 \pm 0.20}}$ | $\frac{{140 \pm 36}}{{1.67 \pm 0.43}}$ | $\frac{{134 \pm 37}}{{0.67 \pm 0.17}}$ | $\frac{{18 \pm 5}}{{0.08 \pm 0.01}}$ |
| Dreissena veliger | $\frac{{6 \pm 2}}{{ < 0.01}}$ | $\frac{{7 \pm 2}}{{ < 0.01}}$ | $\frac{{2.0 \pm 0.5}}{{0.31 \pm 0.18}}$ | $\frac{{10 \pm 3}}{{0.01 \pm 0.001}}$ | $\frac{{4 \pm 2}}{{ < 0.01}}$ | $\frac{{1.0 \pm 0.2}}{{ < 0.01}}$ |
В обоих водоемах численность литорального зоопланктона достоверно превышала количество пелагического в 1.7–2.0 раза (рис. 6а). В среднем в Камском водохранилище она достигала 183 ± 47 тыс. экз./м3, в Воткинском – 155 ± ± 33 тыс. экз./м3. Основу численности литорального сообщества Камского водохранилища составляли копеподы (60%), в Воткинском водохранилище доминировали копеподы (42%) и коловратки (36%). Биомасса прибрежного зоопланктона в обоих водоемах была высокой (1.5–1.7 г/м3) и достоверно не отличалась от наблюдаемой в те же сроки в пелагиали (1.1–1.4 г/м3) (рис. 6б).
Рис. 6.
Численность (а) и биомасса (б) зоопланктона в пелагиали (П) и литорали (Л) Камского и Воткинского водохранилищ в 2014–2019 гг.
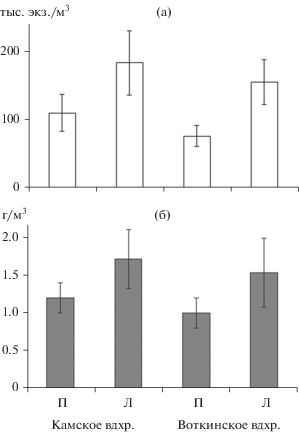
В среднем за период 2014–2019 гг. количество пелагического зоопланктона в Камском водохранилище было на 20–30% выше по сравнению с Воткинским (рис. 6), но эти различия не достоверны. Сходное соотношение обилия наблюдалось и в литорали, численность и биомасса различались на 10–15%.
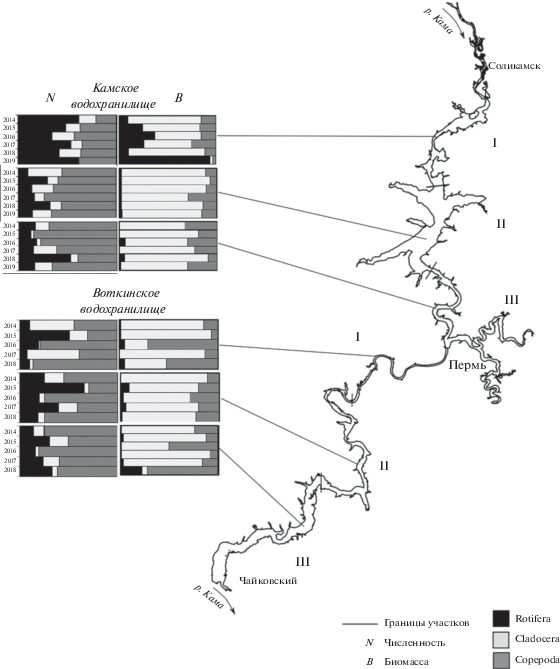
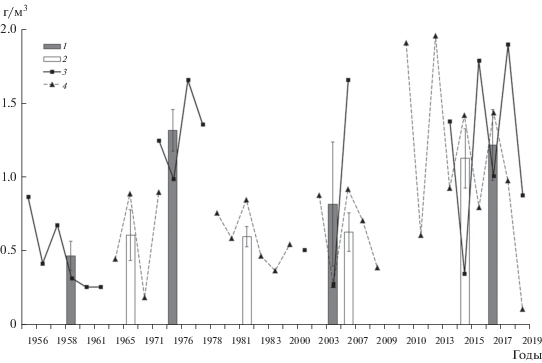
Анализ динамики биомассы зоопланктона Камского водохранилища за 63 года и Воткинского водохранилища за 54 года показал, что в обоих водоемах наблюдались значительные межгодовые колебания во все периоды исследований, а также четкая тенденция к росту биомассы в современный период (рис. 7). В Камском водохранилище биомасса пелагического зоопланктона в 2014–2019 гг. (1.2 ± 0.2 г/м3) оказалась выше отмеченной в первое десятилетие существования водоема (0.5 ± 0.1 г/м3) почти в 2.5 раза. Значения биомассы, сравнимые с современными (1.3 ± 0.1 г/м3), наблюдались в 1970-х годах. В Воткинском водохранилище до 2010 г. биомасса зоопланктона была сравнительно небольшой (0.6 ± 0.1 г/м3). Жарким летом 2010 г. отмечен троекратный подъем биомассы сообщества до 1.9 г/м3. В последующие годы (до 2018 г.) она оставалась чаще всего высокой (1.3 ± 0.2 г/м3), в среднем вдвое выше по сравнению с периодом до 2010 г. Биомасса зоопланктона водохранилища резко снизилась до 0.1 г/м3 в холодном и многоводном 2019 г.
Рис. 7.
Многолетние изменения биомассы зоопланктона Камского (1956–2019 гг.) и Воткинского (1965–2019 гг.) водохранилищ. 1 – Камское водохранилище, среднее за периоды три–шесть лет; 2 – Воткинское водохранилище, среднее за период три–девять лет; 3 – среднее за июль–август каждого года в Камском водохранилище; 4 – среднее за июль–август каждого года в Воткинском водохранилище. Источник: 1956–1959 гг. – (Уломский, 1961); 1961–1962 гг. – (Кортунова, Серкина, 1980; Серкина, 1971); 1965–1966 гг. – (Удалова, 1968); 1971–1975 гг. – (Серкина, 1975; Поскрякова, 1977); 1976–1978 гг. – (Кортунова, Зуева, 1979; Кортунова, Галанова, 1986); 1979–1982 гг. – (Кортунова, Галанова, 1988); 1983–2009 гг. – (Костицын и др., 2011; Селеткова, 2015); 2010–2011 гг. – данные архива ПермНИРО; 2013 г. – (Крайнев, Кузнецова, 2013); 2014–2019 гг. – настоящая работа.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В Камском и Воткинском водохранилищах обитает более 106 видов зоопланктона, почти половину из них составляют Rotifera. С учетом архивных данных ПермНИРО видовое богатство сообщества превышает 180 видов. Наиболее полно состав зоопланктона водохранилищ р. Камы приведен в обзоре Лазаревой (2020). Все семь видов, дополнивших этот список в данной работе, выявлены в неопубликованных материалах архива ПермНИРО. Они обычны для таежной зоны Европейской России немногочисленны (Пидгайко, 1984), и их не рассматривали в качестве вселенцев.
Работа с архивными данными ПермНИРО позволила установить сроки появления в Камском и Воткинском водохранилищах южных теплолюбивых видов-вселенцев, в том числе трех солоновато-водных понто-каспийских ракообразных (табл. 1). Только одна пресноводная южная коловратка Pompholyx sulcata проникла в эти водоемы еще в 1980-х гг., два вида рода Brachionus появились в начале 2000-х, остальные вселенцы – в 2010-х (Кузнецова, 2015; Селеткова, 2015; Крайнев и др., 2018; Лазарева, 2020).
Особый интерес представляет расселение в р. Каме понто-каспийских ракообразных. Самые первые находки копеподы Eurytemora caspica в обоих исследованных водоемах датируются 2012 г., вид определяли как E. cf. affinis (Кузнецова, 2015). Для сравнения, в р. Волге этот вид зарегистрирован с середины 1980-х гг. (Тимохина, 2000). В настоящее время морфологическими и молекулярными методами установлено (Сухих и др., 2020), что в водохранилищах рек Волги и Камы обитает E. caspica, тогда как типичная E. affinis (Poppe, 1880) нигде не обнаружена.
Местообитания двух других понто-каспийских видов достоверно установлены только с 2016 г. (Лазарева, 2020). В 2016‒2019 гг. хищная кладоцера Cercopagis pengoi отмечена ежегодно, но только в приплотинном участке Камского водохранилища. Сравнительно многочисленная во второй половине лета копепода Heterocope caspia предположительно появилась в обоих водохранилищах еще в начале 2000-х гг. Однако вселенца смешивали с обычной для таежной зоны H. appendiculata Sars, 1863. В водохранилищах р. Волги (Волгоградское, Саратовское и Куйбышевское) H. caspia многочисленна с середины 1960-х гг. (Волга…, 1978; Тимохина, 2000).
В водохранилищах Верхней Волги, как результат потепления, отмечено увеличение обилия циклопоидных копепод родов Mesocylops и Thermocyclops, особенно T. crassus (Лазарева, Соколова, 2015). Сходные изменения в планктоне были зарегистрированы в водоемах Западной Европы (Adrian et al., 2006). До середины 2000-х гг. в Камском водохранилище вклад Cyclopoida в биомассу пелагического зоопланктона варьировал в пределах 25–50%, в Воткинском – 8–30% (Поскрякова, 1977; Кортунова, 1983; Кортунова, Галанова, 1988; Преснова, Хулапова, 2015; Селеткова, 2015). В 2014–2019 гг. Cyclopoida формировали 10–20% биомассы в Камском водохранилище и 7–35% в Воткинском. До 2010 г. в планктоне обоих водоемов доминировал преимущественно Mesocyclops leuckarti (Кортунова, 1983; Кортунова, Галанова, 1988; Преснова, Хулапова, 2015; Селеткова, 2015). В 2014–2019 гг. в Камском водохранилище вклад Thermocyclops crassus в отдельные годы достигал до 24–30% численности ракообразных и до 20% биомассы зоопланктона.
Количество зоопланктона обоих изученных водохранилищ характеризуется большими колебаниями год от года (Кортунова, 1983; Кортунова, Галанова, 1988; Селеткова, 2015). Эту закономерность подтверждают и наши данные за 2014–2019 гг. (рис. 7). В современный период биомасса зоопланктона водохранилищ увеличилась в 2.5–3.0 раза по сравнению с 1950–1960-ми гг. В Камском водохранилище ее максимумы отмечены в 1970-е и 2010-е гг. В Воткинском водохранилище сильное увеличение биомассы наблюдалось только в 2010-х гг. Рост количества летнего зоопланктона в последнее десятилетие регистрируют и для других водохранилищ каскада, в частности, для водоемов Верхней и Средней Волги (Kopylov et al., 2012; Lazareva et al., 2014). Причиной считается эвтрофирование экосистем водохранилищ, вызванное глобальным потеплением.
Выводы. За шесть лет (2014–2019 гг.) в Камском и Воткинском водохранилищах были обнаружены 100 и 69 видов зоопланктона соответственно. Уточнены сроки появления десяти видов-вселенцев южного происхождения, 70% которых в обоих водохранилищах зарегистрированы с 2012 по 2016 гг. Из них три вида ракообразных (южная кладоцера Diaphanosoma orghidani и понто-каспийские копеподы Eurytemora caspica и Heterocope caspia) встречались ежегодно в >30% проб и локально образовывали до 30% численности ракообразных. Из южных коловраток в отдельные годы были многочисленны Conochiloides coenobasis, Pompholyx sulcata и Asplanchna henrietta (до 24% Nrot). Установлено увеличение вклада в сообщество (до 24–30% численности ракообразных) термофильной копеподы Thermocyclops crassus, которая ранее не доминировала. В период 2014–2019 гг. в обоих водохранилищах наибольшая численность зоопланктона (>150 тыс. экз./м3) наблюдалась в прибрежье, в пелагиали она была ниже в 1.7–2.0 раза. Биомасса прибрежного и пелагического зоопланктона фактически не различалась и варьировала в пределах 1.1–1.7 г/м3. Выявлена отчетливая тенденция увеличения биомассы зоопланктона водохранилищ в 2010-х гг., уровень биомассы в 2014–2019 гг. стал в 2.5–3.0 раза выше отмеченного в 1950–1960-х гг.
Список литературы
Беляева П.Г., Минеева Н.М., Сигарева Л.Е. и др. 2018. Содержание растительных пигментов в воде и донных отложениях водохранилищ р. Камы // Тр. Инст. биол. внутр. вод РАН. Вып. 81(84). С. 97. https://doi.org/10.24411/0320-3557-2018-10007
Волга и ее жизнь. 1978. Ленинград: Наука.
Кортунова Т.А. 1983. Зоопланктон Камского водохранилища и его продукция // Комплексные исследования рек и водохранилищ Урала. Пермь: Пермск. гос. ун-т. С. 68.
Кортунова Т.А., Галанова А.А. 1986. Зоопланктон Камских водохранилищ как кормовая база рыб // Биол. рес. Зап. Урала. Межвуз. сб. науч. тр. Пермь. С. 50.
Кортунова Т.А., Галанова А.А. 1988. Зоопланктон // Биол. Воткинского вдхр. Иркутск: Иркутск. гос. ун-т. С. 50.
Кортунова Т.А., Зуева Е.Ю. 1979. Состояние зоопланктона Камского водохранилища (по данным 1976 г.) // Сб. науч. трудов Пермской лаборатории Гос. НИИ озер. и реч. рыб. хоз-ва. Ленинград: Гос. НИИ озер. и реч. рыб. хоз-ва. Вып. 2. С. 22.
Кортунова Т.А., Серкина Р.А. 1980. Распределение и сезонная динамика зоопланктона Камского водохранилища // Биол. рес. водоемов Зап. Урала. Пермь: Пермский гос. нац. исслед. ун-т. С. 15.
Костицын В.Г., Поздеев И.В., Истомина А.М. и др. 2011. Биологическое обоснование к прогнозу вылова на 2012 год в основных рыбохозяйственных водоемах Пермского края и в Удмуртском секторе Воткинского водохранилища. Пермь: Пермское отделение Гос. НИИ озер. и реч. рыб. хоз-ва.
Крайнев Е.Ю., Кузнецова Е.М. 2013. Видовое разнообразие и распределение зоопланктона Воткинского водохранилища // Современные проблемы водохранилищ и их водосборов. Тр. Межд. науч.-практ. конф. Пермь: Пермск. гос. ун-т. С. 137.
Крайнев Е.Ю., Целищева Е.М., Лазарева В.И. 2018. Американская коловратка Kellicottia bostoniensis (Rousselet, 1908) (Rotifera: Brachionidae) в Камском водохранилище (река Кама, Россия) // Биол. внутр. вод. № 1. С. 55. https://doi.org/10.7868/S0320965218010072
Кузнецова Е.М. 2015. Зоопланктон Камского водохранилища в летний период 2013 г. // Фундаментальные и прикладные исследования в биологии и экологии: Матер. рег. студенческой науч. конф. 14–19 апр. 2014 г. Пермь: Пермский гос. нац. исслед. ун-т. С. 57.
Лазарева В.И. 2020. Многолетние изменения состава и обилия зоопланктона водохранилищ р. Камы // Биол. внутр. вод. № 3. С. 1. https://doi.org/10.31857/S0320965220030110
Лазарева В.И., Соколова Е.А. 2015. Метазоопланктон равнинного водохранилища в период потепления климата: биомасса и продукция // Биол. внутр. вод. № 3. С. 30. https://doi.org/10.7868/S0320965215030092
Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 286 с.
Пидгайко М.Л. 1984. Зоопланктон водоемов Европейской части СССР. М.: Наука.
Поскрякова Н.П. 1977. Зоопланктон Камы в августе 1975 г. // Биол. внутр. вод: Информ. бюл. Ленинград: Наука. № 36. С. 57.
Преснова Е.В., Хулапова А.В. 2015. Структура и распределение зоопланктона в центральном районе Воткинского водохранилища // Вест. Перм. ун-та. Биология. Вып. 4. С. 366.
Селеткова Е.Б. 2015. Зоопланктон Камского водохранилища // Биоценозы рек и водохранилищ Западного Урала. Санкт-Петербург: Гос. НИИ озер. и реч. рыб. хоз-ва. С. 136.
Серкина Р.А. 1971. Сезонная динамика зоопланктона Камского водохранилища // Уч. зап. Пермь: Пермск. гос. ун-т. № 261. С. 126.
Серкина Р.А. 1975. О состоянии зоопланктона в районе загрязнения Воткинского водохранилища // Биол. рес. Камских водохранилищ. Пермь: Пермск. гос. ун-т. С. 143.
Сухих Н.М., Лазарева В.И., Алексеев В.Р. 2020. Копепода Eurytemora caspica Sukhikh et Alekseev, 2013 (Crustacea, Calanoida) в водохранилищах Волги и Камы // Биология внутренних вод. № 2. С. 129. https://doi.org/10.31857/S0320965220020163
Тимохина А.Ф. 2000. Зоопланктон как компонент экосистемы Куйбышевского водохранилища. Тольятти: Ин-т экологии волж. бассейна РАН.
Удалова Н.Н. 1968. Зоопланктон Воткинского водохранилища на р. Каме в открытый период 1965–1966 гг. // Тезисы докладов конференции по изучению водоемов бассейна Волги. Тольятти: Ин-т экологии Волж. бассейна РАН. С. 125.
Уломский С.Н. 1961. Сырой вес массовых форм низших ракообразных Камского водохранилища и некоторых озер Урала и Зауралья // Тр. Уральск. отдел. Гос. науч.-исслед. ин-та озер. и реч. рыб. хоз-ва. Т. 5. Свердловск: Гос. НИИ озер. и реч. рыб. хоз-ва. С. 200.
Эдельштейн К.К. 1998. Водохранилища России: экологические проблемы и пути их решения. Москва: ГЕОС.
Adrian R., Wilhelm S., Gerten D. 2006. Life-history traits of lake plankton species may govern their phenological response to climate warming // Global Change Biol. V. 12. P. 1652. https://doi.org/10.1111/j.1365-2486.2006.01125.x
Kopylov A.I., Lazareva V.I., Mineeva N.M. et al. 2012. Influence of anomalous high water temperatures on the development of the plankton community in the Middle Volga Reservoirs in summer 2010 // Dokl. Biol. Sci. V. 442. P. 11. https://doi.org/10.1134/S0012496612010012
Lazareva V.I., Mineeva N.M., Zhdanova S.M. 2014. Spatial Distribution of Plankton from the Upper and Middle Volga Reservoirs in Years with Different Thermal Conditions // Biol. Bull. V. 41. № 10. P. 869. https://doi.org/10.1134/S1062359014100070
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод