

Биология внутренних вод, 2021, № 5, стр. 511-517
Сравнительный анализ пищевого спектра леща Abramis brama L. (Cyprinidae, Pisces) на разнотипных участках Рыбинского водохранилища
Г. Х. Щербина *
Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл., Россия
* E-mail: gregory@ibiw.ru
Поступила в редакцию 29.05.2019
После доработки 08.10.2020
Принята к публикации 15.01.2021
Аннотация
Приведены данные по кормовой базе и питанию леща из разных по кормности участков Рыбинского водохранилища осенью 2013 г. Всего в пищевом спектре леща обнаружен 21 вид беспозвоночных, преобладали хирономиды и моллюски – 9 и 6 видов соответственно. Показано, что на высококормных участках Шекснинского плёса, находящихся в зоне продолжительного локального промышленного загрязнения, снижается интенсивность питания леща, что приводит к уменьшению его упитанности, по сравнению с особями из условно чистых высококормных участков Моложского и Волжского плёсов. Сравнительный анализ с ранее проведенными исследованиями показал, что роль полисапробных личинок Chironomus f. l. plumosus в питании леща Рыбинского водохранилища в последние годы возросла.
ВВЕДЕНИЕ
Рыбинское водохранилище – один из крупнейших искусственных водоемов России. В северо-восточной части водохранилища (Шекснинский плёс) находится Череповецкий промышленный комплекс – крупнейший индустриальный центр, в который входят предприятия химической промышленности и черной металлургии. В район Шекснинского плеса поступает наибольшее количество сточных вод с расположенных там предприятий, поэтому этот участок считается самым загрязненным (Флеров и др., 2000; Чуйко и др., 2010; Заботкина, 2014). Основная масса загрязняющих веществ оседает в донные отложения, где аккумулируются различные тяжелые металлы, полихлорированные бифенилы и полиароматические углеводороды (Козловская, Герман, 1997; Флеров и др., 2000; Герман, Законнов, 2003; Чуйко и др., 2010).
Известно, что тяжелые металлы и органические загрязняющие вещества, будучи устойчивыми соединениями, поступают в организм рыб преимущественно по пищевым цепям, поэтому наиболее подвержены влиянию этих веществ рыбы-бентофаги (Герман, Козловская, 1999, 2001; Голованова, 2008). Лещ Abramis brama (Linnaeus, 1758) – самый массовый и распространенный в Рыбинском водохранилище вид, питается бентосными организмами, обитающими в поверхностном слое донных отложений (Житенева, 1980).
Питание бентосоядных рыб, в том числе леща, в Рыбинском водохранилище изучали с момента заполнения водоема (Ключарева, 1960). Наиболее подробно питание леща в водоеме было изучено Т.С. Житеневой (1958, 1959, 1980). Связь питания леща с кормовой базой Рыбинского водохранилища дана в работе (Баканов, Стрижникова, 1979). По наблюдениям Житеневой (1980), в Рыбинском водохранилище хирономиды являются основной пищей леща в течение всего вегетационного периода с максимальным их потреблением осенью, причем, основным кормовым объектом служат личинки и куколки мотыля Chironomus f. l. plumosus.
Цель работы – изучить пищевой спектр леща из разных по кормности участков Рыбинского водохранилища и сравнить его результатами ранее проведенных исследований.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материалом для исследования были лещи, выловленные в конце нагульного периода (конец сентября – начало октября 2013 г.) с помощью трала НИС “Академик Топчиев”. Рыбу отбирали на станциях Рыбинского водохранилища, различающихся по степени промышленного загрязнения (рис. 1).
Рис. 1.
Карта-схема Рыбинского водохранилища. Станции: 1 – Коприно, 2 – Волково, 3 – Первомайка, 4 – Брейтово, 5 – Городок, 6 – Ягорба, 7 – Мякса, 8 – Любец.
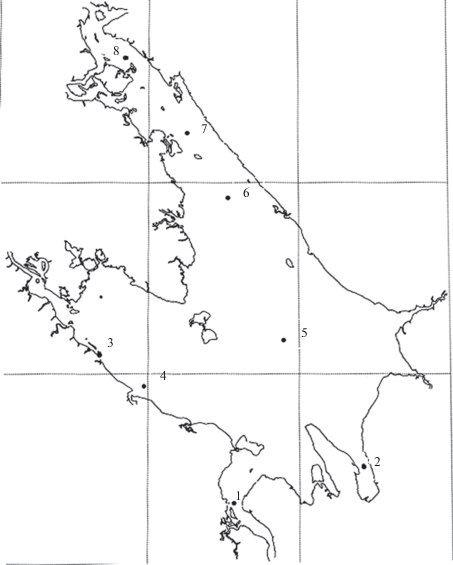
Выбор станций, где проводили отлов лещей обусловлен многолетними исследованиями ряда авторов. Станции 1–5 находились в условно чистых районах Волжского, Моложского и Главного плёсов, станции 6–8 – в относительной близости от г. Череповца, в зоне продолжительного локального промышленного загрязнения.
Глубина на станциях колебалась от 10 до 13 м и лишь на ст. 2 достигала 18 м.
На каждой станции отлавливали по 10 половозрелых и примерно одноразмерных и близких по массе лещей (табл. 1). Рыбу доставляли в лабораторию судна, где проводили биологический анализ. Коэффициенты упитанности по Фультону рассчитывали по работе (Флёрова, 2014). Для изучения кормовой базы леща на каждой станции в местах вылова рыбы собирали макрозообентос (Безматерных, Щербина, 2017). На всех станциях донные грунты были представлены серыми илами, на которых основной биоценоз – Chironomus f. l. plumosus (Перова, Щербина, 1998). Величину кормности участков на водохранилище определяли по работе (Пидгайко и др., 1968).
Таблица 1.
Основные характеристики, ЧИП и ОИП лещей, биомасса кормового зообентоса на различных станциях Рыбинского водохранилища
| Показатель | Номер станции | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| Возраст рыб | +8–+11 | +8–+11 | +8–+11 | +8–+12 | +8–+11 | +8–+11 | +6–+11 | +6–+10 |
| Длина рыб, см | $\frac{{35.9 \pm 0.7}}{{32.9 - 38.9}}$ | $\frac{{36.7 \pm 0.5}}{{33.1 - 38.1}}$ | $\frac{{35.6 \pm 0.7}}{{32.8 - 39.5}}$ | $\frac{{36.8 \pm 0.7}}{{31.9 - 40.8}}$ | $\frac{{36.1 \pm 0.6}}{{34.1 - 39.8}}$ | $\frac{{35.7 \pm 0.6}}{{32.8 - 38.7}}$ | $\frac{{32.7 \pm 1.2}}{{28.6 - 40.5}}$ | $\frac{{29.1 \pm 1.2}}{{25.2 - 38.0}}$ |
| Масса рыб, г | $\frac{{790 \pm 48}}{{607 - 1060}}$ | $\frac{{913 \pm 32}}{{676 - 1064}}$ | $\frac{{793 \pm 55}}{{606 - 1120}}$ | $\frac{{858 \pm 47}}{{591 - 1129}}$ | $\frac{{828 \pm 48}}{{611 - 1116}}$ | $\frac{{741 \pm 61}}{{615 - 1014}}$ | $\frac{{622 \pm 63}}{{425 - 1040}}$ | $\frac{{397 \pm 63}}{{230 - 929}}$ |
| Коэффициент упитанности по Фультону | 1.69 ± 0.04 | 1.85 ± 0.02 | 1.73 ± 0.03 | 1.71 ± 0.03 | 1.74 | 1.62 ± 0.10 | 1.74 ± 0.04 | 1.57 ± 0.09 |
| Число кишечников с пищей/пустых | 4/6 | 9/1 | 6/4 | 10/0 | 2/8 | 9/1 | 10/0 | 6/4 |
| Биомасса зообентоса, г/м2 | 48.28 | 30.45 | 44.81 | 101.1 | 10.83 | 6.05 | 29.23 | 31.30 |
| Доля в биомассе зообентоса, %: | ||||||||
| хирономиды олигохеты |
73 21 |
52 45 |
54 21 |
84 15 |
0 93 |
23 77 |
80 18 |
71 10 |
| ЧИП, ‱: | ||||||||
| хирономиды* | $\frac{{19.81 \pm 29.96}}{{99.9}}$ | $\frac{{30.39 \pm 15.11}}{{98.3}}$ | $\frac{{36.41 \pm 26.96}}{{99.7}}$ | $\frac{{36.05 \pm 33.31}}{{99.3}}$ | $\frac{{0.22}}{{95.7}}$ | $\frac{{0.36 \pm 0.48}}{{29.8}}$ | $\frac{{25.65 \pm 18.25}}{{99.7}}$ | $\frac{{1.75 \pm 0.94}}{{83.7}}$ |
| моллюски | 0.01 | 0.49 | 0.00 | 0.26 | 0.01 | 0.24 | 0.00 | 0.17 |
| олигохеты | 0.00 | 0.03 | 0.05 | 0.00 | 0.00 | 0.61 | 0.09 | 0.17 |
| ОИП, ‱ | $\frac{{19.82 \pm 29.97}}{{0.89 - 64.50}}$ | $\frac{{30.91 \pm 15.69}}{{6.48 - 50.15}}$ | $\frac{{36.46 \pm 26.98}}{{0.09 - 80.26}}$ | $\frac{{36.31 \pm 33.38}}{{2.92 - 100.71}}$ | $\frac{{0.23}}{{0.05 - 0.41}}$ | $\frac{{1.21 \pm 1.24}}{{0.03 - 4.23}}$ | $\frac{{25.74 \pm 18.29}}{{4.03 - 62.44}}$ | $\frac{{2.09 \pm 1.14}}{{0.21 - 3.22}}$ |
Сбор, хранение, полевую и цифровую обработку материалов по питанию леща проводили согласно существующей методике (Методическое…, 1974). Каждый кишечник леща помещали в морозильную камеру в отдельном полиэтиленовом пакете с указанием места и даты отлова рыбы, ее порядкового номера и массы. Пищевой комок каждой особи просматривали под бинокуляром целиком (все фрагменты беспозвоночных выбирали в камере Богорова). Под микроскопом определяли, измеряли и подсчитывали все потребленные организмы. Массу хирономид восстанавливали по ширине головной капсулы (Безматерных, Щербина, 2015). Массу остальных донных беспозвоночных восстанавливали по таковой из бентосных проб, собранных одновременно с материалом по питанию рыб. ЧИП рассчитывали как отношение восстановленной массы отдельных кормовых объектов к массе рыбы. ОИП определяли, суммируя все ЧИП. При расчете среднего значения ОИП и ЧИП за данный период определяли отношением суммы всех индексов потребления к числу исследованных рыб. Лещей с пустыми кишечниками не учитывали (Баканов и др., 1980). Данные представлены в виде средних значений и их ошибок (M ± m).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
В пищевом спектре леща Рыбинского водохранилища осенью 2013 г. обнаружен 21 вид донных беспозвоночных, из них наиболее широко представлены хирономиды, моллюски и олигохеты – 9, 6 и 3 вида соответственно. Кроме того, зарегистрировано по одному виду остракод, пиявок и ручейников. Наибольшее число видов (9) отмечено в пищевом рационе леща на ст. 6, наименьшее (2) – на ст. 1. На ст. 1, 3, 4 число обнаруженных в составе макрозообентоса видов было существенно выше, чем в пищевом спектре леща. На остальных станциях эти различия незначительны (табл. 2).
Таблица 2.
Частота встречаемости (%) видов донных беспозвоночных в пищевом комке леща на различных станциях Рыбинского водохранилища
| Вид | Номер станции | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| Cincina piscinalis (O.F. Müeller) | – | $\frac{{44.4}}{{1 - 7}}$ | – | – | – | $\frac{{11.1}}{{30}}$ | – | – |
| C. depressa C. Pfeiffer | – | – | – | – | – | – | – | $\frac{{16.7}}{{2 - 8}}$ |
| Euglesa casertana (Poli) | $\frac{{25}}{1}$ | $\frac{{11.1}}{6}$ | – | $\frac{{20}}{{1 - 10}}$ | $\frac{{50}}{1}$ | $\frac{{22.2}}{{2 - 8}}$ | $\frac{{10}}{2}$ | $\frac{{16.7}}{2}$ |
| Neopisidium torguatum (Stelfox) | – | $\frac{{22.2}}{{2 - 6}}$ | – | – | – | – | – | $\frac{{66.7}}{{2 - 12}}$ |
| Pisidium inflatum (Muhlfeld in Porro) | – | – | – | – | – | $\frac{{11.1}}{1}$ | – | – |
| Dreissena polymorpha (Pallas) | – | – | – | $\frac{{10}}{1}$ | – | – | – | – |
| Guistadrilus multisetosus (Smith) | – | – | – | – | $\frac{{11.1}}{3}$ | – | – | |
| Limnodrilus hoffmeisteri Claparede | – | $\frac{{11.1}}{{25}}$ | – | – | – | – | $\frac{{10}}{2}$ | – |
| Tubifex newaensis (Michaelsen) | – | – | – | – | – | $\frac{{11.1}}{4}$ | $\frac{{20}}{1}$ | – |
| Erpobdella octoculata (L.) | – | – | – | – | – | $\frac{{11.1}}{1}$ | – | – |
| Cypria curtifurcata Klie | – | $\frac{{11.1}}{{11}}$ | $\frac{{50}}{{10 - 12}}$ | – | – | – | – | $\frac{{33.3}}{{2 - 58}}$ |
| Brachycentrus subnubilus Curtis | – | – | – | – | – | – | – | $\frac{{16.7}}{1}$ |
| Ablabesmyia monilis (L.) | – | – | $\frac{{16.7}}{1}$ | – | – | – | – | – |
| A. phatta (Eggert) | – | – | – | $\frac{{20}}{{2 - 8}}$ | – | – | – | – |
| Procladius choreus (Meigen) | – | $\frac{{77.8}}{{1 - 3}}$ | $\frac{{50}}{{1 - 7}}$ | $\frac{{80}}{{2 - 27}}$ | – | $\frac{{22.2}}{1}$ | $\frac{{30}}{{1 - 2}}$ | $\frac{{16.7}}{1}$ |
| P. simplicistilus Freeman | – | – | – | – | – | $\frac{{11.1}}{1}$ | – | – |
| Chironomus plumosus (L.) | $\frac{{100}}{{2 - 134}}$ | $\frac{{100}}{{12 - 151}}$ | $\frac{{83.3}}{{65 - 246}}$ | $\frac{{100}}{{28 - 452}}$ | $\frac{{50}}{1}$ | $\frac{{22.2}}{{2 - 7}}$ | $\frac{{100}}{{7 - 172}}$ | $\frac{{100}}{{2 - 5}}$ |
| Cryptochironomus obreptans (Walker) | – | – | $\frac{{66.7}}{8}$ | $\frac{{30.0}}{{14}}$ | – | – | $\frac{{10}}{2}$ | $\frac{{16.7}}{4}$ |
| Einfeldia dissidens (Walker) | – | – | $\frac{{16.7}}{1}$ | – | – | – | – | – |
| Stictochironomus crassiforceps (Kieffer) | – | $\frac{{11.1}}{2}$ | $\frac{{16.7}}{1}$ | – | – | – | – | – |
| Cladotanytarsus wexionensis Brundin | – | – | – | – | $\frac{{50}}{1}$ | – | – | – |
| Всего видов: в пище леща |
2 | 8 | 7 | 6 | 3 | 9 | 6 | 8 |
| в макрозообентосе | 20 | 9 | 29 | 19 | 5 | 7 | 9 | 11 |
Повышенное видовое богатство в спектре питания леща на ст. 6 связано, по-видимому, с кормовыми условиями – минимальная среди всех исследованных станций Рыбинского водохранилища биомасса кормового макрозообентоса (6.05 г/м2) и очень низкая доля хирономид (23%) в его составе (табл. 1).
Хотя число питающихся на ст. 6 лещей достигало 90%, среднее значение ОИП было 1.21‱, а на долю ЧИП хирономид приходилось ~30%. Минимальное значение ОИП (0.23‱) среди всех исследованных станций водохранилища отмечено на ст. 5, где в составе макрозообентоса обнаружены только олигохеты и моллюски. Несмотря на это, коэффициент упитанности по Фультону и доля хирономид в питании леща здесь составили 1.74‱ и ~96% соответственно. В то же время число питающихся рыб на данном участке было самым низким (20%) среди станций (табл. 1). Такие особенности можно объяснить миграцией лещей. Известно, что лещ в водохранилище представлен одной популяцией, состоящей из локальных стад. Они различаются масштабами перемещения в процессе нереста и нагульного периода, и делятся на оседлых особей и мобильных, которые могут перемещаться в поисках пищи на большие расстояния (Герасимов и др. 2015).
Основные причины такой низкой пищевой активности леща на ст. 5, 6, по-видимому, заключаются в повышенном содержании продуктов трансформации загрязняющих веществ (Козловская, Герман, 1997; Флеров и др., 2000; Голованова, 2008; Чуйко и др., 2010), негативно влияющих на развитие донных беспозвоночных. Следует отметить, что наибольшая биомасса кормового макрозообентоса зарегистрирована на станциях, расположенных недалеко от впадающих в водохранилище малых и средних рек: ст. 1 – р. Сутка, ст. 2 – реки Согожа и Ухра, ст. 3 – р. Себла, ст. 4 – р. Сить, ст. 7 – р. Мякса.
На шести станциях водохранилища доля хирономид в составе макрозообентоса достигала 52–84%, в составе пищевого комка леща она была существенно выше – 83.7–99.9% (табл. 1). Наибольшие средние значения ОИП у лещей Рыбинского водохранилища выявлены на двух станциях (ст. 3 и 4) Моложского плеса (рис. 2), который по величине индекса сапробности Пантле–Букка самый чистый (Перова, Щербина, 1998). Высокое значение ОИП отмечено также на приплотинной ст. 2, расположенной на наиболее продуктивном участке Рыбинского водохранилища, здесь отмечены максимальные для водоема биомассы макрозообентоса 143– 198 г/м2 (Щербина, 2002). Относительно высокое среднее значение ОИП лещей на ст. 7 (относится к загрязненному участку Рыбинского водохранилища), связано с тем, что на этой станции высокая биомасса макрозообентоса (~30 г/м2), и в его составе доминировали хирономиды (80%) со значительным преобладанием личинок Chironomus plumosus. Личинки данного вида встречались в пище лещей на всех восьми станциях. Их число в кишечниках леща колебалось от 1 до 452 экз., а на пяти станциях частота их встречаемости была максимальной – 100% (табл. 2). Все это подтверждает вывод о доминировании мотыля в питании леща Рыбинского водохранилища на различных биотопах с максимальным значением в осенний период (Житенева, 1980). При изучении питания леща Рыбинского водохранилища осенью 2000 г. обнаружено 13 пищевых компонентов, средняя доля хирономид была 58%, по 21% приходилось на группы “моллюски” и “прочие”. Причем, число личинок Ch. plumosus в кишечниках лещей колебалась от 1 до 82 экз. Незначительное число потребленных личинок мотыля связано, по-видимому, с небольшим числом (6 экз.) исследованных в этот период рыб (Щербина, 2012).
Рис. 2.
Величина общего индекса потребления (‱) на различных станциях Рыбинского водохранилища. 1 – хирономиды, 2 – моллюски, 3 – олигохеты.
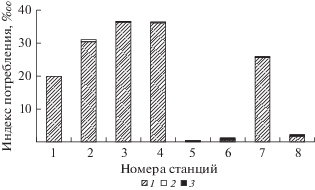
Рис. 3.
Доля (%) основных групп макрозообентоса в питании леща на различных станциях Рыбинского водохранилища. Обозначения, как на рис. 2.

Выводы. При изучении питания леща на восьми станциях Рыбинского водохранилища установлено, что на семи из них хирономиды значительно преобладали в его пищевом рационе, и только на малопродуктивной ст. 6 доминировала группа “прочие”, в основном, представленная олигохетами. В целом, среднее по водоему значение ЧИП хирономид у леща Рыбинском водохранилище осенью 2013 г. было 77%, что выше, чем в аналогичный период 2000 г. 58%. Среди хирономид существенно преобладали личинки и куколки Ch. f. l. plumosus, частота их встречаемости на большинстве станций была максимальной.
На условно чистых станциях водохранилища ОИП и ЧИП хирономид значительно превышают таковые на загрязненных станциях Шекснинского плеса, подверженного сбросу сточных вод с расположенных в Череповецком промышленном комплексе предприятий. Минимальные размеры, массу и коэффициент упитанности по Фультону исследованных лещей наблюдали на ст. 8, расположенной ближе всего к г. Череповец.
Список литературы
Баканов А.И., Стрижникова Л.Н. 1979. О связи изменения кормовой базы и питанием леща Abramis brama orientalis Berg в Волжском плесе Рыбинского водохранилища // Вопр. ихтиол. Т. 19. № 1(114). С. 134.
Баканов А.И., Краснопер Е.В., Стрижникова Л.Н. 1980. Об использовании индексов при изучении питания рыб-бентофагов // Ин-т биол. внутр. вод АН ССCР. Рукоп. деп. в ВИНИТИ. № 4984-80-деп.
Безматерных В.В., Щербина Г.Х. 2015. Размерно-массовая характеристика старших возрастов личинок наиболее распространенных видов хирономид (Diptera, Chironomidae) в бассейне Верхней Волги // Биология внутр. вод. № 2. С. 105.https://doi.org/10.1134/S1995082915010046
Безматерных В.В., Щербина Г.Х. 2017. Об оптимальном числе проб для оценки количественных показателей макрозообентоса на участке водоема // Биология внутр. вод. № 3. С. 83. https://doi.org/10.1134/S199508291703004X
Герасимов Ю.В., Бражник С.Ю., Стрельников А.С. и др. 2015. Лещ // Рыбы Рыбинского водохранилища: популяционная динамика и экология. Ярославль: Филигрань. С. 198.
Герман А.В., Козловская В.И. 2001. Гепатосоматический индекс и биохимический состав печени леща Abramis brama L. Шекснинского плеса Рыбинского водохранилища при различных уровнях накопления органических токсикантов // Вопр. ихтиол. Т. 41. № 2. С. 249.
Герман А.В., Козловская В.И. 1999. Содержание полихлорированных бифенилов в леще Abramis brama Рыбинского водохранилища // Вопр. ихтиол. Т. 39. № 1. С. 139.
Герман А.В., Законнов В.В. 2003. Аккумуляция полихлорированных бифенилов в Шекснинском плесе Рыбинского водохранилища // Вод. ресурсы. Т. 30. № 5. С. 571.
Голованова И.Л. 2008. Влияние тяжелых металлов на физиолого-биохимический статус рыб и водных беспозвоночных // Биология внутр. вод. № 1. С. 99.https://doi.org/10.1007/s12212-008-1014-1
Житенева Т.С. 1958. О питании леща в Рыбинском водохранилище // Тр. биол. ст. “Борок”. Вып. 3. С. 25.
Житенева Т.С. 1959. Питание молоди леща в Рыбинском водохранилище // Тр. Инст. биол. водохр. Т. 1(4). С. 259.
Житенева Т.С. 1980. Питание леща на разных биотопах Рыбинского водохранилища // Биология внутр. вод: Информ. бюл. № 48. С. 37.
Заботкина Е.А. 2014. Оценка здоровья леща Abramis brama L. из разных по уровню загрязнения плесов Рыбинского водохранилища по цито- и гистологическим показателям селезенки // Матер. V Всерос. конф. по водной экотоксикологии “Антропогенное влияние на водные организмы и экосистемы”. Ярославль: Филигрань. С. 52.
Ключарева О.А. 1960. Питание бентосоядных рыб Рыбинского водохранилища // Тр. Дарвинского заповед. Вып. 6. С. 159.
Козловская В.И., Герман А.В. 1997. Полихлорированные бифенилы и полиароматические углеводороды в экосистеме Рыбинского водохранилища // Вод. ресурсы. Т. 24. № 5. С. 563.
Методическое пособие по изучению питания и пищевых отношений рыб в естественных условиях. 1974. Москва: Наука.
Перова С.Н., Щербина Г.Х. 1998. Сравнительный анализ структуры макрозообентоса Рыбинского водохранилища в 1980 и 1990 гг. // Биология внутр. вод. № 2. С. 52.
Пидгайко М.П., Александров Б.М., Иоффе Ц.И. и др. 1968. Краткая биолого-продукционная характеристика водоемов Северо-Запада // Изв. ГосНИИ озер. и реч. рыб. хоз-ва. Т. 68. С. 205.
Флеров Б.А., Томилина И.И., Кливленд Л. и др. 2000. Комплексная оценка состояния донных отложений Рыбинского водохранилища // Биол. внутр. вод. № 2. С. 148.
Флёрова Е.А. 2014. Физиолого-биохимические методы исследования рыб. Учебно-методическое пособие. Ярославль: Изд-во Ярославская гос. c.-х. акад.
Чуйко Г.М., Законнов В.В., Морозов А.А. и др. 2010. Пространственное распределение и качественный состав полихлорированных бефинилов (ПХБ), хлорорганических пестицидов (ХОП) в донных отложениях и леще (Abramis brama L.) Рыбинского водохранилища // Биол. внутр. вод. № 2. С. 98.https://doi.org/10.1134/S199508291002015X
Щербина Г.Х. 2002. Сравнительный анализ структуры макрозообентоса на участках верхнего и нижнего бьефов Рыбинского гидроузла // Биология внутр. вод. № 3. С. 44.
Щербина Г.Х. 2012. Роль донных макробеспозвоночных в питании рыб-бентофагов в Волжских водохранилищах // Матер. Всерос. конф. “Бассейн Волги в ХХI веке: структура и функционирование экосистем водохранилищ”. Ижевск: Пермяков С.А. С. 366.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод