

Биология внутренних вод, 2022, № 3, стр. 233-244
Два новых для науки вида хромадорид (Nematoda, Chromadorida) из водных объектов Вьетнама
Фан Ке Лонг a, В. Г. Гагарин b, *, Нгуен Динь Ты c
a Вьетнамский национальный музей природы, Вьетнамская академия наук и технологий
Ханой, Вьетнам
b Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл, Россия
c Институт экологии и биологических ресурсов Вьетнамской Академии наук и технологий,
Отдел нематологии
Ханой, Вьетнам
* E-mail: gagarin@ibiw.ru
Поступила в редакцию 17.02.2021
После доработки 24.11.2021
Принята к публикации 12.01.2022
- EDN: CDPPLX
- DOI: 10.31857/S0320965222030032
Аннотация
Приведено иллюстрированное описание двух новых для науки видов нематод из водных объектов Вьетнама. Dichromadora agilis sp. n. обнаружена в пруду для выращивания креветок. Морфологически наиболее близка к D. apapillata Timm, 1961 и D. weddellensis Vermeeren, Vanreusel, Vanhoves, 2004, но отличается от обоих видов структурой пищевода, более длинными головными щетинками и формой рулька. Приведена таблица основных морфологических признаков валидных видов рода Dichromadora, самцы которых не имеют преклоакальных супплементов. Chromadorina affinis sp. n. обнаружена в устье р. Ван Ак. Вид близок к Сh. macropunctata (Wieser, 1954) и Ch. cervix (Wieser, 1951). От Сh. macropunctata отличается относительно более коротким хвостом у самцов, более короткими головными щетинками и наличием преклоакальнных супплементов, от Ch. cervix – относительно более толстым телом, более короткими головными щетинками и наличием глазков. Приведена таблица основных морфологических признаков самцов валидных видов рода Chromadorina.
ВВЕДЕНИЕ
Фауна свободноживущих нематод пресных и солоноватых водоемов Вьетнама исследуется с 2002 г. (Fauna ..., 2007; Gusakov, Gagarin, 2017; Gagarin, 2018; Гагарин, 2021). Выявлено >250 видов нематод, из них более половины описаны как новые для науки.
Род Dichromadora Kreis, 1929 довольно богат видами, широко распространенными в морских и солоноватых водах (Gerlach, Riemann, 1973). Виды рода Chromadorina Filipjev, 1918 также обычны в морских и солоноватых водоемах, четыре вида встречаются и в пресных водоемах (Gerlach, Riemann, 1973).
Цель работы — описать два новых для науки вида: Dichromadora agilis sp. n. и Chromadorina affinis sp. n.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
В 2016 г. в устье р. Ван Ак (прибрежье Южно-Китайского моря) и в искусственных водоемах для выращивания креветок исследовали фауну свободноживущих нематод. Пробы отбирали на глубине 0.3–0.7 м с помощью пластикового цилиндра диаметром 3.5 см и длиной 10 см, промывали через сачок из газа с диаметром ячеи 0.08 мм и фиксировали горячим (60–70°С) 4%-ным раствором формальдегида. Затем пробу помещали в емкость объемом 200 мл, добавляли раствор Ludox-TM50 (1 : 1) и центрифугировали пять раз по 3–5 мин. Нематод переводили в чистый глицерин по методу Зайнхорста (Seinhorst, 1959), монтировали в небольшой капле глицерина на предметных стеклах и опечатывали кольцом из парафина–воска. Для измерения и определения особей, а также для фотографирования и изготовления рисунков использовали световой микроскоп Nikon Eclipse 80i с дифференциально-интерференционным контрастом и цифровую камеру Nikon DS-Fil. Для анализа данных использовали программу NIS-Elements D3.2.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Описание видов
Отряд Chromadorida Chitwood, 1933
Семейство Chromadoridae Filipjev, 1917
Род Dichromadora Kreis, 1929
Диагноз (по: Huang & Zhang, 2010; Tchesunov, 2014). Кутикула гомогенная, орнаментирована, с боковыми полями в форме двух продольных рядов увеличенных точек. Лабиальные сенсиллы в форме папилл, четыре головные сенсиллы в форме тонких щетинок. Фовеи амфидов в форме поперечной щели или петли. Ротовая полость с крупным, неподвижным полым дорсальным зубом и двумя более мелкими субвентральными зубами. Фаринкс обычно не расширен в области ротовой полости или ассиметрично расширен дорсально. Базальный бульбус фаринкса хорошо развит. Преклоакальные супплементы имеются или отсутствуют.
Типовой вид Dichromadora cephalata (Steiner, 1916) Kreis, 1929 (=Chromadora cephalata Steiner 1916).
В настоящее время в род Dichromadora входят 32 валидных вида (Venekey et al., 2019). Самцы 20 видов не имеют преклоакальных супплементов. В водоемах Вьетнама обнаружено пять видов этого рода (D. affinis Gagarin, Nguyen Vu Thanh, 2011, D. apapillata Timm, 1961, D. arcospiculum Timm, 1961, D. rigida Nguyen Vu Thanh, Nguyen Dinh Tu, Gagarin, 2016, D. simplex Timm, 1961 (Gagarin, 2018).
Dichromadora agilis sp. n. (рис. 1, 2).
Рис. 1.
Детали строения голотипа самца (а, в, г, д, е) и паратипа самки (б) Dichromadora agilis sp. n.: а, б – общий вид; в – передний конец тела; г – спикула и рулек; д – тело в области базального бульбуса фаринкса; е – задний конец тела. an. – анус; a.o. – передний яичник; b.b. – бульбус фаринкса; ca. – кардий; c.g. – каудальные железы; ch. – хейлостома; cl. – клоака; c.s. – головные щетинки; d.t. – дорсальный зуб; eg. – яйцо; f.amph. – фовея амфид; gub. – рулек; p.o. – задний яичник; spic. – спикула; spin. – спиннерета; t. – хвост, tes. – семенник; v. – вульва.
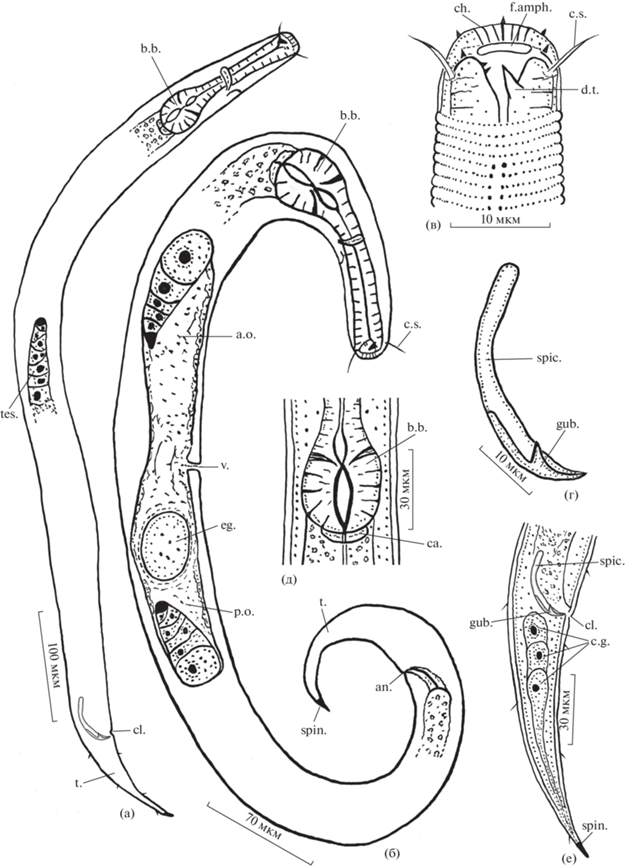
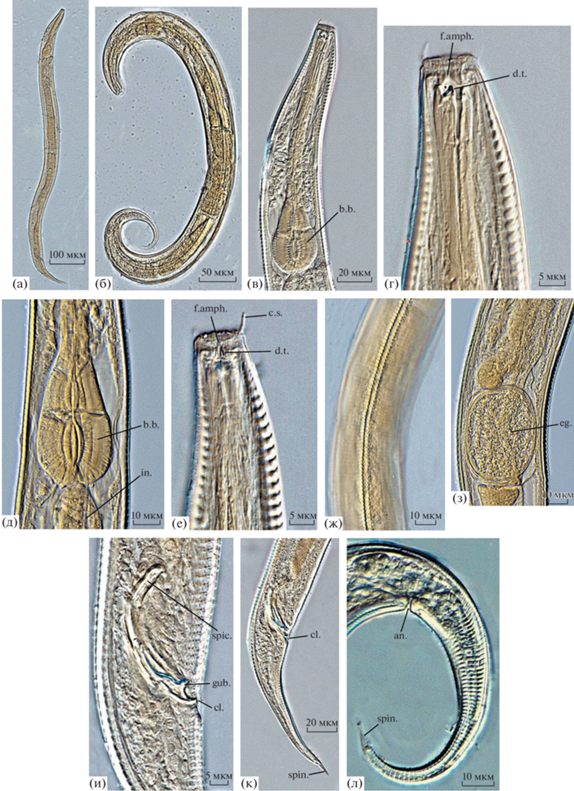
Рис. 2.
Микрофотографии голотипа самца (а, в, г, д, ж, и, к) и паратипа самки (б, е, з, л) Dichromadora agilis sp.n. а, б – общий вид; в – передний конец тела; г, е – голова; д – тело в области базального бульбуса пищевода; ж – строение кутикулы в среднем отделе тела; з – тело в области вульвы; и – тело в области клоаки; к, л – задний конец тела. in. – средняя кишка, остальные обозначения, как на рис. 1.
Материал. Голотип: самец (инвентарный номер препарата HD 4.1.13); паратипы 5♂♂, 2♀♀. Препарат голотипа хранится в коллекции Музея природы Вьетнамской академии наук и технологий (Ханой, Вьетнам), препараты паратипов – в коллекции нематод Института экологии и биологических ресурсов Вьетнамской академии наук и технологий (г. Ханой, Вьетнам).
Местонахождение. Северный Вьетнам, провинция Куангнинь (Quang Ninh), искусственный пруд для выращивания креветок. Координаты: 21°16′15″ с.ш., 107°23′65″ в.д. Глубина 0.5 м, грунт – заиленная дерновина. Соленость 16.8‰.
Описание. Морфометрическая характеристика голотипа и паратипов приведена в табл. 1.
Таблица 1.
Морфометрическая характеристика Dichromadora agilis sp. n.
| Признак | Голотип 1♂ |
Паратипы | |||
|---|---|---|---|---|---|
| 5♂♂ | 2♀♀ | ||||
| диапазон | среднее | ♀ | ♀ | ||
| L, мкм | 808 | 694–807 | 759 | 829 | 792 |
| a | 19 | 17–20 | 18 | 16 | 17 |
| b | 5.9 | 5.8–6.1 | 5.9 | 6.6 | 6.6 |
| c | 7.9 | 7.1–7.9 | 7.4 | 6.8 | 6.8 |
| c' | 3.8 | 3.8–4.4 | 4.0 | 4.8 | 4.4 |
| V, % | – | – | – | 47.2 | 46.8 |
| Ширина, мкм: | |||||
| области губ | 12 | 12–14 | 13 | 12 | 12 |
| тела в его среднем отделе | 43 | 39–43 | 41 | 51 | 48 |
| тела в области клоаки или ануса | 27 | 24–27 | 26 | 26 | 27 |
| Длина, мкм: | |||||
| фарингеального бульбуса | 42 | 36–43 | 39 | 45 | 43 |
| головных щетинок | 7 | 7–8 | 7 | 8 | 8 |
| фаринкса | 136 | 119–139 | 129 | 126 | 120 |
| спикул (по дуге) | 37 | 37–39 | 38 | – | – |
| рулька | 22 | 20–23 | 21 | – | – |
| хвоста | 102 | 97–105 | 102 | 122 | 120 |
| Расстояние, мкм: | |||||
| от конца фаринкса до вульвы | – | – | – | 265 | 251 |
| от конца фаринкса до клоаки | 570 | 478–568 | 528 | – | – |
| от вульвы до ануса | – | – | – | 316 | 301 |
Примечание. Здесь и в табл. 2–4: а – отношение длины тела к его максимальной ширине, b – отношение длины тела к длине фаринкса, c – отношение длины тела к длине хвоста, с' – отношение длины хвоста к диаметру тела в области ануса или клоаки, L – длина тела, V – отношение расстояния от переднего конца тела до вульвы к длине тела; “–” – отсутствие данного признака.
Самцы. Стройные черви среднего размера. Кутикула кольчатая. Толщина кутикулы в среднем отделе тела 1.0–1.5 мкм. Латеральные поля состоят из двух продольных рядов довольно крупных точек, начинаются на уровне переднего отдела фаринкса и тянутся вдоль тела до середины хвоста. Ширина полей 4–5 мкм. Шейные щетинки не обнаружены. Соматические щетинки редкие и короткие. Передний конец тела сужен. Область губ не обособлена от остального тела. Внутренние и внешние губные сенсиллы в форме мелких едва заметных папилл. Четыре головные сенсиллы в форме тонких щетинок длиной 7–8 мкм. Фовеи амфидов в форме поперечной щели и расположены на уровне переднего отдела фарингостомы. Хейлостома с продольными кутикулярными ребрами. Фарингостома в форме воронки и вооружена сравнительно крупным, полым дорсальным зубом и двумя мелкими субвентральными зубами. Фаринкс мускулистый, с хорошо развитым базальным бульбусом, внутренняя полость которого разделена плазматической перегородкой на два отдела, причем задний отдел более крупный, чем передний. Кардий маленький.
Семенник один, прямой, расположен справа от средней кишки. Спикулы стройные, изогнуты вентрально. Длина спикул в 1.3–1.4 раза превышает диаметр тела в области клоаки. Рулек в форме удлиненного желоба, причем, в дистальной его части вентрально расположен треугольный заостренный выступ. Длина рулька 20–23 мкм. Преклоакальные супплементы не обнаружены. Хвост удлиненно-конический, постепенно сужается. Каудальные железы хорошо развиты. Спиннерета удлиненно-коническая, “клювовидная”, длиной 8–9 мкм.
Самки. По общей морфологии подобны самцам. Строение кутикулы и переднего конца тела как у самцов. Кутикула кольчатая, гомогенная. Боковые поля в форме двух продольных рядов крупных точек. Внутренние и внешние губные сенсиллы в форме папилл. Четыре головные сенсиллы в форме щетинок длиной 8 мкм. Фовеи амфидов в форме поперечной изогнутой щели, расположенной на уровне переднего отдела фарингостомы. Хейлостома с кутикулярными ребрами. Фарингостома в форме воронки, вооружена крупным полым дорсальным зубом и двумя мелкими субвентральными зубами. Фаринкс мускулистый, с хорошо развитым базальным бульбусом. Внутренняя полость бульбуса разделена плазматическими перегородками на два отдела. Задний отдел более крупный, чем передний.
Яичники парные, загнутые, сравнительно короткие. Вульва слегка преэкваториальная, в форме поперечной щели. Ее губы не склеротизированы и не выступают за контуры тела. Передний яичник расположен справа от средней кишки, задний – слева. Вагина короткая, с толстыми стенками. Обе матки заполнены многочисленными сперматозоидами. Хвост удлиненно-конический. Три каудальные железы хорошо развиты. Спиннерета удлиненно-коническая, длиной 8 мкм.
Дифференциальный диагноз. Dichromadora agilis sp. n. относится к группе видов рода, самцы которых не имеют преклоакальных супплементов (табл. 2). По длине тела и длине спикул D. agilis sp. n. близка к D. apapillata и D. weddellensis. От первого вида отличается структурой базального бульбуса фаринкса (внутренняя полость бульбуса фаринкса разделена на два отдела, у D. apapillata она единая, цельная), сравнительно коротким фаринксом (b = 5.8–6.6 против b = 5.4–5.5 у D. apapillata), более длинными головными щетинками (их длина 7–8 мкм или 53–57% ширины области губ, у D. apapillata 6 мкм или 50% соответственно) и более длинным и иной формы рульком (его длина 20–25 мкм и имеет заостренный вентральный выступ на дистальном конце, у D. apapillata длина рулька 16 мкм и он лишен заостренного треугольного выступа на дистальном конце (Timm, 1961)). От второго вида, D. weddellensis, новый вид отличается структурой базального бульбуса фаринкса (он разделен на два отдела, у D. weddellensis он единый, цельный), более толстым телом (а = 16–20 против а = 20–31 у D. weddellensis), более длинными головными щетинками (их длина 7–8 мкм, 53–57% ширины области губ против 6 мкм длины и 36–38% ширины области губ) и иной структурой рулька (на дистальном конце рулька имеется заостренный вентральный выступ, отсутствующий на рульке у самцов D. weddellensis (Vermeeren et al., 2004).
Таблица 2.
Морфометрическая характеристика валидных видов рода Dichromadora, самцы которых не имеют супплементов (по: Vermeeren et al., 2004, с добавлением)
| Вид | L, мкм | a | b | c | V, % | Ширина области губ, мкм | Длина, мкм | ||
|---|---|---|---|---|---|---|---|---|---|
| головные щетинки | спикула | рулек | |||||||
| D. abnormis Gerlach, 1953 | 761–1103 | 24–34 | 5.4–6.1 | 7.0–7.5 | 45–47 | 22–23 | 15–18 | 24 | – |
| D. affinis Gagarin, Nguyen Vu Thanh, 2011 | 580–874 | 20–33 | 5.3–7.1 | 5.7–9.3 | 45–51 | 11–13 | 4.0–5.0 | 24–26 | 13–16 |
| D. amphidiscoides Kito, 1981 | 398–612 | 18–31 | 6.1–7.5 | 4.5–5.3 | – | 5–7 | – | – | – |
| D. antarctica (Cobb, 1914) | 420–680 | 23–36 | 5.4–7.0 | 3.5–5.2 | 45–58 | 6–8 | 4 | 14 | – |
| D. apapillata Timm, 1961 | 729–820 | 15–17 | 5.4–5.5 | 5.5–7.2 | 48 | 12 | 6 | 38 | 16 |
| D. arcospiculum Timm, 1961 | 540–578 | 18–30 | 5.2–6.1 | 6.0–7.1 | 47–49 | 8–10 | 1.1–2.0 | 23–28 | 15–18 |
| D. gathuai Muthumbi, Vincx, 1998 | 509–817 | 26–33 | 5.5–7.5 | 5.3–7.8 | 43–47 | 8–10 | – | – | – |
| D. geophila (de Man, 1876) | 583–958 | 13–24 | 5.7–8.0 | 5.6–12.0 | 44–47 | 13–14 | 5 | 28–34 | 16–18 |
| D. gracilis (Kreis, 1929) | 1108–1218 | 30–33 | 8.3–8.4 | 7.3–10.9 | 45–49 | – | 11–12 | 37–41 | 19–21 |
| D. islandica Kreis, 1963 | 764–989 | 18–22 | 6.3–7.2 | 6.5–7.6 | 47–49 | – | – | – | – |
| D. longicaudata Muthumbi, Vincx, 1998 | 425–590 | 30–45 | 6.7–8.6 | 2.5–4.0 | 39–43 | 5 | 2–3 | 18.4 | 7 |
| D. multisetosa Huang, Zhang, 2010 | 470–535 | 17–21 | 5.0–6.0 | 5.8–7.1 | 49–51 | 14–17 | 7–14 | 34–38 | 10–13 |
| D. parasimplex Dashchenko, 2002 | 838–1043 | 18–21 | 5.4–6.7 | 6.7–8.4 | 48–50 | 13–15 | 6–7 | 41–46 | 17–18 |
| D. polarsternis Vermeeren, Vanreusel, Vanhove, 2004 | 964–1183 | 21–30 | 6.3–7.4 | 6.7–9.4 | 47–56 | 12 | 7 | 28 | – |
| D. punctata Schuurmans, Stekhoven, 1950 | 904 | 23 | 7.0 | 7.5 | – | 20 | 3 | 20–24 | 10 |
| D. simplex Timm, 1961 | 480–543 | 15–20 | 5–6 | 6.5–7.8 | 48–49 | 10–12 | 3 | 20–25 | 12–13 |
| D. strandi Allgen, 1940 | 1700–1875 | 31 | 8.1–8.3 | 10.3–10.4 | – | – | – | – | – |
| D. tobaensis Schneider, 1937 | 337–700 | 16–28 | 4.6–6.5 | 6.5–9.0 | 46–47 | – | – | – | – |
| D. weddellensis Vermeeren, Vanreusel, Vanhove, 2004 | 687–951 | 24–31 | 6.3–8.0 | 7.6–8.7 | 47–57 | 11 | 6 | 37 | 21 |
| D. agilis sp. n. | 694–829 | 16–20 | 5.8–6.6 | 6.8–7.9 | 47 | 12–14 | 7.8 | 37–39 | 20–23 |
Этимология. Видовое название нового вида означает “подвижный, проворный”.
Род Chromadorina Filipjev, 1918.
Диагноз (по: Tchesunov, 2014). Кутикула гомогенная с поперечными рядами точек, но без их латеральной дифференцировки. Фовеи амфидов, когда они видимы, в форме поперечной овальной щели. Фарингостома с тремя плотными зубами примерно равного размера. Глазки могут присутствовать. У большинства видов имеются преклоакальные кубковидные супплементы. Хвост удлиненно-конический с железами и хорошо развитой спиннеретой.
Типовой вид Chromadorina obtusa Filipjev, 1918.
В настоящее время в состав рода Chromadorina входят 27 валидных видов (Venekey, Gheller et al., 2019). Большинство их обитает в море, но четыре вида (Ch. astacicola, Ch. bercziki, Ch. bioculata и Ch. viridis) в пресной воде и три вида (Ch. erythrophthalma, Ch. germanica и Ch. laeta) и в пресной, и в соленой воде.
Chromadorina affinis sp. n. (рис. 3, 4).
Рис. 3.
Детали строения голотипа самца (а–д) и паратипа самки (е, ж) Chromadorina affinis sp. n.: а – общий вид; б – передний конец тела; в – спикула и рулек; г – тело в области базального бульбуса фаринкса; д, ж – задний конец тела; е – тело в области вульвы. in. – средняя кишка, oc. – глазок, sup. – cупплемент. Остальные обозначения, как на рис. 1.
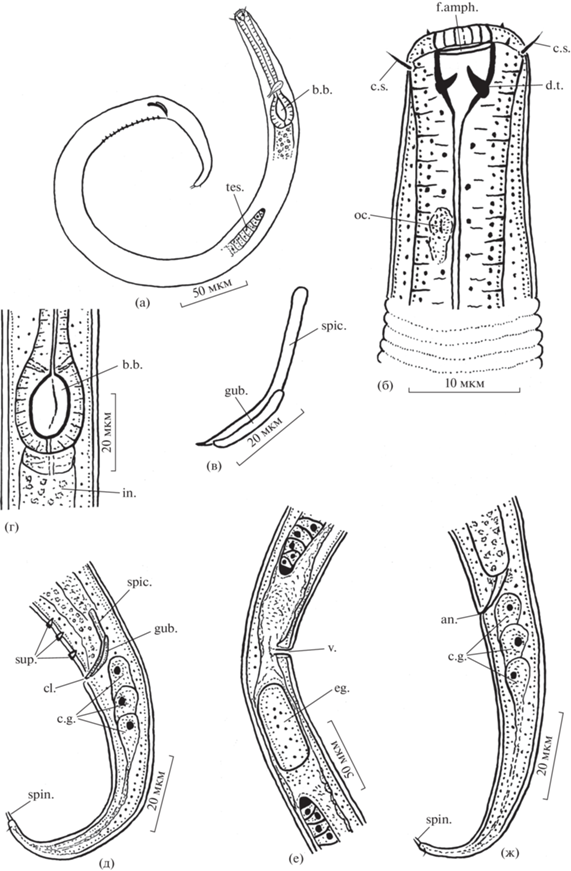
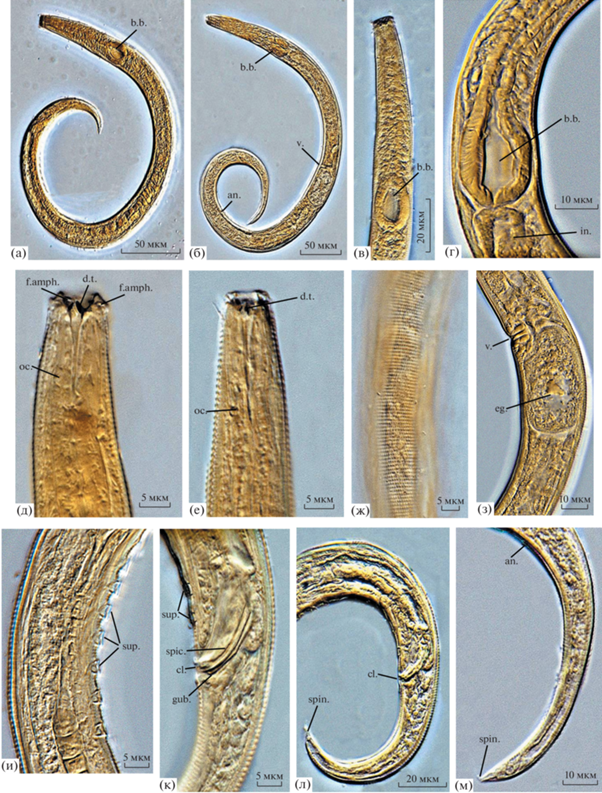
Рис. 4.
Микрофотографии голотипа самца (а, в, г, д, ж, и, к, л) и паратипа самки (б, е, з, м) Chromadorina affinis sp. n.; а, б – общий вид; в – передний конец тела; г – тело в области базального бульбуса фаринкса; д, е – головной конец; ж – строение кутикулы в среднем отделе тела; з – тело в области вульвы; и – тело в области супплементов; к – тело в области клоаки; л, м – задний конец тела. Буквенные обозначения как на рис. 1 и рис. 3.
Материал. Голотип: самец (инвентарный номер препарата HТ 1.2.5); паратипы 15♂♂, 12♀♀. Препараты голотипа и шести паратипов (4♂♂, 2♀♀) хранятся в коллекции Музея природы Вьетнамской академии наук и технологий (Ханой, Вьетнам), препараты остальных паратипов – в коллекции нематод Института экологии и биологических ресурсов Вьетнамской академии наук и технологий (г. Ханой, Вьетнам).
Местонахождение. Северный Вьетнам, провинция Куангнинь (Quang Ninh), устье р. Ван Ак (Van Ac River). Координаты: 21°16′20″ с.ш., 107°25′88″ в.д. Глубина 1.0–1.5 м, грунт – заиленный песок, мангровые заросли. Соленость воды 18.2‰.
Описание. Морфометрическая характеристика голотипа и паратипов приведена в табл. 3.
Таблица 3.
Морфометрическая характеристика Chromadorina affinis sp. n.
| Признак | Голотип | Паратипы | |
|---|---|---|---|
| 1♂ | 15♂♂ | 12♀♀ | |
| L, мкм | 474 | 481± 34 (410–545) | 489 ± 18 (462–526) |
| a | 24 | 22 ± 2 (18–28) | 19 ± 1 (18–22) |
| b | 5.6 | 5.8 ± 0.4 (5.1–6.4) | 5.9 ± 0.2 (5.5–6.2) |
| c | 7.5 | 7.7 ± 0.5 (7.0–8.6) | 6.8 ± 0.5 (6.2–7.2) |
| c' | 4.1 | 3.9 ± 0.3 (3.4–4.4) | 5.3 ± 0.4 (4.8–5.9) |
| V, % | – | – | 48.1 ± 1.1 (46.2–50.8) |
| Ширина, мкм: | |||
| области губ | 9 | 10 ± 0.1 (9–10) | 10 ± 1 (9–11) |
| тела в его среднем отделе | 20 | 22 ± 2 (20–26) | 25 ± 1 (24–27) |
| тела в области ануса или клоаки | 15 | 16 ± 1 (14–17) | 15 ± 1 (14–16) |
| Длина, мкм: | |||
| базального бульбуса | 26 | 24 ± 1 (22–26) | 22 ± 2 (20–26) |
| головных щетинок | 3.0 | 2.5 ± 0.1 (2.0–3.0) | 2.5 ± 0.1 (2.0–3.0) |
| фаринкса | 85 | 83 ± 3 (78–90) | 83 ± 2 (80–88) |
| спикул (по дуге) | 24 | 24 ± 1 (22–26) | – |
| рулька | 11 | 12 ± 1 (10–13) | – |
| хвоста | 63 | 63 ± 3 (56–68) | 74 ± 4 (66–80) |
| Расстояние, мкм: | |||
| от конца фаринкса до вульвы | – | – | 152 ± 12 (136–179) |
| от конца фаринкса до клоаки | 326 | 335 ± 31 (272–390) | – |
| от вульвы до ануса | – | – | 178 ± 10 (156–189) |
| Число супплементов | 13 | 13 ± 1 (12–15) | – |
Самцы. Мелкие, сравнительно стройные черви. Кутикула кольчатая, гомогенная, без латеральных полей. Соматические щетинки редкие и короткие. Губная область не обособлена от остального тела. Шесть внутренних и шесть внешних губных сенсилл в форме мелких папилл. Четыре головные сенсиллы в форме щетинок длиной 2.0–3.0 мкм. Пигментные пятна (глазки) имеются, расположены на расстоянии 15–18 мкм от переднего конца тела. Фовеи амфидов в форме поперечной овальной щели, расположены на уровне верхнего отдела фарингостомы; едва видны на препаратах. Хейлостома с кутикулярными ребрами. Фарингостома в форме воронки со сравнительно тонкими кутикулярными стенками, вооружена тремя плотными зубами, причем дорсальный зуб слегка крупнее обоих субвентральных зубов. Фаринкс мускулистый; его базальный бульбус хорошо развит, длиной 22–26 мкм (28–30% длины фаринкса). Внутренняя полость бульбуса обширная, с сильно кутикулярными стенками.
Семенник один, прямой и расположен справа от средней кишки. Спикулы стройные, изогнутые, длиной 22–26 мкм. Рулек в форме удлиненной пластинки и в ~2 раза короче спикул. Перед клоакой расположено 12–15 кубковидных супплементов. Хвост удлиненно-конический. Три каудальные железы. Спиннерета удлиненно-коническая, длиной 5 мкм.
Самки. По общей морфологии подобны самцам. Строение кутикулы и переднего конца тела, как у самцов. Кутикула кольчатая, гомогенная, без латеральной дифференцировки. Внутренние и внешние губные сенсиллы в форме папилл. Четыре головные сенсиллы в форме щетинок длиной 2.0–3.0 мкм. Глазки имеются, расположены на расстоянии 14–17 мкм от переднего конца тела. Фовеи амфидов в форме поперечной овальной щели расположены на уровне верхнего отдела фарингостомы. Хейлостома с кутикулярными ребрами. Фарингостома в форме воронки и вооружена тремя плотными зубами, дорсальный зуб слегка крупнее субвентральных. Фаринкс стройный, мускулистый. Его базальный бульбус хорошо развит, длиной 22–27 мкм (28–30% длины фаринкса). Внутренняя полость бульбуса обширная. Длина ректума равна или немного больше диаметра тела в области ануса.
Яичники парные, загнутые, сравнительно короткие. Передний яичник расположен справа от средней кишки, задний – слева. Вульва в форме поперечной щели. Ее губы не склеротизированы. Вагина с толстыми стенками. Матки обширные. В матках найдены созревшие яйца размером 43–48 × 22–24 мкм. Хвост удлиненно-конический, постепенно сужается. Спиннерета удлиненно-коническая, длиной 5–7 мкм.
Дифференциальный диагноз. По количеству преклоакальных супплементов новый вид близок к Ch. bercziki Andrássy, 1962, Ch. erythrophthalma, Ch. salina и Сh. viridis (табл. 4). От первого вида он отличается меньшей длиной тела (♂♂ L = 410–545 мкм против ♂♂ L = 700–840 мкм у Ch. bercziki), наличием глазков (у Ch. bercziki они отсутствуют), более короткими головными щетинками (их длина 2.0–3.0 мкм против 5.0–6.0 мкм у Ch. bercziki), более короткими спикулами и рульком (их длина соответственно равна 22–26 и 10–13 мкм против 34–38 и 22–24 мкм у Ch. bercziki) и отсутствием отростка у спикул (у Ch. bercziki спикулы имеют сравнительно длинный шиповидный отросток) (Andrássy, 1962). От Ch. erythrophthalma новый вид отличается меньшей длиной тела (♂♂ L = 410–545 мкм против ♂♂ L = 810–1000 мкм у Ch. erythrophthalma), более короткими головными щетинками (их длина 2.0–3.0 мкм, 28–30% диаметра области губ против 7.5 мкм, 50% диаметра области губ у Ch. erythrophthalma), более короткими спикулами и рульком (их длина соответственно 22–26 и 10–13 мкм против 36 и 22 мкм у Ch. erythrophthalma (Schneider, 1906)). От Ch. salina новый вид отличается более коротким телом (♂♂ L = 410–545 мкм против ♂♂ L = 950–1050 мкм у Ch. salina), относительно более длинным и стройным хвостом (с = = 7.0–8.6, с' = 3.4–4.4 против с = = 9.7–11.6, с' = 3.0 у Ch. salina), более короткими головными щетинками (их длина 2–3 против 5–8 мкм у Ch. salina) и более короткими спикулами и рульком (их длина соответственно равна 22–26 и 10–13 мкм против 43–57 и 24–30 мкм у Ch. salina (Белогуров, 1978). От Ch. viridis новый вид отличается меньшими размерами тела (♂♂ L = 410–545 мкм против ♂♂ L = 600–900 мкм у Ch. viridis), более короткими головными щетинками (их длина 2–3 мкм, 28–30% диаметра области губ против 5–7 мкм, 65–80% диаметра области губ у Ch. viridis), более короткими спикулами и рульком (их длина соответственно равна 22–26 и 10–13 мкм против 35–38 и 17–20 мкм у Ch. viridis) и наличием глазков (у Ch. viridis они отсутствуют (Andrássy, 1984)). По размерам тела и размерам спикул Ch. affinis sp. n. близок к Ch. macropunctata (Wieser, 1954) и Ch. cervix (Wieser, 1951). От первого вида он отличается относительно коротким хвостом у самцов (♂ с = = 7.0–8.6 против с = 4.1–6.6 у Ch. macropunctata), более короткими головными щетинками (их длина 2.0–3.0 мкм, 25–33% диаметра области губ против 8.0 мкм или 110% ширины области губ у Ch. macropunctata) и наличием кубковидных супплементов у самцов (их 12–15 штук, у Ch. macropunctata они отсутствуют (Wieser, 1954)). От Ch. cervix новый вид отличается более толстым телом (а = 18–28 против а = 32–35 у Ch. cervix), более короткими головными щетинками (их длина 2.0–3.0 мкм, 25–33% ширины области губ против 4.5 мкм или 60–70% ширины области губ у Ch. cervix) и наличием глазков (у Ch. cervix они отсутствуют (Wieser, 1951).
Таблица 4.
Основные морфометрические признаки самцов валидных видов рода Chromadorina
| Вид | L, мкм | a | b | c | с' | Диаметр области губ, мкм | Длина головных щетинок, мкм | Длина спикул, мкм | Длина рулька, мкм | Число преклоакальных супплементов |
|---|---|---|---|---|---|---|---|---|---|---|
| Ch. amata (Allgen, 1933) | 1050 | 35 | 8.1 | 8.1 | – | – | – | – | – | 11 |
| Ch. astacicola (Schneider, 1932) | 1000 | 30 | 8.7 | 9.6 | 12–13 | 7.0 | 29–30 | 22 | 0 | |
| Ch. bergensis (Allgen, 1932) | 747 | 28 | 6.4 | 8.3 | – | – | – | – | – | 8 |
| Ch. bercziki Andrássy, 1962 | 700–840 | 24–32 | 6.7–7.8 | 8.0–9.0 | 4.2–4.6 | 15 | 5.0–6.0 | 34–38 | 22–24 | 11–12 |
| Ch. bioculata (Schultze in Carus, 1857) | 560–830 | 17–23 | 6.4–8.5 | 6.0–7.8 | 4.4–5.0 | – | 7.5–8.5 | 21–28 | 16–18 | 0 |
| Ch. cervix (Wieser, 1951) | 520–560 | 32–35 | 6.0–6.4 | 7.5–8.4 | 4.0 | 27–32 | 4.5 | 23 | – | 7 |
| Ch. demani Inglis, 1962 | 630–700 | 21–22 | 5.8–6.3 | 7.3–7.5 | 3.1–3.4 | 29–33 | 6.0 | 28–34 | 25–28 | 3 |
| Ch. epidemos Hopper, Meyer, 1967 | 610–780 | 26–37 | 5.5–6.6 | 5.3–6.0 | 6.7–8.7 | 8.0–9.0 | 5.0–6.0 | 26–30 | 14–15 | 0 |
| Ch. erythrophthalma (Schneider, 1906) | 810–1000 | 23 | 5.4 | 8.5 | 3.5 | 15.0 | 7.5 | 36 | 22 | 15 |
| Ch. germanica (Butschli, 1874) | 780–1000 | 21–25 | 6.4–7.6 | 7.0–9.4 | 3.5–4.0 | 11–15 | 5.5–8.0 | 26–27 | 16–21 | 17–18 |
| Ch. granulopigmentata (Wieser, 1951) | 560–650 | 23–24 | 5.6–5.8 | 6.4–6.5 | 4.5 | 10 | 6.5 | – | – | 0 |
| Ch. hiromii Kito, Nakamura, 2001 | 633–670 | 33–35 | 6.1–6.4 | 8.9–9.2 | 4.0–4.4 | 9.9–10.5 | 5.1–5.9 | 22–25 | 11–15 | 5–6 |
| Ch. incurvata Wieser, 1951 | 920 | 34 | 7.6 | 7.3 | 5.7 | 14.5 | 5.0 | 29 | – | 0 |
| Ch. inversa Wieser, 1955 | 330 | 22 | 4.3 | 4.9 | 7.0 | 7.5 | 3.5 | 11 | – | 0 |
| Ch. longisetosa (de Coninck, Sch. Stekhoven, 1950) | 1720 | 47 | 9.3 | 6.5 | 17 | 25 | 8.8 | 29 | 15 | 9 |
| Ch. longispiculum Paster de Ward, 1985 | 1050–1680 | 18–24 | 6.5–9.3 | 8.4–10.2 | 3.0 | 30 | 12 | 45 | 30 | 17–22 |
| Ch. macropunctata (Wieser, 1954) | 440–500 | 17–21 | 5.2–5.4 | 4.1–6.6 | 4.0–5.0 | 7.5 | 8.0 | 20–22 | 10–11 | 0 |
| Ch. majae Wieser, 1968 | 2040–2100 | 50–53 | 11.0–11.4 | 12.0–13.2 | 3.5–4.0 | 14.0–15.5 | 5.0–6.5 | 41–44 | – | 0 |
| Ch. metulata Aissa, Vitiello, 1977 | 897–1076 | 28–42 | 6.8–8.4 | 10.0–13.0 | 3.2–4.7 | – | 5.0–7.0 | 20–22 | – | 4 |
| Ch. micoletzkii Inglis, 1962 | 570–660 | 21–25 | 4.6–5.3 | 7.4–7.6 | – | – | – | – | – | 3 |
| Ch. nuda Wieser, 1954 | 710 | 28 | 7.4 | 7.7 | 5.0 | 10 | – | 19 | 8.5 | 2 |
| Ch. obtusa Filipjev, 1918 | 675 | 19 | 5.6 | 7.0 | 3.2 | 14 | 6.0 | 32 | 30 | 2 |
| Ch. paradoxa Timm, 1961 | 1300 | 37 | 9.1 | 8.4 | 6.7 | – | 6.5 | 26 | 15 | 3 |
| Ch. rognoeensis (Allgen, 1932) | 785 | 29 | 6.3 | 8.7 | – | – | – | – | – | 0 |
| Ch. salina Belogurov, 1978 | 950–1050 | 24–25 | 5.9–6.5 | 9.7–11.6 | 3.0 | 14–15 | 5–8 | 43–57 | 24–30 | 14–15 |
| Ch. supralitoralis Lonrezen, 1969 | 590–605 | 24–25 | 6.0–6.4 | 6.0–8.1 | 3.5–3.8 | 10 | 5–6 | 22 | – | 0 |
| Ch. viridis (Linstow, 1876) | 600–900 | 19–24 | 5.6–7.0 | 6.0–6.9 | 3.0–4.0 | 8.0–9.0 | 5.0–7.0 | 35–38 | 17–20 | 14–16 |
| Ch. affinis sp. n. | 410–545 | 18–28 | 5.1–6.4 | 7.0–8.6 | 3.4–4.4 | 8–10 | 2.0–3.0 | 22–26 | 10–13 | 12–15 |
Морфологические замечания. Длина тела видов рода Chromadorina колеблется от 330 до 1720 мкм. Самый крупный из них Ch. majae (длина самцов 2040–2100), самый мелкий Ch. inversa (330 мкм) (табл. 4). Большинство видов имеют глазки. Длина головных щетинок достигает 18–80% диаметра области губ. Длина спикул колеблется от 11 до 57 мкм (1.0–1.5 диаметра тела в области клоаки). У большинства видов самцы имеют преклоакальные супплементы (табл. 4), лишь у четырех видов они отсутствуют.
Список литературы
Белогуров О.И. 1978. Изучение свободноживущих нематод литорали острова Щикотан // Животный и растительный мир шельфовой зоны Курильских островов. Владивосток: Дальневосточный научный центр. С. 139.
Andrássy I. 1962. Nematoden aus dem Psammon des Adige-Flusses II // Memorie del Museo Civica de Storia Naturale di Verona. V. 10. S. 1.
Andrássy I. 1984. Klasse Nematoda (Ordnungen Monhysterida, Araeolaimida, Chromadorida, Desmoscolecida, Rhabditida). Berlin: Akademie-Verlag.
Fauna of Vietnam. Dong Vat Chi Viet Nam. 2007. V. 22. Hanoi: Science and Technics Publishing House. (In Vietnamese).
Gagarin V.G. 2018. An annotated checklist of the free-living nematodes from mangrove thickets of Vietnam // Zootaxa. № 4403(2). P. 261.
Gagarin V.G. 2021. Two New Species of Free-living Nematodes (Nematodes, Monhysterida) from the Cam River, Vietnam // Inland Water Biology. V. 14. № 1. https://doi.org/10.31857/S0320965221010022
Gerlach S.A., Rieman F. 1973. The Bremerhaven checklist of aquatic Nematodes. A Catalogue of Nematoda Adenophorea excluding the Dorylaimida // Veröffenntlichungen Instituts fur Meeresforschung in Bremerhaven. Suppl. 4. P. 1–404.
Gusakov V.A., Gagarin V.G. 2017. An annotated checklist of the main represantives of meiobentos from inland water bodies of Central and Southern Vietnam. I. Rounworms (Nematoda) // Zootaxa. № 3. V. 1. P. 1.
Huang Y., Zhang Z.H. 2010. Three new species of Dichromadora (Nematoda: Chromadorida: Chromadoridae) from Ho Yellow Sea, China // J. Natural History. V. 44. № 9–10. P. 545.
Schneider G. 1906. Beitrag zur Kenntnis der im userschlamm des Finischen Meerbussens freilebenden Nematoden // Acta Societalis pro Fauna et Flora Fennica. T. 27. № 7. S. 1.
Seinhorst J.V. 1959. A rapid method for the transfer of nematodes from fixative to anhydrous glycerin // Nematologica. V. 4. P. 67.
Tchesunov A.V. 2014. Order Chromadorina Chitwood, 1933 // Handbook of Zoology. V. 2. (Nematoda). P. 373.
Timm R.W. 1961. The marine nematodes of the Bay of Bengal // Proceedings of the Pakistan Academy of Sciences. V. 1. № 1. P. 1.
Venekey V., Gheller P.F., Kondratavicius N. et al. 2019. The state of the art of Chromadoridae (Nematoda, Chromadorida): a historical review, diagnoses and comments about valid and dubious genera and a list of valid species // Zootaxa. V. 4578. № 1. P. 1.
Vermeeren H., Vanreusel A., Vanhove S. 2004. Species distribution within the free-living marine nematode genus Dichromadora in the Weddell Sea and adjacent areas // Deep-Sea Research. Part II. V. 51. P. 1643.
Wieser W. 1951. Untersuchungen ueber die algenbewohnende Microfauna mariner Haryboden, I. Zur Oecologie ubd Systematik der Nematodenfauna von Plymouth // Oesterreichische zoologische Zeitschrift. Bd 3. № 3–4. S. 425.
Wieser W. 1954. Free-living marine nematodes. II. Chromadoroidea // Acta Universitets Lund (N. F. 2). V. 50. № 16. P. 1.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод