

Биология внутренних вод, 2021, № 6, стр. 574-585
Многолетняя динамика хлорофилла в планктоне различных участков крупного равнинного водохранилища
Н. М. Минеева *
Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл., Россия
* E-mail: mineeva@ibiw.ru
Поступила в редакцию 12.01.2021
После доработки 03.06.2021
Принята к публикации 21.06.2021
Аннотация
По данным натурных наблюдений 2009–2019 гг. рассматривается сезонная и многолетняя динамика хлорофилла на шести стандартных станциях Рыбинского водохранилища. Определение хлорофилла выполнено флуоресцентным методом. Среднее для станций содержание ƩХл изменялось от минимальных 3.5–10.8 до максимальных 16.9–40.5 мкг/л в отдельные годы и от 12.0 на ст. 5 (Средний Двор) до 21.6 мкг/л на ст. 2 (Молога) в среднем за весь период. Сезонная динамика ƩХл характеризуется весенним, летним и, в отдельные годы, осенним максимумами. При однотипной сезонной динамике сезонный ход ƩХл на станциях в разной степени скоррелирован в годы наблюдения и чаще всего изменяется синхронно на соседних участках. Существенные различия количества хлорофилла на станциях прослеживаются при съемках, выполненных в течение одних суток, и обусловлены сложной гидрологической структурой водохранилища, активными динамическими процессами, а в весенний период – еще и режимом прогрева. Наличие устойчивых крупных циркуляционных зон сглаживает пространственные различия хлорофилла, среднесезонные концентрации которого в годы со средней водностью достоверно не различаются на всех шести станциях, а в экстремально многоводные годы – на четырех станциях Главного плеса. Тесная корреляционная связь в многолетней динамике выявлена для средних за вегетационный сезон концентраций ƩХл на станциях. Результаты работы подтверждают надежность полученных данных для оценки экологического состояния Рыбинского водохранилища.
ВВЕДЕНИЕ
Роль фитопланктона в функционировании пресноводных экосистем и формировании их биологической продуктивности хорошо известна. Развитие водорослей определяет кормовую базу, продукционный потенциал биотопов и экологическое состояние водных объектов (Винберг, 1960; Китаев, 2007). К наиболее актуальным аспектам исследований фитопланктона относится изучение его пространственной и временной динамики, а также экологических факторов, с которыми непосредственно связанно развитие сообщества (Padisák, 2004). При изучении автотрофного звена водных экосистем широко используют фотосинтетические пигменты – универсальные эколого-физиологические характеристики развития и фотосинтетической активности водорослей, обладающие высокой информативностью. Определение пигментов служит полезным инструментом для исследования длительных трендов в развитии фитопланктона и оценки состояния пресных, морских и океанических вод. Многолетние ряды наблюдений получены для многих водоемов мира (Ruggiu et al., 1998; Kangur et al., 2002; Chen et al., 2003; Babanazarova, Lyashenko, 2007; Mendesab et al., 2011; Canfield et al., 2018; Lamont et al., 2019; Gao et al., 2020 и др.).
Исследования растительных пигментов в воде Рыбинского водохранилища проводят в Институте биологии внутренних вод РАН с середины XX в. (Фитопланктон…, 1999; Экологические…, 2001; Структура…, 2018). Обширные данные, полученные стандартным спектрофотометрическим методом (SCOR-UNESCO, 1966), с 2009 г. дополняются флуоресцентной диагностикой (Минеева, 2016; Минеева, Семадени, 2020), позволяющей проводить определение хлорофилла непосредственно в природной воде, оценивать ряд характеристик фитопланктона без воздействия на его целостность и оперативно анализировать большой объем материала. Все эти годы исследования проводят на шести постоянных станциях, используя полученные данные как основу для анализа состояния и сукцессионных изменений экосистемы водохранилища. В этой связи важно знать, в какой степени сопряжено развитие фитопланктона различных участков акватории и как оно отражает ситуацию в водоеме в целом. Сравнительный анализ сезонной и многолетней динамики хлорофилла на стандартных станциях Рыбинского водохранилища определил цель настоящей работы.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материал собран на шести станциях Волжского и Главного плесов Рыбинского водохранилища в течение вегетационных сезонов 2009–2019 гг. с периодичностью 1–2 раза в месяц. Пробы отбирали метровым батометром Элгморка из слоев водной толщи 0–2, 2–6 и 6 м – дно. В работе использовано среднее для этих слоев содержание хлорофилла, которое определяли флуоресцентным методом (Гольд и др., 1986). Метод позволяет оценивать суммарное количество пигмента (ƩХл) по его содержанию у основных представителей пресноводного фитопланктона – цианопрокариот (ХлCyan), диатомовых (ХлBac) и зеленых (ХлChl) водорослей. Флуоресценцию хлорофилла измеряли на стационарном флуориметре ПФЛ-3004, изготовленном в Красноярском госуниверситете. Процедура анализа описана ранее, результаты флуоресцентного определение хлорофилла хорошо совпадают с результатами стандартного спектрофотометрического метода (Минеева, 2016).
На всех станциях измеряли прозрачность, температуру и цветность воды. Для оценки вклада фитопланктона в суммарное количество взвешенного вещества использовали соотношение прозрачности, измеренной диском Секки (ZС) и рассчитанной по содержанию хлорофилла (ZХл) (ZХл = 5.7 ƩХл–0.44 (Бульон, 1985)). Для характеристики светового статуса водорослей вычисляли глубину эвфотной зоны (Zeu), превышающую ZС в 2.6 раза (Минеева, 2009), а также соотношение Zeu и слоя перемешивания Hmix, за который в условиях гомотермии принимали глубину станций.
При статистической обработке данных для расчета средних показателей, их погрешностей, корреляционного, регрессионного и дисперсионного анализа, построения графиков использовали стандартные программные пакеты для персонального компьютера. Для оценки тесноты связи между переменными рассчитывали коэффициент ранговой корреляции Спирмена.
Рыбинское водохранилище, третья ступень волжского каскада, – крупный относительно мелководный водоем замедленного водообмена (средний коэффициент условного водообмена 1.9 год–1), расположенный в подзоне южной тайги (58°00′–59°05′ с.ш., 37°28′–39°00′ в.д.). При площади зеркала 4500 км2 и средней глубине 5.6 м их соотношение (~800) свидетельствует о высокой степени открытости водоема (Китаев, 2007). Акваторию водохранилища подразделяют на четыре разнородных участка (плеса), занятых водными массами со специфическими гидрофизическими и гидрохимическими характеристиками. Три плеса расположены по затопленным руслам основных притоков – рек Волга, Молога и Шексна. Речные воды постепенно трансформируются в водную массу собственно водохранилища, которая заполняет озеровидную центральную часть – Главный плес, составляющий ~70% общей площади (Рыбинское…, 1972).
Годы наблюдения в многолетнем ряду в целом характеризовались как теплые, но существенно различались погодными особенностями (Структура…, 2018). Температура воды в мае–октябре 2016–2019 гг. близка к средней многолетней (13.6°С), а в 2009–2015 гг. превышала средние показатели. Максимальная летняя температура воды в основном была 20–24°С, достигая аномально высоких значений 25–27°С в 2010 и 2018 гг. По условиям водности 10 из 11 лет наблюдения относились к многоводным с объемом притока, существенно превосходящим норму в 2017 г., а в экстремально маловодном 2014 г. приток резко сократился. Уровень водохранилища превышал отметку НПУ 101 м БС в 2009–2012, 2016 и 2017 гг. и был очень низким в 2014 г. (http://www.rushydro.ru/hydrology/informer/?date).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Стандартные станции охватывают основную часть акватории Рыбинского водохранилища (рис. 1) и отражают переход речных вод в водную массу водохранилища. Более глубокие станции расположены на затопленных руслах рек Волги (ст. 1, 2), Шексны (ст. 5) и Мологи (ст. 6), ст. 3, 4 – в центральной части Главного плеса с глубиной, близкой к средней для водохранилища. Средние за период исследования гидрологические и гидрохимические показатели на станциях характеризуются близкими величинами (табл. 1). На общем фоне более высокой температурой, прозрачностью, минерализацией и содержанием биогенных веществ выделяется ст. 1, принимающая воды Иваньковского и Угличского водохранилищ и сохраняющая проточные условия. Минимальная прозрачность отмечена на ст. 6, которой достигают наиболее окрашенные и наименее минерализованные моложские воды, а самое высокое содержание общего азота из-за вероятного влияния Череповецкого промышленного комплекса – на ст. 5. Средняя глубина фотосинтезирующего слоя ~3 м и уступает глубине перемешивания в 4.1–4.9 раза на русловых станциях и в 2.1–2.7 раза на станциях поймы. Отношение ZС/ZХл, которое значительно < 1, свидетельствует о невысоком вкладе водорослей в общее содержание взвешенного вещества (табл. 1). Коэффициенты вариации перечисленных показателей ≤35%.
Рис. 1.
Карта-схема Рыбинского водохранилища с расположением станций наблюдения. Цифры – номера станций (Коприно, Молога, Наволок, Измайлово, Средний Двор, Брейтово – 1–6 соответственно), линии – границы плесов.

Таблица 1.
Характеристика стандартных станций Рыбинского водохранилища
| Показатель | Коприно (1) | Молога (2) | Наволок (3) | Измайлово (4) | Средний двор (5) | Брейтово (6) |
|---|---|---|---|---|---|---|
| Координаты c.ш. | 58°04.23′ | 58°12.47′ | 58°22.38′ | 58°27.44′ | 58°30.31′ | 58°19.20′ |
| в.д. | 38°17.56′ | 38°27.34′ | 38°23.22′ | 38°28.47′ | 38°21.01′ | 37°56.58′ |
| Глубина, м | 11.6 ± 0.1 | 12.9 ± 0.1 | 7.5 ± 0.1 | 5.9 ± 0.1 | 13.1 ± 0.2 | 12.2 ± 0.1 |
| Температура, °С | 16.1 ± 0.5 | 15.4 ± 0.6 | 15.1 ± 0.6 | 15.2 ± 0.6 | 15.2 ± 0.6 | 16.0 ± 0.6 |
| ZС, м | 1.19 ± 0.04 | 1.15 ± 0.04 | 1.12 ± 0.03 | 1.13 ± 0.03 | 1.13 ± 0.03 | 1.02 ± 0.03 |
| Цветность, град | 55 ± 2 | 55 ± 2 | 57 ± 2 | 55 ± 1 | 55 ± 1 | 68 ± 3 |
| Zeu, м | 3.0 ± 1.0 | 3.0 ± 1.8 | 2.9 ± 1.6 | 2.9 ± 1.3 | 2.9 ± 1.3 | 2.6 ± 1.3 |
| Hmix/Zeu, отн. ед. | 4.1 ± 0.1 | 4.7 ± 0.1 | 2.7 ± 0.1 | 2.1 ± 0.1 | 4.8 ± 0.2 | 4.9 ± 0.1 |
| ZС/ZХл, отн. ед. | 0.54 ± 0.24 | 0.66 ± 0.28 | 0.57 ± 0.19 | 0.61 ± 0.19 | 0.54 ± 0.22 | 0.54 ± 0.23 |
| Общий азот, мг/л* | 0.95 | 0.98 | 0.76 | 0.79 | 1.08 | 0.83 |
| Общий фосфор, мкг/л* | 95 | 62 | 50 | 54 | 60 | 60 |
| Минерализация, мг-экв./л** | 4.88 | 4.54 | 4.48 | 4.50 | 4.47 | 4.29 |
Содержание ƩХл на всех станциях изменяется от <1 до 50–130 мкг/л, средние для отдельных лет величины соответственно от минимальных 3.5–10.8 до максимальных 16.9–40.5 мкг/л, осредненные за весь период наблюдения показатели – от 12.0 до 21.6 мкг/л (табл. 2).
Таблица 2.
Содержание хлорофилла на стандартных станциях Рыбинского водохранилища в 2009–2019 гг.
| Номерстанции | ƩХл | ХлCyan | ХлBac | ХлChl | |||
|---|---|---|---|---|---|---|---|
| мкг/л | мкг/л | % | мкг/л | % | мкг/л | % | |
| 1 | 15.3 ± 1.7 (104) | 8.5 ± 1.3 (151) | 55.6 | 6.4 ± 0.9 (137) | 41.8 | 0.4 ± 0.1 (148) | 2.6 |
| 2 | 21.6 ± 2.1 (95) | 12.2 ± 1.4 (112) | 56.5 | 8.8 ± 1.3 (142) | 40.7 | 0.6 ± 0.2 (284) | 2.8 |
| 3 | 14.6 ± 1.4 (89) | 10.1 ± 1.3 (117) | 69.2 | 4.1 ± 0.7 (165) | 28.1 | 0.4 ± 0.1 (181) | 2.7 |
| 4 | 15.5 ± 1.2 (75) | 11.2 ± 1.1 (94) | 72.2 | 3.9 ± 0.6 (141) | 25.2 | 0.4 ± 0.1 (146) | 2.6 |
| 5 | 12.0 ± 0.9 (74) | 8.9 ± 0.9 (98) | 74.2 | 2.9 ± 0.3 (100) | 24.2 | 0.3 ± 0.0 (128) | 2.6 |
| 6 | 15.3 ± 1.3 (80) | 9.7 ± 1.2 (113) | 63.4 | 5.3 ± 0.5 (83) | 34.6 | 0.4 ± 0.1 (136) | 2.5 |
Примечание. Приведены средние величины со стандартной ошибкой. В скобках – коэффициент вариации, %. Номера станций в соответствии с табл. 1.
Самыми низкими концентрациями ƩХл характеризуется ст. 5, самыми высокими – ст. 2. На каждой станции отмечена значительная вариабельность ƩХл при коэффициентах вариации Cv ~ 100% в Волжском плесе и 74–90% в Главном. Судя по частоте встречаемости, на ст. 1, 3, 5 преобладают концентрации ƩХл < 10 мкг/л, типичные для мезотрофных вод, на ст. 2, 4, 6 – концентрации 10–30 мкг/л, типичные для эвтрофных условий. Величины >30 мкг/л составили небольшую часть общей выборки (7–12%) в Главном плесе и более заметную долю (23%) в Волжском плесе на ст. 2 (рис. 2). В отдельные сроки наблюдения содержание ƩХл в разной степени менялось по акватории водохранилища. Чаще всего (в 62% случаев по данным 91 съемки) максимальная и минимальная величины различались в 2–5 раз, реже (29%) – в 6–10 раз, в 9% случаев более существенно.
Рис. 2.
Частота встречаемости концентраций хлорофилла (ƩХл) на стандартных станциях Рыбинского вдхр. в 2009–2019 гг. (% общего числа наблюдений n). ◻ –<10; ■ – 10–30; ■ – >30 мкг/л. На ст. 1, 2 – n = 276, ст. 3, 5, 6 – n = 269, ст. 4 – n = 176.

Содержание хлорофилла основных отделов водорослей также характеризуется широким диапазоном значений (табл. 2). Коэффициенты вариации ХлCyan, ХлBac и ХлChl за небольшим исключением >100%. Среднее для станций количество ХлCyan меняется от 8.5 до 12.2 мкг/л, ХлBac – от 2.9 до 8.8 мкг/л, ХлChl представлен сходными низкими величинами. Максимальные значения всех перечисленных показателей получены на ст. 2. Содержание ХлCyan и ХлBac определяет общее количество хлорофилла, составляя в среднем 55.6–74.4 и 24.2–41.8% его суммарного фонда (табл. 2). Этот вклад меняется в межгодовом и сезонном аспекте. Существенную (>90%) часть фонда ƩХл весной и поздней осенью может составлять ХлBac, а в летние месяцы и в начале осени – ХлCyan, сезонная динамика которых не скоррелирована (Минеева, Семадени, 2020).
При значительном диапазоне концентраций ƩХл их распределение в сезонном цикле в годы исследования ассиметрично на всех станциях. Это отражают планки погрешностей на диаграммах размаха, а также медианы, которые отличаются от среднего арифметического в 1.3–1.5 раза (рис. 3, табл. 2). Выброс данных за пределы нижнего 25% квартиля в большинстве случаев невелик и наиболее заметен только в 2013 г. на ст. 1–4 и 6, в 2014 г. на ст. 2. Выброс за пределы верхнего 75%-ного квартиля гораздо существеннее, особенно на ст. 1, 3, 4, 6 в 2011 г., на ст. 1–3, 5, 6 в 2013 г. и на ст. 1, 3–6 в 2018 г.
Рис. 3.
Диаграммы размаха концентраций хлорофилла на станциях 1–6 (а–е, соответственно) в 2009–2019 гг. 1 – границы 25%-ного и 75%-ного квартилей, 2 – медиана, 3 – максимум, 4 – минимум.

Сезонная динамика хлорофилла характеризуется чередованием подъемов и спадов. Весенний максимум ƩХл со средними за весь период наблюдения концентрациями 24–34 мкг/л на станциях 1–4 и 13–14 мкг/л на ст. 5, 6 формируется в мае при температуре воды от 15.3°C на ст. 1 до 6.8°C на ст. 5 (рис. 4). На всех шести станциях одновременно он отмечен только в 2012 г., на двух станциях Волжского плеса и ст. 6 – также в 2013, 2015, 2016, 2018 гг., на двух-трех станциях центральной части – в 2016, 2017, 2019 гг. В ряде случаев весной зафиксированы и низкие (3–5 мкг/л) концентрации ƩХл. Они наблюдались на всех станциях в 2010 г., на центральных – в 2015 г., в южной и западной части в 2017 г. и на самой северной ст. 5 в 2013, 2015, 2018 гг. Температура воды в эти периоды в основном составляла 5.5–10.1°C, а в 2010 г. в центре водохранилища – ~3°C. Ранним летом в июне при температуре 16–17°C содержание ƩХл снижалось до 9–11 мкг/л на ст. 1, 2, 6 и до 7–8 мкг/л на остальных участках.
Рис. 4.
Сезонная динамика хлорофилла на стандартных станциях Рыбинского водохранилища в 2009–2019 гг: а – станции 1, 2; б – ст. 3, 4; в – ст. 5, 6.
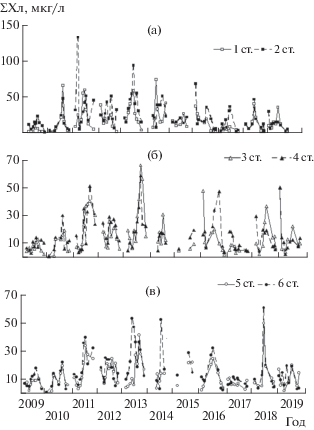
В период прогрева до 20°C и выше в июле–августе формируется летний максимум фитопланктона со средними за все годы концентрациями ƩХл 35–40 мкг/л на ст. 1, 2, 6 и 26–30 мкг/л на ст. 3–5. Летний максимум в основном превалирует в сезонном цикле, но в отдельные годы (чаще в 2016–2019 гг.) уступает весеннему, а в 2009, 2017 и 2019 гг. летний подъем не наблюдался (рис. 4). В конце лета – начале осени при начавшемся выхолаживании водной массы содержание пигмента сначала снижается незначительно, а при температуре воды ниже 10°C достигает минимальных величин 2–7 мкг/л. Однако поздней осенью могут наблюдаться и высокие показатели: в октябре 2014 г. – 23 и 41 мкг/л в Волжском плесе при температуре 7–10°C, в октябре 2018 г. – 10–15 мкг/л при такой же температуре на ст. 2, 3, 5, а на ст. 4 в ноябре при 3.6°C.
В целом на всех станциях наблюдается однотипная сезонная динамика фитопланктона, которая характеризуется сменой подъемов и спадов ƩХл. Однако эти явления происходят не синхронно, и изменение ƩХл на станциях в течение вегетационного сезона в разной степени сопряжено в годы наблюдения. Максимальное число достоверных связей (8–10 из 15 возможных для каждого года) выявлено в 2010, 2012 и 2018 гг., минимальное (3 и 1) – в 2009 и 2017 гг., промежуточное (5–7) в остальные годы, а самое большое количество высоких коэффициентов корреляции >0.7 (9 из 10) – в 2018 г. Чаще всего (в 5–7 случаях из 11 лет наблюдения) сезонный ход хлорофилла скоррелирован на сопредельных (соседних) участках водохранилища, о чем свидетельствуют достоверные коэффициенты корреляции, которые получены для ст. 2 и ст. 3, 6; для ст. 3 и ст. 4, 5, 6 (табл. 3).
Таблица 3.
Годы наблюдения, в которые получены достоверные коэффициенты корреляции между содержанием хлорофилла на станциях Рыбинского водохранилища
| Станции | Годы наблюдения |
|---|---|
| 1–2 | 2010, 2013, 2016, 2018 |
| 1–3 | 2010, 2011, 2016, 2018 |
| 1–4 | 2010 |
| 1–5 | 2018 |
| 1–6 | 2012, 2013, 2015, 2018 |
| 2–3 | 2010, 2012, 2014, 2015, 2016, 2018, 2019 |
| 2–4 | 2010, 2012, 2015, 2019 |
| 2–5 | 2010, 2012, 2014, 2018 |
| 2–6 | 2009, 2012, 2013, 2014, 2018 |
| 3–4 | 2010, 2011, 2012, 2013, 2015, 2019 |
| 3–5 | 2010, 2011, 2012, 2013, 2014, 2015, 2018, 2019 |
| 3–6 | 2009, 2010, 2011, 2012, 2014, 2017, 2018 |
| 4–5 | 2011, 2013, 2015 |
| 4–6 | 2016 |
| 5–6 | 2009, 2011, 2014, 2015, 2016, 2018, 2019 |
При отсутствии устойчивой синхронной сезонной динамики хлорофилла на разных участках водохранилища, результаты кластерного анализа показывают достаточно тесную связь количественного развития фитопланктона на станциях в годы исследования (рис. 5). Единый кластер чаще всего образуют станции Главного плеса (в 2010–2012, 2017, 2019 гг. – все, в 2013–2015 – ст. 3–5), расположенные в центральной части водохранилища и только в 2010 г. – две станции Волжского плеса. Распространенa ситуация, когда особняком отстоит одна станция, в основном – ст. 1, а также ст. 2, 4, 6; ст. 2 часто примыкает к станциям центральной части водохранилища; в 2013 и 2015 гг. с волжскими станциями объединяется ст. 6.
Рис. 5.
Дендрограмма сходства стандартных станций Рыбинского водохранилища по содержанию хлорофилла в 2009–2019 гг. (а–л). По оси ординат – евклидово расстояние, по оси абсцисс – номера станций.

Несмотря на специфику сезонной динамики и количественного развития фитопланктона на разных участках водохранилища, результаты дисперсионного анализа демонстрируют отсутствие значимых различий содержания ƩХл на станциях для всех лет наблюдения (табл. 4). Различия выявляются лишь в 2009 и 2017 гг. при контрастных концентрациях ƩХл на станциях Волжского плеса, но становятся незначимыми при исключении этих станций из расчетов. Сравнение средних за вегетационный сезон концентраций ƩХл на станциях показывает наличие тесной связи между ними в многолетней динамике. Лишь в трех случаях коэффициенты корреляции <0.70 (табл. 5). Содержание ƩХл на ст. 1 объясняет 76% вариации среднего показателя для водохранилища, на ст. 2 и ст. 6 – 85 и 88%, на ст. 3–5 – 53–59%.
Таблица 4.
Результаты сравнения средних за вегетационный сезон концентраций хлорофилла на стандартных станциях Рыбинского водохранилища в годы исследования с помощью однофакторного дисперсионного анализа (ANOVA)
| Год | Станции | SS | df | F | p |
|---|---|---|---|---|---|
| 2009 | 1–6 | 1090 | 53 | 3.53 | 0.008 |
| 3–6 | 472 | 35 | 0.97 | 0.420 | |
| 2010 | 1–6 | 6694 | 46 | 0.27 | 0.928 |
| 2011 | 1–6 | 24 068 | 53 | 1.48 | 0.213 |
| 2012 | 1–6 | 4820 | 53 | 1.95 | 0.103 |
| 2013 | 1–6 | 20 092 | 59 | 1.87 | 0.116 |
| 2014 | 1–6 | 8154 | 33 | 1.55 | 0.206 |
| 2015 | 1–6 | 988 | 21 | 1.29 | 0.316 |
| 2016 | 1–6 | 9928 | 53 | 0.59 | 0.706 |
| 2017 | 1–6 | 2167 | 53 | 2.51 | 0.049 |
| 3–6 | 471 | 35 | 1.86 | 0.156 | |
| 2018 | 1–6 | 8504 | 53 | 0.08 | 0.996 |
| 2019 | 1–6 | 5383 | 53 | 0.33 | 0.891 |
Таблица 5.
Коэффициент корреляции между средними за вегетационный сезон концентрациями хлорофилла на стандартных станциях
| Станция | Коприно | Молога | Наволок | Измайлово | Средний двор | Брейтово |
|---|---|---|---|---|---|---|
| Коприно | 1.00 | – | – | – | – | – |
| Молога | 0.92 | 1.00 | – | – | – | – |
| Наволок | 0.75 | 0.73 | 1.00 | – | – | – |
| Измайлово | 0.55 | 0.71 | 0.77 | 1.00 | – | – |
| Средний двор | 0.69 | 0.76 | 0.59 | 0.77 | 1.00 | – |
| Брейтово | 0.80 | 0.85 | 0.72 | 0.75 | 0.95 | 1.00 |
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В зависимости от особенностей водного объекта определяющими факторами развития гидробионтов могут быть физические, химические или биотические (Chen et al., 2003; Reynolds, 2006; Yang et al., 2016). Фитопланктон Рыбинского водохранилища не испытывает дефицита биогенных веществ (Минеева и др., 2021), сезонные изменения которых происходят в пределах естественных колебаний и свидетельствуют о стабилизации их режима в последние 40 лет (Степанова и др., 2013). Стандартные станции расположены на морфометрически разнородных участках. При сходстве средних многолетних гидрооптических показателей (табл. 1) прозрачность, цветность и глубина эвфотной зоны значительно меняются в течение вегетационного сезона (Минеева, 2009), и фактически фитопланктон станций обитает при различающихся гидрооптических условиях. Именно световой режим оказывает существенное влияние на развитие и продуктивность альгоценозов и становится лимитирующим фактором при достаточной обеспеченности водорослей биогенным питанием и/или при низкой прозрачности воды (Kalff, 2002; Nascimento Moura et al., 2012; Burson et al., 2018). Световой режим, характеристикой которого служит прозрачность (Довгий, 1977), относится к числу основных факторов, влияющих на многолетнюю динамику фитопланктона Рыбинского водохранилища (Мineeva, 2021).
Соотношение зоны перемешивания (в условиях гомотермии – глубины станции) и эвфотной зоны свидетельствует о разном световом статусе водорослей (доступности для них световой энергии). Для всех без исключения наблюдений это соотношение существенно больше единицы, и ситуация, при которой альгоценоз постоянно находился бы в освещенном слое (Hmix/Zeu = 1) (Bormans et al., 2005), не выявлена ни на одной станции. Прозрачность воды в Рыбинском водохранилище в основном определяется общим количеством взвешенного вещества, в составе которого доля водорослевой компоненты невелика (отношение ZС/ZХл <1) (Минеева, 2009). Поступление взвеси происходит при взмучивании донных отложений в результате ветрового воздействия, которому часто подвергается открытая акватория крупного мелководного водоема. Взмучивание донных отложений меняет не только подводный световой климат, но также и концентрацию питательных веществ (Öterler et al., 2015), обеспечивая их дополнительное поступление в толщу воды. Следовательно, погодные условия (ветровое воздействие) могут быть одним из факторов, влияющих на развитие фитопланктона и, соответственно, вариации трофического статуса водохранилища.
Рыбинское водохранилище – водоем со сложной гидрологической структурой, в котором присутствуют водные массы различного генезиса со специфическими физическими и химическими показателями. К особенностям гидродинамики водохранилища относится наличие ветровых и стоковых течений, а также циркуляционных зон. Морфометрические и морфологические характеристики водоема, взаимодействие водных масс, гидрологические процессы влияют на пространственное распределение организмов (Бакастов, Литвинов, 1971; Рыбинское…, 1972; Литвинов, Ривьер, 1991; Экологические…, 1993). Сложная гидрологическая структура обусловливает макромасштабную неоднородность горизонтального распределения фитопланктона, с которой связано формирование полей с различной продуктивностью и трофностью (Экологические…, 1993; Структура…, 2018). В результате прослеживаются существенные различия количества хлорофилла на станциях при съемках, выполненных в течение одних суток. Аналогичная разница для участков, удаленных друг от друга на расстояние, сопоставимое с расположением стандартных станций Рыбинского водохранилища, отмечена для ряда озер и водохранилищ (Antenucci et al., 2005; Bormans et al., 2005; Rakočević, 2012; Yang et al., 2020).
Пространственная неоднородность указывает на различие факторов, контролирующих рост фитопланктона и ход его сезонной сукцессии на разных участках, что присуще мелководным водоемам с большой площадью акватории (Wetzel, 2001). Локальные погодные условия (поступление солнечной радиации, скорость и направление ветра) в пределах большой акватории могут различаться даже при сходной синоптической ситуации в регионе, а пятнистость фитопланктона – быть результатом пространственных вариаций биологических процессов (роста, выедания, регулируемой плавучести, вертикальных миграций) или адвективного переноса (Wiedner et al., 2002; Reynolds, 2006).
Распределение планктона в Рыбинском водохранилище регулируется наличием крупных циркуляционных зон и более мелких разнонаправленных круговоротов (Буторин и др., 1982; Экологические…, 1993). Согласно наблюдениям и расчетам, перемещение скоплений зоопланктона в водохранилище по направлению интегрального переноса может достигать 7–9 км за месяц с учетом двухслойного течения и быть на порядок больше без учета последнего (Бакастов, Литвинов, 1971). Диаметр скоплений зоопланктона оценивается в 10–12 км, что сопоставимо с размерами ядер основных циркуляционных зон. Наибольшая неоднородность в распределении планктона наблюдается при длительной штилевой погоде (Литвинов, Ривьер, 1991). Циркуляционные потоки, возникающие под действием ветра и рельефа дна, могут выносить в открытую часть скопления водорослей, занесенных на мелководья сгонно-нагонным ветровым воздействием (Экологические…, 1993), что часто наблюдается при массовой вегетации цианопрокариот. Из шести стандартных станций ближе к берегу расположены ст. 4, 6, на которых летний подъем хлорофилла может быть связан с ветровым нагоном. Исходя из схемы интегральной циркуляции вод (Буторин и др., 1982), можно предположить, что ст. 3 находится в зоне взаимодействия трех крупных циркуляционных образований, а ст. 5 – в центре антициклонального вихря в северной части Главного плеса. По-видимому, циркуляционные процессы сглаживают пространственную неоднородность фитопланктона, и содержание хлорофилла на сопредельных участках водохранилища часто скоррелировано. В годы со средней водностью крупные циркуляционные зоны довольно устойчивы в течение вегетационного периода, но могут быть нарушены в экстремальные по водности годы (Буторин и др., 1982). Вероятно, наличие этих зон нивелировало и делало незначимым различия среднесезонных концентраций хлорофилла на станциях в течение девяти из 11 лет наблюдения. Достоверные различия выявлены в 2009 и 2017 гг., первый год из которых на общем фоне характеризовался повышенной, второй – экстремальной водностью с объемом притока в мае–октябре 18.47 и 23.0 км3 соответственно (Mineeva, 2021). В эти годы сезонный ход хлорофилла на станциях скоррелирован слабо.
Среднесезонные концентрации хлорофилла за 11-летний период различались в 2.6–4.7 раза на ст. 2–6 и в 8 раз на ст. 1. Этот участок Волжского плеса, с точки зрения гидрологии, рассматривается как большой залив, где в зависимости от направления ветра происходит или аккумуляция, или рассеяние водорослей (Поддубный и др., 1990). При неравномерном распределении фитопланктона по акватории водохранилища максимальным его обилием (биомассой, количеством ΣХл и хлорофилла основных отделов водорослей) традиционно характеризуется ст. 2 (Структура…, 2018), которая расположена на границе Волжского и Главного плесов и представляет собой зону экотона. Известно, что в зонах трансформации и смешения водных масс в пресноводных и морских экосистемах происходит интенсификация физико-химических и биологических процессов, повышение биомассы, численности и видового разнообразия гидробионтов (Экологические…, 1993; Крылов и др., 2010; Сахарова, 2017; Cadier et al., 2017). Самые низкие концентрации хлорофилла отмечены на ст. 5, расположенной севернее остальных.
Сезонное развитие планктона – ежегодно повторяющийся процесс, испытывающий влияние внешних факторов и внутренних взаимодействий (Reynolds, 2006). В больших неглубоких водоемах, представляющих собой активную динамичную среду, ход сезонной сукцессии фитопланктона подвержен частому нарушающему внешнему воздействию (Honti et al., 2007, Sommer et al., 2012; Yang et al., 2016), к которому относится ветровое перемешивание (Bormans et al., 2005; Nascimento Moura et al., 2012), а в водохранилищах еще и работа гидросооружений. Сезонный ход хлорофилла в Рыбинском водохранилище соответствует классической модели (Sommer et al., 2012) и характеризуется двумя–тремя подъемами. Однако сроки их наступления, продолжительность и концентрации хлорофилла на станциях меняются не только в годы с разными погодными условиями, но не всегда синхронны в один и тот же год (Минеева, 2016; Минеева, Семадени, 2020). Вероятные причины этого обсуждены выше. Развитие весеннего сообщества связано еще и с режимом прогрева, что подтверждается разницей сроков наступления максимумов хлорофилла на станциях и температурой воды, при которой эти максимумы формируются и которая в Волжском плесе выше, чем в Главном (Минеева, 2004).
При отсутствии синхронной динамики фитопланктона на разных участках водохранилища локальные низкие, (чаще высокие) концентрации хлорофилла определяют их ассиметричное распределение в сезонном цикле. Выбросы за пределы верхнего 75%-ного квартиля на диаграммах размаха (рис. 3) обусловлены сезонными максимумами фитопланктона – высокими концентрациями ХлCyan в летне-осенний период, ХлBac весной и поздней осенью. Наиболее выраженные сезонные пики ХлCyan и ХлBac наблюдали в водохранилище в 2011, 2013 и 2018 гг. (Mineeva, 2021), т.е. в годы с большим размахом ƩХл. При низких концентрациях одного из этих компонентов разброс ƩХл уменьшается.
Средние концентрации ƩХл на станциях в основном соответствовали мезотрофной категории в 2009 и 2017 гг., умеренно эвтрофной в 2010 и 2019 гг., эвтрофной в 2011–2014 и 2018 гг., а на ст. 2 в 2011 и 2013 гг. – гипертрофной. В 2015 и 2016 гг. трофический статус участков менялся от мезотрофного до эвтрофного. Межгодовые вариации хлорофилла в зависимости от гидроклиматических условий выявлены для водоемов разных регионов. В высоко эвтрофных они не выходят за пределы своей категории трофности (Babanazarova, Lyashenko, 2007; Rusanov et al., 2020), в умеренно эвтрофных, включая Рыбинское водохранилище, в разные годы наблюдаются величины, присущие разным трофическим типам (Kangur et al., 2002; Минеева, 2016; Минеева, Семадени, 2020).
Оценка внешнего влияния на развитие фитопланктона водохранилища осложняется наличием трудно учитываемых или неконтролируемых факторов, о чем косвенно свидетельствует существенная разница между средними за вегетационный сезон концентрациями хлорофилла и их медианам. В действительности развитие биологических сообществ искусственных водоемов испытывает влияние не только локальных погодных условий и климата, но и режима работы гидросооружений. Максимальное развитие фитопланктона отмечается в маловодные годы при штилевой погоде, повышенных инсоляции и температуре воды, минимальное – при противоположных условиях (Пырина, 2000; Пырина и др., 2006). В отдельные годы (2009, 2010, 2014, 2015, 2018 гг.) выявлена умеренная положительная связь сезонной динамики ƩХл с температурой воды. Для средних за вегетационный сезон концентраций ƩХл прослеживается положительная зависимость от температуры и показателя солнечной активности – чисел Вольфа, а факторами, ограничивающим развитие фитопланктона, выступают условия водности (большое количество осадков и объем притока, высокий уровень воды) (Mineeva, 2021). Поскольку годы исследования различались погодными условиями и водностью, то, вероятно, ведущими становились и разные факторы, определившие специфику развития и сезонной динамики фитопланктона на отдельных участках водохранилища. В результате станции наблюдения по-разному сгруппированы на дендрограммах в годы исследования (рис. 5). Тем не менее, отсутствие значимой разницы содержания ƩХл на станциях, а также тесная корреляционная связь средних за вегетационный сезон величин в многолетней динамике подтверждают надежность полученных данных для оценки экологического состояния Рыбинского водохранилища.
Выводы. Содержание хлорофилла на стандартных станциях Рыбинского водохранилища характеризуется широким диапазоном и значительной вариабельностью величин. Сезонный ход хлорофилла на всех станциях представлен двумя–тремя подъемами, сроки наступления которых, продолжительность и концентрации пигмента меняются не только в годы с разными погодными условиями, но не всегда синхронны в один и тот же год. Существенные различия количества хлорофилла на станциях прослеживаются при съемках, выполненных в течение cуток, и обусловлены сложной гидрологической структурой водохранилища, активными динамическими процессами, а в весенний период – еще и режимом прогрева. Наличие устойчивых крупных циркуляционных зон сглаживает различия пространственно-временной динамики хлорофилла, среднесезонные концентрации которого в годы со средней водностью достоверно не различаются на всех стандартных станциях, а в экстремально многоводные годы – на станциях Главного плеса. Отсутствие значимой разницы среднесезонного содержания ƩХл на станциях в годы наблюдения, а также тесная корреляционная связь средних за вегетационный сезон величин в многолетнем аспекте подтверждают надежность полученных данных для оценки экологического состояния Рыбинского водохранилища.
Список литературы
Бакастов С.С., Литвинов А.С. 1971. Опыт расчета горизонтального переноса сообществ планктона // Тр. Ин-та биол. внутр. вод АН СССР. Вып. 22(25). С. 217.
Бульон В.В. 1985. Закономерности первичной продукции в лимнических экосистемах: Автореф. дис. … докт. биол. наук. Ленинград: Зоол. ин-т АН СССР.
Буторин Н.В., Литвинов А.С., Фомичев И.Ф., Поддубный С.А. 1982. Горизонтальная циркуляция вод в Рыбинском водохранилище и возможные ее изменения при перераспределении стока // Тр. Ин-та биол. внутр. вод АН СССР. Ленинград: Наука. Вып. 43(46). С. 150.
Винберг Г.Г. 1960. Первичная продукция водоемов. Минск: Изд-во АН БССР.
Гольд В.М., Гаевский Н.А., Шатров И.Ю. и др. 1986. Опыт использования флуоресценции для дифференциальной оценки содержания хлорофилла a у планктонных водорослей // Гидробиол. журн. Т. 22. № 3. С. 80.
Довгий Т.Н. 1977. Подводная солнечная радиация на Байкале. Новосибирск: Наука.
Китаев С.П. 2007. Основы лимнологии для гидробиологов и ихтиологов. Петрозаводск: Карельск. науч. центр РАН.
Корнева Л.Г. 2015. Фитопланктон водохранилищ бассейна Волги. Кострома: Дом печати.
Крылов А.В., Цветков А.И., Малин М.И. и др. 2010. Сообщества гидробионтов и физико-химические параметры устьевой области притока равнинного водохранилища // Биол. внутр. вод. № 1. С. 65. https://doi.org/10.1134/S1995082910010086
Литвинов А.С., Ривьер И.К. 1991. Влияние гидрологических процессов на распределение зоопланктона в Рыбинском водохранилище // Вод. ресурсы. № 6. С. 73.
Минеева Н.М. 2004. Растительные пигменты в воде волжских водохранилищ. Москва: Наука.
Минеева Н.М. 2009. Первичная продукция планктона в водохранилищах Волги. Ярославль: Принтхаус.
Минеева Н.М. 2016. Сезонная и межгодовая динамика хлорофилла в планктоне Рыбинского водохранилища по данным флуоресцентной диагностики // Тр. Ин-та биол. внутр. вод им. И.Д. Папанина РАН. Вып. 76(78). С. 75.
Минеева Н.М., Семадени И.В. 2020. Сезонная и межгодовая динамика хлорофилла в планктоне Рыбинского водохранилища (2015–2019 гг.) // Тр. Ин-та биол. внутр. вод им. И.Д. Папанина РАН. Вып. 92(95). С. 12.
Минеева Н.М., Степанова И.Э., Семадени И.В. 2021. Биогенные элементы и их роль в развитии фитопланктона водохранилищ Верхней Волги // Биол. внутр. вод. № 1. С. 24. https://doi.org/5221010095 https://doi.org/10.31857/S032096
Поддубный С.А., Корнева Л.Г., Минеева Н.М. 1990. Влияние горизонтальной циркуляции вод на распределение фитопланктона в Рыбинском водохранилище // Вод. ресурсы. № 2. С. 148.
Пырина И.Л. 2000. Многолетняя динамика и цикличность межгодовых колебаний содержания хлорофилла в Рыбинском водохранилище // Озерные экосистемы: биологические процессы, антропогенная трансформация, качество воды: Матер. междунар. науч. конф. Минск: Белорус. гос. ун-т. С. 375.
Пырина И.Л., Литвинов А.С., Кучай Л.А. и др. 2006. Многолетние изменения первичной продукции фитопланктона в Рыбинском водохранилище в связи с действием климатических факторов // Состояние и проблемы продукционной гидробиологии. Москва: КМК. С. 38.
Рыбинское водохранилище и его жизнь. 1972. Ленинград: Наука.
Сахарова Е.Г. 2017. Фитопланктон экотонных зон Рыбинского водохранилища: Автореф. дис. … канд. биол. наук. Борок.
Степанова И.Э., Бикбулатова Е.М., Бикбулатов Э.С. 2013. Закономерности динамики содержания биогенных элементов в водах Рыбинского водохранилища за годы его существования // Вода: химия и экология. № 1. С. 15. https://doi.org//watchemec.ru/article/25349
Структура и функционированием экосистемы Рыбинского водохранилища в начале XXI века. 2018. Москва: РАН.
Фитопланктон Волги. Экология фитопланктона Рыбинского водохранилища. 1999. Тольятти: Самар. науч. центр РАН.
Экологические проблемы Верхней Волги. 2001. Ярославль: Ярослав. гос. тех. ун-т.
Экологические факторы пространственного распределения и перемещения гидробионтов. 1993. Санкт-Петербург: Гидрометеоиздат.
Antenucci J.P., Ghadouani A., Burford M.A. Romero J.R. 2005. The long-term effect of artificial destratification on phytoplankton species composition in a subtropical reservoir // Freshwater Biol. V. 50. № 6. P. 1081. https://doi.org/10.1111/j.1365-2427.2005.01374.x
Babanazarova O.V., Lyashenko O.A. 2007. Inferring long-term changes in the physical–chemical environment of the shallow, enriched Lake Nero from statistical and functional analyses of its phytoplankton // J. Plankton Res. V. 29. № 9. P. 747. https://doi.org/10.1093/plankt/fbm055
Bormans M., Ford P.W., Fabbro L. 2005. Spatial and temporal variability in cyanobacterial populations controlled by physical processes // J. Plankton Res. V. 27. № 1. P. 61.
Burson A., Stomp M., Greenwell E. et al. 2018. Competition for nutrients and light: testing advances in resource competition with a natural phytoplankton community // Ecology. V. 99. № 5. P. 1108. https://doi.org/10.1002/ecy.2187
Cadier M., Sourisseau M., Gorgues T. et al. 2017. Assessing spatial and temporal variability of phytoplankton communities' composition in the Iroise Sea ecosystem (Brittany, France): A 3D modeling approach: Part 2: Linking summer mesoscale distribution of phenotypic diversity to hydrodynamism // J. Marine Systems. V. 169. May. P. 111. https://doi.org/10.1016/j.jmarsys.2017.01.004
Canfield D.E., Bachmann R.W., Hoyer M.V. 2018. Long-term chlorophyll trends in Florida lakes // J. Aquat. Plant Manage. V. 56. P. 47.
Chen Y., Qin B., Teubner K., Dokulil M.T. 2003. Long-term dynamics of phytoplankton assemblages: Microcystis-domination in Lake Taihu, a large shallow lake in China // J. Plankton Res. V. 25. № 1. P. 445. https://doi.org/10.1093/plankt/25.4.445
Gao N., Ma Y., Zhao M. et al. 2020. Quantile analysis of long-term trends of near-surface chlorophyll-a in the Pearl River plume // Water. V. 12. № 6. P. 1662. https://doi.org/10.3390/w12061662
Honti M., Istvanovics V., Osztoics A. 2007. Stability and change of phytoplankton communities in a highly dynamic environment – the case of large, shallow Lake Balaton (Hungary) // Hydrobiologia. V. 581. P. 225. https://doi.org/10.1007/s10750-006-0508-2
Kalff J. 2002. Limnology. Inland Water Ecosystems. Upper Saddle River. N.Y.: Prentice Hall.
Kangur K., Milius A., Mols T. et al. 2002. Lake Peipsi: Changes in nutrient elements and plankton communities in the last decade // Aquat. Ecosyst. Health Manage. V. 5. № 3. P. 363. https://doi.org/10.1080/14634980290001913
Lamont T., Barlow R.G., Brewin R.J.W. 2019. Long-term trends in phytoplankton chlorophyll a and size structure in the Benguela upwelling system // JGR Oceans. V. 124. № 2. P. 1170. https://doi.org/10.1029/2018JC014334
Mendesab C.R., Saa C., Vitorinoc J. et al. 2011. Spatial distribution of phytoplankton assemblages in the Nazaré submarine canyon region (Portugal): HPLC-CHEMTAX approach // J. Marine System. V. 87. № 1. P. 90. https://doi.org/10.1016/j.jmarsys.2011.03.005
Mineeva N.M. 2021. Long-term dynamics of photosynthetic pigments in plankton of the large plain reservoir // Biosystem Diversity. V. 29. № 1. P. 10. https://doi.org/10.15421/012102
Nascimento Moura A., Nascimento E.C., Dantas E.W. 2012. Temporal and spatial dynamics of phytoplankton near farm fish in eutrophic reservoir in Pernambuco, Brazil // Rev. Biol. Trop. V. 60. № 2. P. 581.
Öterler B., Albay M., Çamur-Elipek B. et al. 2015. Spatial and temporal distribution of phytoplankton in Lake Gala (Edirne/Turkey) // Trakya University Journal of Natural Sciences. V. 16. № 2. P. 71. https://doi.org/dergipark.ulakbim.gov.tr/trkjnat/index
Padisák J. 2004. Phytoplankton // The Lakes Handbook 1. Limnology and Limnetic Ecology. Oxford: Blackwell Science Ltd. P. 251.
Rakočević J. 2012. Spatial and temporal distribution of phytoplankton in Lake Skadar // Arch. Biol. Sci., Belgrade. V. 64. № 2. P. 585. https://doi.org/10.2298/ABS1202585R
Reynolds C.S. 2006. The Ecology of Phytoplankton. Cambridge: University Press.
Ruggiu D., Morabito G., Panzani P., Pugnetti A. 1998. Trends and relations among basic phytoplankton characteristics in the course of the long-term oligotrophication of Lake Maggiore (Italy) // Hydrobiologia. V. 369/370. P. 243.
Rusanov A., Trifonova I., Ignatyeva N., Pavlova O. 2020. Long-term changes in phytoplankton and macrophyte communities in an eutrophic shallow reservoir and prospects for its restoration // Oceanological and Hydrobiological Studies. V. 49. Iss. 2. P. 168. https://doi.org/10.1515/ohs-2020-0016
SCOR-UNESCO Working Group 17. 1966. Determination of photosynthetic pigments in sea water // Monographs on Oceanographic Methodology. Montreux: UNESCO. P. 9.
Sommer U., Adrian R., De Senerpont Domis L. et al. 2012. Beyond the plankton ecology group (PEG) model: Mechanisms driving plankton succession // Annual Review of Ecology, Evolution, and Systematics. V. 43. P. 429. https://doi.org/10.1146/annurev-ecolsys-110411-160251
Wetzel R.G. 2001. Limnology. Lake and River Ecosystems. San Diego: Acad. Press.
Wiedner C., Nixdorf B., Heinze R. et al. 2002. Regulation of Cyanobacteria and microcystin dynamics in polimictic shallow lakes // Archiv für Hydrobiologie. V. 155. P. 383.
Yang M., Xia J., Cai W. et al. 2020. Seasonal and spatial distributions of morpho-functional phytoplankton groups and the role of environmental factors in a subtropical river-type reservoir // Water Sci. Technol. V. 82. № 11. P. 2316. https://doi.org/10.2166/wst.2020.489
Yang Y., Pettersson K., Padisák J. 2016. Repetitive baselines of phytoplankton succession in an unstably stratified temperate lake (Lake Erken, Sweden): a long-term analysis // Hydrobiologia. V. 764. Iss. 1. P. 211. https://doi.org/10.1007/s10750-015-2314-1
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод